Abstract
Scales, opercular bones, otoliths, vertebrae, and dorsal fin rays were compared to ascertain the best aging structure of Cirrhinus mrigala. All structures showed alternating opaque and translucent bands that were interpreted as annuli; however, structures differed greatly in the clarity of their presumed annual marks. Age estimates were compared by calculating the percentage of agreement (PA), average percentage of error (APE), and coefficient of variation (CV) and by constructing age bias plots. Age estimates were evaluated between readers and between the pairs of aging structures. Among all structures, scales showed the clearest annual rings and had highest (94.6%) PA between readers, followed by opercular bones (88.6%), otoliths (83.9%), vertebrae (79.9%), and dorsal fin rays (62.4%). Due to highest PA and lower minimal aging error, the scales were considered the most reliable bony structure for aging C. mrigala. When scale ages were compared with those of the other structures, the highest PA and lowest APE and CV values were found between scale and opercular bone estimates. Mean values of age estimate from scales were comparable (p > 0.05) to the values obtained from opercular bones, otoliths, and vertebrae and were significantly (p < 0.05) different with the dorsal fin ray age estimates. Scales were the most suitable structures for age estimation in C. mrigala.
Introduction
Fish age determination is an important tool in fishery biology especially for the assessment of life history, growth rate, age at first maturity, and population dynamics. Knowledge on these aspects has important and useful applications in fisheries resource management. Age of fishes can be estimated using a variety of hard parts, including scales, otoliths, opercular bones, fin rays, cleithra, vertebrae, and urohyal bone. Some hard parts provide more accurate and precise estimates of age than others. Age estimation is often accompanied by several sources of error that can have significant effects on many population parameter estimates (Campana Citation2001). Understanding the precision of structures used for age estimation can reduce variation and increase the utility of age and growth information (DeVries and Frie Citation1996) that may provide useful information to managers.
Comparison of age estimates between structures is an alterative technique to validation that may provide useful information on the accuracy and bias of age-estimating structures (Sylvester and Berry Citation2006). Several studies have focused on comparing ages enumerated from different bony structures in an attempt to quantify the precision and to identify possible bias associated with each structure. Comparisons of age estimates from various structures have been studied for many species such as Barbus rajanorum mystaceus (Polat Citation1987a), Capoeta trutta (Polat Citation1987b), Labeo rohita, Catla catla, and Channa marulius (Khan and Khan Citation2009), Cyprinus carpio (Phelps et al. Citation2007), and Salvelinus confluentus (Zymonas and McMahon Citation2009).
Mrigal (C. mrigala) is a moderately fast growing freshwater major carp, and its fishery is of great commercial importance in the Indian subcontinent. It has a good market demand and is renowned for its good taste. River Ganga is considered as the original home of the Indian major carps (L. rohita, C. catla, and C. mrigala) and a number of other economically important freshwater fishes. Catch statistics over many years indicate a declining trend for riverine catches, both in quantitative and qualitative terms. The average yield of major carps in the River Ganga declined from 26.62 to 2.55 kg/ha per year during the past four decades. Biologically and economically desirable fish species have started to be replaced by low-value species (Upare Citation2007). Riverine fisheries play crucial roles for conservation of biodiversity as they are considered the primary source of original fish germplasm. It warrants monitoring and collection of basic biological information on the available fish stocks. In previous studies, age and growth investigations of C. mrigala were conducted using scales only (Jhingran Citation1957; Kamal Citation1969). To date no published information is available on the precision of age estimates from different aging structures in C. mrigala. Therefore, this study was undertaken with the objective to evaluate and compare the five different aging structures (i.e., scales, otoliths, vertebrae, opercular bones, and dorsal fin rays) in order to identify and quantify the differences in precision and bias between readers and between the pairs of aging structures.
Materials and methods
A total of 464 specimens (209 males and 255 females) of C. mrigala were collected monthly during the period January 2008 to January 2010 from Narora (27°0′ N, 78°25′60 E), and Kanpur (26°28′ N, 80°24′ E) sites of the River Ganga, UP, India. Total length (TL) was measured to the nearest centimeter. Body weight was recorded to the nearest gram as total weight (TW), including gut and gonads.
