
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
When it comes to measuring cognitive control and inhibition, the antisaccade paradigm is a popular task to apply. Usually, simple, perceptually and affectively neutral stimuli, e.g. white circles, are used. Recently, researchers also employed a version of the paradigm displaying emotional faces. Differences in cognitive processing due to stimulus size and emotional valence have not been investigated yet. Thus, in the present study, we applied both versions of the antisaccade paradigm in a healthy sample. In addition, we used scrambled faces to control for stimulus size and emotional valence. We hypothesised slower reaction times and higher error rates for emotional face stimuli compared to circular and scrambled ones as well as significant differences between individual emotions. In contrast to our hypotheses, results showed faster reaction times fewer errors for emotional faces compared to circular and scrambled stimuli. Furthermore, ANOVA models showed no meaningful differences between different emotions. Our study shows specific patterns in inhibitory control due to stimulus size and valence in an antisaccade eye-tracking task.
The data that support the findings of this study are openly available in open science framework at https://osf.io/zqh8a, DOI:10.17605/OSF.IO/ZQH8A.
Introduction
A major research topic in psychology and neuroscience is the investigation of inhibitory control and emotional processing. In particular, research aimed to understand how emotional content affects inhibitory control performance. One frequently used paradigm is the antisaccade task (e.g. Aichert et al., Citation2013; Chen et al., Citation2014; García-Blanco et al., Citation2013). During this task, subjects are instructed to look opposite of a suddenly appearing peripheral stimulus and avoid eye-movements towards it. In contrast, prosaccades only demand alertness and gaze reorienting, as stimuli just need to be fixated as fast as possible. In general, one can observe slower reaction times (RTs) and higher error rates (ERs) during antisaccades compared to prosaccades. RTs represent the latency between stimulus onset and saccadic initiation (Antoniades et al., Citation2013). In the classic version of the paradigm, simple visual cues (e.g. circles; ∼1° of visual angle) serve as stimuli, whereas in the emotional version emotional faces (∼8°×6° of visual angle) are displayed. Both versions of the paradigm are used with the implicit notion that they both measure inhibitory control, ignoring differences in size, emotional valence, and complexity.
Nevertheless, emotional stimuli have repeatedly been shown to undermine inhibitory control (Rebetez et al., Citation2015; Schimmack & Derryberry, Citation2005). Earlier studies found that the presentation of emotional stimuli prolonged RTs (Herbert & Sütterlin, Citation2011) and interrupted ongoing cognitive control processes (Verbruggen & De Houwer, Citation2007). Inhibition was also more difficult in an emotional than in a non-emotional go/no-go task (Schulz et al., Citation2007). On the other hand, emotional stimuli improved the capacity to resist the effect of proactive interference (Levens & Phelps, Citation2008). Thus, emotional content either impairs or facilitates inhibitory control, depending on the underlying cognitive function. Contrasting effects might be due to different cognitive mechanisms being at work (Friedman & Miyake, Citation2004), namely either pre-potent response inhibition (Rebetez et al., Citation2015; Schulz et al., Citation2007), resistance to distractor interference (Verbruggen & De Houwer, Citation2007), or resistance to proactive interference (Levens & Phelps, Citation2008: Rebetez et al., Citation2015). Antisaccades mainly involve the inhibition of pre-potent responses (Noorani & Carpenter, Citation2013).
Therefore, the first objective of this study is to investigate whether there is an interference effect of emotional stimuli on response inhibition (Herbert & Sütterlin, Citation2011; Rebetez et al., Citation2015; Verbruggen & De Houwer, Citation2007) in the antisaccade paradigm. Former studies mainly applied RT tasks, which involve motoric processes. Thus, cognitive and motoric processes are not separable from each other. Therefore, we plan to measure response inhibition in different versions of the antisaccade paradigm with eye tracking technology. To date, no study compared inhibitory control performance between different kinds of stimuli in the antisaccade task. If emotional faces indeed negatively influence response inhibition, we would await strong effects of emotional valence on inhibitory control performance, which are diminished in less complex or smaller, geometrical stimuli.
