Abstract
Understanding population dynamics of invasive species is critical for formulating effective management plans but dependent upon the reliability of age estimations. We compared age estimates derived from dorsal fin spines and pectoral fin rays from common carp (Cyprinus carpio), a widespread invasive species that imposes deleterious ecosystem effects. Age estimates derived from each structure were highly correlated with one another. Percentage agreement (PA) was higher (57.9%) and coefficient of variation (CV) was lower (6.6%) initially for dorsal fin spines compared to pectoral fin rays (43.5%, 9.9%, respectively), but both structures had high PA (∼95%) and low CV (0.3%) following a joint examination. Thus, both dorsal fin spines and pectoral fin rays provide similar age estimates of common carp and can be used to understand population dynamics of this species.
Introduction
Common carp (Cyprinus carpio) is highly adaptable to a suite of environmental conditions (Edwards and Twomey Citation1982; Weber et al. Citation2010), which in part has led to its widespread distribution worldwide. Although a highly prized sport fish in its native range (Panek Citation1987; Arlinghaus and Mehner Citation2003), common carp has become a nuisance in many areas of introduction as a result of its ability to achieve high densities and impose deleterious effects on aquatic ecosystems and native fauna (Parkos et al. Citation2003; Weber and Brown Citation2009). In its native range, common carp is rigorously managed as a sport fish (Panek Citation1987; Arlinghaus and Mehner Citation2003), whereas intensive management effort has been directed at control or eradication in the areas of introduction (Koehn Citation2004; Weber and Brown Citation2009; Weber et al. forthcoming). In either case, accurate and precise age information is needed to evaluate common carp population dynamics (recruitment, growth, and mortality) for successful management.
In general, otoliths are considered the preferred aging structure for a wide array of fishes due to the precision and accuracy associated with these structures (Secor et al. Citation1995). However, the use of otoliths requires the fish to be killed, which is not always feasible due to research objectives (e.g., tagging, telemetry studies) or socio-political reasons (e.g., Arlinghaus et al. Citation2007). As an alternative to otoliths, several non-lethal structures have been used to age adult common carp – primarily scales, pectoral fin rays, and dorsal fin spines (Lubinski et al. Citation1984; Jackson et al. Citation2007; Phelps et al. Citation2007). Scales appear to provide unacceptable precision for aging common carp, but both pectoral fin rays (Phelps et al. Citation2007) and dorsal fin spines (Jackson et al. Citation2007) may offer a non-lethal aging alternative to otoliths. However, the precision of these structures has not been compared. Therefore, our objective was to compare the precision of pectoral fin rays and dorsal fin spines as non-lethal aging alternatives for common carp.
Materials and methods
Aging structures from common carp were collected in 2009 from approximately 100 individuals each in Patterson Reservoir and Bowman-Haley Reservoir, North Dakota, following the application of rotenone and Jamestown Reservoir, North Dakota, using pulsed DC-boat electrofishing. Pectoral fin rays and dorsal fin spines were removed at the base of the structures near the body of the fish. Both structures were cleaned and allowed to air dry for a minimum of 2 weeks. Dermal tissue was removed, and structures were sectioned with a low-speed diamond saw (Buehler Isomet, Lake Bluff, Illinois). Two 0.8 mm cross-sections were cut from the anterior portion of the structures and secured to glass microscope slides with cyanacrylic glue.
Ages were estimated for each structure by two independent readers who were familiar with aging common carp pectoral fin rays and dorsal fin spines using a dissecting microscope. Each structure was aged independently without knowledge of fish size or age estimates derived from the other reader or paired structure. Pearson's correlation analysis was used to quantify relationships between ages derived between structures and readers. Precision was determined using exact percentage agreement (PA) and PA within 1 year. Aging precision between readers and structures was assessed using age-bias plots and the coefficient of variation (CV; Campana et al. Citation1995). The CV was calculated as
Results and discussion
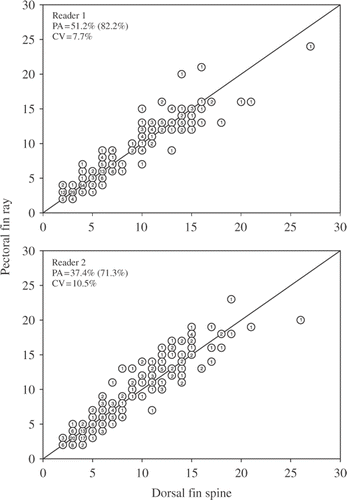
We aged 309 common carp that ranged in total length from 380 to 850 mm. Ages assigned to pectoral fin rays ranged from 2 to 24 years and dorsal fin spine ages ranged from 2 to 27 years (). Age estimates obtained from dorsal fin spines were highly correlated with those obtained from pectoral fin rays for both reader 1 (r2 = 0.97, p < 0.0001) and reader 2 (r2 = 0.97, p < 0.0001). Reader 1 obtained higher exact PA and PA within 1 year and lower CV between structures than reader 2. Exact PA between dorsal fin spine and pectoral fin ray age estimates was 51.2% for reader 1 and 37.4% for reader 2, whereas PA within 1 year was 82.2% for reader 1 and 71.3% for reader 2. CV between dorsal fin spine and pectoral fin ray estimates was 7.7% for reader 1 and 10.5% for reader 2. Both structures appeared to provide similar age estimates and did not consistently under- or over-estimate ages in comparison to the other structure.
