
ABSTRACT
Long-term memory manages its contents to facilitate adaptive behaviour, amplifying representations of information relevant to current goals and expediting forgetting of information that competes with relevant memory traces. Both mnemonic selection and inhibition maintain congruence between the contents of long-term memory and an organism’s priorities. However, the capacity of these processes to modulate affective mnemonic representations remains ambiguous. Three empirical experiments investigated the consequences of mnemonic selection and inhibition on affectively charged and neutral mnemonic representations using an adapted retrieval practice paradigm. Participants encoded neutral cue words and affectively negative or neutral associates and then selectively retrieved a subset of these associates multiple times. The consequences of selection and inhibitory processes engaged during selective retrieval were evaluated on a final memory test in which recall for all studied associates was probed. Analyses of memory recall indicated that both affectively neutral and negative mnemonic representations experienced similar levels of enhancement and impairment following selective retrieval, demonstrating the susceptibility of affectively salient memories to these mnemonic processes. These findings indicate that although affective memories may be more strongly encoded in memory, they remain amenable to inhibition and flexibly adaptable to the evolving needs of the organism.
Introduction
Memory is a highly adaptive and flexible cognitive function that enables an individual to recall information from the past in the service of future goals. Despite the vast amount of information contained in human memory, individuals are often able to recall a specific event, detail, or fact when it would support current behaviour. Searching memory for a particular mnemonic representation mobilises multiple control processes, both to amplify relevant mnemonic representations and to mitigate interference from other information also stored in memory. This second process inhibits irrelevant mnemonic representations, minimising competition from other related traces in the memory cache during selective retrieval and facilitating successful recall. Critically, this inhibition may have lasting consequences, such that items that generate competition during selective retrieval will be forgotten over time. Forgetting, therefore, can be an adaptive consequence of attempts to remember.
It remains unclear, however, what role forgetting plays in governing affective memory. On the one hand, emotionally laden memory traces may have special status in memory, making them less amenable to executive control functions. The affective salience and potential biological relevance of emotional stimuli may buffer them from forgetting. Conversely, the capacity to govern affective representations is critical to guiding cognitive processes along goal-relevant pathways. Successful resolution of mnemonic interference is especially vital when mnemonic competitors are of an affective nature, and the ability to manage the contents of long-term memory in accordance with an organism’s goals may hinge upon successful inhibition of affective mnemonic competitors. The importance of resolving affective mnemonic interference therefore suggests that emotional representations in memory may be amenable to inhibitory processing. The present article addresses this question by adapting a retrieval practice paradigm to examine the influence of mnemonic inhibition on emotional memory traces. We propose that interference generated by affective memory representations is resolved by a similar mechanism to that evoked by affectively neutral memories, resulting in robust forgetting of emotional information following selective retrieval of related information and promoting adaptive cognitive processing in healthy individuals.
Memory, interference, and forgetting
Memory enables individuals to marshal past information in the service of current goals or to shape future responding. However, because of the vast amount of information humans are capable of storing, memories can also interfere with one another, competing for representation during encoding and retrieval. This interference can impede retrieval and degrade memory representations (Anderson, Citation1974; Underwood, Citation1957). Various control mechanisms are often recruited to help compensate for and overcome the effects of interference, quelling inappropriate or maladaptive retrieval (Anderson, Citation2003; Kuhl & Wagner, Citation2009). Guided by these control mechanisms, memory allows the retrieval of specific, relevant information and can prevent irrelevant memories from interfering with ongoing cognitive functions. Thus, the resolution of mnemonic interference provides a mechanism for forgetting and other memory errors.
Forgetting may occur not only due to intentional efforts to purge information stored in memory (Anderson & Green, Citation2001) but also as an unintentional consequence of inhibitory processes mobilised by executive control during retrieval (Anderson et al., Citation1994). This phenomenon of retrieval-induced forgetting has been rigorously examined utilising variants of the retrieval practice paradigm (Anderson et al., Citation1994). In the retrieval practice paradigm, participants study word pairs in which one of the words involved is associated with multiple other items (e.g. FRUIT – banana, FRUIT – orange, TOOL – hammer, TOOL – wrench). After encoding, participants’ memory is tested for a subset of the items, often employing a cue plus letter stem format (e.g. FRUIT – b____). When attempting to retrieve the appropriate target (i.e. banana), other items sharing the same cue (e.g. orange) may compete for awareness and impede retrieval. To facilitate retrieval of the appropriate target, relevant mnemonic representations are amplified. Importantly, to mitigate interference generated by competitors, irrelevant but competing mnemonic representations are inhibited. After selective retrieval, memory for all items is tested to examine the long-term consequences of this mnemonic selection and inhibition. At final test, memory for practiced items (e.g. banana) is often enhanced, reflecting the consequences of selection. Critically, memory for the competing items (e.g. orange) is impaired not only relative to the practiced items but also with respect to unpracticed items that did not share an association with the practiced cues (e.g. TOOL – hammer, TOOL – wrench). This second class of items should not have competed during selective retrieval and thus would not have been the targets of either amplification or inhibition during retrieval; as they are retained over a similar temporal interval, this class of items serves as a useful baseline against which to judge the level of inhibition exerted over the unpracticed, competing items (e.g. orange). The demonstration that recall of these unpracticed, competing items is lower than that for these baseline items therefore demonstrates the long-term consequences of mnemonic control mechanisms invoked during selective retrieval to mitigate interference.
The conditions that promote inhibition in memory and its consequences indicate that mnemonic inhibition is goal-driven and robust. Inhibitory processes are theorised to play a central role in driving forgetting (Anderson, Citation2003; Levy & Anderson, Citation2002; Storm & Levy, Citation2012), although some theories emphasise the contribution of other mnemonic interference processes, such as associative blocking and output interference (Butler et al., Citation2001; Camp et al., Citation2007; Williams & Zacks, Citation2001). Several lines of evidence demonstrate that mnemonic interference processes drive forgetting and that inhibition is mobilised when mnemonic competition disrupts selective retrieval. The inhibition enacted to resolve mnemonic interference during selective retrieval results in lasting degradation of the competing mnemonic representation, as evidenced by the fact that impaired recall of inhibited items extends to tests utilising independent cues (Anderson & Spellman, Citation1995) or recognition formats (Gomez-Ariza et al., Citation2005; Hicks & Starns, Citation2004; Roman et al., Citation2009; Spitzer et al., Citation2009). Inhibition therefore targets not merely the association between the retrieval cue and the competitor but the representation of the competitor itself. Critically, inhibition is contingent of the presence of mnemonic competition; in cases where competition is not evoked, suppression of mnemonic representations will not occur. For instance, weak associates of cues that are low frequency and less likely to be retrieved in response to cues are also less likely to be inhibited (Anderson et al., Citation1994). Moreover, the mere act of retrieval is insufficient to generate inhibition, as retrieving cues and not associates (e.g. F____ – banana) does not lead to suppression of unpresented, related associates (Anderson et al., Citation2000), highlighting the importance of competition during retrieval in prompting suppression. Neural network models have supported the importance of competition in generating inhibition, as only strong competitors are likely to become the targets of inhibitory processes (Norman et al., Citation2007). Mnemonic interference can generate inhibition even in the absence of successful retrieval (Storm et al., Citation2006), demonstrating that the effects of inhibition are dissociable from the consequences of selection. That is to say, retrieval-induced forgetting is not merely a function of greater strengthening of the retrieved targets but is also shaped by inhibitory processes that are mobilised during selective retrieval. Retrieval-induced forgetting is impaired in populations with degraded inhibitory control, such as individuals with attention-deficit/hyperactivity disorder (Storm & White, Citation2010), further implicating control in implementing forgetting under conditions of mnemonic competition. While the present experiments did not specifically test an inhibitory account of retrieval-induced forgetting (Anderson, Citation2003), the preponderance of evidence has extensively demonstrated that inhibition contributes to retrieval-induced forgetting effects (see Storm & Levy, Citation2012).
Mnemonic competition during selective retrieval generates interference, which control processes mobilise to resolve. This mnemonic interference resolution not only amplifies the representations of the targets of mnemonic search but also inhibits the representations of competing memory traces, with both processes exerting lasting mnemonic consequences.