Emotional valence, stimulus size and complexity constitute important factors overlooked so far in antisaccade research. Nevertheless, research indicates that task-evoked potentials are directly related to stimulus size (Busch et al., Citation2004). Simple geometrical shapes, e.g. circles and squares with sizes of 1.5° (small), 4° (medium) or 8° (large) of visual angle served as stimuli in a choice reaction task here. The authors observed the fastest RTs for medium-sized stimuli, while small and big ones did not differ from each other. Fischer and Weber (Citation1997) found only modest effects of stimulus size in the antisaccade task, when investigating varying sizes of circular stimuli. More complex stimuli e.g. emotional faces have never been compared to neutral geometrical stimuli. Nevertheless, there is reason to assume that humans process faces and inanimate objects differently. Carmel and Bentin (Citation2002) demonstrated the domain specificity of the visual mechanism implicated in processing faces, while a more general visual processor accomplishes the processing of other objects, which is sensitive to manipulations and attention. Moreover, evoked potentials are stronger and faster to faces than to objects (Jeffreys, Citation1996).
Thus, our second goal is to prove that inhibitory control differences are directly related to stimulus size, complexity as well as emotional valence. Therefore, we compare performance in an emotional antisaccade task to a classic version and one version including scrambled faces, which are the same size as the emotional faces, but the displayed emotion is not recognisable. Indeed, research already proved an influence of emotional valence on eye-movement patterns. For example, angry faces are detected faster than happy or sad faces in crowds (e.g. Calvo et al., Citation2006). In general, emotional content was found to attract more attention than neutral content in dot-probe tasks (Mogg & Bradley, Citation1999) and cueing paradigms (Khalid et al., Citation2017).
According to earlier findings concerning inhibition performance during the processing of emotional stimuli, we would thus (1) await meaningful differences between neutral and emotional faces during antisaccade execution. Moreover, emotional faces are fixated faster than neutral, scrambled or geometrical stimuli as measured by prosaccades. Furthermore, as emotional content attracts more attention than neutral content, we hypothesise (2) slower RTs and higher ERs in the emotional than in the classic paradigm. As we assume this to be emotion-specific, RTs are furthermore faster in the scrambled paradigm, but the fastest RTs will be observed in the classic paradigm. (3) For prosaccades, we would assume faster RTs for emotional faces compared to scrambled or geometrical stimuli, as they attract more attention than neutral content.
Method
Participants and procedure
Thirty-seven volunteers (Mage = 21.78, SDage = 1.89) participated with informed consent in the present study, which was conducted in line with the guidelines of the Ethics Committee of the University of the first author. They received no compensation, as the experimental session was part of a psychology seminar within their bachelor studies. Exclusion criteria were visual impairment corrected with glasses or any current psychological problem. To rule out the presence of mental disorders in controls, we applied a screening questionnaire (DIA-X-SSQ) (Wittchen & Perkonigg, Citation1996). None of the subjects suffered from a relevant disease. Participants were individually tested in a quiet laboratory. All subjects (29 female) were entered into the final analyses.
Antisaccade tasks
In the classic antisaccade paradigm, a neutral cue (white circle, 1° diameter, line thickness 0.3°) was presented in the centre of the display for a variable interval of 1000 – 2000ms (Tobii TX-300 screen-unit; resolution: 1920 × 1080; refresh rate: 60 Hz; viewing distance: 60 cm; Dell Precision T-5610). Viewing distance was kept stable via chin rest. After the variable interval, this cue moved either 12° left or right from the centre. After another 1000 ms, it moved back to the centre (no gap or overlap). Each position appeared equally often (randomised order). Subjects were instructed to fixate on the central cue. As soon as it appeared on another location, subjects were asked to fixate on the mirror position of the respective target as fast and accurate as possible. As a second condition, prosaccades were performed. Stimulus presentation was the same as in antisaccades. However, subjects were instructed to follow the stimulus as fast as possible and fixate on it. Pro- and antisaccades were presented in separate blocks.
