Abstract
Pampean streams are characterized by herbaceous riparian vegetation and a great abundance of primary producers. In the past decades, the banks of many water courses in the Pampa Ondulada (Buenos Aires, Argentina) have been invaded by Gleditsia triacanthos. Therefore, the original herbaceous vegetation has been replaced by trees, leading to possible consequences in the functioning of the system. We compared the rate of litter breakdown and the structure of the communities of zoosporic true fungi and heterotrophic straminipiles associated with the litter in sites invaded and not invaded by G. triacanthos. Litter bags with two different mesh sizes were placed at three invaded and three non-invaded sites in three streams. In the invaded sites, leaves in the bags with larger mesh size decomposed at a faster rate than those in non-invaded sites, but the community structure and species composition of zoosporic true fungi and heterotrophic straminipiles did not differ. In addition, some variables related to the particulate material were significantly different between invaded and non-invaded sites. In non-invaded sites, where fine sediments were abundant, leaf burial may have played an important role by diminishing the fragmentation of the leaves or changing the environmental conditions for detritivorous invertebrates, thus determining the slow breakdown rates observed in the non-invaded sites.
Introduction
Riparian vegetation is one of the primary controlling factors in the ecology of streams. It can strongly modify the amount of solar energy that reaches the water surface and hence influence stream temperature; it can help stabilize banks; and, it functions as a buffer zone for the stream, filtering the input of particulate matter and controlling the flow of nutrients toward the stream channel. As a consequence, riparian vegetation plays a significant role in stream ecosystem functioning (Cummins et al. Citation1984; Lecerf et al. Citation2005), particularly in riparian forests where it supplies the food web with organic matter for heterotrophic organisms (Barling and Moore Citation1994; Hood and Naiman Citation2000; Sabater et al. Citation2000).
Litter breakdown is the result of physical and chemical factors, which cause mechanical fragmentation and leaching, and of the activities of invertebrates and microorganisms such as aquatic fungi and bacteria (Hieber and Gessner Citation2002). The fungal assemblages involved in leaf breakdown in streams include representatives of Ascomycota (especially the Hyphomycetes), zoosporic true fungi (Blastocladiomycota and Chytridiomycota), heterotrophic straminipiles (Peronosporomycota, Hyphochytriomycota, and Thraustochytriomycota), and to a lesser extent, Zygomycota and Basidiomycota (Dix and Webster Citation1995; Gessner et al. Citation2007). Although the aquatic Hyphomycetes have traditionally been considered the group most commonly found during leaf decomposition (Bärlocher Citation1992; Suberkropp Citation1992; Baldy et al. Citation1995; Chauvet and Suberkropp Citation1998; Nikolcheva and Bärlocher Citation2004; Seena et al. Citation2008; Gulis et al. Citation2009), Marano et al. (Citation2011) have recently emphasized the relative importance of zoosporic fungi and straminipiles in the decomposition of leaves, and until now very few studies have focused on the communities of zoosporic organisms during litter decomposition in fresh waters (Schoenlein-Crusius and Milanez 1989, 1998; Schoenlein-Crusius et al. 1990, 1992; Schoenlein-Crusius et al. Citation1999).
Microbial activity and the leaf decomposition rate in streams are affected by environmental factors such as dissolved oxygen concentration, temperature, concentration of nutrients, and pH (Kuehn and Suberkropp Citation1998; McLatchey and Reddy Citation1998; Lee and Bukaveckas Citation2002; Parnrong et al. Citation2002). Since there are strong linkages among terrestrial producers, aquatic consumers, and microbial decomposers (Vanotte et al. Citation1980; Webster and Benfield Citation1986; Cummins et al. Citation1989; Bärlocher Citation1992; Graça et al. Citation2001), changes in the diversity and in the composition of riparian forests disrupt the functioning of streams and the structure of the communities developed in them. In particular, changes in environmental conditions may alter the activity of microbes and invertebrates and thus the rate of litter decomposition (Bunn et al. 1999; Bärlocher and Graça Citation2002; Laitung and Chauvet Citation2005; Lecerf et al. Citation2007).
Many studies have analyzed the decomposition process of leaf litter in streams that have suffered alterations in their riparian vegetation. However, the conclusions obtained in these studies are somewhat contradictory, especially regarding leaf decomposition rates and the microbiota associated with decomposing leaves. For example, Webster and Benfield (Citation1986) observed a decrease in the rate of decomposition and changes in the species composition, associated with deforestation; in turn, Bärlocher and Graça (Citation2002) did not find any differences in decomposition rate, while they did find that the structure of the communities of hyphomycetes differed between forested and non-forested streams. In contrast, Lecerf et al. (Citation2005) found that the diversity of the riparian vegetation influenced both the diversity of the fungal communities developed on the leaves and the decomposition rate.
