
ABSTRACT
Purpose: The aim of this study was fourfold: (1) to quantify acceleration, velocity, and phase overlap for each phase of the stroke cycle (SC) during 200 m front crawl; (2) for each variable, to identify any differences between the four SC phases; (3) to investigate changes in variables during the 200 m; (4) to explore any association between performance and each variable. Methods: Ten swimmers performed a 200 m maximum swim. Four SCs were analyzed, one for each 50 m, using three-dimensional methods. Each SC was split into four phases: entry, pull, push, and recovery. Center of mass (CM) acceleration; maximum, minimum, and average CM velocity; phase duration, and, overlap of a phase of one arm with each phase of the opposite arm were calculated. Results and Conclusion: Phase velocities were positively correlated with performance and decreased during the 200 m. The acceleration data showed high within and between-swimmer variability. When the entry of one arm overlapped with the pull, and sometimes push, phase of the opposite arm, it was propulsive for the whole body. The pull was the slowest phase and overlapped predominantly with the opposite arm’s recovery. The push phase was often propulsive for the whole body, regardless of the overlaps with the other arm, and together with the entry were the fastest phases. The recovery of each arm was mostly resistive for the whole body, except the short period of overlap with the opposite arm’s push phase.
Swimming performance is affected by propulsive and resistive forces, which are often interlinked. For example, an increase in propulsion could cause an increase in the body’s velocity, but the higher velocity will subsequently also increase the resistance experienced. Examination of propulsive and resistive forces can therefore advance our knowledge of swimming technique and facilitate performance enhancement. There is, however, no direct method of measuring propulsive and resistive forces during free swimming. Although these forces can be measured directly during assisted and resisted swimming, such conditions compromise ecological validity, and therefore further examination of the overall interplay between propulsive and resistive forces for the whole body is warranted. An indirect method of exploring this interplay between propulsion and resistance during free swimming is to calculate the center of mass (CM) acceleration in different phases of the stroke cycle (SC) (Takagi et al., Citation2021). If acceleration is positive, then propulsive forces are higher than resistive forces and the phase is “propulsive,” while the reverse is true when acceleration is negative (Gourgoulis et al., Citation2018). Therefore, some authors have discussed the usefulness of whole body acceleration data in identifying the propulsive or resistive nature of different phases of the SC (Figueiredo, Kjendlie, et al., Citation2012; Gourgoulis et al., Citation2018). Despite its usefulness, the acceleration of the CM in different phases of the SC, has not been calculated with three-dimensional methods during free swimming in any studies. This is perhaps partly due to the complexity of methods and the sophisticated equipment required for its calculation (for example, above and below water three-dimensional analysis and accurate anthropometric data for estimation of the body’s CM).
Researchers typically split the front crawl SC into four different phases, defined by the arms’ movements: entry, pull, push and recovery (McCabe et al., Citation2011). Because the arm is not applying propulsive forces during the entry and recovery, researchers often refer to these phases as nonpropulsive, with pull and push described as propulsive. Although this terminology is often used primarily for the arm actions, it may sometimes be implied or assumed that the pull and push phases are propulsive for the whole body, with the entry and recovery assumed to be nonpropulsive. This assumption, however, overlooks important factors affecting propulsion and resistance, and is therefore prone to errors that could lead to misconceptions. For example, propulsive forces are generated also by the kicking actions during the SC (Bartolomeu et al., Citation2018). Moreover, resistive forces depend on body position, which changes throughout the SC, but also on factors such as the magnitude of swimming velocity. It is therefore unknown if the propulsive forces of the arms during the pull and push phases are larger than the resistance experienced by the body, to allow characterization of these phases as propulsive for the whole body. Likewise, it is not known if the resistive forces are higher than the propulsive forces during entry and recovery and, thus, one could not simply assume that these phases are nonpropulsive for the whole body. Analysis of whole-body acceleration for all phases would therefore be very useful in providing information on the balance between propulsion and resistance. Consideration of velocity data during these phases could assist in explaining the observed acceleration patterns and any between-phase differences in acceleration.
It is also important to note that with the alternate arm motions in front crawl and backstroke, when one arm is applying propulsive forces, the other arm may not be applying any forces. For example when one arm is pulling, the opposite arm may be recovering. As the actions of the opposite arm are affecting body position and acceleration, it would be very useful to quantify the duration of overlaps between different phases of the opposite arms, i.e. how much each phase of one arm overlaps with each of the phases of the opposite arm. Although some studies have explored overlaps between pull of one arm and push of the opposite arm (Seifert et al., Citation2007), the overlaps between all four phases of the SC have yet to be investigated. Such phase calculations should be conducted for both the dominant and nondominant arms, as hand dominance has sometimes been shown to be linked to bilateral asymmetries in technique (Psycharakis & Sanders, Citation2008). Finally, it is known that performance and technique often deteriorate with the fatigue caused during a race (Figueiredo, Kjendlie, et al., Citation2012). Thus, it would be interesting to identify if acceleration, velocity and phase overlap data would change during the course of an event.
