
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
Conflict adaptation reflects the increase in cognitive control after previous conflict between task-relevant and task-irrelevant information. Tonic (sustained) arousal elicited by emotional words embedded in a conflict task has previously been shown to increase conflict adaptation. However, the role of phasic (transient) emotional arousal remains unclear. In Experiment 1 (N = 55), we therefore investigated the effect of phasic arousal using a colour flanker task with negative, positive, and neutral words as stimuli. We hypothesised that phasic arousal elicited in this context will increase conflict adaptation in the subsequent trial. Indeed, when the words were positive or negative as compared to neutral, we observed increased conflict adaptation. In Experiment 2 (N = 54), we examined the role of the self-relevance by presenting words with a self-related pronoun (“my”) or sender-related pronoun (“his”/“her”). We expected that emotional words with high self-relevance would lead to stronger effects of emotional arousal on conflict adaptation. Confirming this hypothesis, results showed that emotional words within a self-related context again increased conflict adaptation, whereas this effect was not observed in the sender-related context. Taken together, these results are the first to show that phasic arousal elicited by emotional words increases conflict adaptation, in particular when these words have high self-relevance.
In modern society, people are constantly confronted with many streams of information competing for our attention. Being able to ignore irrelevant information while focusing on the relevant parts is essential for goal-directed behaviour. When goal-relevant and distracter information is conflicting, it becomes more difficult to select the appropriate action. Cognitive control helps to resolve conflict in these situations, serving goal-relevant action. In the lab, conflict is usually created by presenting participants simultaneously with target and distracter information while measuring changes in performance. Famous examples of conflict tasks are the Stroop, Simon, and flanker tasks (Eriksen & Eriksen, Citation1974; Simon & Rudell, Citation1967; Stroop, Citation1935). For example, in the arrow flanker task, participants have to indicate the direction of a centrally presented arrow that is flanked by arrows facing in a direction that is either the same (congruent) or different (incongruent) from the central arrow. It has consistently been observed that this task is more difficult for incongruent trials than for congruent trials due to the interference of the distracting arrows during incongruent conditions. This effect is called the congruency effect and is reflected by slower reaction times and more errors on incongruent as compared to congruent trials. Interestingly, the interference of the conflicting information becomes smaller when a trial is preceded by another incongruent trial. This sequential effect is commonly referred to as the congruency-sequence effect or conflict-adaptation effect (Botvinick, Braver, Barch, Carter, & Cohen, Citation2001; Gratton, Coles, & Donchin, Citation1992).
One of the most influential theories that explains why conflict adaptation occurs is the conflict monitoring theory (Botvinick et al., Citation2001). This theory assumes that the occurrence of conflict will trigger increased cognitive control. Accordingly, when conflict is detected during an incongruent trial, a neural conflict monitoring system signals to prefrontal areas that more cognitive control is needed to respond adequately to the situation. Consequently, the conflict is resolved by an increase in cognitive control, leading to the facilitation of attentional selection of the target during the next trial. This adaptive increase in cognitive control is thought to produce the conflict-adaptation effect. Although previous research has shown that congruency sequence effects can also be driven partly by alternative mechanisms, including feature integration (Hommel, Proctor, & Vu, Citation2004) and contingency learning (Schmidt & De Houwer, Citation2011), conflict adaptation can still be observed when carefully controlling for these factors (Braem et al., Citation2019; Duthoo, Abrahamse, Braem, Boehler, & Notebaert, Citation2014; Schmidt & Weissman, Citation2014).
Research from the last decade has started to focus on studying affective (van Steenbergen, Citation2015) and motivational factors (Braem, Verguts, Roggeman, & Notebaert, Citation2012; Stürmer, Nigbur, Schacht, & Sommer, Citation2011) that influence adaptive cognitive control. For example, recent findings have emphasised the role of integral emotions in conflict tasks, suggesting that conflict itself is aversive and that negative valence and/or increased arousal drives conflict adaptation (Botvinick, Citation2007; Dreisbach & Fischer, Citation2015; Inzlicht, Bartholow, & Hirsh, Citation2015; van Steenbergen, Citation2015). Confirming this idea, conflict has been shown to induce physiological arousal (e.g. Brown et al., Citation1999; Laeng, Ørbo, Holmlund, & Miozzo, Citation2011; van der Wel & van Steenbergen, Citation2018; van Steenbergen & Band, Citation2013), primes negative affect (e.g. Dreisbach & Fischer, Citation2012), and the presumed negative valence of conflict can be counteracted by positive valence induced by performance non-contingent reward, reducing the conflict-adaptation effect (van Steenbergen, Band, & Hommel, Citation2009; but also see Dignath, Janczyk, & Eder, Citation2017; Padmala, Bauer, & Pessoa, Citation2011; Yamaguchi & Nishimura, Citation2019, Exp. 2 & 3). In addition, affective manipulations incidental to the task at hand modulate conflict adaptation, such that negative relative to positive affect increases conflict adaptation (Kuhbandner & Zehetleitner, Citation2011; Schuch & Koch, Citation2015; Schuch, Zweerings, Hirsch, & Koch, Citation2017; van Steenbergen, Band, & Hommel, Citation2010; van Steenbergen, Band, Hommel, Rombouts, & Nieuwenhuis, Citation2015; but see also Dreisbach, Fröber, Berger, & Fischer, Citation2019), suggesting that when the valence of affective states and the conflict itself are congruent, the aversive conflict signal is boosted which consequently increases conflict adaptation.
In contrast to the relatively consistent literature on valence, reported findings on the effects of experimentally induced emotional arousal on conflict adaptation are rather mixed. Although phasic (transient) arousal has been predicted to drive conflict adaptation (Verguts & Notebaert, Citation2009), studies that manipulate phasic arousal using a stimulus that is not embedded in the main task typically do not alter conflict adaptation (Brown, van Steenbergen, Kedar, & Nieuwenhuis, Citation2014; Dignath et al., Citation2017), or only impair task switching (Demanet, Liefooghe, & Verbruggen, Citation2011). Studies using tonic (sustained) manipulations of emotional arousal via mood induction have reported null-effects (Kuhbandner & Zehetleitner, Citation2011; van Steenbergen et al., Citation2010) or only impairments of top-down control of distractors (Kuhbandner & Zehetleitner, Citation2011). A pharmacological manipulation of the noradrenergic system using propranolol also did not impact conflict adaptation (de Rover et al., Citation2015). However, a recent study employing transcutaneous vagus nerve stimulation, which is assumed to activate the locus coeruleus-norepineprine (LC-NE) system, did result in increased conflict adaptation (Fischer, Ventura-Bort, Hamm, & Weymar, Citation2018; but also see van Bochove et al., Citation2018), although the exact contribution of phasic and tonic arousal to this stimulation is not clear yet (Warren et al., Citation2019).