Pampean streams are characterized by having low current velocities because of low slope, high nutrient levels (Feijoó et al. Citation1999), and herbaceous riparian vegetation and therefore high light availability. In correspondence with these characteristics, the streams belonging to the Luján River basin (Buenos Aires, Argentina) have been described as presenting great abundance and variety of phytobenthic species (Giorgi et al. Citation2005) and of submersed, free-floating, and emergent macrophytes (Gantes and Sánchez Caro Citation2001), biota that makes them mainly autotrophic systems (Vilches Citation2005).
Gleditsia triacanthos (honey locust) is an exotic and invasive species (Ghersa et al. Citation2002; Kalesnik and Malvárez Citation2003; Chaneton et al. Citation2004) that has colonized the banks of many watercourses in the northeastern Buenos Aires province. This invasion is particularly significant at the main course of the Luján River and is likely to continue along the basin (Cappello and de la Peña Citation2007).
Biological invasions are known to alter the composition and community structure of invaded areas (Vitousek Citation1990). In the case of the pampean streams, this alteration is particularly significant since colonization by honey locust constitutes a change not from one tree species to another (as is most common), but from herbaceous vegetation to trees. Hence, on one hand, the alteration decreases the levels of incident light and, thus, as a consequence, decreases the abundance of producers. On the other hand, it creates a new input of allochthonous organic matter (leaves). It is unclear how an ecosystem based on autochthonous production would respond to these changes.
The decomposition rate can be used as a variable to evaluate the impact of alterations on stream ecosystem functioning (Gessner and Chauvet Citation2002). It can be expected that an invasion of honey locust would influence ecosystem function (i.e., decomposition rates) through changes in environmental conditions as well as in the decomposer communities. In this study, we compared the decomposition rates of leaves of G. triacanthos and the structure of the assemblages of zoosporic fungi and straminipiles associated with the decomposing material in sites invaded and non-invaded by G. triacanthos.
Materials and methods
Study area
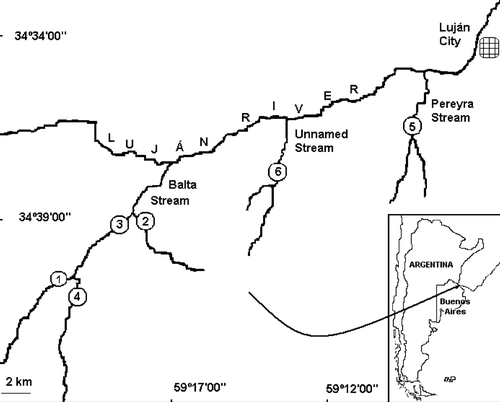
The Luján River basin is located in the Pampa Ondulada subregion in east-central Argentina. The pampa is an extensive grassland ecosystem, most of which is subjected to agriculture and cattle farming (Viglizzo et al. Citation2006). The Luján River basin covers an area of 2190 km2, has a total length of 102 km, and has an average slope for the middle basin of 0.45 m km−1 (Instituto Nacional del Agua Citation2007). Six sites on three streams in the middle part of the Luján River basin were studied (). Gleditsia triacanthos is extensively and densely established along the Luján River itself but is also incipient along the banks of the basin's streams. For this reason, the sites invaded by honey locust (shaded sites) were in one unique sub-basin on the Balta Stream (third-order) and were identified as sites 1, 2, and 3. Non-invaded sites (unshaded) were located on another tributary of the Balta Stream (site 4; first-order), in the Pereyra Stream (site 5; third-order), and in an unnamed stream (site 6; third-order).
Figure 1. Study area and locations of sampling sites. Invaded sites: 1–3; non-invaded sites: 4–6.
Analysis of environmental variables
The following physicochemical variables were measured at each site in fall, winter, spring, and summer: stream flow and discharge, photosynthetic active radiation, particulate organic matter (POM), sediment fractions, temperature, pH, conductivity, and dissolved oxygen, nitrate, and soluble reactive phosphorus (SRP) concentrations. Discharge was measured by adding a solute as a single pulse to the stream (Elosegi et al. Citation2009). Stream flow velocity was estimated as the ratio between the reach length and the elapsed time when the flowing water reached its maximum conductivity at the downstream measuring point. At site 2, on two of the sampling occasions, the method was not applied due to low discharge; instead, the complete flow of water was collected in a container over 1 min, and the accumulated volume was recorded; this procedure was then repeated three times, and the results were averaged. In these cases, stream velocity was considered to be zero. Photosynthetic active radiation was measured at the surface and subsurface of the water using a spherical quantum sensor (Li-cor 250), and from this data the percentage of light transmitted in water was estimated.
Three fractions of sestonic POM were measured – coarse >2 mm (CPOM), fine >50 µ (FPOM), and ultrafine particulate matter and organic matter >0.70 µ (UFPM and UFPOM). The concentrations of CPOM and FPOM were estimated by collecting the material in drift nets placed into the stream for 1 h. The concentration of UFPM/UFPOM was obtained by filtering a water sample of 500 mL. The dry weight (at 105°C) and the ash-free dry weight (AFDW) (at 500°C) of the collected material were then estimated.