The aim of the present study was fourfold: (1) quantify acceleration, velocity and phase overlaps for each of the four SC phases during a 200 m maximum front crawl swim; (2) for each variable, identify any differences between the four SC phases; (3) identify any changes in variables during the course of the 200 m; (4) explore any association between performance (as indicated by average SC velocity) and each of the variables measured. Based on previous evidence, it was hypothesized that the phase velocities would decrease during the 200 m and that they would be associated with performance.
Materials and methods
Participants
The participants were 10 male swimmers of national or international level (defined as having qualified and competed at national or international level competitions within the preceding year), specialized in 200 m front crawl (17.1 ± 0.9 years, 181.0 ± 5.4 cm, 72.4 ± 5.7 kg, 200 m personal best: 121.5 ± 4.9 s). All swimmers were training competitively and undertaking a minimum of five training sessions per week. They were free from injury and illness and avoided stressful training in the days prior to testing. The institutional ethics committee approved the study and all participants signed informed consent forms.
Experimental protocol and data analysis
All testing took place in a 25 m indoors pool. The swimmers performed their own personalized warm up, followed by a 20-min passive rest, to simulate a race scenario. Each swimmer then performed a 200 m maximal front crawl swim alone, with no other swimmers in the pool at the same time. All swimmers started with a push start, in order to prevent the dive influencing the kinematics of the first SC that was used for the analysis. Swimmers were instructed to use the same pacing and strategy that they would use in competition. To ensure that test performance would be at a similar level to competition performance, the 200 m time had to be within 105% of a swimmer’s personal best performance of the season (also considering the effect of the push start on the final time). All swimmers satisfied this criterion when first tested, with test time being on average 3% slower than their personal best time.
Before testing commenced, handedness was determined with a laterality questionnaire (Annett, Citation1970; Oldfield, Citation1971) and anthropometric data with the elliptical zone method (Deffeyes & Sanders, Citation2005; Jensen, Citation1978; Psycharakis et al., Citation2010). The latter allowed the calculation of the position of the CM and, hence, all acceleration and velocity variables in the present study represent CM movement. Although all methods that estimate the segment volume and CM rely on some literature-informed assumptions for these variables that may affect accuracy of CM calculations, the elliptical zone method has been shown to produce very small errors and to have very good accuracy and reliability (Psycharakis et al., Citation2010; Sanders et al., Citation2015).
For the 200 m swim, performance was recorded with six stationary, synchronized and genlocked cameras (JVC-KY32CCD, 50 Hz): two mounted above water level and four underwater (more details and illustration of this set-up have been described elsewhere (Psycharakis et al., Citation2010). The length of the space recorded by the six cameras was 6.5 m, allowing for data collection of a full SC for each 50 m length of the 200 m swim (SC1 to SC4; approximately at 20 m, 70 m, 120 m and 170 m). To eliminate effects of breathing on variables (Psycharakis & McCabe, Citation2011), swimmers avoided breathing while swimming through the 6.5 m calibrated space. The camera and calibration set-up have been shown to have negligible image distortion and refraction, high accuracy and reliability (RMS errors in all directions for 20 control points above and 20 below water ranged from 3.3–5.2 mm, representing 0.1–0.5% of calibrated space), and to produce small and acceptable calculation errors, which are similar to or better than other studies that used comparable calibration volumes (de Jesus et al., Citation2015; Psycharakis et al., Citation2005; Psycharakis et al., Citation2010).
The 19 body landmarks that were digitized for each frame of the SC were as follows: vertex; shoulders, elbows, wrists, hips, knees, ankles, metatarsophalangeal joints; end of middle fingers and big toes. The calculation of 3D coordinates relied on the Direct Linear Transformation method (Abdel-Aziz & Karara, Citation2015) incorporated into APAS. The above- and below-water sequences were digitized and transformed separately, and then combined into a single file because they used the same global coordinate system of a single calibration frame. A Fourier transform and inverse transform were used to filter and smooth the raw displacement data by retaining harmonics up to 6 Hz in the inverse transform. The use of the Fourier series transform is regarded as highly appropriate when analyzing periodic data, such as in swimming (Bartlett, Citation1997), and avoids the problem of distortion at the ends of the data set encountered when using other types of filters.
A SC was defined as two consecutive entries of the same hand, and was split into four phases: entry, pull, push, and recovery. In line with other studies in the literature (McCabe et al., Citation2011), the four phases were defined with the use of five distinct events of horizontal displacement of finger and shoulder: X1: finger entry, X2: beginning of finger moving horizontally backwards, X3: finger vertically aligned with the shoulder, X4: end of backward movement, X5: finger reentry. Entry was then defined as the period between X1 and X2, pull as the period between X2 and X3, push as the period between X3 and X4, and recovery as the period between X4 and X5.