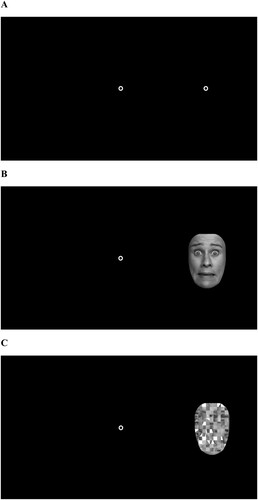
In addition, we applied the same paradigm with emotional faces (happy, fearful, sad, and neutral). Faces were taken from the FACES database (Ebner et al., Citation2010). Non-facial features (e.g. hair, neck) were removed, and faces were resized to a size of 6°x8°. For each expression, five black and white pictures were used. The stimuli were balanced for gender (50% male faces); faces were Caucasian. Emotions were presented randomised within blocks. As for the selection of individual emotions for the emotional antisaccade task, we have been guided by previous studies applying the task (Aichert et al., Citation2013; Chen et al., Citation2014; García-Blanco et al., Citation2013). We decided to use the neutral, one positive (happy) and one negative (sad) emotion as well as one that signals danger (fear). Therefore, it would be possible to evaluate whether any of these categories is able to facilitate inhibitory control as shown in earlier studies (Baumeister et al., Citation2001; Rebetez et al., Citation2015). As a third condition, we used scrambled faces in order to control for stimulus size and complexity. A Photoshop filter was applied to make those emotional faces, displaying either a neutral, happy, sad or a fearful emotion, unrecognisable (see for stimuli presentation).
Figure 1. Stimuli as presented in the three different pro- and antisaccade paradigms applying classic circles (A), emotional faces (B) and scrambled faces (C) as target cues.
All blocks were preceded by 10 practice trials, the data from which were discarded. After each 75 trials, subjects got the possibility to take a break. The order of the blocks was randomised across participants. In total, we recorded over 100,000 saccades. The experimental session took about 3–3.5 h per participant, depending on individual breaks within and between blocks. We recorded the following blocks:
Block 1 (classic prosaccades): 150 trials
Block 2 (classic antisaccades): 375 trials
Block 3 (scrambled faces prosaccades): 150 trials
Block 4 (scrambled faces antisaccades): 375 trials
Block 5–6 (emotional faces prosaccades): 600 trials (150 per emotion)
Block 7–10 (emotional faces antisaccades): 1500 trials (375 per emotion)
As prosaccades serve as a control condition (cf. Noorani & Carpenter, Citation2013) and our main research question is more concerned with antisaccades, we decided to measure only 150 repetitions of prosaccades per paradigm. Moreover, we wanted to ensure that the experiment would not take more than 3 h, as participants fatigue and data quality diminishes over time.
Central performance indicators for pro- and antisaccades were (1) saccadic RTs and (2) ERs (saccades in direction of the target cue or corrective saccades). The applicability of those parameters has been validated both in healthy and clinical samples (Ettinger et al., Citation2003). Saccadic and antisaccadic eye-movements were defined by criteria of amplitude ≥1.5°, velocity ≥30°/s and latency between 100 and 900 ms (e.g. Ettinger et al., Citation2003). To ensure spatial accuracy, we only included saccades, which landed on the target cue (prosaccade) or on its exact mirror position (antisaccade). Artifact-affected trials (e.g. blink saccades, measurement failures) were removed from analyses (0.02% of trials in the classic paradigm; 0.02% of trials in the scrambled paradigm; 0.02% of trials in the emotional paradigm).
RTs were calculated only on correct responses (99.85% of prosaccade trials and 86.81% of antisaccade trials in the classic paradigm; 99.39% of prosaccade trials and 89.06% of antisaccade trials in the scrambled paradigm; 98.58% of prosaccade trials and 93.32% of antisaccade trials in the emotional paradigm). Binocular eye-movement registration was carried out using an infrared-based eye-tracking system (Tobii TX-300, Tobii AB, Danderyd, Sweden), which allows for quantification of horizontal eye-movements up to +/−25° with a sampling rate of 300 Hz. Before data acquisition, we calibrated and validated the eye-tracker with a 5-point calibration. Artifact correction and calculation of saccade parameters was performed with a custom developed interactive analysis software. This software allowed evaluating the RTs based on a linear regression as well as eye-movement interval classification. Within the tool, the automated detection was adjusted manually and directly exported into excel files for further statistical analyses.