Sediment fractions were determined by taking the upper 2 cm of two cores (3.5 cm in diameter) at each site. The fraction <1 mm was separated using a sieve, and its percentage of dry weight in relation with the whole sample was determined.
Conductivity, pH, and dissolved oxygen concentration were measured with field electrodes (Hanna Electronics, HI 9023, and HI 9033). Nitrate concentrations were measured by colorimetric methods using a Hach DR 2000 spectrophotometer, and SRP was determined according to APHA (Citation1998).
Decomposition analysis
Leaf breakdown rates were estimated using the litter bag technique (Boulton and Boon Citation1991). Leaf litter of G. triacanthos was collected at the study sites. The leaves (leaflets) were picked from the ground soon after abscission. The leaves were rinsed with water and dried at 50°C. Fifteen bags (20 × 20 cm) with 1.60 ± 0.01 g of leaflets of each of the mesh sizes employed (1 and 0.3 mm) were prepared. Since the average width of the leaflets was 5 mm, a mesh size of 1 mm was selected as the biggest in order to prevent the loss of leaflets. At each site, bags were placed in the stream, floating sub-superficially, and tethered to bricks. Three litter bags were collected on each of four occasions after 80, 156, 187, and 216 days of immersion and transported to the laboratory in polyethylene bags. Litterbag contents were washed under tap water to remove the sediment. Litter was dried at 70°C to constant weight, and the AFDW was determined to the nearest 0.1 mg after combustion at 500°C. The remaining weight of the material in the bags after 8 days of exposure was considered as the initial AFDW. The AFDWs were adjusted to a negative exponential model in order to estimate breakdown rates (k) (Olson Citation1963).
Analysis of the fungal communities
The communities of zoosporic fungi and straminipiles associated with the remaining litter material in invaded and non-invaded sites were assessed after 216 days of exposure from the bags of 0.3 mm mesh size. This choice was made based on the assumption that, with finer mesh-size bags, loss of litter is less and the potential for shredders which feed on mycelia and zoospores to enter the bags is lower. Litter bags were processed within 1 h after collection. The material of each bag was placed in a plastic sieve of 1000 μ mesh and washed repeatedly with tap water to remove sediment. The baiting technique was used (Sparrow Citation1960; Stevens Citation1974; Barr Citation1987). Twenty leaflets of similar size were placed in a Petri dish with 30 mL of sterile deionized water and incubated with five discs (5 mm in diameter) of Zea mays leaves or five Sesamum indicum seeds. Each dish was considered a sample unit, and three replicates were made for each substrate. Dishes were incubated at room temperature for 4–60 days. The species composition of zoosporic true fungi and heterotrophic straminipiles (Chytridiomycota and Peronosporomycota) was determined. Taxonomic identifications were made according to Coker (Citation1923), Sparrow (of environmental variables between invaded and non-invaded sites was performed 1960), Karling (Citation1977), and Rocha and Pires-Zottarelli (Citation2002).
Statistical analysis
The comparison of environmental variables between invaded and non-invaded sites was performed using the Mann–Whitney test (Zar Citation1999). A principal component analysis (PCA) was also performed with the environmental variables. Some variables were log(x + 1), squareroot or, arcsine-squareroot transformed to achieve normality and reduce skew and kurtosis.
We compared the rate of decomposition (k) and the percentage of leaf mass remaining for invaded and non-invaded sites using the Mann–Whitney test.
The number of isolations (abundance) of zoosporic fungi and straminipiles was calculated according to Marano et al. (Citation2008) and Marano et al. (Citation2011). Community structure was analyzed by species richness, Shannon diversity index, and Bray-Curtis (Sorensen quantitative) similarity index (Magurran Citation2004; Colwell Citation2005). The Mann–Whitney test was used to compare richness, number of isolations, and Shannon diversity indexes of zoosporic fungi and straminipiles between invaded and non-invaded sites.
Results
The UFPM and UFPOM sestonic fractions, the percentage of transmitted light, the water conductivity, and the percentage of fine sediments were significantly different between invaded and non-invaded sites ().
Table 1. Environmental variables measured in invaded and non-invaded sites.