For each SC phase, CM velocities (m/s) were calculated by dividing CM displacement by phase duration, and then average, maximum and minimum values were computed. Average CM acceleration (m/s (Gourgoulis et al., Citation2018) was calculated by subtracting the final minus the initial phase velocity and dividing by phase duration. Durations of each phase were reported as a percentage of the whole SC duration. Phase “overlaps” were calculated, to provide an indication of how much overlap exists between different phases of the opposite arms. The phase overlaps were calculated by first isolating a phase of one arm, for example, the entry phase of the dominant arm, and calculating how much time the nondominant arm spent in different phases during that period. These times were then expressed as a percentage of the duration of the “reference” phase of the first; to provide an example, during the entry phase of the dominant arm in the present study, the nondominant arm spent 8% in the pull, 64% in the push and 28% in the recovery phase.
To assess digitizing reliability, one complete SC of one swimmer was digitized 10 times for all cameras and the standard deviation (SD) and coefficient of variation (CV) were calculated for the acceleration and velocity variables during the entry phase. The calculations indicated good reliability: acceleration: SD = 0.08 m/s2, CV = 6.4%; average velocity: SD < 0.01 m/s, CV = 0.4%; maximum velocity: SD = 0.03 m/s, CV = 2.1%; minimum velocity: SD = 0.03 m/s, CV = 1.3%;
Statistical analysis
Statistical analysis was performed with the Statistical Package for Social Sciences (SPSS, version 24). Descriptive statistics are reported as mean group values ± standard deviation (SD). Data normality was checked with the Shapiro–Wilk test. For each variable, significance of bilateral differences between dominant and nondominant sides was checked through paired samples t-tests or Wilcoxon matched pairs signed rank tests. Because of the large number of tests performed (N = 160; four phases × two sides × four pool position stroke cycles for five dependent variables), the α-level was set at 0.01 to mitigate the effect of experimental error-rate. A full Bonferroni correction would require an α-level of 0.0003, but, also considering the sample size, a value this low could lead to a large number of Type 2 errors (false negatives). The α-level of 0.01 may have still “allowed” a small number of Type 1 errors, but this was deemed an acceptable compromise. As nearly all (157 of 160) comparisons indicated no significant bilateral differences, subsequent analyses were performed using the average value of the dominant and nondominant sides.
To identify changes in each variable during the course of the 200 m, a one-way repeated measures ANOVA was performed for normally distributed data to compare the four SCs. There were no violations of the sphericity assumption, so no adjustments to the p values were needed. Post hoc t-tests with Bonferroni adjustments for the alpha level (p < .05, automatically adjusted by SPSS) were carried out between all pairs of SCs. Friedman nonparametric tests were used for non-normally distributed data, with post hoc Wilcoxon matched pairs signed rank tests. Significance for the latter was accepted for p < .01, again to allow for possible Type 1 error-rate inflation.
For each variable, in order to compare all pairs of phases for every SC, paired samples t-tests or Wilcoxon matched pairs signed rank tests were used. Effect sizes (ES) were calculated as an indication of magnitude of differences. The significant differences that led to subsequent calculation of ESs all came from parametric tests, and ES of 0.2, 0.5 and 0.8 were considered small, moderate and large respectively (Cohen, Citation1988; Field, Citation2009). Finally, to assess correlations between performance (indicated by average SC velocity) and each variable, the Pearson’s correlation coefficient (r) was calculated. Because of the large number of tests performed for phase comparisons and correlations, the α-level was also set at 0.01. For the correlations, the ES of the r value was interpreted in line with Hopkins’ suggestions (Hopkins, Citation2006), as follows: < 0.1, very small; 0.1 < 0.3, small; 0.3 < 0.5, moderate; 0.5 < 0.7, large; 0.7 < 0.9 very large; 0.9 < 1.0, nearly perfect.
Results
Variable patterns and phase comparisons
shows the data for acceleration, velocity and phase durations. The mean group data shows that the push phase had positive acceleration and the recovery negative acceleration throughout the 200 m. Positive mean group values for acceleration were also found for the entry and pull phases (except entry in SC1 and pull in SC4). Relatively high SD was observed in all phases, indicating noteworthy intersubject variation. There were no significant between-phase differences in acceleration. Nevertheless, there was a tendency for acceleration to be lower during recovery.
Table 1. Acceleration, velocity, and phase duration data for the 200 m maximum front crawl swim (mean ± standard deviation).
The average velocity data indicated that swimmers were always significantly slower during the pull than all other phases (range of p and d for pairs with significant differences: p ≤ .008, 0.40 ≤ d ≤ 1.62), and that they were sometimes slower in the recovery phase than in the entry (SC1, SC2, mean 200 m values; 0.004 < p ≤ .008, 0.32 ≤ d ≤ 0.68) and in the push phase (SC1; p = .007, d ≤ 0.77). Maximum velocity was significantly lower in the pull phase than in other phases, in one third of all pairwise comparisons (p ≤ .008, 0.72 ≤ d ≤ 1.33). For minimum velocity, significantly higher values were sometimes found in the push than in the pull and recovery phases (SC1, mean 200 m values; p ≤ .002, 0.69 ≤ d ≤ 1.50), and in the entry compared to the pull phase (SC1, p < .001, d = 1.07).