Emotion modulation of memory
Although several studies of mnemonic interference resolution highlight the importance of understanding forgetting of emotional memories (e.g. Anderson, Citation2006), empirical evidence on the topic is scant. Findings from the emotional memory literature indicate that affectively salient memories are more strongly encoded than affectively neutral memories, giving them a mnemonic advantage not only in the short term but also at long retention intervals (Bradley et al., Citation1992; Dolcos et al., Citation2005). Given the mnemonic advantage of emotional memories and their longevity, are they less susceptible to inhibition and forgetting than affectively neutral memories are? Specifically, are emotional memory representations robust to mnemonic interference resolution?
There is some theoretical basis for the hypothesis that emotional memories would be insensitive to mnemonic control processes, particularly inhibition. Emotional memories are strongly encoded, which may help buffer them from the effects of inhibition. Evidence from intentional directed forgetting of negative materials reveals reduced inhibition of negative materials (Minnema & Knowlton, Citation2008; Nowicka et al., Citation2011). A meta-analysis shows diminished inhibition of emotional memories in directed forgetting paradigms (Hall et al., Citation2021), supporting the notion that negative mnemonic representations do not suffer from intentional inhibition. This observation from intentional forgetting may extend to the goal-driven, unintentional forgetting that occurs as a function of the interference resolution processes triggered during selective retrieval. Additionally, negative stimuli are often resistant to attentional control processes (Reeck & Egner, Citation2011, Citation2015), suggesting they might be similarly robust to mnemonic control processes.
Conversely, there is also theoretical basis for the hypothesis that emotional memories may be amenable to inhibitory processes. As interference resolution processes are mobilised by competition, the enhanced memory strength of emotional memories may facilitate their retrieval and cause them to generate competition during selective retrieval. This competition would therefore mobilise interference resolution processes, leading to ample inhibition of affective memories. This position is consistent with computational models of executive control in memory, which emphasise that strong competitors during selective retrieval should experience robust levels of inhibition (Norman et al., Citation2007). Several studies of intentional forgetting indicate that negative memories can be inhibited (Depue et al., Citation2007; Joormann et al., Citation2005), with one investigation finding that emotional memory representations may be more susceptible to mnemonic control processes than neutral memories (Depue et al., Citation2006). However, it is important to note that this previous investigation only compared recall performance between representations that were selected and inhibited, showing that affective representations exhibited larger differences between conditions of selection and inhibition. Recall was not explicitly compared to performance on baseline items or between emotionally negative and neutral inhibition conditions, leaving open the possibility that emotional memory traces may experience similar levels of inhibition when compared to baseline memories as neutral memory traces. Moreover, these previous experiments examined attempts at intentional, instructed forgetting, which may be sensitive to strategic shifts and demand characteristics, and left unexplored the consequences of mnemonic interference resolution processes on affective memories.
The possibility that emotional memories may be amenable to mnemonic control processes engaged in response to interference has been contentious in the literature. One investigation found no overall differences in retrieval-induced forgetting across affective stimulus classes, yet did find that more negatively arousing stimuli were less likely to experience inhibition (Kuhbandner et al., Citation2009). Another experiment observed that retrieval-induced forgetting did not occur for affective mnemonic representations (Dehli & Brennen, Citation2009), but this finding did not replicate in a separate investigation (Blix & Brennen, Citation2012). Some investigations examining autobiographical memories have indicated that mnemonic inhibition occurs for both emotional and non-emotional memories (Barnier et al., Citation2004), whereas others have produced evidence that inhibition does not occur for positive memories (Wessel & Hauer, Citation2006).
These mixed findings and divergent theoretical perspectives may arise from a number of variations in experimental procedure and materials. For instance, adapting the retrieval practice paradigm to accommodate selective retrieval of autobiographical memories necessitated deviations from the general procedure, such as testing details of a central episode (Wessel & Hauer, Citation2006), which may have altered the underlying executive control and mnemonic processes. Investigations using verbal stimuli employed very few emotional stimuli (Kuhbandner et al., Citation2009) or employed emotional stimuli that differed from neutral stimuli not only in their affective qualities but also in their basic semantic features (Dehli & Brennen, Citation2009), which may have impacted results. Semantic features have been shown to alter affective modulation effects in memory processes in general (Dillon et al., Citation2006; Talmi & Moscovitch, Citation2004) and retrieval-induced forgetting effects in particular (Anderson, Citation2003; Goodmon & Anderson, Citation2011), which likely contributed to the mixed findings from previous experiments. A series of empirical investigations employing established procedures and appropriately-controlled stimuli is therefore necessary in order to evaluate the capacity of mnemonic interference resolution to enact long-term consequences on affectively charged mnemonic representations and establish the consistency of these effects.
The present research
The present experiments examine whether the emotional salience of a mnemonic representation modulates its susceptibility to control processes in long-term memory. Specifically, the impact of goal-driven interference resolution processes that guide selective retrieval on affective and neutral mnemonic competitors is examined. In light of the mixed findings from previous investigations employing unique procedures and varied stimuli, the present experiment sought to mirror the traditional retrieval practice paradigm as closely as possible to examine the impact of mnemonic inhibition on affective representations. Additionally, the verbal stimuli employed in the present experiments were carefully selected and controlled to mitigate the contributions of semantic features to the present findings. We sought to investigate whether inhibitory processes mobilised to resolve mnemonic interference during selective retrieval would exert similar control over both affective and neutral mnemonic competitors or would spare affective mnemonic competitors. Elucidating whether or not affective status of mnemonic competitors modulates retrieval-induced forgetting would clarify the capacity of control signals to modulate the emotional contents of long-term memory and potentially provide insight into clinically-relevant phenomena. Given the importance of forgetting negative memories for clinical applications related to mood disorders and trauma, the following experiments focused on characterising mnemonic interference resolution processes with respect to negatively valenced stimuli in healthy participants.
Experiment 1
The present experiment employed an adapted version of a retrieval practice paradigm designed to examine the consequences of control engaged during selective retrieval on affectively salient and neutral memory representations. Participants initially encoded neutral cue words paired with negative and neutral associates. Subsequently, participants’ memory for a subset of the items was tested multiple times. We anticipated that during this selective retrieval phase, control processes would improve the accessibility of tested target items while impairing the accessibility of potential mnemonic competitors. The consequences of these control processes were evaluated on a final memory test, which probed participants’ memory for all initially encoded items. We expected that mnemonic competitors would experience inhibition regardless of their affective salience.
Methods
Participants
Forty-one participants were recruited from the Duke University community to participate in the present experiment for either course credit or payment. Participants were eligible to participate if they were native English speakers or learned English before the age of six and if they were between the ages of 18 and 40. Four participants were excluded due to a technical error that resulted in a failure to record data during the final recall test and two participants were excluded due to a technical error during the retrieval practice phase that terminated the phase prematurely. Data from all six participants were excluded from all analyses, leaving a sample size of thirty-five (19 females) with an average age of 21.2 years old (s.d. = 3.4). 40.9% of participants identified as Caucasian, 15.9% as African or African-American, 15.9% as Asian or Asian-American, 2.3% as Hispanic or Latino, and 4.6% with multiple ethnicities. This sample size was consistent with prior retrieval-induced forgetting studies (e.g. Gomez-Ariza et al., Citation2005; Hicks & Starns, Citation2004; Spitzer et al., Citation2009), including studies that examined emotional modulation of retrieval induced forgetting (e.g. Blix & Brennen, Citation2012; Dehli & Brennen, Citation2009; Wessel & Hauer, Citation2006). Sensitivity analyses indicated it was sufficiently powered to detect an effect of f = .18. All participants provided informed consent and all experimental procedures were approved by the Duke University Medical Center Institutional Review Board.
Materials
Forty nouns served as cue words in this experiment. Each cue had six noun associates, resulting in 240 cue-associate word pairs. Half of the associates for each cue were affectively neutral and the other half were affectively negative, with each set of associates serving as a separate subset for counterbalancing purposes. To verify that the negative and neutral nouns employed as associates in the present experiment differed in their emotional content, a separate sample of participants (N = 46) rated these associates using the Self-assessment Manikin (SAM) scale (Bradley & Lang, Citation1999). Participants rated each associate’s valence (negative to positive) and arousal (calm to excited) on a 5-point scale. Affectively negative words were rated as more negative in valence (M = 1.7) compared to neutral items (M = 3.3), t(238) = 33.63, p < .001, d = 4.43, and more arousing (M = 3.1) than neutral items (M = 2.4), t(238) = 11.94, p < .001, d = 1.54. Thus, the affectively negative verbal stimuli employed in the present study were both more negatively valenced and more arousing than the affectively neutral stimuli employed.