In the present study, we investigate the role of emotional valence and arousal following a different approach that was originally developed by Kanske and Kotz (Citation2010, Citation2011a, Citation2011b, Citation2011c). In their experiments, these authors used conflict tasks that embedded the emotional stimuli in the primary task. For example, using a colour flanker task, the stimulus involved an array of the same emotional word while manipulating the colour of the central target and the distractors (see for an example). Although the emotional words were irrelevant to the primary task, emotional versus neutral words increased cognitive control (reduced conflict effect) in the colour flanker task, an effect observed for both positive (Kanske & Kotz, Citation2011b, Citation2011c) and negative stimuli (Kanske & Kotz, Citation2010, Citation2011a). These findings clearly suggest that phasic arousal, but not valence, increases cognitive control provided that the emotional stimulus is embedded in the task itself (for reviews, see Cohen & Henik, Citation2012; Kanske, Citation2012). Following up on this line of research, Zeng et al. (Citation2017) recently used the same colour flanker task to investigate the effect of valence and arousal on conflict adaptation in the same study. Consistent with a role of arousal on conflict adaptation, they observed increased conflict adaptation when stimuli were emotional as opposed to neutral. This increased conflict adaptation was observed for both the positive and negative conditions, suggesting that the valence of emotional words do not modulate conflict adaptation. Critically, in the study of Zeng and colleagues phasic and tonic effects of arousal could not be dissociated because positive, negative, and neutral emotional words were presented in different blocks. So it is yet unknown whether the effect they reported is due to a phasic arousal effect, a tonic arousal effect, or a combination of both. Therefore, the present study aimed to further investigate the effect of phasic arousal, by using a trial-wise manipulation of emotional words.
Figure 1. Overview of a congruent trial (left) and an incongruent trial (right) in Experiment 1.
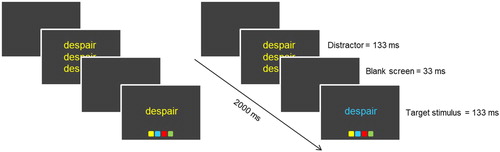
In Experiment 1, we used an adapted version of the task developed by Zeng et al. (Citation2017). We manipulated valence and arousal by randomly presenting colour flanker stimuli with emotional (negative or positive) and neutral content within blocks. Given previous findings, we predicted that irrespective of valence, emotional versus neutral stimuli in the current trial will increase conflict adaptation in the next trial. In Experiment 2, we further investigated whether the self-relevance of the emotional words alters the effect of emotion on control processes, by adding a self-related or sender-related pronoun to the words. Task designs in both experiments experimentally controlled for priming, repetition, and contingency learning effects which otherwise could invalidate the interpretation of conflict-adaptation effects (Schmidt & Weissman, Citation2014).
Experiment 1
Method
Participants
We aimed to collect data for 60 participants so that our analysis would have sufficient power (>80%) to detect small-to-medium within-subject effects with Cohen's d ≥ .41, which has been described to reflect the average effect size in psychology (Lakens & Evers, Citation2014). Following this calculation, 61 right-handed participants (22 men) with a mean age of 21.54 (SD = 2.03) took part in the study. All participants were native Dutch speakers with no self-reported history of trauma or psychiatric illness. Participants were compensated for taking part in the study using either course credits or money. The study was approved by the local ethics committee at Leiden University. As done in our earlier work (e.g. van Steenbergen, Weissman, Stein, Malcolm-Smith, & van Honk, Citation2017), we first did an initial screening of the data on individual aggregated accuracy rates using boxplots. This revealed participants that were outliers on either (or both) congruent or incongruent conditions (i.e. more than 1.5 interquartile ranges below/above the 25th/75th percentile). Removing these outliers resulted in the exclusion of six participants, which were outliers due to making errors in over 20% of (in)congruent trials. When we include the data of these subjects (ignoring those with empty cells due to missing observations) the direction and significance of the key effects reported below did not change.
Flanker task
We adapted the colour flanker task that was developed by Kanske and Kotz (Citation2010), see . Participants had to indicate the colour of a target word. Before the participants had to respond to the target stimulus, a distractor display was shown in which the target word was flanked by the same word both above and underneath it. The words on the distractor display could be in either the same colour (congruent) or a different colour (incongruent) from the target stimulus. There were two possible colour combinations of the flankers and target stimuli, which were yellow/blue and red/green, following the design of Zeng et al. (Citation2017). On congruent trials, only one of the colours was used. Critically, in order to prevent priming, repetition, and contingency learning effects, the two combinations were presented in alternating order, such that no current-trial colour or response was repeated on the subsequent trial (for a similar design, see Schmidt & Weissman, Citation2014).
Participants had to indicate the colour of the target word with a key press. Each of the four colours was assigned to a key. This colour-key mapping was counterbalanced across participants. The participants had to press the “D” and “F” keys with the left middle and index fingers and the “J” and “K” keys with their right middle and index fingers, respectively. Throughout the experiment, as a reminder, the order of the colours were displayed on the bottom of the screen in the corresponding order of the keys on the keyboard.
Each trial lasted exactly 2000 ms. The distractor was displayed for 133 ms, followed by a blank screen of 33 ms. The target word was displayed for 133 ms followed by a window to respond that lasted for a maximum of 1374 ms. A blank time fill screen was presented for the remaining time, to make sure that there was exactly 2000 ms between each distractor display.
The target words could be either emotional or neutral. Words of each of these categories were presented equally often in each block in random order. The valence of emotional words was counterbalanced between subjects, with half of the participants being presented with positive emotional and neutral words, and the other half being presented with negative emotional and neutral words. The stimuli were Dutch words taken from the dataset of Moors et al. (Citation2013). For each word type, 30 nouns were chosen. Each word was presented 8 times during the experiment. All positive, negative, and neutral words were compared on valence, arousal, number of letters, frequency (Moors et al., Citation2013), and concreteness (Brysbaert, Stevens, De Deyne, Voorspoels, & Storms, Citation2014). See for an overview and the supplementary material for the stimuli used.
Table 1. Mean scores and standard deviations of valence, arousal, concreteness, frequency and number of letters of each word group.
Procedure
After giving informed consent, participants started the experiment with a practice block, which consisted of 32 trials using another set of words. Participants were instructed to respond as fast as possible while avoiding mistakes. During the practice trials, textual feedback was shown for 200 ms after each trial, informing the participant whether their response was correct, incorrect or too slow. Aside from the feedback, practice trials were the same as experimental trials. Participants had to achieve an overall accuracy of at least 90% to advance to the experimental blocks. When participants had lower accuracy, they were instructed to inform the experimenter and had to redo the practice block. During the experiment, 17 out of 61 participants had to redo the practice block one or more times (M = 1.34, range = 1-4). After having completed the practice block, participants performed two experimental blocks. Both blocks consisted of 240 trials, each block taking eight minutes to complete. Between the two blocks, there was a break as well as a reminder of the instructions. After completing the flanker task, participants filled out the State-Trait Anxiety Inventory (Spielberger, Gorsuch, & Lushene, Citation1970). This questionnaire was added for a student project and not included in the analyses reported here.