With respect to phase durations, entry was significantly longer (p ≤ .008, 1.57 ≤ d ≤ 4.64) and pull was in most cases significantly shorter (p ≤ .009, 1.06 ≤ d ≤ 4.64) than all other phases. The push and recovery phases had similar durations, except SC1 when recovery was longer (p = .008, d = 1.39).
displays the data for phase duration overlaps. During the entry phase of one arm, the opposite arm spent significantly more time pushing (64% of the reference arm’s entry phase duration, on average) than pulling (11% duration) or recovering (25% duration) (p ≤ .007, 1.01≤ d ≤ 5.07), and significantly more time recovering than pulling (except SC3; p ≤ .009, 2.11≤ d ≤ 3.67). During the pull phase of one arm, the opposite arm was mostly recovering [76% of the pull phase duration; 0.002 ≤ p ≤ .008 and 1.62 ≤ d ≤ 10.27, when compared to “overlap” time the opposite arm spent in entry (23% duration) and push (1% duration)], and there was almost no overlap with the push phase of the opposite arm. Nearly all of the push phase of one arm overlapped with the entry of the opposite arm [94% of push phase duration; 0.002 ≤ p ≤ .004 and 8.20 ≤ d ≤ 21.58, when compared to “overlap” time the opposite arm spent in pull (1% duration) and recovery (5% duration)]. Finally, during the recovery phase of one arm, the opposite arm was mostly in the entry and pull phases (37% and 59% of recovery phase duration, respectively; p ≤ .008 and 1.80 ≤ d ≤ 4.71, when compared to 4% “overlap” time the opposite arm spent in the push phase). provides an example of the SC phases of both arms, and their overlaps, for the velocity of the CM of a single swimmer.
Figure 1. Velocity of the CM of a single swimmer throughout a stroke cycle. The start points of each of the four phases of the stroke cycle are indicated in the graph for both the right arm (RA, continuous vertical lines) and the left arm (LA, dashed vertical lines). The periods of overlaps between the phases of the two arms are indicated at the bottom of the graph.
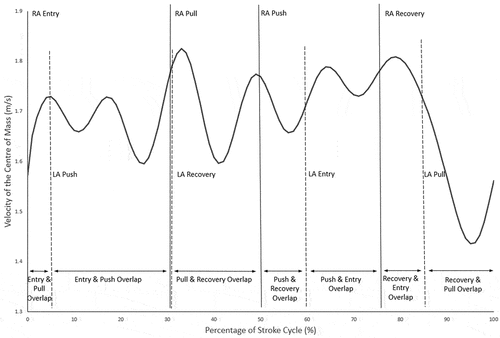
Table 2. Phase overlaps (mean ± standard deviation).
Changes during the 200 m
The values for all variables and all SCs are shown in . There were no significant changes in acceleration during the 200 m (0.2 ≤ F ≤ 1.4, 0.266 ≤ p ≤ .905). Average, maximum and minimum velocity decreased during the 200 m (17.3 ≤ F ≤ 159.9, p ≤ .001). They were all significantly higher in SC1 than in the other SCs (p ≤ .043, 1.71 ≤ d ≤ 4.70), for all four phases. All velocity variables for the entry and push phases were also significantly higher in SC2 than in SC3 and SC4 (p ≤ .049, 0.57 ≤ d ≤ 1.32).
Push duration increased (F = 3.6, p = .029) and recovery duration decreased during the 200 m (F = 14.1, p < .001). The only significant pairwise comparisons were for SC4, which had a longer push than SC3 (p = .005, d = 0.37) and a shorter recovery than SC1 (p = .004, d = 0.89). Phase overlaps were consistent throughout the 200 m, except a decrease in the overlap between one arm’s entry and the opposite arm’s recovery (F = 4.2, p = .015; no significant pairwise comparisons).
Correlations with performance
There were no significant correlations between performance and: acceleration (−0.61 ≤ r ≤ 0.51, 0.060 ≤ p ≤ .989), phase duration (−0.41 ≤ r ≤ 0.63, 0.049 ≤ p ≤ .902; except pull phase duration in SC3: r = −0.78 and p = .007) and phase duration overlaps (−0.60 ≤ r ≤ 0.74, 0.020 ≤ p ≤ .961).
Performance had a positive significant correlation with average (0.765 ≤ r ≤ 0.989, p ≤ .010) and maximum velocity (0.779 ≤ r ≤ 0.983, p ≤ .008) for all phases throughout the 200 m and for the mean 200 m values (except for maximum velocity in SC1 pull, SC1 push and SC4 push phases), with the ESs ranging from very large to nearly perfect. Performance was also positively and significantly associated with minimum velocity for all phases in SC2, SC3 and SC4, as well as the push in SC1 and the mean 200 m values of the pull (0.791 ≤ r ≤ 0.916, p ≤ .006), with very large to nearly perfect ESs.