Preparation of aging structures and reading techniques
Scales were removed using forceps from above the lateral line near the tip of the pectoral fin. Scales were washed, cleaned, and studied as dry mounts after removing the extraneous matter and mucous by washing them in tap water and rubbing in between the finger tips. To make scales more clear and soft, the large scales were dipped in weak solution (1%) of KOH for about 5–10 min, and then washed in tap water and dried in air (Khan and Khan Citation2009). Since the scales attain a fairly large size and are translucent, they were read with the help of a magnifying glass. However, the scales of smaller fishes were observed under a microscope (Khan and Siddiqui Citation1973).
The opercular bones were detached with a scalpel and dipped in boiling water for a few minutes to remove extraneous tissue. A bristled brush was used to remove tissue that boiling water did not loosen. Cleaned opercular bones were examined under transmitted fluorescent light with the naked eye (Phelps et al. Citation2007).
Sagittal otoliths (the largest of their kind) were removed from otic capsules. Otoliths were washed, cleaned, and read whole by immersion in alcohol and examined under microscope using reflected light. Otoliths with unclear annual rings were ground with sandpaper to make the annuli more distinct (Khan and Khan Citation2009).
Vertebrae (4th to 10th) were removed and placed in boiling water for 10–15 min to clear the attached muscles. All processed vertebrae were then dried for 2 weeks to count annual rings under a dissecting microscope (Phelps et al. Citation2007).
The first four dorsal fin rays were removed from each specimen. Fin rays were sectioned using jeweler's saw. If required, some of the sections were polished with sandpaper, and a drop of immersion oil was used to improve the clarity (Phelps et al. Citation2007). Sections were placed on a microscope slide and viewed under a dissecting microscope.
All the otoliths, scales, vertebrae, opercular bones, and dorsal fin rays were aged independently by two readers without the knowledge of fish length, weight, or date of collection. In case of disagreement between readers, the aging structures were re-examined together by the two readers until a consensus was reached (Khan and Khan Citation2009).
Calculations and statistical analyses
Precision was measured by calculating the PA, coefficient of variation (CV), and average percentage of error (APE; Beamish and Fournier Citation1981) between the readers and between the pairs of aging structures. Age bias graphs (Campana et al. Citation1995) were constructed to examine potential biases between readers. PA was estimated as the proportion of each age on which both readers agreed.
Mean age readings (consensus data) obtained from various hard anatomical parts were subjected to one-way analysis of variance (ANOVA) followed by Duncan's multiple range test (DMRT) (Gomez and Gomez Citation1984) in order to explain whether the readings from different hard anatomical parts of the fish showed significant differences among themselves. Although mean age estimate is not an indicator for the reliability of aging structure, it may provide useful information regarding over or under estimation of age by a structure irrespective of fish size-class. This may prove useful in selecting the structures which may give statistically indifferent readings when size-class is not taken into account. All statistical analyses were done using MS-Excel and SPSS (version 12.0).
Results
PA of ages between the age readings of two readers was highest for scales (94.6%) followed by opercular bones (88.6%), otoliths (83.9%), vertebrae (79.9%), and dorsal fin rays (62.4%). CV (2.78%) and APE (1.96%) were lowest for age estimates from scales (). Age bias graphs between two readers for scales, opercular bones, otoliths, vertebrae, and dorsal fin rays are presented in . In case of scale and opercular bones, no age bias was found between readers, while small error was present in otoliths and vertebrae. Disagreement between readers increased with age for dorsal fin rays, as indicated by larger standard error bars for older fishes. In the case of dorsal fin rays, there was also a consistent pattern of underestimation of ages. When scale age estimates were compared with other alternative structures, highest PA was found between scales-opercular bones (94%), followed by scales-otoliths (86.2%), scales-vertebrae (69.2%), and scales-dorsal fin rays (67.2%). Scales and opercular bones showed lowest values of APE and CV (). Mean values of age estimates from different structures, when compared using ANOVA followed by DMRT, showed that maximum age estimates obtained from scales were significantly (p < 0.05) higher from dorsal fin rays, but comparable (p > 0.05) to the values obtained from opercular bones, vertebrae, and otoliths ().