As the present study sought to examine the effects of affective salience on mnemonic executive control, steps were taken to ensure that the affectively neutral and affectively negative associates differed in affective salience but otherwise had similar semantic properties. The means for frequency as indexed by the Corpus of Contemporary American English (Davies, Citation2008) were similar for affectively neutral (M = 14,405.7) and negative associates (M = 11,360.5), t(238) = 1.15, p = .252, and affectively neutral (M = 6.0) and negative (M = 5.9) associates were a similar length, t(238) = 0.19, p = .850. When selecting associates, words with a moderate relationship with the cue were selected, and we sought to avoid selecting words with either dominant or idiosyncratic associations with the cue word. To evaluate if associates across affective categories had a similar level of relatedness to the cue word, latent semantic analysis was used (http://lsa.colorado.edu). Both affectively neutral (M = 0.22) and negative (M = 0.19) associates were similarly related to the cue words, t(238) = 1.41, p = .160. As semantic integration has been proposed to present a potential boundary condition on retrieval-induced forgetting (Goodmon & Anderson, Citation2011), the degree of relatedness among the neutral associates that shared each cue and among the negative associates that shared each cue was also assessed using latent semantic analysis. This analysis indicated that both affectively neutral (M = 0.16) and negative (M = 0.18) associates exhibited a similar level of relatedness with the other associates from the same affective class sharing the same cue, t(238) = 1.25, p = .213. Overall, the affectively neutral and negative associates employed in the present study were similar on the dimensions of frequency, length, relatedness to the cue, and relatedness to other associates sharing the same cue from the same affective category. The full list of stimuli is available in the Supplement.
To facilitate counterbalancing, the forty cues were divided into four separate lists. Follow-up analyses indicated that the associates within these four lists did not differ in frequency, length, relatedness to the cues, relatedness to other associates from the same affective category sharing the cue, rated valence, or rated arousal (p’s > .15). For each list, the affectively negative associates and the affectively neutral associates comprised a sub-list. The mnemonic condition to which each sub-list was assigned was counterbalanced across participants. Each list of cues could be assigned to one of three circumstances: affectively neutral associates serving as targets during the retrieval practice phase (Neutral RP+) and negative associates serving as competitors (Negative RP−), affectively negative associates serving as targets during the retrieval practice phase (Negative RP+) and neutral associates serving as competitors (Neutral RP−), or associates were untested during the retrieval phase (Negative NRP, Neutral NRP).
Procedure
Participants completed an adapted version of the retrieval practice paradigm (Anderson et al., Citation1994) across five phases (). Presentation of experimental stimuli and data collection were controlled using Psychophysics Toolbox Version 3 (Brainard, Citation1997; Pelli, Citation1997). During the initial encoding phase of the experiment, participants studied 240 cue-associate word pairs. Each word pair was presented individually on a trial, with the cue appearing in capital letters and the associate appearing in lowercase letters (e.g. AIR – cloud, AIR – pollution). Each trial began with a one-second fixation, followed by a four-second presentation of the word pair. Participants were instructed to memorise each word pair in preparation for an upcoming memory test. The presentation order in this phase was constrained such that word pairs that shared a cue never appeared consecutively to minimise relational binding across associates.
Figure 1. Retrieval Practice Paradigm. In the initial encoding phase, participants memorise cue – associate word pairs. Each cue has three negative and three neutral associates. During the retrieval practice phase, participants’ memory for a subset of the encoded items is probed. On each trial, participants are provided with the cue word plus a letter stem and have four seconds to retrieve the full associate from memory. In Experiment 1, participants were provided with a 1-letter stem. In Experiment 2 and the supplemental experiment, participants were provided with a 2-letter stem as shown here to facilitate self-monitoring of performance. During the final test, recall for all original items is tested. Not depicted in this figure is the visuospatial distracter task in the interval between retrieval practice and test and the final rating phase.

Following this encoding phase, participants then completed a retrieval practice phase. During this phase, participants were asked to retrieve a subset of the studied items. Each trial began with a one-second fixation, followed by the four-second presentation of a retrieval cue: the capitalised cue word along with the first letter of the associate being tested (e.g. AIR – c___). While the cue was presented on the screen, participants attempted to recall the associate (e.g. cloud). Participants provided a response via keyboard presses indicating their retrieval success, pressing the “1” key if they could successfully recall the associate or the “2” key if they could not successfully recall the associate. All responses were covert, and feedback on retrieval accuracy was not provided to participants. In order to facilitate participant responding on each trial, during the final second of the trial the presentation font colour changed from white to red, serving as a reminder for the participant to register a response. The retrieval phase spanned two blocks, and each cued associate was practiced three times during this phase. The presentation order was controlled so that associates sharing a cue were never probed consecutively. Upon completion of the retrieval practice phase, participants then completed a fifteen-minute visuospatial filler task involving making judgments about abstract images of fractals in order to mitigate the influence of short-term memory effects on recall performance during the final test phase.
The final test had a similar presentation structure to the retrieval practice phase. The trial structure was identical to that employed during the retrieval practice phase, but with all items tested instead of a subset. Participants were instructed to recall the probed associate and vocally respond with the target word if they retrieved it. A microphone recorded verbal responses during this phase, which were coded offline for accuracy. Presentation order of retrieval cues was again constrained such that associates sharing a cue were never tested consecutively.
During the final phase of the experiment, participants provided affective ratings of the associates employed in the experiment. Associates appeared individually (i.e. without their studied cue) and participants provided sequential judgments of the word’s valence (negative to positive) and arousal utilising a 5-point SAMS scale (Bradley & Lang, Citation1999). Ratings were self-paced, and the presentation order was constructed such that an equal number of items from each affective mnemonic condition appeared in each third of the list. Data from this experiment are available at https://osf.io/45uqv/?view_only = 730710271e3a4c7f920230764f26d877.
Results
Affective ratings
During the rating phase, affectively negative associates were rated as more negative (M = 1.55, s.d. = 0.34) than neutral (M = 3.21, s.d. = 0.20) associates, t(34) = 24.48, p < .001, d = 4.14. Affectively negative associates (M = 2.74, s.d. = 0.92) were also rated as more arousing than neutral associates (M = 2.15, s.d. = 0.75), t(34) = 3.95, p < .001, d = 0.67. Therefore, participant ratings confirmed that negative verbal stimuli were more emotionally engaging than neutral verbal stimuli.
Retrieval practice phase performance
Performance during the retrieval practice phase was examined to assess the benefits of executive control during selective retrieval. Initial repeated-measures ANOVAs of retrieval success and response times on successful trials revealed no main effects of emotion or interactions between emotion and retrieval attempt (all p’s > .15), so these variables were collapsed across affective stimulus category (). A repeated-measures ANOVA of retrieval success revealed no effects of retrieval attempt (first, second, or last), F(2, 68) = 0.56, p = .576, indicating that retrieval success did not become more likely over the course of the retrieval practice phase, possibly due to the large size of the mnemonic set. Successful retrieval did become faster, however, over the course of retrieval practice phase as indicated by a repeated-measures ANOVA, F(2, 68) = 87.91, p < .001, η2 = .72. Follow-up t-tests indicated that successful retrieval was slower on the first attempt than either the second, t(34) = 8.36, p < .001, d = 1.41, or the third attempt, t(34) = 12.85, p < .001, d = 2.17. Successful retrieval was also faster on the final attempt than on the second attempt, t(34) = 4.79, p < .001, d = 0.81. Thus, although retrieval success did not become more likely over the course of the retrieval practice phase, it did become faster and more efficient, consistent with the operation of mnemonic control processes mobilised during selective retrieval.
Table 1. Performance from Retrieval Practice Phase of Experiment 1. Means (standard deviations). Accuracy rates and response times (seconds) from the first, second, and third retrieval attempts during the retrieval practice phase.