Discussion
Whole body acceleration data can reveal which phases of the SC are propulsive for the whole body. Exploration of phase overlaps between the opposite arms could assist in explaining the differences in the balance between propulsive and resistive forces in these phases. To our knowledge, this is the first study to report whole body acceleration and phase overlaps for the four phases of the front crawl SC. The results, which are based on rigorous data collection and analysis methods, provide a new insight on technique, and could affect performance and practice.
In summary, the push phase was often propulsive and, together with the entry, were the fastest SC phases. There were large overlaps between the push and entry phases of opposite arms. Contrary to previous assumptions, the entry phase of each arm was not resistive for the whole body, which may be partially explained by its substantial overlap with the opposite arm’s push phase. The pull was the slowest phase and overlapped predominantly with the opposite arm’s recovery. Recovery was resistive for the whole body throughout the 200 m. High SD of the group values for mean acceleration of all phases suggested a degree of interswimmer variance. Velocity decreased during the 200 m and was positively correlated with performance.
Variable patterns and phase comparisons
When discussing the acceleration data, two findings should be taken into consideration. First, that the digitizing reliability for mean acceleration was 0.08 m/s2. Thus, group values lower than 0.08 m/s2 need to be interpreted with caution. Second, that high SD was observed in mean acceleration of all phases, indicating a degree of variability between swimmers, not only in the magnitude of acceleration, but also on characterizing a phase propulsive or resistive for the whole body. Based on these two findings, any group acceleration values between −0.08 and 0.08 m/s2 in the present study were interpreted conservatively and considered “neutral” (i.e. not clearly propulsive or resistive for the swimmer). Ten of the 20 mean group accelerations calculated for all SC phases fell into this category, including four out of five values for both the entry and pull phases.
The acceleration data revealed that the push phase of each arm was propulsive for the whole body in SC1, SC4 and for the average 200 m values (acceleration was positive but within the reliability limits of 0.08 m/s2 in SC2 and SC3). For nearly all of the push phase of one arm (94%), the opposite arm was in the entry phase. The entry of each arm was not a resistive phase for the whole body, contrary to what has been often assumed, and was in fact propulsive in SC3. This may be partly explained by the fact that for about two thirds of the entry duration of one arm, the opposite arm was in the push phase. Moreover, the entry had the longest duration among all four phases and, thus, it may benefit more from propulsive kicking forces. Figueiredo, Sanders, et al. (Citation2012). also found the entry to be substantially longer than all other phases during 200 m front crawl.
In the present study, the entry and push were the phases with the highest velocities in the great majority of cases. Isometric strength measures on land (Mayer et al., Citation1994) have shown that the arm is normally able to apply a larger torque around the shoulder during 0°–90° degrees of shoulder extension (the range that normally corresponds to the push phase), compared to 90°–180° of shoulder extension (range normally corresponding to pull phase). Even though the underwater arm movements in swimming are not isometric, if the arms were able to apply larger torques during the dynamic movements of the push phase, this could be one reason that swimmers were reaching higher velocities in the push than the pull phase. It has also been reported that hand speed is generally faster in the push than in the pull phase (Koga et al., Citation2020; McCabe et al., Citation2015). Although hand propulsive forces are also affected by factors such as the orientation and shape of the hand, higher hand speeds could theoretically increase the propulsive forces of the hand, and partially contribute to the higher velocities in the push than in the pull phase. Further research on other aspects affecting velocity, such as the position, orientation and speed of individual segments and the whole body, could shed further light into the reasons for between-phase differences in velocity and acceleration.
Recovery of each arm was resistive throughout for the whole body, as expected. The pull phase of the arms was propulsive for the whole body only in SC2 and was consistently the slowest of the four phases. A number of factors may have contributed to this, such as that the pull duration was shorter, and thus likely to benefit less from propulsive kicking forces, and that it overlapped primarily with the opposite arm’s recovery. It would also be interesting to explore if factors contributing to increased resistance were present during the pull phase. For example, a difference between the roll of the shoulders and hips would create a “trunk twist,” which could increase frontal surface area and, thus, potentially increase resistance. To check this possibility, the trunk twist for each phase was calculated, as suggested by Psycharakis & Sanders, (Citation2008). The trunk twist of the pull (18.0°) was not however found to be significantly different from that of the push (19.3°) and entry (17.7°) phases. Nevertheless, the frontal surface area may also be affected by the body segments that are above the water. For example, the maximum trunk twist during the pull phase would likely occur when one arm is recovering, and, therefore, the effects of frontal surface area on resistance are more complex to estimate. Thus, consideration of the position of other body segments above and below water, and research on other factors that may affect forces, could help explain further the between- and within-phase differences.