Data analyses
We analysed both reaction times and error rates. We used the following trial-level exclusions that are standard in conflict adaptation studies. For the RT analysis, we excluded the first trials of each block (0.4% of all trials), inaccurate trials (4.2%), as well as trials following errors (4.2%). Furthermore, the data was inspected for normality. Subsequently, all trials per subject were split for all factors involved for the subsequent ANOVA (e.g. previous-trial emotion, previous-trial congruency, and current-trial congruency for the main analysis). Trials with a RT deviating more than 2.5 standard deviations from the individual condition-specific mean were excluded from the analyses (2.0%). For the accuracy analyses, we excluded the first trial of a block, trials following errors, and trials with no responses from the analysis. Analyses on the RT data were used to test our hypotheses whereas analyses on accuracy were used to confirm that observed effects in RT were not driven by shifts in speed-accuracy trade-off.
Repeated measures ANOVAs were used on reaction times and error rates of the current trial using the within-subject factors previous-trial emotion (neutral vs. emotional), previous-trial congruency (congruent vs. incongruent), and current-trial congruency (congruent vs. incongruent). The valence of the emotions (positive or negative) was added as a between-subject factor.
Greenhouse-Geisser correction was used when the assumption of sphericity was violated. All main and interaction effects with p-values < .1 are reported.
Results
Reaction times
provides an overview of all reaction times per combination of conditions, as well as the congruency and conflict-adaptation effects separate for both levels of previous-trial emotion. Analyses revealed a significant main effect of current congruency, which showed that incongruent trials resulted in slower RTs than congruent trials (F(1,53) = 689.54, p < .001, MSE = 1126.9, eta²p = .929). In addition, there was an effect of previous congruency on RT, showing that trials following previous incongruent trials were slower than trials that followed congruent trials (F(1,53) = 4.06, p = .049, MSE = 190.2, eta²p = .071). Importantly, there was also a significant interaction between current-trial and previous-trial congruency, indicating our task produced the conflict-adaptation effect (F(1,53) = 69.24, p < .001, MSE = 177.0, eta²p = .566).
Table 2. Behavioral data of Experiment 1 (N = 55).
Most importantly, we observed the predicted interaction between previous-trial emotion, previous-trial congruency and current-trial congruency (F(1,53) = 6.86, p = .011, MSE = 96.1, eta²p = .115). As shown in , conflict adaptation was stronger for emotional words (26 ms, SE = 3.2) than for neutral words (16 ms, SE = 3.3). Post-hoc tests did not clearly reveal whether the difference was driven by the effect of previous conflict, previous no-conflict or their combination, as the interference effect after congruent trials did not differ between the previous neutral trials versus emotional trials (t(54) = −1.870, p = .067) and the interference effect after incongruent trials was also not significantly different between previous neutral versus emotional trials (t(54) = 1.209, p = .232).
Figure 2. Reaction times as a function of previous congruency (x-axis) and current congruency (lines) for neutral (left) and emotional (right) previous-trial words. The error bars indicate the 95% confidence intervals of the between-subject standard errors.
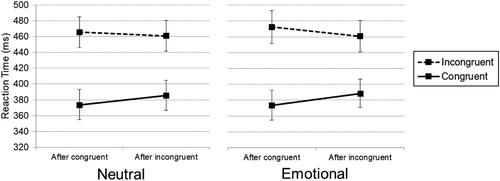
Additionally, we observed faster reaction times after previous neutral trials as compared to previous emotional trials (F(1,53) = 4.63, p = .036, MSE = 134.5, eta²p = .080). We also observed a non-significant interaction between the between-subject factor valence and conflict adaptation, (F(1,53) = 3.49, p = .067, MSE = 117.0, eta²p = .062), in the direction suggesting that a context of mixed negative and neutral relative to mixed positive and neutral words numerically increases conflict adaptation. There were no overall between-subject effects of the valence of the emotion (positive or negative), (F(1,53) = .03, p = .871, MSE = 38783.1, < .001), nor any other significant interactions between valence and any other (combinations of) conditions. This indicates that there was no evidence that the valence of the emotional words influenced the effect of previous-trial emotion on conflict adaptation.
Error rates
An overview of the mean error rates per condition can be found in . A repeated-measures ANOVA on accuracy showed that participants made more errors on incongruent trials than on congruent trials (F(1,53) = 7.984, p = .007, MSE < .1, = .131). After previously incongruent trials error rates reduced (F(1,53) = 13.47, p = .001, MSE < .1,
= .203). Critically, there was no interaction between emotion and conflict adaptation (F(1,53) = .241, p = .626, MSE < .1,
= .005), indicating that the differences in reaction time cannot be explained by a speed-accuracy trade-off. No other effects were observed (ps > .206).
Discussion
The aim of Experiment 1 was to test whether emotional versus neutral words embedded in a colour flanker task increase conflict-driven upregulations of cognitive control when they are presented in random order. Consistent with the hypothesis, conflict adaptation was increased by positive and negative emotional words compared to neutral words, irrespective of the between-subject factor valence. These findings extend the findings of Zeng et al. (Citation2017) that revealed increased conflict adaptation for emotional words in a blocked design that did not allow the disentanglement of the phasic and tonic effects of arousal. The present results show for the first time that phasic arousal elicited by emotional words increases conflict adaptation in the subsequent trial.
Experiment 2
When participants read emotional words in experimental settings, it is important to realise that emotions could simply label the emotion expressed by the sender, rather than that they reflect one’s own emotions. In order to make emotional words self-related, Herbert and colleagues (Citation2011a) therefore developed a new paradigm in which the affective noun was preceded by the pronoun my (“e.g. my despair”); compared to sender-related pronoun (“e.g. his despair”). Emotional nouns preceded by self-related pronouns have been shown to facilitate deeper emotional processing and accompany larger physiological responses in several studies, suggesting that they change the level of experienced arousal (e.g. Bayer, Ruthmann, & Schacht, Citation2017; Herbert, Herbert, Ethofer, & Pauli, Citation2011a; Herbert, Pauli, & Herbert, Citation2011b). Building on this work, in Experiment 2 we combined this approach with the emotional flanker task used in Experiment 1 to test whether self-relevance is associated with stronger effects of emotional arousal on conflict adaptation.
Method
Participants
The sample consisted of 60 new participants (21 male) with a mean age of 21.72 (SD = 2.53). The exact same exclusion criteria from the first experiment were used. Initial screening of the data on individual aggregated accuracy rates using boxplots showed participants that were outliers on either (or both) congruent or incongruent conditions (i.e. more than 1.5 interquartile ranges below/above the 25th/75th percentile). Removing these outliers resulted in the exclusion of six participants, which were outliers due to making errors in over 22% of (in)congruent trials. When we include the data of these subjects (ignoring those with empty cells due to missing observations) the direction and significance of the key effects reported below did not change.