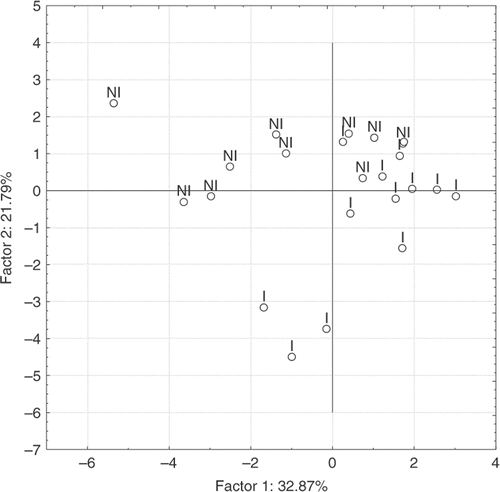
The PCA of environmental variables showed that axis 1 and axis 2 accounted for 54.7% of the total variance () and that both were related to the transportation of particulate material. UFPM, UFPOM, and CPOM were the variables that contributed the most to the variation on axis 1, whereas, those that contributed most to the variation on axis 2 were discharge, FPOM, CPOM, and percentage of transmitted light. A certain degree of separation between invaded (I) and non-invaded sites (NI) was observed in the space defined by the first two axes. ().
Figure 2. PCA ordination of the environmental variables.
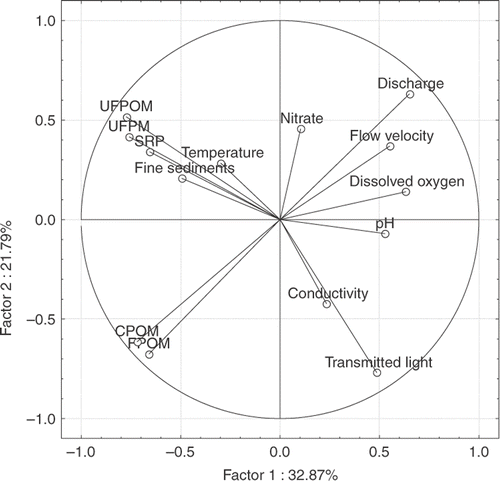
Figure 3. Sites distribution along the first two axes of PCA ordination diagrams.
At the end of the exposure period (216 days), the mean remaining leaf weight in the bags of larger mesh size was significantly lower in invaded (2%) than in non-invaded sites (45%) (p < 0.05), whereas the remaining dry weight in the bags of fine mesh size was not significantly different between invaded (30.5%) and non-invaded sites (42%). In the bags of larger mesh size, the leaves of G. triacanthos decomposed significantly faster (p < 0.05) in invaded (k = 0.0110/day−1) than in non-invaded sites (k = 0.0060 day−1), whereas, no differences of statistical significance were found in the bags of fine mesh size in invaded (k = 0.0047 day−1) or non-invaded sites (k = 0.0040 day−1) (). The rates of decomposition were different (p < 0.05) between bags of larger and smaller mesh size in invaded sites, but not in non-invaded sites ().
Figure 4. Decomposition rates of the leaves of G. triacanthos (k) estimated from invaded (I) and non-invaded (NI) sites and from bags of big mesh (BS) and small mesh (SS) size.
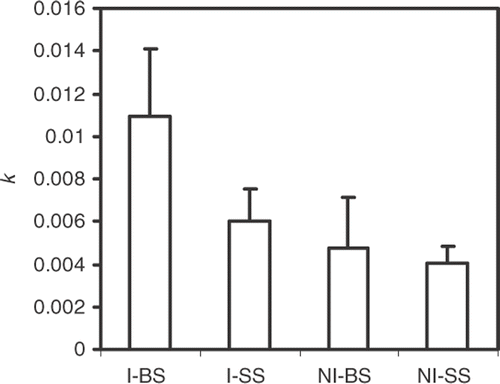
Table 2. Decomposition rates (k) and regression coefficients (R 2) from invaded (1–3) and non-invaded (4–6) sites, and from bags of two mesh sizes.
We recorded 15 taxa – six Chytridiomycota and nine Peronosporomycota. Eleven of them were recorded in the invaded sites, whereas only seven were recorded in the non-invaded sites. Some species were exclusive of invaded sites (Achlya sp., Cladochytrium replicatum, Protoachlya paradoxa, and Rhizophlyctis sp.), while some others were exclusive of non-invaded sites (Saprolegnia sp.) (). The values for the Bray–Curtis similarity index (IS) were, on average, low for both kinds of sites (ISI = 0.31 and ISNI = 0.29) and also for the totality of the samples (IS = 0.30). Neither mean richness (S) nor the mean Shannon diversity (H′) index differed between invaded and non-invaded sites (SI = 8.0 ± 2.60, SNI = 6.30 ± 0.58; = 1.64 ± 0.41,
= 1.61 ± 0.05). Although the number of isolations of each taxon was greater in invaded than in non-invaded sites (), the total number of isolations (Ni) of zoosporic organisms did not differ significantly (p > 0.05) between invaded (I = 554) and non-invaded sites (NI = 453).
Figure 5. Number of isolations (NI) of the taxa in G. triacanthos invaded and non-invaded sites.