As mentioned above, the high SD in the acceleration values, together with the mean group data being often close to zero, showed that the propulsive or resistive patterns were not always consistent between swimmers. For example, two swimmers had negative accelerations for the push phase throughout the 200 m, and positive accelerations for the recovery for two SCs, i.e. the opposite of what the group values showed for these phases. This considerable individual variation suggests that the consideration of a full SC phase may still be too broad to reveal strong group patterns, and even more in-depth breakdown of acceleration data would be necessary. Given that there are phase overlaps between the two arms, separate calculation of acceleration for each of those overlaps may provide more consistent and informative data. We therefore decided to calculate the acceleration also for all phase overlaps, with the data presented in . The SD remained high, indicating that inter and intraswimmer variability still exists, but some stronger group patterns with much higher mean acceleration values emerged. First, the periods of the SC when the entry of one arm overlaps with the pull of the other arm were propulsive for the whole body (mean acceleration between 0.47 and 1.02 m/s2). These overlaps take up about 8% of the total SC duration. The overlap between push and pull phases of opposite arms also seemed propulsive, but this overlap was present for one to two swimmers and for 0.4% of total SC duration only, so no substantial conclusions can be drawn. There was some evidence of overall propulsion in the overlaps between entry and push phases of the arms, but the values exceeded 0.08 m/s2 for SC3 only. The overlaps between recovery and one other phase showed different patterns. The periods of the SC when recovery of one arm overlapped with entry or pull of the other arm, were generally resistive for the whole body. However, for the short period of the SC when recovery overlapped with the push phase (c. 2.3% of whole SC duration) it was only resistive for the whole body in SC3, but propulsive in the other SCs, showing both within and between swimmer variability.
Table 3. Acceleration data (m/s2) for overlapping phases of the opposite arms during the 200 m maximum front crawl swim (mean ± standard deviation).
In the future, research design could perhaps expand to different ways of breaking down the underwater part of the SC, other than the traditional distinction of entry, pull and push phases. Considering that these phases are defined by arm movements, some patterns in variables may be masked by the use of this phase distinction and may be revealed by exploratory analysis, or by approaches that include consideration of phase overlaps, leg movements and body position. Even with a different breakdown of the SC though, it is not certain that any additional or stronger group patterns would be evident in the acceleration data. Visual observation of our own data suggested several periods of acceleration and deceleration within the SC (e.g., see ), which were highly variable between and within swimmers, with no particular standout sections of the SC that would be different to those analyzed and worth investigating further. Although the individual velocity data were much more consistent and in line with the group data, additional testing of more swimmers and SCs (considering that the 3D methods used in the present study could allow the analysis of just one SC per 50 m), including breathing SCs, would be needed to confirm the acceleration and velocity patterns shown in the present study.
It has been previously suggested that the catch (i.e. when the finger starts moving horizontally backwards, indicating start of pull phase) should be performed early, to minimize entry duration and initiate the pull sooner (Sanders, Citation2000). However, in the present study, the entry was in fact faster than the pull phase. “Catching” sooner would not necessarily mean that pull phase duration (with the arm applying propulsive forces) would be extended or that the early catch would affect performance positively, as there was evidence in SC3 that faster swimmers sometimes spend less time in the pull phase. Given that previous suggestions for an early catch did not specify how early the catch should be performed (Sanders, Citation2000), it is worth taking into consideration the acceleration data for the phase overlaps, shown in . When entry overlapped with the pull and push phases, acceleration was positive (although often within the reliability limits for the overlaps with the pull phase). However, the overlap between entry and recovery showed mostly negative acceleration. The entry phase duration was about a third of the whole SC duration, and the overlap with the recovery occurred in the final quarter of the duration of the entry phase. Thus, shortening the duration of the entry phase by a quarter (i.e. performing the catch at 25% of the whole SC), may in theory avoid the overlap between entry and recovery and potentially improve performance. Nevertheless, it is not known how this reduction may affect body position and performance of subsequent underwater phases, for example an earlier catch may disrupt hydrodynamic position, increasing resistance. Thus, the effect of such an earlier catch on performance should be experimentally explored before any recommendations for technique changes are made.
Changes during the 200 m
Acceleration and phase overlap data were consistent and were not affected by fatigue developed during the race. Velocity variables generally decreased in all phases during the course of the 200 m. This was expected and in line with data for full SCs in other studies (Figueiredo, Kjendlie, et al., Citation2012). Swimmers extended the underwater part of the SC during the course of the race, by spending slightly more time pushing and slightly less time recovering. Figueiredo, Sanders, et al., (Citation2012). also reported that recovery duration decreased in the later stages of a 200 m swim. The corresponding increase in the underwater part of the stroke in that study, however, was observed primarily in the entry and pull phases. The longer duration of the underwater phase may be related to the sub-maximal arm fatigue that has been reported in 200 m events (Figueiredo et al., Citation2013), which S. G. Psycharakis & Yanai, (Citation2018) suggested that may cause the slower underwater movement of the arms during the later stages of this event.