Participants’ responses were analyzed to assess if recall accuracy on the final test corresponded with self-reported retrieval during the retrieval practive phase. There was a main effect of the number of times an item was endorsed as successfully retrieved during the retrieval practice phase, F(3, 99) = 72.32, p < .001, η2 = .69. Participants were least likely to accurately recall an associate if it was never endorsed as successfully retrieved during the retrieval practice phase (M = 2.91%, s.d. = 4.42%), more likely after one indication of successful retrieval (M = 10.78%, s.d. = 14.75%), more likely after two indications of successful retrieval (M = 41.27%, s.d. = 29.19%), and most likely if the item was always endorsed as successfully retrieved (M = 61.59%, s.d. = 27.44%). All pairwise comparisons were significant, p’s < .01. Thus, responses during the retrieval practice phase provided a valid index of retrieval success.
Final recall performance
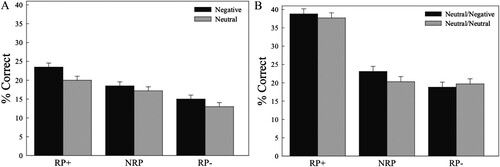
Overall, participants recalled an average of 17.86% of the studied items (s.d. = 9.16%). A repeated-measures ANOVA was conducted to investigate the within-subjects factors of emotion (negative or neutral) and retrieval practice condition (RP+, NRP, or RP−) ((A)). This analysis yielded a main effect of emotion, F(1, 34) = 5.30, p = .028, η2 = .14, as negative associates were more likely to be successfully recalled than neutral associates. Importantly, there was also a main effect of retrieval practice condition, F(2, 68) = 21.07, p < .001, η2 = .38. Follow-up t-tests indicated that RP+ items were more likely to be recalled at final test than either NRP items, t(34) = 3.16, p = .003, d = 0.53, or RP− items, t(34) = 5.98, p < .001, d = 1.01. Moreover, RP− items were less likely to be successfully recalled at final test than NRP items, t(34) = 3.73, p = .001, d = 0.63, indicating a significant retrieval-induced forgetting effect. Critically, the interaction between emotion and retrieval practice condition was not significant, F(2, 68) = 0.95, p = .393, indicating that emotion did not modulate the long-term effects of executive control on memory. Given that establishing whether negative and neutral stimuli experienced different levels of retrieval-induced forgetting was a central aim of the present research, we followed up on this null result by conducting a Bayesian analysis to assess evidence in favour of the null hypothesis. The analysis produced a Bayes Factor of 6.96 in favour of the null hypothesis, which should be interpreted as moderate evidence for the null hypothesis. Thus, although affectively negative associates were more likely to be recalled overall, there was no difference in their susceptibility to either practice enhancement or retrieval-induced forgetting, signalling their amenability to mnemonic selection and inhibition, respectively.
Figure 2. Free recall performance for Experiments 1 (Panel A) and 2 (Panel B). Note that in Experiment 2, for cues in the Neutral/Negative condition, RP+ items were always neutral and RP− items were always negative. Analyses in Experiment 1 revealed a main effect of emotion, with negative items recalled more frequently than neutral items. In both Experiments 1 and 2, recall rates on the final test indicated significant enhancement effects for practiced items (RP+) and retrieval-induced forgetting of unpracticed competitors (RP−) regardless of their affective salience.
Item analysis of final recall performance
Analyses of participant recall performance on the final memory test indicated that neither the practice enhancement nor the retrieval-induced forgetting effects were modulated by affect. Seeking to confirm this interpretation at the level of individual items, an item analysis was conducted. For each item included in the present study, its average valence and arousal rating was computed based on participant ratings provided at the end of the experiment. The probability of successful retrieval was also computed for each item, both overall and separately for each condition it appeared in: RP+, NRP, and RP−. A suppression score was computed for each item by calculating the difference between recall rates in the NRP and RP− condition and dividing that difference by the overall NRP recall rate [(NRP − RP−)/NRP] (Kuhl et al., Citation2007). An enhancement score was also computed by calculating the difference between recall rates in the RP+ and NRP conditions and dividing that difference by the overall NRP recall rate [(RP+ − NRP)/NRP]. Correlation analyses were then conducted to examine whether either valence or arousal correlated with suppression score or enhancement score. Neither valence nor arousal correlated with either observed enhancement or retrieval-induced forgetting for items (all p’s > .5). Thus, the findings from the item analysis also reveal no evidence for affective modulation of mnemonic control processes evoked during selective retrieval. We followed up on these findings by conducting the corresponding Bayesian analyses to assess the extent to which there was evidence in favour of a null hypothesis of no affective modulation of memory. The Bayes Factors of these analyses were all greater than 16.62 in favour of the null hypothesis, indicating strong evidence for the null hypothesis of no affective modulation. Thus, the affective nature of the stimuli did not alter their susceptibility to either practice enhancement or retrieval-induced forgetting.
Discussion
The findings from Experiment 1 indicate that mnemonic interference resolution processes enact similar consequences on both affectively negative and neutral mnemonic representations. Following selective retrieval, the targets of retrieval were better recollected compared to items from unpracticed categories retained over the same temporal interval. Items that would have generated interference during selective retrieval were less likely to be recalled on the final memory test, relative not only to practiced targets but also to items from unpracticed categories. Thus, mnemonic competitors exhibited enhanced forgetting relative to items retained over the time interval. Importantly, while affectively negative associates were more likely to be recalled than neutral associates, neither the practice enhancement effect nor the retrieval-induced forgetting effects were modulated by the affective salience of the targets of control. Thus, executive processes mobilised in response to mnemonic interference exert similar consequences on both negative and neutral mnemonic representations.
Experiment 2
The previous experiment provided evidence that negative mnemonic representations are amenable to control processes in long-term memory. This pattern was replicated in an additional supplemental experiment (see Supplement). Memory for affectively negative verbal stimuli was enhanced by practice and impaired by retrieval-induced forgetting at similar levels as memory for affectively neutral verbal stimuli. In Experiment 1, participants practiced both negative and neutral associates during the retrieval practice phase, albeit the negative and neutral associates were derived from different cues. This design feature could have impacted the findings in two ways. First, the recall of negative associates during the retrieval practice phase could alter participant mood, which may alter mnemonic control processes. Specifically, negative mood states have been associated with diminished retrieval-induced forgetting (Baeuml & Kuhbandner, Citation2007; Koessler et al., Citation2009) and could modulate the retrieval process. Second, the counterbalancing structure of Experiment 1 resulted in negative items serving as competitors for neutral items and neutral items serving as competitors for negative items. The finding that both negative and neutral competitors experienced similar levels of forgetting might actually mask differences in the dynamics of interference between neutral and negative items. For example, when negative items are practiced, they might simply be easier to retrieve, leading to less interference from competitors overall (in this case, neutral items) and less subsequent forgetting of those competitors (reduced forgetting). Meanwhile, when neutral items are practiced, negative items may generate substantial interference, but be less amenable to forgetting (reduced forgetting). In this example, therefore, both negative and neutral associates would exhibit similar levels of forgetting, but for different reasons. To more clearly examine the impact of mnemonic interference on negative competitors, in Experiment 2 participants only practiced retrieval of neutral associates during the retrieval practice phase.
Methods
Participants
Forty-six participants were recruited to participate in the experiment. One participant did not complete the task as instructed, and was thus excluded from all analyses. Participant recruitment criteria excluded individuals who were not native English speakers or had a history of substance abuse, major psychological illness, or neurological illness. Participants (27 females) were 18–30 years old (M = 21.8, s.d. = 3.2) and provided informed consent. The sample was ethnically diverse, with 43.2% self-identifying as Caucasian, 29.6% as Asian or Asian-American, 15.9% as African or African-American, 2.3% as American Indian or Native Alaskan, 2.3% as Hispanic or Latino, and 6.8% as mixed ethnicity. This sample size was consistent with prior retrieval-induced forgetting studies, including studies that examined emotional modulation of retrieval induced forgetting. Sensitivity analyses indicated it was sufficiently powered to detect an effect of f = .16. All procedures were approved by the Duke University Medical Center Institutional Review Board.