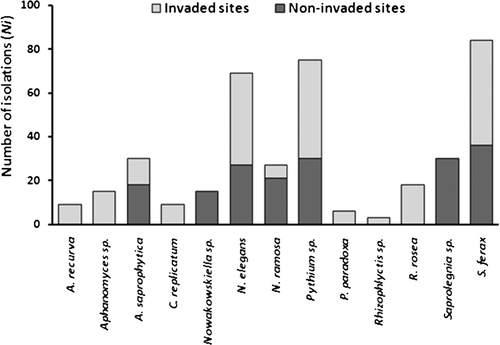
Table 3. Presence of species (*) of zoosporic true fungi and heterotrophic straminipiles in sites invaded and not invaded by G. triacanthos.
Discussion
The results show that the invasion of streambanks by G. triacanthos constitutes a factor of stress that has led to an increase in the leaf breakdown rates at invaded sites. Gessner and Chauvet (Citation2002) observed that breakdown rates may increase or decrease with disturbance, so these changes could be used as indicators of functional stream integrity.
The replacement of herbaceous vegetation by G. triacanthos in the streambanks alters the shading and the input of allochthonous organic matter (i.e., leaves). However, the variables that significantly contributed to the differentiation between invaded and non-invaded sites were those related to the transportation of particulate materials. In sites invaded by honey locust a smaller amount of fine sediments was found. In agreement with these results, Sweeney et al. (Citation2004) found that deforestation increases the deposition of sediments, and Micheli and Kirchner (Citation2002) observed that different types of riparian vegetation have different capacities for enhancing streambank stability. On several occasions, we observed that the litter bags in non-invaded sites became buried by sediments, and thus factors associated with the transport and deposition of particulate materials might explain the differences found in decomposition rates. Sedimentation reduces the surface area for microbial colonization and invertebrate activity, limits physical abrasion, and decreases oxygen availability (Bunn Citation1988, Sponseller and Benfield Citation2001); consequently, the decomposition process becomes slower. Webster and Benfield (Citation1986) observed a decrease in the rates of decomposition associated with deforestation that led to an increase in litter burial. We observed, as well as Pascoal et al. (Citation2005), that in the sites with herbaceous banks, when the litter bags had been buried by sediments, the material remained intact up to the end of the sampling period. In this way, as a consequence of the burial of the leaves, low rates of decomposition of G. triacanthos's leaf litter could be expected to occur over extended reaches in pampean streams. However, Gantes et al. (Citation2005) performed field decomposition assays in streams of the same basin, using macrophyte leaves as substrate, and found higher rates of decomposition (e.g., k = 0.0203/day−1). Since the main natural source of organic matter in these streams is autochthonous in origin and this material decomposes at a fast rate due to its chemical composition, burial, and the consequent delay in litter breakdown are unlikely to happen.
Differences in the decomposition rate between both types of sites were only found for the bags of larger mesh size. One plausible explanation for this result is that it might be related to the activity of shredder invertebrates (Lecerf et al. Citation2005). However, this functional group is not well-represented in pampean streams, and it was rarely observed in the studied sites (Rigacci Citation2009). Apart from probable differences in the organisms that had access to each type of bag, leaves in fine mesh bags stay more confined and thus, less oxygenated and less vulnerable to leaching, abrasion, and fragmentation (Triska Citation1970). In contrast, leaves in larger mesh bags are more exposed to physical abrasion and more subjected to loss of particles, both of which contribute to accelerate mass loss. Breakdown could be primarily microbial and physical in streams where, shredders are not an important component of the system (Benfield et al. Citation1977). Furthermore, differences in decomposition rates from bags of coarse and fine mesh size were observed only in shaded sites.
Zoosporic fungi and straminipiles were well represented on the leaves from both invaded and non-invaded sites. These groups exhibited some differences in taxa composition but not in their abundance, richness, or diversity between sites. This suggests that differences in litter breakdown would not be related to the activity of zoosporic organisms. Instead, they could possibly be related to invertebrate activity and/or to the mechanical fragmentation of the leaves. The conditions inside the buried bags (particularly low oxygen availability) might have affected invertebrate activity.
The perturbation produced by the presence of the forest riparian vegetation increased the breakdown rates. We did not find any changes in the structure of the communities of zoosporic organisms that could be related to the differences observed in the decomposition rates. Hence, either the differences regarding the physical fragmentation of the leaves, the leaf burial, or a limited invertebrate activity may have played a role in the breakdown rates in non-invaded sites.
Acknowledgements
The authors thank Aníbal Sánchez Caro for constructive comments on earlier versions of the manuscript and Violet Corace for help with the English version of this article. They also thank Marcelo Sciorra and Jonatan Gómez for field assistance. This research was funded by Universidad Nacional de Luján.
References
- APHA . 1998 . Standard methods for the examination of water and wastewater , Washington, DC : American Public Health Association .
- Baldy , V , Gessner , MO and Chauvet , E . 1995 . Bacteria, fungi and the breakdown of leaf litter in a large river . Oikos , 74 : 93 – 102 .
- Barling , RD and Moore , ID . 1994 . Role of buffer strips in management of waterway pollution: a review . Environmental Management , 18 : 543 – 558 .