Figure 1. Age-bias plots of dorsal fin spines and pectoral fin rays used to age common carp (N = 309) sampled from three North Dakota lakes. Precision between structures was evaluated as exact PA, PA within 1 year (value within parentheses), and mean CV. Numbers in circles represent the number of common carp of each estimated age.
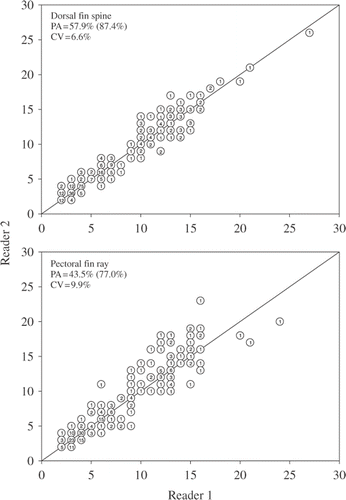
Precision of age estimates between readers was higher for dorsal fin spines than pectoral fin rays (). Ages estimated by reader 1 were highly related to ages estimated by reader 2 for both dorsal fin spines (r2 = 0.98, p < 0.0001) and pectoral fin rays (r2 = 0.97, p < 0.0001). Exact PA of age estimates between readers was 43.5% for pectoral fin rays and 57.9% for dorsal fin spines, whereas PA within 1 year was 77.0% for pectoral fin rays and 87.4% for dorsal fin spines. CV was lower for dorsal fin spines (6.6%) than pectoral fin rays (9.9%). Ages assigned to pectoral fin rays tended to vary more than those assigned to dorsal fin spines. For instance, pectoral fin rays estimated to be age-9 by reader 1 were assigned ages varying from age-5 to age-13 by reader 2. Reader 2 tended to assign older ages to pectoral fin rays than reader 1, whereas no patterns were detected with dorsal fin spines. Most disagreements between readers occurred for structures expressing at least eight annuli.
Figure 2. Age-bias plots for dorsal fin spines and pectoral fin rays used to age common carp (N = 309) sampled from three North Dakota lakes. Precision between readers was evaluated as exact PA, PA within 1 year (value within parentheses), and mean CV. Numbers in circles represent the number of common carp of each estimated age.
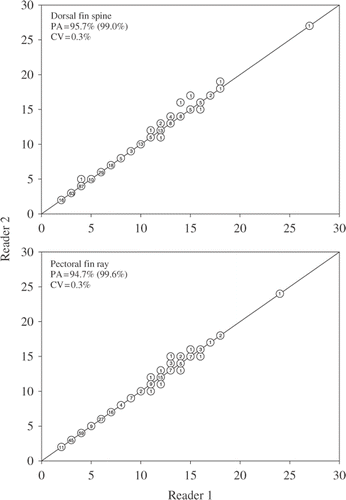
After a joint examination by both readers, PA increased and CV decreased for both dorsal fin spines and pectoral fin rays. Readers were unable to agree upon age estimates for 13 dorsal fin spines and pectoral fin rays following the joint examination. Ages estimated by reader 1 were highly correlated with those of reader 2 for both dorsal fin spines (r2 = 0.99, p < 0.0001) and pectoral fin rays (r2 = 0.99, p < 0.0001; ). Exact PA and PA within 1 year increased to 95.7% and 99.0% for dorsal fin spines and 94.7% and 99.6% for pectoral fin rays, respectively. Additionally, overall CV decreased to 0.3% for both dorsal fin spines and pectoral fin rays.
Figure 3. Age-bias plots for dorsal fin spines and pectoral fin rays used to age common carp (N = 309) sampled from three North Dakota lakes based on a joint viewing. Precision between readers was evaluated as exact PA, PA within 1 year (value within parentheses), and mean CV. Numbers in circles represent the number of common carp of each estimated age.