Materials
Twenty-eight nouns served as cue words, each with six noun associates. To allow associate-specific cueing, all of the associates linked with a given cue began with a unique first letter. In the present experiment, negative associates were not retrieved during the retrieval practice phase and therefore never appeared in the RP+ condition. To accommodate this design change, half of the cues had six neutral associates while the other half of the cues had three neutral and three negative associates. For the cue words with only neutral associates, separate sub-lists of three items each were constructed for counterbalancing purposes. When the cue word was practiced during the retrieval practice phase, either sub-list could serve as the targets for retrieval. For the other half of the cues, when the cue word was practiced during the retrieval practice phase, only the neutral associates served as targets and the negative associates were always competitors. Overall, 126 of the associates in the stimulus set were neutral and 42 were negative. Analyses indicated that neutral and negative associates did not differ on semantic features. Both negative (M = 15,552.6) and neutral (M = 17,048.2) associates had similar word frequencies, t(166) = 0.35, p = .726, and negative (M = 5.9) and neutral (M = 6.0) associates were a similar length, t(166) = 0.30, p = .766. Negative (M = .22) and neutral (M = .22) associates were also similarly related to the cue words as indexed by latent semantic analysis, t(166) = 0.08, p = .939, and exhibited a similar degree of relatedness to other associates that shared the same cue within a given sub-list, t(166) = 0.61, p = .541 (Negative M = .17, Neutral M = .15). Ratings provided by an independent group of participants (N = 15) indicated that negative and neutral words were also judged as similarly imageable, t(166) = 1.198, p = .234 (Negative M = 4.04, Neutral M = 4.30), and concrete, t(166) = 1.182, p = .239 (Negative M = 4.68, Neutral M = 4.99). Overall, the negative and the neutral associates were highly similar on relevant semantic dimensions. Four separate counterbalancing lists were constructed, and analyses indicated that these lists did not differ from one another on word frequency, length, relatedness between associates and cues, or relatedness among the associates within a sub-list for each cue (p’s > .3). The full list of stimuli is available in the Supplement.
Procedure
The experimental protocol was nearly identical to that utilised in the previous experiment with a few minor differences. First, the presentation order was controlled in each experimental phase so that an equal number of items in each condition appeared in each third of the list to minimise any potential order effects. Second, to facilitate participant monitoring of accuracy during the retrieval practice phase, the retrieval cues consisted of the cue word in capital letters followed by the first two letters of the probed associate in lowercase letters (e.g. AIR – cl____). Third, the retrieval practice and final test phases spanned three blocks instead of two. Finally, at the end of the rating phase, participants also rated the cue words in addition to the associates employed in the experiment individually. Except for these minor differences, the procedure was otherwise identical to that employed in Experiment 1. Data from this experiment are available at https://osf.io/45uqv/?view_only = 730710271e3a4c7f920230764f26d877.
Results
Affective ratings
Participants’ ratings of the valence and arousal of the associates indicated that they found the negative associates more affectively charged than the neutral associates. Negative associates (M = 1.6, s.d. = 0.2) were rated as more negatively valenced than neutral associates (M = 3.4, s.d. = 0.2), t(44) = 33.48, p < .001, d = 4.99, and participants also indicated they found the negative items (M = 3.2, s.d. = 0.8) more arousing than the neutral items (M = 2.0, s.d. = 0.6), t(44) = 9.12, p < .001, d = 1.36.
Accounts of retrieval-induced forgetting have suggested that selective retrieval may generate associative interference by altering the meaning of the cue words such that unpracticed associates share a weaker association with the cue (Anderson, Citation2003). In the present experiment, this account could be particularly important, as practicing neutral associates could make the cue words seem more neutral in nature and weakening the association between the cue word and unpracticed negative associates. To assess this possible interference mechanism, the ratings of the cue words were examined. Cues with both neutral and negative associates were rated as similarly valenced, t(44) = 0.51, p = .616, and arousing, t(44) = 1.33, p = .189, regardless of whether the cue was unpracticed during the retrieval practice phase or the neutral associates were practiced. Thus, the cue’s affective qualities do not seem to be modified by practicing the neutral associates, indicating that alterations in cue affect do not likely contribute to retrieval-induced forgetting in the present experiment.
Retrieval practice phase performance
Performance during the retrieval practice phase was examined to assess if the affective nature of mnemonic competitors influenced selective retrieval (). Recall success rates from the retrieval practice phase were examined using a two-way repeated-measured ANOVA with the within-subjects factors of affective status of mnemonic competitors (negative or neutral) and retrieval attempt (first, second, or third). As this analysis yielded no main effects or interactions with affective status (p’s > .5), this analysis was re-run collapsing across the factor of affective status of mnemonic competitors. This analysis revealed a main effect of retrieval attempt, F(2, 88) = 9.63, p = .002, ηp2 = .18. Follow-up t-tests indicated that retrieval success was less likely on the first retrieval attempt compared to either the second, t(44) = 4.41, p < .001, d = 0.66, or the third, t(44) = 3.10, p = .003, d = 0.46, retrieval attempt. However, retrieval success was similarly likely for the second and third retrieval attempts, t(44) = 1.06, p = .294. Retrieval success therefore became more likely following the first attempt but stabilised after the second retrieval attempt.
Table 2. Performance from Retrieval Practice Phase of Experiment 2. Means (standard deviations). Accuracy rates and response times (seconds) from the first, second, and third retrieval attempts during the retrieval practice phase.
Response latencies indicating successful retrieval were analyzed using a two-way repeated-measures ANOVA, with the within-subjects factors of affective status of mnemonic competition (negative or neutral) and retrieval attempt (first, second, or third). This analysis yielded a main effect of competitor affective status, F(1, 44) = 12.21, p = .001, ηp2 = .22, and a main effect of retrieval attempt, F(2, 88) = 62.87, p < .001, η2 = .59. These main effects were qualified, however, by a significant interaction, F(2, 88) = 4.91, p = .009, ηp2 = .10. Separate repeated-measures ANOVAs for items with neutral competitors, F(2, 88) = 60.86, p < .001, ηp2 = .58, and negative competitors, F(2, 88) = 34.02, p < .001, ηp2 = .44, revealed significant effects of retrieval attempt for both types of items. Follow-up t-tests to explore the interaction between competitor affective status and retrieval attempt indicated that items with negative mnemonic competitors were recalled faster on the first retrieval attempt compared to items with neutral mnemonic competitors, t(44) 4.19, p < .001, d = 0.63. However, this difference in response latency was not present at either the second, t(44) = 0.94, p = .352, or the third, t(44) = 1.40, p = .170, retrieval attempt, indicating that this difference in retrieval success latency dissipated after the first retrieval attempt. After the initial retrieval attempt, therefore, successful retrieval exhibits similar response latency regardless of whether mnemonic competitors are negative or neutral.
Reponses during the retrieval practice phase were assessed to determine if they tracked recall accuracy on the final test. Consistent with all the previous experiment, there was a main effect of the number of times an item was endorsed as successfully retrieved during the retrieval practice phase, F(3, 126) = 99.18, p < .001, ηp2 = .71. Participants were least likely to accurately recall an associate if it was never endorsed as successfully retrieved during the retrieval practice phase (M = 4.2%, s.d. = 6.9%), more likely after one indication of successful retrieval (M = 24.3%, s.d. = 23.8%), more likely after two indications of successful retrieval (M = 47.4%, s.d. = 28.4%), and most likely if the item was always endorsed as successfully retrieved (M = 70.7%, s.d. = 21.0%). All pairwise comparisons were significant, p’s < .001. Thus, participants’ responses provided a valid measure of retrieval success and provided a reliable index of mnemonic selection processes.