Figure 1. Age bias graphs between two independent readers for scales, opercular bones, otoliths, vertebrae, and dorsal fin rays age estimates. Each error bar represents the 95% confidence interval, and the solid line indicates the theoretical 1 : 1 agreement line of age estimates between readers. Points above the line indicate ages that were overestimated, whereas points below the line indicate ages that were underestimated.
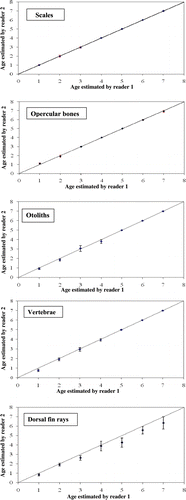
Table 1. Comparison of PA, APE, and CV between the age readings of two independent readers in C. mrigala.
Table 2. Comparison of PA, APE, and CV of the age readings between hard anatomical structures in C. mrigala.
Table 3. Comparison of mean values of age estimates from different bony parts in 464 C. mrigala.
Discussion
Selection of the appropriate method for age and growth determination in fishes often requires balancing precision and accuracy of the method with sample size limitations (DeVries and Frie Citation1996). In this study, variations were observed in the age estimates from different aging structures. Scales showed clearer and sharper growth rings in comparison to other structures. In the published literature available on Indian freshwater fishes as well as in most of the cyprinids, scales have been exclusively used for age studies in C. catla (Natarajan and Jhingran Citation1963), C. mrigala (Kamal Citation1969), and L. rohita (Khan and Siddiqui Citation1973). According to Jhingran (Citation1957), Natarajan and Jhingran (Citation1963), and Kamal (Citation1969), in C. mrigala and C. catla scales showed clear annulations. In the first published report on precision of age estimates from different aging structures of Indian freshwater fishes, it was observed that in L. rohita, C. catla, and C. marulius, among different structures (scales, otoliths, opercular bones, vertebrae, and dorsal fin rays), scales were the most suitable structure for aging L. rohita and C. marulius, and opercular bones were best for C. catla (Khan and Khan Citation2009). In addition to having clear and sharp annuli, scales also have the advantages of easy collection, which is non-destructive to the fish. Several researchers have reported that scales can provide unreliable estimates of fish age (DeVries and Frie Citation1996; Maceina and Sammons Citation2006), which has often been attributed to reabsorption and deposition of false annuli due to stress and food limitation and also because clarity of annuli is affected with increase in fish age. Annuli in scales of C. mrigala tended to show gradual decrease in clarity and sharpness beyond the age of 7 years, even though the annuli were quite readable. Almost similar observations were also found for opercular bones.
Opercular bones provided age readings very close to the scales as evident from the PA, APE, and CV values between structures and the age bias graph between readers. Opercular bones were reported to be superior to all other structures in common carp, (C. carpio; McConnell Citation1952) and in C. catla (Khan and Khan Citation2009). Sharp and Bernard (Citation1988) reported that opercles provided age estimates nearly identical to those from otoliths and suggested the use of opercles in place of otoliths for many age and growth studies on lake trout (Salvelinus namaycush) from Alaskan lakes. Khemiri et al. (Citation2005) found opercular bones unsuitable for age estimation in Boops boops from Tunisian waters. Jimenez-Badillo (Citation2006) mentioned that the extraction, preparation, and reading of opercular bones in comparison with scales were relatively easier and cited that the rings on scales and opercular bones were due to periods of fast or slow growth imposed by reproductive energetic demands in blue tilapia (Oreochromis aureus).
In C. mrigala, PA and APE for age estimates using otoliths between readers were 83.9% and 9.48%, respectively. Clarity and sharpness of annuli on otoliths did not seem to be affected by age group. Otoliths have often been used for accurate age estimation as they do not show reabsorption and have acellular growth. Otoliths are also reported to be metabolically inert and thus do not reflect physiological changes that may occur throughout the life of fish (Phelps et al. Citation2007). Since size of otoliths in C. mrigala is relatively smaller than other fishes, the readability was easier and accurate for larger fishes compared to smaller ones. Otoliths were reported to be inferior to scales and vertebrae for age estimation in Leuciscus cephalus orientalis (Ozdemir and Sen Citation1986). Sylvester and Berry (Citation2006) reported that otoliths were more accurate than scales and pectoral fin rays for age estimation in white sucker (Catostomus commersonii). Isermann et al. (Citation2003), while comparing the scales, sagittal otoliths, and dorsal spines, reported that otoliths provided the most time-efficient and precise approach for age estimation in walleye (Sander vitreum).