Flanker task
Experiment 2 used the same paradigm as Experiment 1, but we now manipulated relevance by adding a self-related pronoun (“my”) or a sender-related pronoun (either “his” or “her”, matched to participant’s gender) to the nouns, following up on the earlier work by Herbert and colleagues (Citation2011b).
Procedure
Additional practice trials were added to the experiment, so that the practice consisted of a self-related block and sender-related block with 16 trials each. During the experiment, 23 out of 60 participants had to redo the practice block (M = 1.6, range = 1-8) one or more times until they received an overall accuracy of at least 90%. The order of the experimental blocks was counterbalanced between participants using an ABBA design (A = Self, B = Sender, or vice versa). Because we manipulated relevance within participants, the amount of trials was doubled from 480 to 960, again with 240 trials per block. After completing the flanker task, participants filled out the State-Trait Anxiety Inventory (Spielberger et al., Citation1970) and the Empathy Quotient (Baron-Cohen & Wheelwright, Citation2004) questionnaire. Both questionnaires were added for a student project. Following a reviewer’s suggestion, we performed an exploratory analysis that correlated the Empathy Quotient with the effect of self-relevance on the interaction of previous emotion and conflict adaptation (Herbert, Sfärlea, & Blumenthal, Citation2013; Meixner & Herbert, Citation2018). No significant effect was observed (r = -.077; p = .580).
Data analyses
Data analyses were identical to Experiment 1, except that personal relevance was added as an additional within-subject factor. For the RT analysis, we excluded the first trials of each block (0.4% of all trials), inaccurate trials (4.5%), as well as trials following errors (4.5%), and trials with a RT deviating more than 2.5 standard deviations from the individual condition-specific mean were excluded from the analyses (2.1%).
Results
Reaction times
and provide an overview of all reaction times per combination of conditions. As in Experiment 1, analyses revealed a significant main effect of current congruency, which showed that incongruent trials resulted in slower RTs than congruent trials (F(1,52) = 1007.86, p < .001, MSE = 1531.6, eta²p = .951). In addition, there was an effect of previous congruency on RT, showing that trials following previous incongruent trials were slower than trials that followed congruent trials (F(1,52) = 4.92, p = .031, MSE = 253.2, eta²p = .086). Again, there was also a significant interaction between current-trial and previous-trial congruency, again indicating that our task produced the standard conflict-adaptation effect (F(1,52) = 77.91, p < .001, MSE = 274.6, eta²p = .600). The interaction between previous-trial emotion and current-trial congruency (F(1,52) = 2.98, p = .090, MSE = 234.6, eta²p = .054) was not significant. However, we did observe a significant interaction between the between-subject factor valence and current-trial congruency (F(1,52) = 4.74, p = .034, MSE = 1531.6, eta²p = .084), suggesting that the congruency effect – but not the conflict-adaptation effect, F(1,52) = 2.19, p = .145, MSE = 274.6, eta²p = .040 – was larger in the a context of mixed negative and neutral relative to mixed positive and neutral words.
Table 3. Behavioral data of Experiment 2 of self-related blocks (N = 54).
Table 4. Behavioral data of Experiment 2 of sender-related blocks (N = 54).
Most importantly, in contrast to Experiment 1, there was no overall interaction between previous emotion, previous congruency and current congruency (F(1,52) = 1.92, p = .172, MSE = 209.8, eta²p = .036). Instead, this interaction was moderated by the relevance indicated by the pronoun, as indicated by a significant four-way interaction between relevance, previous emotion, previous congruency, and current congruency (F(1,52) = 5.55, p = .022, MSE = 213.9, eta²p = .096). Relevance did not significantly modulate overall conflict adaptation (F(1,52) = .19, p = .667, MSE = 215.9, eta²p = .004). Furthermore, there were no between-subject effects of the valence of the emotion (positive or negative) (F(1,52) = .30, p = .586, MSE = 81514.8, eta²p = .006) nor any significant interactions between the valence and the interaction between the relevance, emotion, and conflict adaptation (F(1,52) = .10, p = .753, MSE = 213.9, eta²p = .002), indicating that the valence of the emotions did not influence the effects of the relevance and emotion on the conflict-adaptation effect.
To further investigate the interaction between relevance, previous emotion, and conflict adaptation, ANOVAs were run separately for the self-related and sender-related blocks. Analysis on the self-related blocks again showed effect of current congruency (F(1,52) = 833.52, p < .001, MSE = 957.0, eta²p = .941), previous congruency (F(1,52) = 4.10, p = .048, MSE = 202.1, eta²p = .073), and their interaction (F(1,52) = 42.99, p < .001, MSE = 227.7, eta²p = .453). Additionally, we observed faster reaction times after previous neutral trials as compared to previous emotional trials (F(1,52) = 9.67, p = .003, MSE = 137.8, eta²p = .157). Most importantly, self-related blocks did show a significant interaction between emotion and conflict adaptation (F(1,52) = 6.82, p = .012, MSE = 218.1, eta²p = .116), which replicates the effects observed in Experiment 1; as shown in (top panels), we observed increased conflict adaptation after emotional words (26 ms, SE = 4.0) compared to neutral words (12 ms, SE = 3.7), t(53) = 2.630, p = .011. Post-hoc tests showed that interference after congruent trials did not differ between the neutral trials and emotional trials (t(53) = .154, p = .878). However, the interference effect after incongruent trials was lower for previous emotional trials as opposed to neutral trials (t(53) = 4.180, p < .001).
Figure 3. Reaction times as a function of previous congruency (x-axis) and current congruency (lines) for neutral (left panels) and emotional (right panels) previous-trial words, separate for self-related blocks (top panels) and sender-related blocks (bottom panels). The error bars indicate the 95% confidence intervals of the between-subject standard errors.
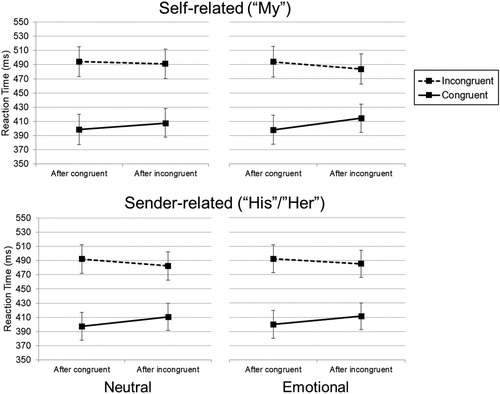
On the other hand, analyses on the sender-related blocks did not show a significant interaction between previous emotion and conflict adaptation (F(1,52) = .50, p = .482, MSE = 205.7, eta²p = .010), see (bottom panels). This analysis also confirmed the effect of current congruency (F(1,52) = 876.82, p < .001, MSE = 851.2, eta²p = .944), the interaction between previous and current congruency (F(1,52) = 44.33, p < .001, MSE = 262.8, eta²p = .460), and the effect of the between-subject factor valence on the congruency effect (F(1,52) = 6.00, p = .018, MSE = 851.2, eta²p = .103).