Final recall performance
Final recall performance was analyzed to assess the impact of affect and retrieval practice status on memory ((B)). Each cue word was associated either exclusively with neutral items or with both neutral and negative items. In the latter circumstance, the negative associates were never practiced during the retrieval practice phase and could appear only in the NRP or RP− conditions. A repeated-measures ANOVA with the within-subjects factors of cue-type (neutral only or mixed-valence associates) and mnemonic condition (RP+, NRP, RP−). Note that for cues with both negative and neutral associates, only neutral items appeared in the RP+ condition and only negative items appeared in the RP− condition. This analysis yielded a main effect of mnemonic condition, F(2, 88) = 77.04, p < .001, ηp2 = .64. Follow-up t-tests indicated that recall for RP+ items was significantly higher than for either NRP items, t(44) = 9.48, p < .001, d = 1.41, or RP− items, t(44) = 9.61, p < .001, d = 1.43, reflecting practice enhancement effects. Moreover, recall rates were higher for NRP items than for RP− items, t(44) = 2.09, p = .043, d = 0.31, indicating significant retrieval-induced forgetting. Neither the main effect of cue type, F(1, 44) = 0.90, p = .347, nor the interaction, F(2, 88) = 1.51, p = .228 was significant. Thus, cue type and affective status did not significantly modulate the effects of selection or inhibition on memory. To assess the evidence for the null hypothesis of no affective modulation, we conducted the corresponding Bayesian analyses. This analysis produced a Bayes Factor of 1.35 in favour of the null hypothesis, providing weak evidence in favour of the null. As a confirmatory follow-up, recall rates specifically to the negative associates were separately probed to confirm that retrieval-induced forgetting indeed occurred for negative items. The analysis comparing recall rates for negative associates in the NRP condition and the RP− condition indicated that significant retrieval-induced forgetting did indeed occur, t(44) = 3.88, p < .001, d = 0.58, consistent with the findings from the previous experiments presented here.
Item analysis of final recall performance
Analyses of participants’ memory performance on the final recall test did not yield evidence of significant modulation of mnemonic control effects by affect. To complement these analyses, an item analysis was also conducted to evaluate whether modulation might occur at the level of individual items. For each item, the average valence and arousal ratings were calculated from the ratings provided by participants at the end of the experiment. The overall recall rate for each item as well as its enhancement and retrieval-induced forgetting score were also computed. Correlation analyses revealed no correlations between either valence or arousal and any of the memory-related indices (p’s > .18), indicating that affect did not modulate either selection or inhibition effects in the present experiment. The corresponding Bayesian analysis was again conducted. These analyses all produced Bayes Factors greater than 5.71 in favour of the null hypothesis, which should be interpreted as moderate evidence for the null hypothesis of no affective modulation of memory.
Discussion
The findings from Experiment 2 were consistent with the conclusions from the previous experiments. Both practice enhancement effects and retrieval-induced forgetting effects were not modulated by the affective nature of mnemonic competitors. Control processes mobilised as a result of interference generated by competitors during selective retrieval acted upon both affective classes of mnemonic representations. In this experiment, negative items were not cued during the retrieval practice phase, yet the findings were concordant with those from the previous experiment that cued both negative and neutral items. Therefore, there does not appear to be support for the notion that mood effects generated during the retrieval practice phase influenced mnemonic executive control processes. Mnemonic control processes therefore appear to exert similar levels of influence across both negative and neutral mnemonic representations.
General results
Across the experiments reported here and in the supplemental information, mnemonic selection and inhibition evoked during selective retrieval exerted similar effects on negative and neutral mnemonic representations. Although none of our experiments produced a significant interaction between mnemonic status and affective salience, it is possible that each experiment was underpowered to detect such an interaction. To address this concern, we combined data across all three experiments and conducted a repeated-measures ANOVA to assess the within-subjects effects of affect and mnemonic condition (RP+, NRP, RP−). This approach would enable us to detect subtle effects across the three experiments. Please note that this approach gives us a larger sample size than prior studies examining affective modulation of retrieval-induced forgetting. A sensitivity analysis indicates that this approach allows for the detection of effect sizes of f = .09, which corresponds to a small effect size. Consistent with the retrieval-induced forgetting literature, there was a significant main effect of mnemonic condition, F(2, 246) = 173.19, p < .001, η2 = .59. Subsequent t-tests indicated that recall for RP+ items was significantly higher than recall for either NRP, t(123) = 12.73, p < .001, d = 1.14, or RP− items, t(123) = 14.96, p < .001, d = 1.34, demonstrating an enhancement effect. Importantly, recall of RP− items was significantly lower than recall for NRP items, t(123) = 5.80, p < .001, d = 0.52, providing evidence of retrieval-induced forgetting. The main effect of emotion was not significant, F(1, 123) = 2.18, p = .143, η2 = .02, and the interaction between emotion and memory was not significant, F(2, 246) = 0.40, p = .669, η2 < .01. Thus, the affective salience of the mnemonic representations did not appear to modulate either the mnemonic effects of selection or inhibition. As examining whether negative and neutral stimuli experienced similar levels of retrieval-induced forgetting was a main aim of the present paper, we also conducted the corresponding Bayesian analyses. These analyses revealed a Bayes Factor of 11.37 in favour of the null hypothesis, which should be interpreted as strong evidence in favour of the null hypothesis. Thus, we find no evidence of affective modulation of retrieval-induced forgetting and our analyses support a conclusion of no differences in memory based on affect.
General discussion
Emotional events and experiences can have lasting effects on an organism. The question arises whether the mnemonic consequences of these emotional experiences are permanent or whether executive control processes may modify these mnemonic traces. The present article examined whether the mnemonic interference resolution processes mobilised by selective retrieval enact similar consequences on both affectively negative and neutral mnemonic representations. Three empirical investigations supported the hypothesis that both selection and inhibition act similarly on negative and neutral mnemonic representations, demonstrating that affective memories are amenable to executive control.
The present experiments employed an adapted retrieval practice paradigm, employing both affectively negative and neutral verbal mnemonic associates in order to assess the effects of mnemonic control processes across different affective classes of memories. All three experiments provided strong evidence that the effects of control processes mobilised in response to mnemonic competition are consistent for both negative and neutral memories. Following repeated retrieval attempts, the targets of retrieval were more accurately recalled on a final memory test compared to other items. The increased accessibility of these items reflects selection processes engaged during mnemonic interference resolution that amplify representations of the intended targets of retrieval in order to enable successful mnemonic recovery. Crucially, recall of items that would have generated competition during selective retrieval was impaired on the final memory test compared to other items. Memory performance was diminished relative to items maintained over a similar delay that were unlikely to generate competition, indicating that inhibitory processes played a role in mnemonic interference resolution. Importantly, neither the observed enhancement nor suppression effects was modulated by the affective status of the mnemonic representations being altered. These conclusions were further supported by an item analysis, that demonstrated that neither an item’s valence nor its arousal altered its susceptibility to mnemonic control. Interestingly, Experiment 1 demonstrated that these findings emerged even when an overall memory benefit for negative memories was observed and when the demands on selective retrieval were high. These findings were not the result of negative moods generated by the recall of negative items during selective retrieval, as demonstrated by the fact that an identical pattern of results was observed when only neutral items were tested during the retrieval practice phase in Experiment 2. Experiment 2 also addressed the concern that observed retrieval-induced forgetting might be the result of changes in retrieval cue representation. The process of retrieving either negative or neutral associates of a cue may cause the subjective affective quality of the cue itself to alter to take on the characteristics of its retrieved associates, thus contributing to forgetting effects (Anderson, Citation2003). However, rating data from Experiment 2 demonstrated practicing a subset of its mnemonic associates did not alter the affective quality of the cues for participants, indicating that this route likely did not play a major role in contributing to the present findings. The finding that negative mnemonic representations are amenable to control in general and inhibition in particular was robust and consistent across all three experiments.
The present findings provide important insight into the relationship between emotional memory and the processes that govern long-term memory. The demonstration that retrieval-induced forgetting occurs not only with affectively neutral but also with negatively valenced stimuli is particularly interesting given the importance of understanding the forgetting of negative material to theories of repression and responses to trauma (Anderson, Citation2006). However, it is important to note that the present investigations utilised verbal stimuli, which are more readily controlled but less affectively evocative than life stressors associated with clinical affective disorders. Nonetheless, the finding that mnemonic interference resolution processes act upon negative memories is consistent with evidence indicating that memories of emotional life events are susceptible to decay and forgetting over time (Hirst et al., Citation2009; Talarico & Rubin, Citation2003). The present demonstration extends these findings by demonstrating that forgetting of emotional events is not merely due to passive decay over time but rather can be enacted in an active fashion in response to mnemonic interference generated during selective retrieval.