Statistical analyses
For comparing the three paradigms with one another, we applied a 2 (pro- vs. antisaccades) x 3 (classic vs. scrambled vs. emotional) repeated measures ANOVA model to mean RTs as well as ERs of each of the three paradigms. Another 2 (pro- vs. antisaccades) x 4 (neutral vs. happy vs. sad vs. fearful) repeated measures ANOVA model was then conducted to analyse mean RTs and ERs within the emotional paradigm and thus be able to evaluate performance on an emotion-specific level. In case of significant main effects, we report Bonferroni-corrected pairwise comparisons. For the paradigms, Bonferroni corrections accounted for three comparisons, while for the different emotions, corrections accounted for 6 comparisons. Sphericity was tested using Mauchly’s test and in cases of deviance from sphericity, we adjusted the degrees of freedom using the Greenhouse-Geisser correction. Effect sizes are indicated by partial eta squared. Alpha levels were set at 0.05. Results are reported with original df and corrected p-values. Data were analysed using SPSS 25.
A priori power analysis with G*Power (Faul et al., Citation2009) showed that a sample size of N = 16 would be sufficient to detect an effect of f = 0.44 with a statistical power of 1−β = 0.95 and α = 0.05 in all of the computed repeated measures ANOVA models (within subjects). We only found one effect size for the comparison between different emotional stimuli of = 0.16 (Rebetez et al., Citation2015). Neither Fischer and Weber (Citation1997) nor Verbruggen and De Houwer (Citation2007) did report effect sizes in their publications. Moreover, 3,150 replications per participant and 37 participants should yield sufficiently precise estimates and good confidence interval coverage.
Results
Mean scores and standard deviations on measures for pro- and antisaccade performance are presented in . For comparing the classic, scrambled, and emotional paradigm, we applied a 2 × 3 repeated measures ANOVA model to mean RTs of the three paradigms. A significant main effect for condition (F[1, 36] = 440.69, p < 0.01, = 0.92) as well as paradigm (F[2, 72] = 115.94, p < 0.01,
= 0.76) could be obtained; the interaction also reached significance (F[2, 72] = 10.65, p < 0.01,
= 0.23). Pairwise comparisons showed that RTs were faster in prosaccade than in antisaccade trials (MD = −76.27, SE = 3.363, p < 0.01). Moreover, as can be seen in , RTs were faster for emotional than scrambled (MD = −18.78, SE = 1.79, p < 0.01) or classic stimuli (MD = −32.40, SE = 2.55, p < 0.01).
Figure 2. RTs and ERs across different paradigms (A, B) and emotions within the emotional antisaccade task (C, D).
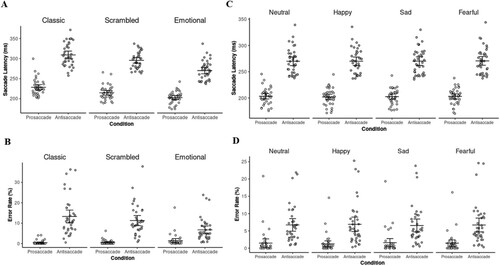
Table 1. Descriptive statistics of antisaccade paradigms and for different emotions.
The same model was conducted for ERs (in %), providing a significant main effect for condition (F[1, 36] = 87.76, p < 0.01, = 0.71) as well as for paradigm (F[2, 72] = 10.72, p < 0.01,
= 0.23) alongside an interaction effect (F[2, 72] = 18.87, p < 0.01,
= 0.34). Pairwise comparisons showed that ERs were lower in prosaccade than in antisaccade trials (MD = −9.61, SE = 1.03, p < 0.01). Moreover, ERs were lower for emotional than scrambled (MD = −1.89 SE = 0.44, p < 0.01) or classic stimuli (MD = −2.80, SE = 0.75, p < 0.01).