- Bärlocher , F . 1992 . “ Community organization ” . In The ecology of aquatic Hyphomycetes , Edited by: Bärlocher , F . 38 – 76 . Berlin (Germany) : Springer-Verlag .
- Bärlocher , F and Graça , MAS . 2002 . Exotic riparian vegetation lowers fungal diversity but not leaf decomposition in Portuguese streams . Freshwater Biology , 47 : 1123 – 1135 .
- Barr , DJS . 1987 . “ Isolation, culture and identification of chytridiales, Spizellomycetales and Hyphochytriales ” . In Zoosporic fungi in teaching and research , Edited by: Fuller , MS and Jaworski , A . 118 – 120 . Athens, GA : Southeastern Publishing Corporation .
- Benfield , EF , Jones , DS and Patterson , MF . 1977 . Leaf pack processing in a pastureland stream . Oikos , 29 : 99 – 103 .
- Boulton , AJ and Boon , PI . 1991 . A review of methodology used to measure leaf litter decomposition in lotic environments: time to turn over an old leaf? . Australian Journal of Marine and Freshwater Research , 42 : 1 – 43 .
- Bunn , SE . 1988 . Processing of leaf litter in two northern jarrah forest streams, Western Australia: II. The role of macroinvertebrates and the influence of soluble polyphenols and inorganic sediment . Hydrobiologia , 162 : 211 – 223 .
- Bunn , SE , Davies , PM and Mosisch , TD . 1999 . Ecosystem measures of river health and their response to riparian and catchment degradation . Freshwater Biology. , 41 : 333 – 345 .
- Cappello , V . de la Peña C. 2007. Propuesta de manejo de acacia negra (Gleditsia triacanthos) en la cuenca del río Luján. Dirección provincial de preservación de la biodiversidad. Secretaría de Política Ambiental. Gobierno de la Provincia de Buenos Aires
- Chaneton , EJ , Mazia , CN , Machera , M , Uchitel , A and Ghersa , CM . 2004 . Establishment of honey locust (Gleditsia triacanthos) in burned pampean grasslands . Weed Technology , 18 : 1325 – 1329 .
- Chauvet , E and Suberkropp , K . 1998 . Temperature and sporulation of aquatic hyphomycetes . Applied and Environmental Microbiology , 64 : 1522 – 1525 .
- Coker , WC . 1923 . The saprolegniaceae with notes on other water molds , Chapel Hill : University of North Carolina Press .
- Colwell , RK . 2005. Estimates: statitical estimations of species richness and shared species from samples. Version 7.5 Persistent URL. Available from: <purl.oclc.org/estimates>
- Cummins , KW , Minshall , GW , Sedell , JR , Cushing , CE and Petersen , RC . 1984 . Stream ecosystem theory . Verhandlung Internationale Vereinigung Limnologie , 22 : 1818 – 1827 .
- Cummins , KW , Wilzbach , MA , Gates , DM , Perry , JB and Taliaferro , WB . 1989 . Shredders and riparian vegetation . Bioscience , 39 : 24 – 30 .
- Dix , NJ and Webster , J . 1995 . Fungal ecology , Cambridge : Chapman and Hall .
- Elosegi , A , Butturini , A and Armengol , J . 2009 . “ El caudal circulante ” . In Conceptos y técnicas en ecología fluvial , Edited by: Elosegi , A and Sabater , S . 51 – 69 . Bilbao (España) : Fundación BBV .
- Feijoó , CS , Giorgi , ADN , García , ME and Momo , FR . 1999 . Temporal and spatial variability in streams of a pampean basin . Hydrobiologia , 394 : 41 – 52 .
- Gantes , P , Momo , FR , Sánchez Caro , A and Gómez , J . 2005 . Descomposición de macrófitas en el arroyo Las Flores (Buenos Aires) . Biología Acuática , 22 : 131 – 136 .
- Gantes , P and Sánchez Caro , A . 2001 . Environmental heterogeneity and spatial distribution of macrophytes in plain streams . Aquatic Botany , 70 : 225 – 236 .
- Gessner , MO and Chauvet , E . 2002 . A case for using litter breakdown to assess functional stream integrity . Ecological Applications , 12 : 498 – 510 .
- Gessner , MO , Gulis , V , Kuehn , KA , Chauvet , E and Subberkrop , K . 2007 . “ Fungal decomposers of plant litter in aquatic ecosystems ” . In The Mycota IV. Environmental and microbial relationships, , 2nd , Edited by: Kubicek , CP and Druzhinina , IS . 301 – 324 . Berlin (Germany) : Springer-Verlag .
- Ghersa , CM , de la Fuente , EB , Suarez , S and León , RJC . 2002 . Woody species invasion in the rolling pampa grasslands, Argentina . Agriculture, Ecosystems and Environment , 88 : 271 – 278 .