Vertebrae provided precise age estimates, similar to otoliths, in burbot (Lota lota; Guinn and Hallberg Citation1990). In this study, the rings on vertebral centra were not very clear and showed numerous minute marks unrelated to cyclic events, which corroborates the observations made by Khemiri et al. (Citation2005) in B. boops and Khan and Khan (Citation2009) in L. rohita, C. catla, and C. marulius. Clark (Citation1987) reported that the time required to process and read vertebrae (20 times as long as scales) made them less practical to use but that precision and accuracy involved with vertebrae made them the best among the three structures (vertebrae, otoliths, and scales) for the age estimation of fall chum salmon (Onchorhynchus keta).
Fin rays are considered a non-lethal alternative to internal bones (DeVries and Frie Citation1996; Zymonas and McMahon Citation2009). Some studies suggested that aging was more precise with fin rays as compared to other structures. Dorsal fin rays were more suitable than other structures for aging B. rajanorum mystaceas (Polat Citation1987a) and C. trutta (Polat Citation1987b). However, other investigators rejected fin rays on account of difficulties with sample preparation, identification of the first annulus, and distinction between true and false annuli (Zymonas and McMahon Citation2009). Quinn and Ross (Citation1982) questioned the reliability of age estimates from pectoral fin rays in fish older than 7 years because of the lack of annulus formation and the difficulty in interpreting fin ray sections. Thus, suitability of fin rays for aging may vary among species and potentially among populations (Zymonas and McMahon Citation2009). Due to difficulty in the correct identification of annuli in dorsal fin rays, fish age may be underestimated, leading to imprecise estimation of growth and other biological parameters. In this investigation, the annuli of dorsal fin rays in C. mrigala showed less clarity than other structures for age estimation.
Scales were found to be the most reliable bony structure for aging C. mrigala up to 7 years of age. If size-class is not taken into account, then all the selected aging structures except dorsal fin rays could be used for mean age estimation in C. mrigala with no significant differences in their readings in comparison to the most suitable aging structures, the scales.
Acknowledgement
The authors thank the Chairman, Department of Zoology, Aligarh Muslim University, Aligarh, India, for providing the necessary facilities for the study.
References
- Beamish , RJ and Fournier , DA . 1981 . A method for comparing the precision of a set of age determinations . Canadian Journal of Fisheries Aquatic Science , 8 : 982 – 983 .
- Campana , SE . 2001 . Accuracy, precision, and quality control in age determination, including a review of the use and abuse of age validation methods . Journal of Fish Biology , 59 : 197 – 242 .
- Campana , SE , Annand , MC and McMillan , JI . 1995 . Graphical and statistical methods for determining the consistency of age determinations . Transactions of the American Fisheries Society , 124 : 131 – 138 .
- Clark , RA . 1987 . Sources of variability in three ageing structures for Yukon river fall chum salmon Onychorhynchus keta walbaums escapement samples , Proceedings of the 1987 Northeast Pacific Pink and Chum Salmon Workshop, Alaska Department of Fish and Game, Juneau, Alaska, p. . 11–121
- DeVries , DR and Frie , RV . 1996 . “ Determination of age and growth ” . In Fisheries Techniques, , 2nd , Edited by: Murphy , BR and Willis , DW . 483 – 512 . Bethesda (MD) : American Fisheries Society .
- Gomez , KA and Gomez , AA . 1984 . Statistical procedures for agricultural research , 680 Singapore : John Wiley and Sons .
- Guinn , DA and Hallberg , JE . 1990 . Precision of estimated ages of burbot using vertebrae and otolith , Juneau, Alaska : Alaska Department of Fish and Game. . Fishery Data Series no. 90-17
- Isermann , DA , Meerbeek , JR , Scholten , GD and Willis , DW . 2003 . Evaluation of three different structures used for walleye age estimation with emphasis on removal and processing times . North American Journal Fisheries Management , 23 : 625 – 631 .