We further performed a 2 × 4 repeated measures ANOVA model to evaluate the effect of emotional stimuli on task performance. For mean RTs within the emotional paradigm, this model provided a significant main effect for condition (F[1, 19] = 72.92, p < 0.01, = 0.79); neither a main effect of emotion nor an interaction was found, indicating no meaningful difference between emotions. The same accounted for ERs, only showing a significant main effect of condition (F[1, 19] = 23.59, p < 0.01,
= 0.55), but no main effect for emotion nor an interaction.
Discussion
The first goal of the current study was to examine the influence of emotional stimuli on inhibitory control in the antisaccade task, thus replicating earlier findings showing interference effects of emotional content on response inhibition. Our results showed no decline of inhibitory control performance in the emotional paradigm compared to the classic or scrambled one. Moreover, there was no meaningful difference between neutral and emotional faces stimuli. Our second goal was to show that inhibitory control performance is directly related to stimulus size, complexity as well as emotional valence. We found major differences concerning cognitive processing performance for both pro- and antisaccades, which speaks for a generally better and faster processing of emotional faces compared to scrambled faces or smaller circular stimuli. Contrary to our hypotheses and earlier research, bigger and more complex stimuli seem to facilitate inhibitory control in the antisaccade paradigm.
Thus, the distinction of the antisaccade task from other tasks applied in earlier studies is crucial. Contrasting results might be due to different cognitive mechanisms being at work (Friedman & Miyake, Citation2004), which is either pre-potent response inhibition (Rebetez et al., Citation2015; Schulz et al., Citation2007), resistance to distractor interference (Verbruggen & De Houwer, Citation2007), or resistance to proactive interference (Levens & Phelps, Citation2008: Rebetez et al., Citation2015). The antisaccade task mainly involves pre-potent response inhibition (Noorani & Carpenter, Citation2013). Moreover, all of the formerly applied tasks involve motoric movements for response execution. Thus, motoric and cognitive processes are not separable from each other. In contrast, the antisaccade task only involves eye-movement, which is not biased by motor-processes during cognitive processing. Moreover, as we used a blocked design, we were able to investigate the inhibition of pre-potent responses separate from other cognitive processes. In the stop signal or go-no/go task, stop and go trials are interleaved, and thus it is likely that not only inhibitory control, but also shifting might have been involved there (Friedman & Miyake, Citation2004).
Regarding the emotional antisaccade task, we found no difference in RTs during emotional antisaccade trials compared to neutral ones, providing no evidence that either positive or negative emotions interfere with the effectiveness of inhibitory control by capturing more attention than neutral faces. Our results show no emotion-specific response behaviour whatsoever, thus contrasting previous evidence (e.g. Herbert & Sütterlin, Citation2011; Rebetez et al., Citation2015). Furthermore, we contrast previous studies showing greater interference for negative than for positive stimuli (Baumeister et al., Citation2001; Rebetez et al., Citation2015).
More specifically, we observed slower RTs for smaller circular than for emotional stimuli. Even scrambled faces, which had the same size as the emotional faces, received slower RTs than emotional face stimuli. In addition, this effect was not only specific to antisaccades and inhibitory control, but also visible in prosaccades, which represent general attentional capacity. This result highlights that bigger and more complex stimuli, especially those that display emotions might facilitate inhibitory control, as they are detected faster and easier than smaller, geometrical stimuli. Our results are in line with earlier research showing that task-evoked potentials are directly related to stimulus size (Busch et al., Citation2004), i.e. bigger stimuli triggering more brain activity as well as faster and stronger responses to faces than objects (Jeffreys, Citation1996). Varying sizes of white circular stimuli only had modest effects on response inhibition (Fischer & Weber, Citation1997). This speaks for a special advantage for more complex stimuli, i.e. emotional faces, which is in line with research demonstrating a domain specificity for face processing (Carmel & Bentin, Citation2002; Jeffreys, Citation1996).