- Giorgi , A , Feijoó , C and Tell , HG . 2005 . Primary producers in a Pampean stream: temporal variation and structuring role . Biodiversity and Conservation , 14 : 1699 – 1718 .
- Graça , MAS , Ferreira , RCF and Coimbra , CN . 2001 . Litter processing along stream gradient: the role of invertebrates and decomposers . Journal of the North American Benthological Society , 20 : 408 – 420 .
- Gulis , V , Kuehn , KA and Suberkropp , K . 2009 . “ Fungi ” . In Encyclopedia of Inland Waters , Edited by: Likens , GE . Vol. 3 , 233 – 243 . Oxford : Elsevier .
- Hieber , MA and Gessner , MO . 2002 . Contribution of stream detrivores, fungi, and bacteria to leaf breakdown based on biomass estimates . Ecology , 83 : 1026 – 1038 .
- Hood , WG and Naiman , RJ . 2000 . Vulnerability of riparian zones to invasion by exotic vascular plants . Plant Ecology , 148 : 105 – 114 .
- Instituto Nacional del Agua. 2007. Diagnóstico del funcionamiento hidrológico-hidráulico de la cuenca del río Luján, Provincia de Buenos Aires, Argentina. Laboratorio de Hidráulica. Informe LHA-248-01-2007
- Kalesnik , F and Malvárez , AI . 2003 . Las especies invasoras exóticas en los sistemas de Humedales. El caso del Delta Inferior del Río Paraná. INSUGEO. CONICET-Universidad de Tucumán . Miscelanea , 12 : 5 – 12 .
- Karling , JS . 1977 . Chytridiomycetarum iconographia , Vaduz : Lubrecht & Cramer .
- Kuehn , KA and Suberkropp , K . 1998 . Decomposition of standing litter of the freshwater emergent macrophyte Juncus effusus . Freshwater Biology , 40 : 717 – 727 .
- Laitung , B and Chauvet , E . 2005 . Vegetation diversity increases species richness of leaf-decaying fungal communities in woodland streams . Archiv für Hydrobiologie , 164 : 217 – 235 .
- Lecerf , A , Dobson , M , Dang , CK and Chauvet , E . 2005 . Riparian plant species loss alters trophic dynamics in detritus-based stream ecosystems . Oecologia , 146 : 432 – 442 .
- Lecerf , A , Patfield , D , Boiché , A , Riipinen , MP , Chauvet , E and Dobson , M . 2007 . Stream ecosystems respond to riparian invasion by Japanese knotweed (Fallopia japonica) . Canadian Journal of Fisheries and Aquatic Sciences , 64 : 1273 – 1283 .
- Lee , AA and Bukaveckas , PA . 2002 . Surface water nutrient concentrations and litter decomposition rates in wetlands impacted by agriculture and mining activities . Aquatic Botany , 74 : 273 – 285 .
- Magurran , A . 2004 . Measuring biological diversity , Oxford : Blackwell Publishing .
- Marano , AV , Barrera , MD , Steciow , MM , Donadelli , JL and Saparrat , MCN . 2008 . Frequency, abundance and distribution of zoosporic organisms from Las Cañas stream (Buenos Aires, Argentina) . Mycologia , 100 : 691 – 700 .
- Marano , AV , Pires-Zottarelli , CLA , Barrera , MD , Steciow , MM and Gleason , FH . 2011 . Diversity, role in decomposition and succession of zoosporic fungi and straminipiles on submerged decaying leaves in a woodland stream . Hydrobiologia , 659 : 93 – 109 .
- McLatchey , GP and Reddy , KR . 1998 . Regulation of organic matter decomposition and nutrient release in a wetland soil . Journal of Environmental Quality , 27 : 1268 – 1274 .
- Micheli , ER and Kirchner , JW . 2002 . Effects of wet meadow riparian vegetation on streambank erosion. 1. remote sensing measurements of streambank migration and erodibility . Earth Surface Processes and Landforms , 27 : 627 – 639 .
- Nikolcheva , LG and Bärlocher , F . 2004 . Taxon-specific primers reveal unexpectedly high diversity during leaf decomposition in a stream . Mycological Progress , 3 : 41 – 49 .
- Olson , JS . 1963 . Energy storage and the balance of producers and decomposers in ecological systems . Ecology , 44 : 322 – 331 .
- Parnrong , S , Buapetch , K and Buathong , M . 2002 . Leaf decomposition rates in three tropical streams of southern Thailand: the influence of land use. Verhandlungen des Internationalen Verein . Limnologie , 28 : 475 – 479 .
- Pascoal , C , Cassio , F , Aranzazu , M , Sanz , B and Gomes , P . 2005 . Role of fungi, bacteria, and invertebrates in leaf litter breakdown in a polluted river . Journal of the North American Benthological Society , 24 : 784 – 797 .