Error rates
An overview of the mean error rates per condition can be found in and . A repeated-measures ANOVA on accuracy showed that the amount of errors on incongruent trials and congruent trials differed, with more errors on incongruent trials (F(1,52) = 7.34, p = .009, MSE < .1, = .124). After previously incongruent trials error rates reduced (F(1,52) = 7.85, p = .007, MSE < .1,
= .131). Critically, there were no interactions between emotion and conflict adaptation (F(1,52) < .01, p = .974, MSE < .1,
< .001), nor did relevance modulate the effect (F(1,52) = .16, p = .692, MSE < .1,
= .003), indicating that the differences in reaction time cannot be explained by a speed/accuracy trade-off. No other effects were observed (ps > .121).
Discussion
In line with predictions, the findings of Experiment 2 show that the pronouns added before the words in our flanker task tended to influence the interaction between previous-trial emotion and conflict adaptation. Irrespective of valence, emotional words that were preceded by the self-related pronoun “my” increased conflict adaptation relative to neutral words, replicating the findings from Experiment 1 in a self-related context. Interestingly, when words were sender-related by using the pronoun “his” or “her” (matched to the participant’s gender), emotional content no longer modulated conflict adaptation.
General discussion
The goal of the present study was to investigate the effect of phasic arousal on cognitive control, by investigating the effect of emotional versus neutral words that are embedded in a colour flanker task. As predicted, Experiment 1 showed that emotional stimuli, irrespective of being positive or negative, increase conflict adaptation in comparison to neutral words. Replicating and extending these findings, Experiment 2 showed the same effect in a self-related context when words were preceded by the pronoun “my”, whereas this effect was not observed in a sender-related context when words were preceded by the pronoun “his” or “her”.
Our results replicate and extend the findings of Zeng et al. (Citation2017) who demonstrated that emotional arousal manipulated at a block-level increases conflict adaptation. Our results clearly show that this effect of arousal also occurred when emotion is manipulated at the trial-level, thus demonstrating that the induction of phasic emotional arousal is sufficient to increase conflict adaptation. The adaptation-by-binding account of Verguts and Notebaert (Citation2009) provides a mechanistic explanation for our findings. According to this theory, phasic arousal increases the binding between the representations of stimuli and task-relevant goals via Hebbian learning triggered by the locus-coeruleus norepinephrine system. This binding serves to improve cognitive control in the next trial, reflected by increased conflict adaptation. However, at the same time several previous studies have not provided consistent evidence for arousal-related increases in conflict adaptation (Brown et al., Citation2014; Dignath et al., Citation2017), or observed reduced adaptation after threat-related images (Padmala et al., Citation2011). Based on these findings, it could be speculated that phasic arousal enhances conflict adaptation only when it has strong overlap in terms of both temporal and spatial features, as our paradigm used emotional words that were embedded in the task itself. Note that the effects observed in the present study could also reflect effects of emotional intensity rather than phasic arousal because we did not use a low-arousal control condition that was matched on valence. However, because emotional stimuli typically show a boomerang-shaped distribution in the two-dimensional space defined by arousal and valence, it is challenging to dissociate phasic arousal from emotional intensity in experimental studies.
Our control analyses (see supplementary material) showed that the emotion-induced increase in conflict adaptation did not accompany overall increases in cognitive control (reduced interference effect) in the trial of the emotional stimulus itself. At first sight, these findings seem to contradict the original findings by Kanske and Kotz (Citation2010, Citation2011a, Citation2011b, Citation2011c) that have repeatedly shown that emotional words, irrespective of valence, improve cognitive control (i.e. reduce the congruency effect) in the trial of the emotional word itself. However, this apparent contradiction can be reconciled by assuming that conflict adaptation reflects a carry-over of an enhanced cognitive control state that started at the previous trial, a suggestion that has been supported by earlier work that has dynamically tracked target and distracter processing over time (Scherbaum, Fischer, Dshemuchadse, & Goschke, Citation2011). Because our paradigm used four response options (to control for common confounds in the conflict-adaptation effects) and quite short inter-trial intervals, it was likely more difficult than the slower two-response paradigms originally employed by Kanske and Kotz. Task differences like these could make it such that participants relied more on reactive cognitive control mechanisms that do not impact behaviour immediately while it does improve performance in a subsequent trial.
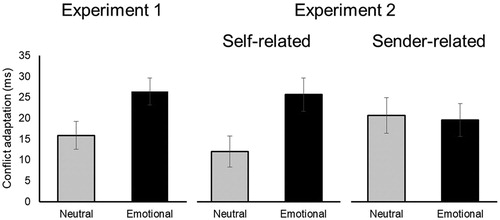
The self-relevance manipulation introduced in Experiment 2 demonstrated that the context in which emotional words are processed influences control processes. As illustrated in , emotion only increased conflict adaptation when words were self-related (mirroring the findings in Experiment 1) but not when they were sender-related. These results dovetail earlier work that focused on neural and physiological indices of language processing. These studies have consistently shown deeper processing of words when they are presented in self-related in comparison to sender-related contexts (Herbert et al., Citation2011a; Herbert et al., Citation2011b; Fields & Kuperberg, Citation2012, Citation2015; Bayer et al., Citation2017; Weis & Herbert, Citation2017). When sender-related, emotional words did not reliably influence control processes in our study, a finding consistent with literature on perspective taking and empathy showing that humans have an egocentric bias, which is stronger under time pressure (Epley, Keysar, Van Boven, & Gilovich, Citation2004), and which is costly to override (Cameron et al., Citation2019; Hutcherson, Bushong, & Rangel, Citation2015; Lockwood et al., Citation2017). However, a closer investigation of the findings in Experiment 2 suggest that this account cannot completely explain the entire pattern of results observed. As illustrated in , participants in the sender-related context actually also showed a general increase in conflict adaptation irrespective of emotion. This pattern was not predicted. One could speculate that cognitively taxing process of perspective taking renders the primary conflict task more difficult at a subjective level, which in turn modulates reactive control processes (van Steenbergen, Band, & Hommel, Citation2015). However this speculation remains to be tested in future studies. Either way, our results highlight the potential of adding a self-relevance manipulation in the context of a cognitive control task with emotional stimuli. This further extends earlier literature that has studied the influence of other person’s errors (Koban & Pourtois, Citation2014) and conflict (e.g. Liepelt, Wenke, Fischer, & Prinz, Citation2011, Citation2013; Spapé & Ravaja, Citation2016; Winkel et al., Citation2009) on cognitive control outside the context of an emotional manipulation.
Figure 4. Conflict adaptation effects measured in RT (ms) after neutral and emotional previous-trial words, separate per experiment and levels of relevance. Error bars represent standard errors of the within-subject conflict adaptation effects (difference score).