Correlations with performance
There was no evidence in the present study to suggest that acceleration or phase overlap patterns are linearly linked to performance. There were several positive correlations between performance and velocity variables, for all SC phases and throughout the 200 m, suggesting that phase velocities are good performance predictors. This was expected and in agreement with Figueiredo, Kjendlie, et al., (Citation2012), who reported that maximum and minimum velocity data for the full SCs were positively associated to performance. The only other variable associated with performance was the pull phase duration, with faster swimmers spending less time in the pull phase, albeit this correlation was only observed in SC3. Considering that the pull phase ends when the finger is at the same vertical position as the shoulder, a “dropped elbow,” which is an indication of poorer technique (Arellano et al., Citation2003), may lengthen the duration of the pull phase. The dropped elbow has been reported to be a common error for slower swimmers in qualitative studies (Arellano et al., Citation2003), and may be associated with the longer duration of their pull phases.
Conclusion
Contrary to previous assumptions, the entry phase of each arm is not an overall resistive phase for the whole body. It is mostly propulsive for the whole body when overlapping with pull/push phases of the opposite arm, and only resistive toward its end, when it overlaps with the opposite arm’s recovery. The pull phase of each arm was overall not clearly propulsive or resistive for the whole body, perhaps because it overlapped predominantly with the opposite arm’s recovery, and it was the slowest phase of the SC. The push phase of each arm was often propulsive for the whole body, and together with the entry are the fastest phases of the SC. The recovery phase was mostly resistive for the whole body, except for a short period when overlapping with opposite arm’s push phase. Velocity was faster in SC1, and there were positive correlations between the velocities reached at each of the four phases and swimming performance. There were no other changes in variables during the 200 m, or significant correlations with performance. The marked deceleration during recovery suggests that swimmers should minimize the time spent recovering the arms. It may be possible that performing the catch slightly earlier in the entry phase (reducing entry phase duration by about a quarter) would avoid the overlap with the recovery of the opposite arm and could improve performance. However, the effects of such a change on technique and performance have to be tested experimentally before any recommendations for changes are made to swimmers. Overall, testing more swimmers, comparing groups of swimmers with distinctly different performance levels, and including on shorter and longer races than 200 m front crawl, would be helpful in confirming and generalizing the findings of the present study.
Ethics approval
The present study was approved by the ethics committee of the Institute of Sport, Physical Education & Health Sciences (University of Edinburgh).
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Abdel-Aziz, Y. I., & Karara, H. M. (2015). Direct linear transformation from comparator coordinates into object space coordinates in close-range photogrammetry. Photogrammetric Engineering & Remote Sensing, 81(2), 103–107. https://doi.org/10.14358/PERS.81.2.103
- Annett, M. (1970). A classification of hand preference by association analysis. British Journal of Psychology, 61(3), 303–321. https://doi.org/10.1111/j.2044-8295.1970.tb01248.x
- Arellano, R., Lopez-Contreras, G., & Sanchez-Molina, J. (2003). Qualitative evaluation of technique in international Spanish junior and pre junior swimmers: An analysis of error frequencies. In J. Chatard (Ed.), Biomechanics and medicine in swimming IX (pp. 87–92). University of St Etienne Publications.
- Bartlett, R. (1997). Introduction to sports Biomechanics; (1st ed.). E & FN Spon.
- Bartolomeu, R. F., Costa, M. J., & Barbosa, T. M. (2018). Contribution of limbs’ actions to the four competitive swimming strokes: A nonlinear approach. Journal of Sports Sciences, 36(16), 1836–1845. https://doi.org/10.1080/02640414.2018.1423608
- Cohen, J. (1988). Statistical power analysis for the behavioral sciences (2nd ed.). L. Erlbaum Associates.
- Deffeyes, J., Sanders, R. (2005). Elliptical zone body segment modeling software: Digitising, modeling, and body segment parameter calculation. In Q. Wang, (Ed.), Proceedings of the XVII International Symposium on Biomechanics in Sports (pp. 749–752). The China Institute of Sport Science.
- de Jesus, K., de Jesus, K., Figueiredo, P., Vilas-Boas, J. P., Fernandes, R. J., & Machado, L. J. (2015). Reconstruction accuracy assessment of surface and underwater 3D motion analysis: A new approach. Computational and Mathematical Methods in Medicine, 2015, 1–8. https://doi.org/10.1155/2015/269264
- Field, A. P. (2009). Discovering statistics using SPSS: And sex, drugs and rock “n” roll (3rd ed.). SAGE Publications.