- Rigacci , L . 2009. Invertebrados bentónicos en arroyos con distinta vegetación ribereña. Tesis de Licenciatura de la Universidad Nacional de Luján, Luján, Buenos Aires, Argentina
- Rocha , M and Pires-Zottarelli , CLA . 2002 . Chytridiomycota e Oomycota da Represa do Guarapiranga, São Paulo . Acta Botanica Brasilica , 16 : 287 – 309 .
- Sabater , F , Butturini , A , Marti , E , Munoz , I , Romani , A , Wray , J and Sabater , S . 2000 . Effects of riparian vegetation removal on nutrient retention in a Mediterranean stream . Journal of the North American Benthological Society , 19 : 609 – 620 .
- Schoenlein-Crusius , IH and Milanez , AI . 1989 . Sucessão fúngica em folhas de Ficus microcarpa L. f. submersas no lago frontal situado no Parque Estadual das Fontes do Ipiranga, São Paulo, SP . Reviews of Microbiology , 20 : 95 – 101 .
- Schoenlein-Crusius , IH and Milanez , AI . 1998 . Fungal succession on leaves of Alchornea triplinervia (Spreng.) Muell. Arg. submerged in a stream of an Atlantic rainforest in the state of São Paulo, Brazil . Revista Brasileira de Botânica , 21 : 253 – 259 .
- Schoenlein-Crusius , IH , Pires-Zottarelli , CLA and Milanez , AI . 1990 . Sucessão fúngica em folhas de Quercus robur L. (Carvalho) submersas em um lago situado no município de Itapecerica da Serra, SP . Revista de Microbiologia , 21 : 61 – 67 .
- Schoenlein-Crusius , IH , Pires-Zottarelli , CLA and Milanez , AI . 1992 . Aquatic fungi in leaves submerged in a stream in the Atlantic rainforest . Revista de Microbiologia , 23 : 167 – 171 .
- Schoenlein-Crusius , IH , Pires-Zottarelli , CLA , Milanez , AI and Humphreys , RD . 1999 . Interaction between the mineral content and the occurrence number of aquatic fungi in leaves submerged in a stream in the Atlantic rainforest, São Paulo, Brazil . Revista Brasileira de Botânica , 22 : 133 – 139 .
- Seena , S , Wynberg , N and Bärlocher , F . 2008 . Fungal diversity during decomposition of oak, maple and linden leaves assessed through clone libraries . Fungal Diversity , 30 : 1 – 14 .
- Sparrow , FK . 1960 . Aquatic phycomycetes , Ann Arbor : University of Michigan Press .
- Sponseller , RA and Benfield , EF . 2001 . Influences of land use on leaf breakdown in southern Appalachian headwater streams: a multiple-scale analysis . Journal of the North American Benthological Society , 20 : 44 – 59 .
- Stevens , RB . 1974 . Mycological guidebook , Seattle : University of Washington Press .
- Suberkropp , KF . 1992 . “ Aquatic hyphomycetes communities ” . In The fungal community: its organization and role in the ecosystem, , 2nd , Edited by: Caroll , CG and Wicklow , DT . 729 – 747 . New York : Marcel Dekker Inc .
- Sweeney , BW , Bott , TL , Jackson , JK , Kaplan , LA , Newbold , JD , Standley , LJ , Hession , WC and Horwitz , RJ . 2004 . Riparian deforestation, stream narrowing, and loss of stream ecosystem services . Proceedings of the National Academy Sciences of the United States of America , 101 : 14132 – 14137 .
- Triska , FJ . 1970. Seasonal distribution of aquatic hyphomycetes in relation to the disappearance of leaf litter from a woodland stream [PhD thesis]. [Pittsburgh (PA)]: University of Pittsburgh
- Vanotte , RL , Minshall , GW , Cummins , KW , Sedell , JR and Cushing , CE . 1980 . The river continuum concept . Canadian Journal of Fisheries and Aquatic Sciences , 37 : 130 – 137 .
- Viglizzo , EF , Frank , FC and Carreño , LV . 2006 . “ La situación ambiental en las ecorregiones Pampa y Campos y Malezales ” . In La situación ambiental argentina 2005 , Edited by: Brown , A , Martinez Ortiz , U , Acerbi , M and Corcuera , J . 263 – 278 . Buenos Aires (Argentina) : Fundación Vida Silvestre Argentina .
- Vilches , CS . 2005. Comparación del metabolismo en productores del arroyo Las Flores. Tesis de Licenciatura de la Universidad Nacional de Luján, Luján, Buenos Aires, Argentina
- Vitousek , PM . 1990 . Biological invasions and ecosystem processes: towards an integration of population biology and ecosystem studies . Oikos , 57 : 7 – 13 .
- Webster , JR and Benfield , EF . 1986 . Vascular plant breakdown in freshwater ecosystems . Annual Review of Ecology and Systematics , 17 : 567 – 594 .
- Zar , JH . 1999 . Biostatistical analysis , 663 New Jersey : Prentice Hall .