There are a few limitations of the present study we mention here. First, although the stimuli were selected on norm scores providing measures of valence and arousal, we did not independently measure word-elicited changes in valence and arousal levels in our sample of participants. Second, we did not give particular instructions about how to interpret the different pronouns in our task, following earlier work (Herbert et al., Citation2011b). Future studies could investigate the effect of framing, for example by instructing participants to imagine a stranger versus a close other when reading sender-related pronoun-noun combinations (Bayer et al., Citation2017).
To conclude, our study revealed for the first time that phasic arousal increases conflict adaptation when using positive and negative emotional versus neutral words embedded in the cognitive control task itself. This effect was observed in Experiment 1 and was replicated in a self-relevant context in Experiment 2. In contrast, when emotional words were not self-relevant, conflict adaptation was no longer modulated by emotional content. These findings not only show how phasic emotional arousal influences control processes, they also demonstrate that self-referential processing can modulate it. This work provides interesting avenues for future research, for example to test independent and interactive effects of empathy, emotional processing, and cognitive control. The paradigm we developed here might also be used to identify aberrations of these functions in a wide range of psychiatric disorders including depression, autism, and sociopathy (e.g. Brazil, van Dongen, Maes, Mars, & Baskin-Sommers, Citation2018; McTeague, Goodkind, & Etkin, Citation2016; Nuske, Vivanti, & Dissanayake, Citation2013).
Data accessibility
Task, data, and analysis scripts are available online at: https://osf.io/8rmsp/.
Supplementary_Material.doc
Download MS Word (38.3 KB)Acknowledgements
We thank Isabelle Blok, Zala Reppman, Bas Rexwinkel, and Nurseda Risvanoglu for their help with data acquisition.
Disclosure statement
No potential conflict of interest was reported by the author(s).
References
- Baron-Cohen, S., & Wheelwright, S. (2004). The empathy quotient: An investigation of adults with Asperger syndrome or high functioning autism, and normal sex differences. Journal of Autism and Developmental Disorders, 34(2), 163–175.
- Bayer, M., Ruthmann, K., & Schacht, A. (2017). The impact of personal relevance on emotion processing: Evidence from event-related potentials and pupillary responses. Social Cognitive and Affective Neuroscience, 12(9), 1470–1479. doi: 10.1093/scan/nsx075
- Botvinick, M. M. (2007). Conflict monitoring and decision making: Reconciling two perspectives on anterior cingulate function. Cognitive, Affective, & Behavioral Neuroscience, 7(4), 356–366. doi: 10.3758/CABN.7.4.356
- Botvinick, M. M., Braver, T. S., Barch, D. M., Carter, C. S., & Cohen, J. D. (2001). Conflict monitoring and cognitive control. Psychological Review, 108(3), 624–652.
- Braem, S., Bugg, J. M., Schmidt, J. R., Crump, M. J. C., Weissman, D. H., Notebaert, W., & Egner, T. (2019). Measuring adaptive control in conflict tasks. Trends in Cognitive Sciences, https://doi.org/10.1016/j.tics.2019.07.002
- Braem, S., Verguts, T., Roggeman, C., & Notebaert, W. (2012). Reward modulates adaptations to conflict. Cognition, 125(2), 324–332. doi: 10.1016/j.cognition.2012.07.015
- Brazil, I. A., van Dongen, J. D. M., Maes, J. H. R., Mars, R. B., & Baskin-Sommers, A. R. (2018). Classification and treatment of antisocial individuals: From behavior to biocognition. Neuroscience and Biobehavioral Reviews, 91, 259–277. doi: 10.1016/j.neubiorev.2016.10.010
- Brown, G. G., Kindermann, S. S., Siegle, G. J., Granholm, E., Wong, E. C., & Buxton, R. B. (1999). Brain activation and pupil response during covert performance of the Stroop color word task. Journal of the International Neuropsychological Society, 5(4), 308–319.
- Brown, S. B. R. E., van Steenbergen, H., Kedar, T., & Nieuwenhuis, S. (2014). Effects of arousal on cognitive control: Empirical tests of the conflict-modulated Hebbian-learning hypothesis. Frontiers in Human Neuroscience, 8, 23. https://doi.org/10.3389/fnhum.2014.00023
- Brysbaert, M., Stevens, M., De Deyne, S., Voorspoels, W., & Storms, G. (2014). Norms of age of acquisition and concreteness for 30,000 Dutch words. Acta Psychologica, 150, 80–84. doi: 10.1016/j.actpsy.2014.04.010
- Cameron, C. D., Hutcherson, C. A., Ferguson, A. M., Scheffer, J. A., Hadjiandreou, E., & Inzlicht, M. (2019). Empathy is hard work: People choose to avoid empathy because of its cognitive costs. Journal of Experimental Psychology. General, 148(6), 962–976. doi: 10.1037/xge0000595
- Cohen, N., & Henik, A. (2012). Do irrelevant emotional stimuli impair or improve executive control? Frontiers in Integrative Neuroscience, 6, 33. doi: 10.3389/fnint.2012.00033
- Demanet, J., Liefooghe, B., & Verbruggen, F. (2011). Valence, arousal, and cognitive control: A voluntary task-switching study. Frontiers in Psychology, 2, 336. doi: 10.3389/fpsyg.2011.00336
- de Rover, M., Brown, S. B. R. E., Band, G. P., Giltay, E. J., van Noorden, M. S., van der Wee, N. J. A., & Nieuwenhuis, S. (2015). Beta receptor-mediated modulation of the oddball P3 but not error-related ERP components in humans. Psychopharmacology, 232(17), 3161–3172. doi: 10.1007/s00213-015-3966-2
- Dignath, D., Janczyk, M., & Eder, A. B. (2017). Phasic valence and arousal do not influence post-conflict adjustments in the Simon task. Acta Psychologica, 174, 31–39. doi: 10.1016/j.actpsy.2017.01.004
- Dreisbach, G., & Fischer, R. (2012). Conflicts as aversive signals. Brain and Cognition, 78(2), 94–98. doi: 10.1016/j.bandc.2011.12.003
- Dreisbach, G., & Fischer, R. (2015). Conflicts as aversive signals for control adaptation. Current Directions in Psychological Science, 24(4), 255–260. doi: 10.1177/0963721415569569
- Dreisbach, G., Fröber, K., Berger, A., & Fischer, R. (2019). Unexpected conflict signals loom larger in a positive context: Evidence from context specific control adjustments. Journal of Experimental Psychology. Learning, Memory, and Cognition, 45(8), 1398–1409. doi: 10.1037/xlm0000662
- Duthoo, W., Abrahamse, E. L., Braem, S., Boehler, C. N., & Notebaert, W. (2014). The heterogeneous world of congruency sequence effects: An update. Frontiers in Psychology, 5, 1001. doi: 10.3389/fpsyg.2014.01001
- Epley, N., Keysar, B., Van Boven, L., & Gilovich, T. (2004). Perspective taking as egocentric anchoring and adjustment. Journal of Personality and Social Psychology, 87(3), 327–339. doi: 10.1037/0022-3514.87.3.327
- Eriksen, B. A., & Eriksen, C. W. (1974). Effects of noise letters upon the identification of a target letter in a nonsearch task. Perception & Psychophysics, 16(1), 143–149. doi: 10.3758/BF03203267
- Fields, E. C., & Kuperberg, G. R. (2012). It’s all about you: An ERP study of emotion and self-relevance in discourse. NeuroImage, 62(1), 562–574. doi: 10.1016/j.neuroimage.2012.05.003
- Fields, E. C., & Kuperberg, G. R. (2015). Dynamic effects of self-relevance and task on the neural processing of emotional words in context. Frontiers in Psychology, 6, 2003. https://doi.org/10.3389/fpsyg.2015.02003
- Fischer, R., Ventura-Bort, C., Hamm, A., & Weymar, M. (2018). Transcutaneous vagus nerve stimulation (tVNS) enhances conflict-triggered adjustment of cognitive control. Cognitive, Affective & Behavioral Neuroscience, 18(4), 680–693. doi: 10.3758/s13415-018-0596-2
- Gratton, G., Coles, M. G., & Donchin, E. (1992). Optimizing the use of information: Strategic control of activation of responses. Journal of Experimental Psychology: General, 121(4), 480–506.