The present experiments provide clear evidence that retrieval-induced forgetting extends to negative mnemonic representations and also resolve conflicting conclusions arising from discrepant reports from previous experiments, some of which observed retrieval-induced forgetting with negative materials (Barnier et al., Citation2004; Kuhbandner et al., Citation2009), and others which did not (Dehli & Brennen, Citation2009; Moulds & Kandris, Citation2006; Wessel & Hauer, Citation2006). Findings of reduced or eliminated retrieval-induced forgetting may have arisen from differences in semantic features between emotional and non-emotional materials employed. For instance, relatedness between associates has been shown to have profound impacts on retrieval-induced forgetting (Goodmon & Anderson, Citation2011) and semantic features have previously been demonstrated to modulate emotional effects in long-term memory (Dillon et al., Citation2006; Talmi & Moscovitch, Citation2004). The present experiment controlled for the influence of these features and found no evidence of an affective modulation of retrieval-induced forgetting, suggesting that earlier observed effects may have been influenced by semantic factors. Moreover, the present observed effects were preserved across three separate experiments and a larger combined sample size than other work in this area, establishing the consistency of the finding. However, the present experiments all employed only affectively negative and neutral stimuli, limiting conclusions to negatively valenced mnemonic representations. Future experiments should examine whether mnemonic interference resolution processes also extend to positive memories.
Affective states have previously been shown to modulate mnemonic inhibitory processes. Negative moods (Baeuml & Kuhbandner, Citation2007) and stress inductions (Koessler et al., Citation2009) eliminate retrieval-induced forgetting, and positive moods minimise directed forgetting (Baeuml & Kuhbandner, Citation2009). The present demonstration that negative items are amenable to retrieval-induced forgetting raises interesting questions regarding potential interactions between affective states and affective stimuli. Demonstrations of mood congruent memory suggest that affective representations relevant to the current mood state are more accessible (Bower, Citation1981), and may be both more easily selected when cued and more likely to compete for awareness during selective retrieval. Mood-congruent affective representations may therefore be more likely to be inhibited, although this effect could be modulated by the modes of cognitive processing promoted by the present affective state. This interaction between emotional materials and affective states could therefore modify the nature of mnemonic competition during retrieval and alter the processes engaged in resolving mnemonic interference.
The present findings may also be relevant to understanding factors contributing to clinical disorders. Researchers have long appreciated that cognitive biases play a role in anxiety and depression, promoting more negative interpretations of events (Mathews & MacLeod, Citation2005). Anxious individuals are not only more likely to interpret an ambiguous event as negative in the moment, but these distorted interpretations also bias memory of the event and lead individuals to later falsely recall more negative details associated with the original experience (Hertel et al., Citation2008). The present experiments explored the effects of mnemonic interference resolution processes on emotional memory representations in individuals without a current or previous psychological disorder. Future research should therefore examine whether similar effects are found in clinical populations or whether deficits in mnemonic inhibition of emotional materials are observed. The disruption of cognitive inhibition of emotional mental representations may play a key role in the etiology and maintenance of clinical disorders (Joormann, Citation2010), and characterising the efficacy of mnemonic interference resolution processes would provide insight into the cognitive mechanisms contributing to psychopathology.
The present experiments demonstrated that retrieval-induced forgetting extends to both neutral and emotional memories. Mnemonic interference resolution processes mobilised by competition generated during selective retrieval govern mnemonic representations regardless of affective status, managing the contents of long-term memory in concert with an individual’s goals. Affective memories are therefore not immutable representations, but rather amenable to adjustment and capable of flexibly updating in an ongoing fashion to meet an organism’s goals.
PCEM_A_Supplemental Information
Download MS Word (62.9 KB)Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Anderson, J. R. (1974). Retrieval of propositional information from long-term memory. Cognitive Psychology, 6(4), 451–474. https://doi.org/10.1016/0010-0285(74)90021-8
- Anderson, M. C. (2003). Rethinking interference theory: Executive control and the mechanisms of forgetting. Journal of Memory and Language, 49(4), 415–445. https://doi.org/10.1016/j.jml.2003.08.006
- Anderson, M. C. (2006). Repression: A cognitive neuroscience approach. Psychoanalysis and Neuroscience, 12, 327–349. https://doi.org/10.1007/88-470-0550-7_14
- Anderson, M. C., Bjork, E. L., & Bjork, R. A. (2000). Retrieval-induced forgetting: Evidence for a recall-specific mechanism. Psychonomic Bulletin & Review, 7(3), 522–530. https://doi.org/10.3758/BF03214366
- Anderson, M. C., Bjork, R. A., & Bjork, E. L. (1994). Remembering can cause forgetting: Retrieval dynamics in long-term memory. Journal of Experimental Psychology: Learning, Memory, and Cognition, 20(5), 1063–1087. https://doi.org/10.1037/0278-7393.20.5.1063
- Anderson, M. C., & Green, C. (2001). Suppressing unwanted memories by executive control. Nature, 410(6826), 366–369. https://doi.org/10.1038/35066572
- Anderson, M. C., & Spellman, B. A. (1995). On the status of inhibitory mechanisms in cognition: Memory retrieval as a model case. Psychological Review, 102(1), 68–100. https://doi.org/10.1037/0033-295X.102.1.68
- Baeuml, K.-H., & Kuhbandner, C. (2007). Remembering can cause forgetting—but Not in negative moods. Psychological Science, 18(2), 111–115. https://doi.org/10.1111/j.1467-9280.2007.01857.x
- Baeuml, K.-H., & Kuhbandner, C. (2009). Positive moods can eliminate intentional forgetting. Psychonomic Bulletin & Review, 16(1), 93–98. https://doi.org/10.3758/PBR.16.1.93
- Barnier, A., Hung, L., & Conway, M. (2004). Retrieval-induced forgetting of emotional and unemotional autobiographical memories.. Cognition & Emotion, 18(4), 457–477. https://doi.org/10.1080/0269993034000392
- Blix, I., & Brennen, T. (2012). Retrieval-induced forgetting after trauma: A study with victims of sexual assault. Cognition & Emotion, 26(2), 321–331. https://doi.org/10.1080/02699931.2011.570312
- Bower, G. H. (1981). Mood and memory. American Psychologist, 36(2), 129–148. https://doi.org/10.1037/0003-066X.36.2.129
- Bradley, M. M., Greenwald, M. K., Petry, M. C., & Lang, P. J. (1992). Remembering pictures: Pleasure and arousal in memory. Journal of Experimental Psychology: Learning, Memory, and Cognition, 18(2), 379–390. https://doi.org/10.1037/0278-7393.18.2.379
- Bradley, M. M., & Lang, P. J. (1999). Affective norms for English words (ANEW): Stimuli, instruction manual and affective ratings. Technical report C-1. The Center for Research in Psychophysiology, University of Florida.