- Jhingran , VG . 1957 . Age determination of the Indian major carps, Cirrhinus mrigala (Hamilton) by means of scales . Nature , 79 : 468 – 469 .
- Jimenez-Badillo , L . 2006 . Age-growth models for tilapia Oreochromis aureu (Perciformes, Cichilidae) of the infiernillo reservoir, Mexico and reproductive behaviour . Revista de Biologia Tropical , 54 : 577 – 588 .
- Kamal , MY . 1969 . Studies on the age and growth of Cirrhina mrigala (Hamilton) from the river Yamuna at Allahabad . Proceedings of National Academy of Sciences India , 35 : 72 – 92 .
- Khan , MA and Khan , S . 2009 . Comparison of age estimates from scale, opercular bone, otolith, vertebrae and dorsal fin ray in Labeo rohita (Hamilton), Catla catla (Hamilton) and Channa marulius (Hamilton) . Fisheries Research , 100 : 255 – 259 .
- Khan , RA and Siddiqui , AQ . 1973 . Studies on the age and growth of rohu, Labeo rohita (Ham.) from a pond (moat) and rivers Ganga and Yamuna . Proceedings of National Institute of Sciences of India , 39 : 583 – 597 .
- Khemiri , S , Gaamour , A , Zylberberg , L , Meunier , F and Romdhane , MS . 2005 . Age and growth of bogue, Boops boops, in Tunisian waters . Acta Adriatica , 46 : 159 – 175 .
- Maceina , MJ and Sammons , SM . 2006 . An evaluation of different structures to age freshwater fish from a northeastern US river . Fisheries Management and Ecology , 13 : 237 – 242 .
- McConnell , WJ . 1952 . The Opercular bone as an indicator of age and growth of the carp, Cyprinus carpio Linnaeus . Transaction of American Fisheries Society , 81 : 138 – 149 .
- Natarajan , AV and Jhingran , AG . 1963 . On the biology of Catla catla (Ham.) from the river Jamuna . Proceedings of National Institute of Sciences India , 29 : 326 – 355 .
- Ozdemir , N and Sen , D . 1986 . Age determination by scale, vertebra and operculum of Leuciscus cephalus orientalis (Nordmann, 1840) in the Euphrates . Journal Firat University , 1 : 101 – 111 .
- Phelps , QE , Edwards , KR and Willis , DW . 2007 . Precision of five structures for estimating age of Common carp . North American Journal of Fisheries Management , 27 : 103 – 105 .
- Polat , N . 1987a . Age determination methods of Barbus rajanorum mystaceus (Heckel) living in Keban Dam Lake (in Turkish) . VIII National Biology Kongress , 2 : 575 – 588 .
- Polat , N . 1987b . Age determination of Capoeta trutta (Heckel) in Keban Dam Lake . Turkish Journal of Zoology , 11 : 155 – 160 .
- Quinn , SP and Ross , MR . 1982 . Annulus formation by white suckers and the reliability of pectoral fin rays for ageing them . North American Journal of Fisheries Management , 2 : 204 – 208 .
- Sharp , D and Bernard , DR . 1988 . Precision of estimated ages of lake trout from five calcified structures . North American Journal of Fisheries Management , 8 : 367 – 372 .
- Sylvester , RM and Berry Jr , CR . 2006 . Comparison of white sucker age estimates from scales, pectoral fin rays, and otoliths . North American Journal of Fisheries Management , 26 : 24 – 31 .
- Upare , MA . 2007 . “ Credit and microfinance programmes in inland capture fisheries in West Bengal and Assam, India ” . In Credit and microfinance needs in inland capture fisheries and conservation in Asia. FAO Fisheries Technical Paper No. 460 , Edited by: Tietze , U , Siar , SV , Marmulla , G and Anrooy , RV . 89 – 107 . Rome : FAO of United Nations .
- Zymonas , ND and McMahon , TE . 2009 . Comparison of pelvic fin rays, scales and otoliths for estimating age and growth of bull trout, Salvelinus confluentus . Fisheries Management and Ecology , 16 : 155 – 164 .