- Herbert, C., Herbert, B. M., Ethofer, T., & Pauli, P. (2011a). His or mine? The time course of self-other discrimination in emotion processing. Social Neuroscience, 6(3), 277–288. doi:10.1080/17470919.2010.523543.
- Herbert, C., Pauli, P., & Herbert, B. M. (2011b). Self-reference modulates the processing of emotional stimuli in the absence of explicit self-referential appraisal instructions. Social Cognitive and Affective Neuroscience, 6(5), 653–661. doi:10.1093/scan/nsq082.
- Herbert, C., Sfärlea, A., & Blumenthal, T. (2013). Your emotion or mine: Labeling feelings alters emotional face perception-an ERP study on automatic and intentional affect labeling. Frontiers in Human Neuroscience, 7, 378. doi: 10.3389/fnhum.2013.00378
- Hommel, B., Proctor, R. W., & Vu, K.-P. L. (2004). A feature-integration account of sequential effects in the Simon task. Psychological Research, 68(1), 1–17. doi: 10.1007/s00426-003-0132-y
- Hutcherson, C. A., Bushong, B., & Rangel, A. (2015). A neurocomputational model of altruistic choice and its implications. Neuron, 87(2), 451–462. doi: 10.1016/j.neuron.2015.06.031
- Inzlicht, M., Bartholow, B. D., & Hirsh, J. B. (2015). Emotional foundations of cognitive control. Trends in Cognitive Sciences, 19(3), 126–132. doi: 10.1016/j.tics.2015.01.004
- Kanske, P. (2012). On the influence of emotion on conflict processing. Frontiers in Integrative Neuroscience, 6, 42. doi: 10.3389/fnint.2012.00042
- Kanske, P., & Kotz, S. A. (2010). Modulation of early conflict processing: N200 responses to emotional words in a flanker task. Neuropsychologia, 48(12), 3661–3664. doi: 10.1016/j.neuropsychologia.2010.07.021
- Kanske, P., & Kotz, S. A. (2011a). Emotion triggers executive attention: Anterior cingulate cortex and amygdala responses to emotional words in a conflict task. Human Brain Mapping, 32(2), 198–208. doi: 10.1002/hbm.21012
- Kanske, P., & Kotz, S. A. (2011b). Positive emotion speeds up conflict processing: ERP responses in an auditory Simon task. Biological Psychology, 87(1), 122–127. doi: 10.1016/j.biopsycho.2011.02.018
- Kanske, P., & Kotz, S. A. (2011c). Conflict processing is modulated by positive emotion: ERP data from a flanker task. Behavioural Brain Research, 219(2), 382–386. doi: 10.1016/j.bbr.2011.01.043
- Koban, L., & Pourtois, G. (2014). Brain systems underlying the affective and social monitoring of actions: An integrative review. Neuroscience and Biobehavioral Reviews, 46(Pt 1), 71–84. doi: 10.1016/j.neubiorev.2014.02.014
- Kuhbandner, C., & Zehetleitner, M. (2011). Dissociable effects of valence and arousal in adaptive executive control. PloS One, 6(12), e29287. doi: 10.1371/journal.pone.0029287
- Laeng, B., Ørbo, M., Holmlund, T., & Miozzo, M. (2011). Pupillary Stroop effects. Cognitive Processing, 12(1), 13–21. doi: 10.1007/s10339-010-0370-z
- Lakens, D., & Evers, E. R. K. (2014). Sailing from the seas of Chaos into the Corridor of stability: Practical recommendations to increase the informational value of studies. Perspectives on Psychological Science: A Journal of the Association for Psychological Science, 9(3), 278–292. doi: 10.1177/1745691614528520
- Liepelt, R., Wenke, D., & Fischer, R. (2013). Effects of feature integration in a hands-crossed version of the social Simon paradigm. Psychological Research, 77(2), 240–248. doi: 10.1007/s00426-012-0425-0
- Liepelt, R., Wenke, D., Fischer, R., & Prinz, W. (2011). Trial-to-trial sequential dependencies in a social and non-social Simon task. Psychological Research, 75(5), 366–375. doi: 10.1007/s00426-010-0314-3
- Lockwood, P. L., Hamonet, M., Zhang, S. H., Ratnavel, A., Salmony, F. U., Husain, M., & Apps, M. A. J. (2017). Prosocial apathy for helping others when effort is required. Nature Human Behaviour, 1(7), 0131. doi: 10.1038/s41562-017-0131
- McTeague, L. M., Goodkind, M. S., & Etkin, A. (2016). Transdiagnostic impairment of cognitive control in mental illness. Journal of Psychiatric Research, 83, 37–46. doi: 10.1016/j.jpsychires.2016.08.001
- Meixner, F., & Herbert, C. (2018). Whose emotion is it? Measuring self-other discrimination in romantic relationships during an emotional evaluation paradigm. PloS One, 13(9), e0204106. doi: 10.1371/journal.pone.0204106
- Moors, A., De Houwer, J., Hermans, D., Wanmaker, S., van Schie, K., Van Harmelen, A.-L., … Brysbaert, M. (2013). Norms of valence, arousal, dominance, and age of acquisition for 4,300 Dutch words. Behavior Research Methods, 45(1), 169–177. doi: 10.3758/s13428-012-0243-8
- Nuske, H. J., Vivanti, G., & Dissanayake, C. (2013). Are emotion impairments unique to, universal, or specific in autism spectrum disorder? A comprehensive review. Cognition & Emotion, 27(6), 1042–1061. doi: 10.1080/02699931.2012.762900
- Padmala, S., Bauer, A., & Pessoa, L. (2011). Negative emotion impairs conflict-driven executive control. Frontiers in Psychology, 2, 192. doi: 10.3389/fpsyg.2011.00192
- Scherbaum, S., Fischer, R., Dshemuchadse, M., & Goschke, T. (2011). The dynamics of cognitive control: Evidence for within-trial conflict adaptation from frequency-tagged EEG. Psychophysiology, 48(5), 591–600. doi: 10.1111/j.1469-8986.2010.01137.x
- Schmidt, J. R., & De Houwer, J. (2011). Now you see it, now you don’t: Controlling for contingencies and stimulus repetitions eliminates the Gratton effect. Acta Psychologica, 138(1), 176–186. doi: 10.1016/j.actpsy.2011.06.002
- Schmidt, J. R., & Weissman, D. H. (2014). Congruency sequence effects without feature integration or contingency learning confounds. PloS One, 9(7), e102337. doi: 10.1371/journal.pone.0102337
- Schuch, S., & Koch, I. (2015). Mood states influence cognitive control: The case of conflict adaptation. Psychological Research, 79(5), 759–772. doi: 10.1007/s00426-014-0602-4
- Schuch, S., Zweerings, J., Hirsch, P., & Koch, I. (2017). Conflict adaptation in positive and negative mood: Applying a success-failure manipulation. Acta Psychologica, 176, 11–22. doi: 10.1016/j.actpsy.2017.03.005
- Simon, J. R., & Rudell, A. P. (1967). Auditory S-R compatibility: The effect of an irrelevant cue on information processing. The Journal of Applied Psychology, 51(3), 300–304.