- Figueiredo, P., Kjendlie, P. L., Vilas-Boas, J. P., & Fernandes, R. J. (2012). Intracycle velocity variation of the body centre of mass in front crawl. International Journal of Sports Medicine, 33(4), 285–290. https://doi.org/10.1055/s-0031-1301323
- Figueiredo, P., Rouard, A., Vilas-Boas, J. P., & Fernandes, R. J. (2013). Upper- and lower-limb muscular fatigue during the 200-m front crawl. Applied Physiology, Nutrition, and Metabolism, 38(7), 716–724. https://doi.org/10.1139/apnm-2012-0263
- Figueiredo, P., Sanders, R., Gorski, T., Vilas-Boas, J., & Fernandes, R. (2012). Kinematic and electromyographic changes during 200 m front Crawl at race pace. International Journal of Sports Medicine, 34(01), 49–55. https://doi.org/10.1055/s-0032-1321889
- Gourgoulis, V., Koulexidis, S., Gketzenis, P., & Tzouras, G. (2018). Intracyclic velocity variation of the center of mass and hip in breaststroke swimming with maximal intensity. Journal of Strength and Conditioning Research / National Strength & Conditioning Association, 32(3), 830–840. https://doi.org/10.1519/JSC.0000000000002333
- Hopkins, W. G. (2006, August 7). A scale of magnitude for effect statistics. A new view of statistics. Retrieved March 12, 2021, from https://sportsci.org/resource/stats/effectmag.html
- Jensen, R. K. (1978). Estimation of the biomechanical properties of three body types using a photogrammetric method. Journal of Biomechanics, 11(8–9), 349–358. https://doi.org/10.1016/0021-9290(78)90069-6
- Koga, D., Gonjo, T., Kawai, E., Tsunokawa, T., Sakai, S., Sengoku, Y., Homma, M., & Takagi, H. (2020, September 29). Effects of exceeding stroke frequency of maximal effort on hand kinematics and hand propulsive force in front crawl. Sports Biomech. Advance online publication. 1–13. https://doi.org/10.1080/14763141.2020.1814852
- Mayer, F., Horstmann, T., Röcker, K., Heitkamp, H. C., & Dickhuth, H. H. (1994). Normal values of Isokinetic maximum strength, the strength/velocity curve, and the angle at peak torque of all degrees of freedom in the shoulder. International Journal of Sports Medicine, 15(S 1), S19–25. https://doi.org/10.1055/s-2007-1021105
- McCabe, C. B., Psycharakis, S., & Sanders, R. (2011). Kinematic differences between front crawl sprint and distance swimmers at sprint pace. Journal of Sports Sciences, 29(2), 115–123. https://doi.org/10.1080/02640414.2010.523090
- McCabe, C. B., Sanders, R. H., & Psycharakis, S. G. (2015). Upper limb kinematic differences between breathing and non-breathing conditions in front crawl sprint swimming. Journal of Biomechanics, 48(15), 3995–4001. https://doi.org/10.1016/j.jbiomech.2015.09.012
- Oldfield, R. C. (1971). The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia, 9(1), 97–113. https://doi.org/10.1016/0028-3932(71)90067-4
- Psycharakis, S. G., & McCabe, C. (2011). Shoulder and hip roll differences between breathing and non-breathing conditions in front crawl swimming. Journal of Biomechanics, 44(9), 1752–1756. https://doi.org/10.1016/j.jbiomech.2011.04.004
- Psycharakis, S. G., Naemi, R., Connaboy, C., McCabe, C., & Sanders, R. H. (2010). Three-dimensional analysis of intracycle velocity fluctuations in frontcrawl swimming. Scandinavian Journal of Medicine & Science in Sports, 20(1), 128–135. https://doi.org/10.1111/j.1600-0838.2009.00891.x
- Psycharakis, S. G., & Sanders, R. H. (2008). Shoulder and hip roll changes during 200-m front crawl swimming. Medicine and Science in Sports and Exercise, 40(12), 2129–2136. https://doi.org/10.1249/MSS.0b013e31818160bc
- Psycharakis, S., Sanders, R., Mill, F. (2005). A calibration frame for 3D swimming analysis. In Q. Wang, (Ed.), Proceedings of XVII International Symposium on Biomechanics in Sports (pp. 901–905). The China Institute of Sport Science.
- Psycharakis, S. G., & Yanai, T. (2018). How does buoyancy affect performance during a 200m maximum front crawl swim? Journal of Sports Sciences, 36(18), 2061–2067. https://doi.org/10.1080/02640414.2018.1436188
- Sanders, R. (2000). Beyond race analysis. In Proceedings of the XVIII International Symposium on Biomechanics in Sports. (pp. 3–14).
- Sanders, R. H., Chiu, C. Y., Gonjo, T., Thow, J., Oliveira, N., Psycharakis, S. G., Payton, C. J., & McCabe, C. B. (2015). Reliability of the elliptical zone method of estimating body segment parameters of swimmers. Journal of Sports Science & Medicine, 14(1), 215–224.
- Seifert, L., Chollet, D., & Chatard, J. C. (2007). Kinematic changes during a 100-m front crawl: Effects of performance level and gender. Medicine and Science in Sports and Exercise, 39(10), 1784–1793. https://doi.org/10.1249/mss.0b013e3180f62f38
- Takagi, H., Nakashima, M., Sengoku, Y., Tsunokawa, T., Koga, D., Narita, K., Kudo, S., Sanders, R., Gonjo, T. (2021, August 23). How do swimmers control their front crawl swimming velocity? Current knowledge and gaps from hydrodynamic perspectives. Sports Biomechanics / International Society of Biomechanics in Sports. 1–20. https://doi.org/10.1080/14763141.2021.1959946