Obtaining accurate age estimates is critical for estimating fish population dynamics (recruitment, growth, and mortality). Spines and fin rays have become increasingly popular alternatives to lethal aging structures for fishes (Koch and Quist Citation2007). Fin rays and spines have been found to provide precise age estimates in many fishes, including white sucker (Catostomus commersonii; Sylvester and Berry Citation2006), chinook salmon (Oncorhynchus tshawytscha; Chilton and Bilton Citation1986), blue throat wrasse (Notolabrus tetricus; Metcalf and Swearer Citation2005), and Pacific cod (Gadus macrocephalus; Lai et al. Citation1987). For common carp, dorsal fin spines have been shown to be a more precise estimator of age than scales (Jackson et al. Citation2007) and indirectly validated (Wichers Citation1976), whereas pectoral fin rays have been shown to provide similar age estimates as otoliths (Phelps et al. Citation2007). Our results demonstrate that both dorsal fin spines and pectoral fin rays provide comparable age estimates for common carp. Although initial PA between structures and readers was relatively low (37.4–57.9%), both dorsal fin spines and pectoral fin rays had high (∼95%) reader agreement following a joint examination. Aging structures should yield a CV of less than 8% to provide reliable age estimates (Campana Citation2001). Initial CV values for dorsal fin spines were <8% for both between structures and readers, whereas CV for pectoral fin rays were ∼10%. Following a joint viewing by both readers, CV was low for both structures (0.3%).
Spines and rays offer some benefits compared to other structures for aging fish. Spines and rays can be quickly and easily removed and processed compared to other structures (Conand et al. Citation1995; Sun et al. Citation2001; Isermann et al. Citation2003), do not require killing the fish, and at times may be preferred over otoliths (Muir et al. Citation2008). However, the use of spines and rays can sometimes be problematic because the expansion of the central lumen can deteriorate early annuli in older fish (Kwak et al. Citation2006). Additionally, we observed that pectoral fin rays are smaller in diameter, more fragile, and were damaged more frequently during processing compared to dorsal fin spines. Additional preparation techniques (i.e., Koch and Quist Citation2007) may improve the ability to process these structures but require little additional cost or time.
Comparisons of dorsal fin spines and pectoral fin rays in the literature are rare. Ihde and Chittenden (Citation2002) found that rays and spines required a long time to process and that spines did not provide a reliable estimation of age, likely due to an enlarged lumen. Layher (Citation1981) recommended using dorsal fin spines to age flathead catfish (Pylodictis olivaris) due to large lumen in pectoral fin rays but warned that dorsal fin spines were smaller and more difficult to prepare and age than pectoral fin rays. Our results reveal that both dorsal spines and pectoral fin rays provide similar age estimates for common carp and can be relied upon when research objectives require a non-lethal aging technique.
Acknowledgements
The authors thank B. Kratz, J. Hendrickson, S. Gangl, G. Van Eeckhout, and other members of the North Dakota Game and Fish Department for collecting common carp aging structures and several technicians who aided in aging structure preparation. Partial funding for this project was provided through the Federal Aid in Sport Fish Restoration Act (Studies 1513 and 1596) administered through South Dakota Department of Game, Fish and Parks.
Related Research Data
References
- Arlinghaus , R , Cooke , SJ , Schwab , A and Cooke , IG . 2007 . Fish welfare: a challenge of the feelings-based approach, with implications for recreational fishing . Fish and Fisheries , 8 : 57 – 71 .
- Arlinghaus , R and Mehner , T . 2003 . Socio-economic characterization of specialized common carp (Cyprinus carpio L.) anglers in Germany, and implications for inland fisheries management and eutrophication control . Fisheries Research , 61 : 19 – 33 .
- Campana , SE . 2001 . Accuracy, precision and quality control in age determination, including a review of the use and abuse of age validation methods . Journal of Fish Biology , 59 : 197 – 242 .
- Campana , SE , Annand , MC and McMillan , JI . 1995 . Graphical and statistical methods for determining the consistency of age determinations . Transactions of the American Fisheries Society , 124 : 131 – 138 .
- Chilton , DE and Bilton , HT . 1986 . New method for ageing chinook salmon (Onchorhynchus tshawytscha) using dorsal fin rays, and evidence of its validity . Canadian Journal of Fisheries and Aquatic Sciences , 43 : 1588 – 1594 .
- Conand , F , Camara , SB and Domain , F . 1995 . Age and growth of three species of Ariidae (Siluriformes) in coastal waters of Guinea . Bulletin of Marine Science , 56 : 58 – 67 .