The current study adds to a growing body of research linking emotional processing and inhibitory control. Indeed, although previous studies found that emotional stimuli interfere with the ability to inhibit response execution (Herbert & Sütterlin, Citation2011; Schulz et al., Citation2007; Verbruggen & De Houwer, Citation2007) but facilitate resistance to proactive interference (Levens & Phelps, Citation2008), our results emphasised that this effect might be specific for certain paradigms including a motoric reaction during response execution. For eye tracking tasks like the antisaccade paradigm, emotional faces seem to facilitate inhibitory control, as faces of a certain size are detected faster and easier than smaller circular stimuli, and are thus easier to avoid. Moreover, when controlling for stimulus size and complexity by using scrambled faces, we find a strong effect that seems to be directly related to the emotional valence of the displayed face stimuli. Most of all, the current study results highlight the same interaction concerning stimulus properties (i.e. stimulus size, complexity, and emotional valence) in two different task properties (pro- and antisaccades).
Open practices
Materials and data associated with this research are available at https://osf.io/zqh8a.
Declarations of interest
None.
Acknowledgements
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors. We thank Stella Ilg and Tobias Holzinger for carrying out data collection in our eye-tracking lab. Furthermore, we thank Ulrich Ettinger for his helpful comments on this manuscript.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Aichert, D. S., Derntl, B., Wöstmann, N. M., Groß, J. K., Dehning, S., Cerovecki, A., Möller, H.-J., Habel, U., Riedel, M., & Ettinger, U. (2013). Intact emotion–cognition interaction in schizophrenia patients and first-degree relatives: Evidence from an emotional antisaccade task. Brain and Cognition, 82(3), 329–336. https://doi.org/https://doi.org/10.1016/j.bandc.2013.05.007
- Antoniades, C., Ettinger, U., Gaymard, B., Gilchrist, I., Kristjánsson, A., Kennard, C., Leigh, R. J., Noorani, I., Pouget, P., Smyrnis, N., Tarnowski, A., Zee, D. S., & Carpenter, R. H. S. (2013). An internationally standardised antisaccade protocol. Vision Research, 84, 1–5. https://doi.org/https://doi.org/10.1016/j.visres.2013.02.007
- Baumeister, R. F., Bratslavsky, E., Finkenauer, C., & Vohs, K. D. (2001). Bad is stronger than good. Review of General Psychology, 5(4), 323–370. https://doi.org/https://doi.org/10.1037/1089-2680.5.4.323
- Busch, N. A., Debener, S., Kranczioch, C., Engel, A. K., & Herrmann, C. S. (2004). Size matters: Effects of stimulus size, duration and eccentricity on the visual gamma-band response. Clinical Neurophysiology, 115(8), 1810–1820. https://doi.org/https://doi.org/10.1016/j.clinph.2004.03.015
- Calvo, M. G., Avero, P., & Lundqvist, D. (2006). Facilitated detection of angry faces: Initial orienting and processing efficiency. Cognition and Emotion, 20(6), 785–811. https://doi.org/https://doi.org/10.1080/02699930500465224
- Carmel, D., & Bentin, S. (2002). Domain specificity versus expertise: Factors influencing distinct processing of faces. Cognition, 83(1), 1–29. https://doi.org/https://doi.org/10.1016/S0010-0277(01)00162-7
- Chen, N. T., Clarke, P. J., Watson, T. L., MacLeod, C., & Guastella, A. J. (2014). Biased saccadic responses to emotional stimuli in anxiety: An antisaccade study. PloS One, 9(2), e86474. https://doi.org/https://doi.org/10.1371/journal.pone.0086474
- Ebner, N. C., Riediger, M., & Lindenberger, U. (2010). FACES—a database of facial expressions in young, middle-aged, and older women and men: Development and validation. Behavior Research Methods, 42(1), 351–362. https://doi.org/https://doi.org/10.3758/BRM.42.1.351