- Brainard, D. H. (1997). The psychophysics toolbox. Spatial Vision, 10(4), 433–436. https://doi.org/10.1163/156856897X00357
- Butler, K. M., Williams, C. C., Zacks, R. T., & Maki, R. H. (2001). A limit on retrieval-induced forgetting. Journal of Experimental Psychology: Learning, Memory, and Cognition, 27(5), 1314–1319. https://doi.org/10.1037/0278-7393.27.5.1314
- Camp, G., Pecher, D., & Schmidt, H. G. (2007). No retrieval-induced forgetting using item-specific independent cues: Evidence against a general inhibitory account. Journal of Experimental Psychology: Learning, Memory, and Cognition, 33(5), 950–958. https://doi.org/10.1037/0278-7393.33.5.950
- Davies, M. (2008). The Corpus of Contemporary American English: 425 million words, 1990-present. http://corpus.byu.edu/coca/
- Dehli, L., & Brennen, T. (2009). Does retrieval-induced forgetting occur for emotional stimuli? Cognition and Emotion, 23(6), 1056–1068. https://doi.org/10.1080/02699930802285221
- Depue, B. E., Banich, M. T., & Curran, T. (2006). Suppression of emotional and nonemotional content in memory. Psychological Science, 17(5), 441–447. https://doi.org/10.1111/j.1467-9280.2006.01725.x
- Depue, B. E., Curran, T., & Banich, M. T. (2007). Prefrontal regions orchestrate suppression of emotional memories via a two-phase process. Science, 317(5835), 215–219. https://doi.org/10.1126/science.1139560
- Dillon, D. G., Cooper, J. J., Grent-'t-Jong, T., Woldorff, M. G., & LaBar, K. S. (2006). Dissociation of event-related potentials indexing arousal and semantic cohesion during emotional word encoding. Brain and Cognition, 62(1), 43–57. https://doi.org/10.1016/j.bandc.2006.03.008
- Dolcos, F., LaBar, K. S., & Cabeza, R. (2005). Remembering one year later: Role of the amygdala and the medial temporal lobe memory system in retrieving emotional memories. Proceedings of the National Academy of Sciences, 102(7), 2626–2631. https://doi.org/10.1073/pnas.0409848102
- Gomez-Ariza, C. J., Lechuga, M. T., Pelegrina, S., & Bajo, M. T. (2005). Retrieval-induced forgetting in recall and recognition of thematically related and unrelated sentences. Memory & Cognition, 33(8), 1431–1441. https://doi.org/10.3758/BF03193376
- Goodmon, L. B., & Anderson, M. C. (2011). Semantic integration as a boundary condition on inhibitory processes in episodic retrieval. Journal of Experimental Psychology: Learning, Memory, and Cognition, 37(2), 416–436. https://doi.org/10.1037/a0021963
- Hall, K. J., Fawcett, E. J., Hourihan, K. L., & Fawcett, J. M. (2021). Emotional memories are (usually) harder to forget: A meta-analysis of the item-method directed forgetting literature. Psychonomic Bulletin & Review, 28(4), 1313–1326. https://doi.org/10.3758/s13423-021-01914-z
- Hertel, P. T., Brozovich, F., Joormann, J., & Gotlib, I. H. (2008). Biases in interpretation and memory in generalized social phobia. Journal of Abnormal Psychology, 117(2), 278-288. https://doi.org/10.1037/0021-843X.117.2.278
- Hicks, J. L., & Starns, J. J. (2004). Retrieval-induced forgetting occurs in tests of item recognition. Psychonomic Bulletin & Review, 11(1), 125–130. https://doi.org/10.3758/BF03206471
- Hirst, W., Phelps, E. A., Buckner, R. L., Budson, A. E., Cuc, A., Gabrieli, J. D. E., & Vaidya, C. J. (2009). Long-term memory for the terrorist attack of September 11: Flashbulb memories, event memories, and the factors that influence their retention. Journal of Experimental Psychology: General, 138(2), 161–176. https://doi.org/10.1037/a0015527
- Joormann, J. (2010). Cognitive inhibition and emotion regulation in depression. Current Directions in Psychological Science, 19(3), 161–166. https://doi.org/10.1177/0963721410370293
- Joormann, J., Hertel, P. T., Brozovich, F., & Gotlib, I. H. (2005). Remembering the good, forgetting the bad: Intentional forgetting of emotional material in depression. Journal of Abnormal Psychology, 114(4), 640–648. https://doi.org/10.1037/0021-843X.114.4.640
- Koessler, S., Engler, H., Riether, C., & Kissler, J. (2009). No retrieval-induced forgetting under stress. Psychological Science, 20(11), 1356–1363. https://doi.org/10.1111/j.1467-9280.2009.02450.x
- Kuhbandner, C., Baeuml, K.-H., & Stiedl, F. (2009). Retrieval-induced forgetting of negative stimuli: The role of emotional intensity. Cognition & Emotion, 23(4), 817–830. https://doi.org/10.1080/02699930802204768
- Kuhl, B., Dudukovic, N. M., Kahn, I., & Wagner, A. D. (2007). Decreased demands on cognitive control reveal the neural processing benefits of forgetting. Nature Neuroscience, 10(7), 908–914. https://doi.org/10.1038/nn1918
- Kuhl, B., & Wagner, A. D. (2009). Forgetting and retrieval. In G. G. Berntson & J. T. Cacioppo (Eds.), Handbook of neurosciences for the behavioral sciences (Vol. 1, pp. 586–605). Wiley.
- Levy, B. J., & Anderson, M. C. (2002). Inhibitory processes and the control of memory retrieval. Trends in Cognitive Sciences, 6(7), 299–305. https://doi.org/10.1016/S1364-6613(02)01923-X
- Mathews, A., & MacLeod, C. (2005). Cognitive vulnerability to emotional disorders. Annual Review of Clinical Psychology, 167–195. https://doi.org/10.1146/annurev.clinpsy.1.102803.143916
- Minnema, M. T., & Knowlton, B. J. (2008). Directed forgetting of emotional words. Emotion, 8(5), 643–652. https://doi.org/10.1037/a0013441
- Moulds, M., & Kandris, E. (2006). The effect of practice on recall of negative material in dysphoria. Journal of Affective Disorders, 91(2–3), 269–272. https://doi.org/10.1016/j.jad.2006.01.003
- Norman, K. A., Newman, E. L., & Detre, G. (2007). A neural network model of retrieval-induced forgetting. Psychological Review, 114(4), 887–953. https://doi.org/10.1037/0033-295X.114.4.887
- Nowicka, A., Marchewka, A., Jednorog, K., Tacikowski, P., & Brechmann, A. (2011). Forgetting of emotional information Is hard: An fMRI study of directed forgetting. Cerebral Cortex, 21(3), 539–549. https://doi.org/10.1093/cercor/bhq117
- Pelli, D. G. (1997). The VideoToolbox software for visual psychophysics: Transforming numbers into movies. Spatial Vision, 10(4), 437–442. https://doi.org/10.1163/156856897X00366
- Reeck, C., & Egner, T. (2011). Affective privilege: Asymmetric interference by emotional distracters. Frontiers in Psychology, 2. https://doi.org/10.3389/fpsyg.2011.00232
- Reeck, C., & Egner, T. (2015). Interactions between attention and emotion. In A. W. Toga (Ed.), Brain mapping: An encyclopedic reference (pp. 269–274). Elsevier.
- Roman, P., Soriano, M. F., Gomez-Ariza, C. J., & Bajo, M. T. (2009). Retrieval-Induced forgetting and executive control. Psychological Science, 20(9), 1053–1058. https://doi.org/10.1111/j.1467-9280.2009.02415.x
- Spitzer, B., Hanslmayr, S., Opitz, B., Mecklinger, A., & Baeuml, K.-H. (2009). Oscillatory correlates of retrieval-induced forgetting in recognition memory. Journal of Cognitive Neuroscience, 21(5), 976–990. https://doi.org/10.1162/jocn.2009.21072
- Storm, B. C., Bjork, E. L., Bjork, R. A., & Nestojko, J. F. (2006). Is retrieval success a necessary condition for retrieval-induced forgetting? Psychonomic Bulletin & Review, 13(6), 1023–1027. https://doi.org/10.3758/BF03213919
- Storm, B. C., & Levy, B. J. (2012). A progress report on the inhibitory account of retrieval-induced forgetting. Memory & Cognition, 40(6), 827–843. https://doi.org/10.3758/s13421-012-0211-7
- Storm, B. C., & White, H. A. (2010). ADHD and retrieval-induced forgetting: Evidence for a deficit in the inhibitory control of memory. Memory (Hove, England), 18(3), 265–271. https://doi.org/10.1080/09658210903547884
- Talarico, J. M., & Rubin, D. C. (2003). Confidence, not consistency, characterizes flashbulb memories. Psychological Science, 14(5), 455–461. https://doi.org/10.1111/1467-9280.02453
- Talmi, D., & Moscovitch, M. (2004). Can semantic relatedness explain the enhancement of memory for emotional words? Memory & Cognition, 32(5), 742–751. https://doi.org/10.3758/BF03195864
- Underwood, B. J. (1957). Interference and forgetting. Psychological Review, 64(1), 49–60. https://doi.org/10.1037/h0044616
- Wessel, I., & Hauer, B. J. A. (2006). Retrieval-induced forgetting of autobiographical memory details. Cognition and Emotion, 20(3–4), 430–447. https://doi.org/10.1080/02699930500342464
- Williams, C. C., & Zacks, R. T. (2001). Is retrieval-induced forgetting an inhibitory process? The American Journal of Psychology, 114, 329–354. https://doi.org/10.2307/1423685