- Edwards , EA and Twomey , KA . 1982 . Habitat suitability index models: common carp , Washington, DC : US Department of the Interior Fish and Wildlife Service. . No.: FWS/OBS-82/10.12
- Ihde , TF and Chittenden , ME . 2002 . Comparison of calcified structures for aging spotted seatrout . Transactions of the American Fisheries Society , 131 : 634 – 642 .
- Isermann , DA , Meerbeek , JR , Scholten , GD and Willis , DW . 2003 . Evaluation of three different structures used for walleye age estimate with emphasis on removal and processing times . North American Journal of Fisheries Management , 23 : 625 – 631 .
- Jackson , ZJ , Quist , MC , Larscheid , JG , Thelen , EC and Hawkins , MJ . 2007 . Precision of scales and dorsal spines for estimating age of common carp . Journal of Freshwater Ecology , 22 : 231 – 239 .
- Koch , JD and Quist , MC . 2007 . A technique for preparing fin rays and spines for age and growth analysis . North American Journal of Fisheries Management , 27 : 728 – 784 .
- Koehn , JD . 2004 . Carp (Cyprinus carpio) as a powerful invader in Australian waterways . Freshwater Biology , 49 : 882 – 894 .
- Kwak , TJ , Pine III , WE and Waters , DS . 2006 . Age, growth, and mortality of introduced flathead catfish in Atlantic rivers and a review of other populations . North American Journal of Fisheries Management , 26 : 73 – 87 .
- Lai , H , Gunderson , DR and Low , LL . 1987 . Age determination of pacific cod, Gadus macrocephalus, using five ageing methods . Fishery Bulletin , 85 : 713 – 722 .
- Layher , WG . 1981 . Comparison of annulus counts of pectoral and dorsal spines in flathead catfish . Progressive Fish-Culturist , 43 : 218 – 219 .
- Lubinski , KS , Jackson , SD and Hartsfield , BN . 1984 . Age structure and analysis of carp populations in the Mississippi and Illinois Rivers , Champaign : Illinois Natural History Survey. . Aquatic Biology Technical Report 1984(9)
- Metcalf , SJ and Swearer , SE . 2005 . Non-destructive ageing in Notolabrus tetricus using dorsal spines with an emphasis on the benefits for protected, endangered and fishes species . Journal of Fish Biology , 66 : 1740 – 1747 .
- Muir , AM , Sutton , TM , Peeters , PJ , Claramunt , RM and Kinnunen , RE . 2008 . An evaluation of age estimation structures for lake whitefish in Lake Michigan: selecting an aging method based on precision and a decision analysis . North American Journal of Fisheries Management , 28 : 1928 – 1940 .
- Panek , FM . 1987 . “ Biology and ecology of carp ” . In Carp in North America , Edited by: Cooper , EL . 1 – 15 . Bethesda, MD : American Fisheries Society .
- Parkos , JJ , Santucci , VJ and Wahl , DH . 2003 . Effects of adult common carp (Cyprinus carpio) on multiple trophic levels in shallow mesocosms . Canadian Journal of Fisheries and Aquatic Sciences , 60 : 182 – 192 .
- Phelps , QE , Edwards , KR and Willis , DW . 2007 . Precision of five structures for estimating age of common carp . North American Journal of Fisheries Management , 27 : 103 – 105 .
- Secor , DH , Dean , JM and Campana , SE . 1995 . Recent developments in fish otolith research , Columbia : University of South Carolina Press .
- Sun , C , Huang , C and Yeh , S . 2001 . Age and growth of the big-eye tuna, Thunnus obesus, in the western Pacific Ocean . Fishery Bulletin , 99 : 502 – 509 .
- Sylvester , RM and Berry Jr , CR . 2006 . Comparison of white sucker age estimates from scales, pectoral fin rays, and otoliths . North American Journal of Fisheries Management , 26 : 24 – 31 .
- Weber , MJ and Brown , ML . 2009 . Effects of common carp on aquatic ecosystems 80 years after ‘Carp as a dominant’: ecological insights for fisheries management . Reviews in Fisheries Science , 17 : 524 – 537 .
- Weber , MJ , Brown , ML and Willis , DW . 2010 . Spatial variability of common carp populations in relation to lake morphology and physicochemical parameters in the upper Midwest United States . Ecology of Freshwater Fish , 19 : 555 – 565 .
- Weber , MJ , Hennen , MJ and Brown , ML . 2011 . Simulated population responses of common carp to commercial exploitation . North American Journal of Fisheries Management , Forthcoming
- Wichers , WJ . 1976 . Age and growth of carp (Cyprinus carpio) from Pathfinder Reservoir, Wyoming, 1974 and 1975 [master's thesis] , Laramie : University of Wyoming .