- Spapé, M. M., & Ravaja, N. (2016). Not my problem: Vicarious conflict adaptation with human and virtual co-actors. Frontiers in Psychology, 7, 606. doi: 10.3389/fpsyg.2016.00606
- Spielberger, C. D., Gorsuch, R. L., & Lushene, R. E. (1970). Manual for the state-trait anxiety inventory. Palo Alto, CA: Consulting Psychologists Press.
- Stroop, J. R. (1935). Studies of interference in serial verbal reactions. Journal of Experimental Psychology, 18(6), 643–662. doi: 10.1037/h0054651
- Stürmer, B., Nigbur, R., Schacht, A., & Sommer, W. (2011). Reward and punishment effects on error processing and conflict control. Frontiers in Psychology, 2, 335. doi: 10.3389/fpsyg.2011.00335
- van Bochove, M. E., De Taeye, L., Raedt, R., Vonck, K., Meurs, A., Boon, P., … Verguts, T. (2018). Reduced distractor interference during vagus nerve stimulation. International Journal of Psychophysiology: Official Journal of the International Organization of Psychophysiology, 128, 93–99. doi: 10.1016/j.ijpsycho.2018.03.015
- van der Wel, P., & van Steenbergen, H. (2018). Pupil dilation as an index of effort in cognitive control tasks: A review. Psychonomic Bulletin & Review, 25(6), 2005–2015. doi: 10.3758/s13423-018-1432-y
- van Steenbergen, H. (2015). Affective modulation of cognitive control: A biobehavioral perspective. In G. Gendolla, M. Tops, & S. Koole (Eds.), Handbook of biobehavioral approaches to self-regulation (pp. 89–107). New York, NY: Springer.
- van Steenbergen, H., & Band, G. P. H. (2013). Pupil dilation in the Simon task as a marker of conflict processing. Frontiers in Human Neuroscience, 7, 215. doi: 10.3389/fnhum.2013.00215
- van Steenbergen, H., Band, G. P. H., & Hommel, B. (2009). Reward counteracts conflict adaptation. Evidence for a role of affect in executive control. Psychological Science, 20(12), 1473–1477. doi: 10.1111/j.1467-9280.2009.02470.x
- van Steenbergen, H., Band, G. P. H., & Hommel, B. (2010). In the mood for adaptation: How affect regulates conflict-driven control. Psychological Science, 21(11), 1629–1634. doi: 10.1177/0956797610385951
- van Steenbergen, H., Band, G. P. H., & Hommel, B. (2015). Does conflict help or hurt cognitive control? Initial evidence for an inverted U-shape relationship between perceived task difficulty and conflict adaptation. Frontiers in Psychology, 6, 974. doi: 10.3389/fpsyg.2015.00974
- van Steenbergen, H., Band, G. P. H., Hommel, B., Rombouts, S. A. R. B., & Nieuwenhuis, S. (2015). Hedonic hotspots regulate cingulate-driven adaptation to cognitive demands. Cerebral Cortex, 25(7), 1746–1756. doi: 10.1093/cercor/bht416
- van Steenbergen, H., Weissman, D. H., Stein, D. J., Malcolm-Smith, S., & van Honk, J. (2017). More pain, more gain: Blocking the opioid system boosts adaptive cognitive control. Psychoneuroendocrinology, 80, 99–103. doi: 10.1016/j.psyneuen.2017.03.002
- Verguts, T., & Notebaert, W. (2009). Adaptation by binding: A learning account of cognitive control. Trends in Cognitive Sciences, 13(6), 252–257. doi: 10.1016/j.tics.2009.02.007
- Warren, C. M., Tona, K. D., Ouwerkerk, L., van Paridon, J., Poletiek, F., van Steenbergen, H., … Nieuwenhuis, S. (2019). The neuromodulatory and hormonal effects of transcutaneous vagus nerve stimulation as evidenced by salivary alpha amylase, salivary cortisol, pupil diameter, and the P3 event-related potential. Brain Stimulation, 12(3), 635–642. doi: 10.1016/j.brs.2018.12.224
- Weis, P. P., & Herbert, C. (2017). Bodily reactions to emotional words referring to own versus other people’s emotions. Frontiers in Psychology, 8, 1277. doi: 10.3389/fpsyg.2017.01277
- Winkel, J., Wijnen, J. G., Ridderinkhof, K. R., Groen, I. I. A., Derrfuss, J., Danielmeier, C., & Forstmann, B. U. (2009). Your conflict matters to me! Behavioral and neural manifestations of control adjustment after self-experienced and observed decision-conflict. Frontiers in Human Neuroscience, 3, 57. doi: 10.3389/neuro.09.057.2009
- Yamaguchi, M., & Nishimura, A. (2019). Modulating proactive cognitive control by reward: Differential anticipatory effects of performance-contingent and non-contingent rewards. Psychological Research, 83(2), 258–274. doi: 10.1007/s00426-018-1027-2
- Zeng, Q., Qi, S., Li, M., Yao, S., Ding, C., & Yang, D. (2017). Enhanced conflict-driven cognitive control by emotional arousal, not by valence. Cognition & Emotion, 31(6), 1083–1096. https://doi.org/10.1080/02699931.2016.1189882