- Ettinger, U., Kumari, V., Crawford, T. J., Davis, R. E., Sharma, T., & Corr, P. J. (2003). Reliability of smooth pursuit, fixation, and saccadic eye movements. Psychophysiology, 40(4), 620–628. https://doi.org/https://doi.org/10.1111/1469-8986.00063
- Faul, F., Erdfelder, E., Buchner, A., & Lang, A. G. (2009). Statistical power analyses using G* power 3.1: Tests for correlation and regression analyses. Behavior Research Methods, 41(4), 1149–1160. https://doi.org/https://doi.org/10.3758/BRM.41.4.1149
- Fischer, B., & Weber, H. (1997). Effects of stimulus conditions on the performance of antisaccades in man. Experimental Brain Research, 116(2), 191–200. https://doi.org/https://doi.org/10.1007/PL00005749
- Friedman, N. P., & Miyake, A. (2004). The relations among inhibition and interference control functions: A latent-variable analysis. Journal of Experimental Psychology: General, 133(1), 101–135. https://doi.org/https://doi.org/10.1037/0096-3445.133.1.101
- García-Blanco, A. C., Perea, M., & Salmerón, L. (2013). Attention orienting and inhibitory control across the different mood states in bipolar disorder: An emotional antisaccade task. Biological Psychology, 94(3), 556–561. https://doi.org/https://doi.org/10.1016/j.biopsycho.2013.10.005
- Herbert, C., & Sütterlin, S. (2011). Response inhibition and memory retrieval of emotional target words: Evidence from an emotional stop-signal task. Journal of Behavioral and Brain Science, 1(3), 153–159. https://doi.org/https://doi.org/10.4236/jbbs.2011.13020
- Jeffreys, D. A. (1996). Evoked potential studies of face and object processing. Visual Cognition, 3(1), 1–38. https://doi.org/https://doi.org/10.1080/713756729
- Khalid, S., Horstmann, G., Ditye, T., & Ansorge, U. (2017). Measuring the emotion-specificity of rapid stimulus-driven attraction of attention to fearful faces: Evidence from emotion categorization and a comparison with disgusted faces. Psychological Research, 81(2), 508–523. https://doi.org/https://doi.org/10.1007/s00426-016-0743-8
- Levens, S. M., & Phelps, E. A. (2008). Emotion processing effects on interference resolution in working memory. Emotion, 8(2), 267–280. https://doi.org/https://doi.org/10.1037/1528-3542.8.2.267
- Mogg, K., & Bradley, B. P. (1999). Orienting of attention to threatening facial expressions presented under conditions of restricted awareness. Cognition & Emotion, 13(6), 713–740. https://doi.org/https://doi.org/10.1080/026999399379050
- Noorani, I., & Carpenter, R. H. S. (2013). Antisaccades as decisions: LATER model predicts latency distributions and error responses. European Journal of Neuroscience, 37(2), 330–338. https://doi.org/https://doi.org/10.1111/ejn.12025
- Rebetez, M. M. L., Rochat, L., Billieux, J., Gay, P., & Van der Linden, M. (2015). Do emotional stimuli interfere with two distinct components of inhibition? Cognition & Emotion, 29(3), 559–567. https://doi.org/https://doi.org/10.1080/02699931.2014.922054
- Schimmack, U., & Derryberry, D. E. (2005). Attentional interference effects of emotional pictures: Threat, negativity, or arousal? Emotion, 5(1), 55–66. https://doi.org/https://doi.org/10.1037/1528-3542.5.1.55
- Schulz, K. P., Fan, J., Magidina, O., Marks, D. J., Hahn, B., & Halperin, J. M. (2007). Does the emotional go/no-go task really measure behavioral inhibition? Convergence with measures on a non-emotional analog. Archives of Clinical Neuropsychology, 22(2), 151–160. https://doi.org/https://doi.org/10.1016/j.acn.2006.12.001
- Verbruggen, F., & De Houwer, J. (2007). Do emotional stimuli interfere with response inhibition? Evidence from the stop signal paradigm. Cognition & Emotion, 21(2), 391–403. https://doi.org/https://doi.org/10.1080/02699930600625081
- Wittchen, H. U., & Perkonigg, A. (1996). DIA-X SSQ. Swetz und zeitlinger. Frankfurt. Swetz Test Services.