Abstract
South Dakota (SD) represents the northwestern edge of gizzard shad's (Dorosoma cepedianum) native range. However, with recent climate warming the gizzard shad is experiencing a natural range expansion northward. Little knowledge exists regarding temperatures that are required for overwinter survival, and less is known regarding population dynamics of gizzard shad in these populations. The objectives of this research were to determine the extent of reproduction, recruitment, and overwinter survival after annual introductions of adult gizzard shad in two northeastern SD glacial lakes relative to other SD systems. Prior to spawning, the gizzard shad was introduced into East Krause Lake and Middle Lynn Lake and successfully spawned during 2008 and 2009. Mean (±SE) peak larval abundances (number/100 m3) were 3.87 (±1.87) and 1.72 (±0.89) in East Krause Lake and 0.79 (±0.22) and 0.17 (±0.11) in Middle Lynn Lake in 2008 and 2009, respectively. Larval abundances were lower than those observed in most other SD populations. Survival to fall age-0 was documented in both years in Middle Lynn Lake, but only during 2008 in East Krause Lake. No overwinter survival was documented. Low water temperatures and rapidly cooling water likely caused overwinter mortality of gizzard shad in these systems. Temperature at the sediment–water interface during the winter of 2009–2010 was below 2.2°C for 114 and 62 consecutive days in East Krause Lake and Middle Lynn Lake, respectively.
Introduction
The gizzard shad (Dorosoma cepedianum) is a highly fecund, warmwater species found throughout the southern USA (Noble Citation1981). Its native range encompasses most of the eastern continental USA and extends from Florida to New Mexico in the south and as far north as North Dakota in the west and New York in the east (Heidinger Citation1983; Pflieger Citation1997). In southern reservoirs, the gizzard shad often grows rapidly, reaches high densities and tends to produce fewer offspring which limits their utility as a prey species (Swingle Citation1950; Smith Citation1959; Noble Citation1981; Michaletz Citation1998). However, in the northern portion of its range, high winter mortality leads to lower overall adult population abundance (Porath Citation2006). As a result, populations in these locations tend to produce more consistent numbers of age-0 gizzard shad, which then are available as a forage for predators (Willis Citation1987; Wuellner et al. Citation2008). To capitalize on the prey value of gizzard shad, fisheries managers in the Great Plains region collect adult brood stock and transfer them to receiving waters in the spring prior to spawning (Eichner and Ellison Citation1983; Porath Citation2006). For example, growth and abundance of walleye (Sander vitreus) in one western South Dakota (SD) reservoir increased following the introduction and establishment of gizzard shad as an alternative prey source (Ward et al. Citation2007).
SD lies near the northwestern edge of the native range of gizzard shad (Pflieger Citation1997; Wuellner et al. Citation2008). At the northern edge of their range, gizzard shad populations often experience excessive winter mortality which can lead to insufficient broodstock densities or even extirpation (Eichner and Ellison Citation1983; Porath Citation2006). The gizzard shad was historically found in the Missouri River and its tributaries as far north as North Dakota; however, since the damming of the Missouri River and subsequent changes in riverine habitat, the range became more restricted on the northwestern edge. In recent years gizzard shad populations in SD reservoirs and glacial lakes have exhibited overwinter survival, and self-sustaining populations have subsequently perpetuated (Hanten Citation2006; Miller et al. 2007; SDGFP Citation2009a). Gizzard shad arrived in these systems via natural range expansion [flooding (SDGFP Citation2009a) and downstream migrations (Hanten Citation2006; Miller et al. 2007)], were identified during annual surveys, and successfully overwintered during mild winters. However, little knowledge exists regarding water temperatures required for over winter survival and even less is known regarding population dynamics of gizzard shad in these populations. To better understand the potential for gizzard shad overwinter survival at the northern edge of its range, adult gizzard shad were stocked into two glacial lakes. Gizzard shad populations in these lakes were then monitored over two subsequent years. The objectives of this research were to determine the extent of reproduction, recruitment, and overwinter survival after annual introductions of adult gizzard shad in two northeastern SD glacial lakes relative to other SD systems.
Methods and materials
Study area
Our research was conducted on Middle Lynn Lake and East Krause Lake in the prairie pothole region (Day County, SD; ). Both lakes are located within the James River drainage, where the gizzard shad is native and has been documented as far north as North Dakota. Middle Lynn Lake has a surface area of 280 ha, and East Krause Lake has a surface area of 70 ha. Both are natural lakes of glacial origin, are polymictic, have a maximum depth of approximately 6 m, are closed basins, and often reach water temperatures of ≥28°C. The predominant land-use in each watershed is agricultural, and both are considered mesotrophic to eutrophic based on the trophic state index [TSI (Carlson Citation1977); TSIsecchi = 54 and 55, respectively]. The fish assemblage of both lakes consists primarily of walleye, yellow perch (Perca flavescens), fathead minnow (Pimephales promelas), and low densities of bluegill (Lepomis macrochirus), black bullhead (Ameiurus melas), and johnny darter (Etheostoma nigrum). Additionally, East Krause has a low-density black crappie (Pomoxis nigromaculatus) population.
Figure 1. Water bodies assessed for gizzard shad survival and reproduction. Shading represents mean annual temperature (°C) from 1971 to 2000 (US Department of Agriculture 2010).
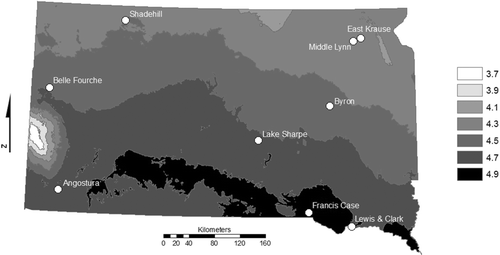
Gizzard shad dynamics were also assessed in several other systems (; ). Lake Byron is a glacial lake that has a connection to the James River during high-water years. In August of 2007 during a standardized gill net and trap net survey, South Dakota Department of Game, Fish and Parks (SDGFP) personnel collected naturally occurring adult and juvenile (age-0) gizzard shad from the lake (SDGFP Citation2009a). To assess if any gizzard shad had survived over winter and successfully spawned the following year, Lake Byron was trawled during the summer of 2008. During 2008 Lake Byron was not connected to the James River; therefore, any larval gizzard shad collected in 2008 would be the progeny of adults that survived over winter. Additional systems from which larval densities were compared included three irrigation reservoirs in western SD, and three mainstem Missouri River reservoirs in SD. These systems spanned more than 450 km from east to west and represented systems with a variety of ecological and climatological conditions (US Environmental Protection Agency Citation2010).
Table 1. Characteristics for all South Dakota waterbodies from which gizzard shad reproduction was compared.
Gizzard shad collection and stocking
During early May of 2008 and 2009, pre-spawn adult gizzard shad were collected from Lake Sharpe, SD, a mainstem Missouri River reservoir, and transported to the study lakes. In 2009, all fish received a pelvic fin clip; those stocked in 2008 were unclipped. In 2008, 167 adult gizzard shad ranging from 370 to 475 mm total length (TL) were collected and stocked into East Krause Lake (50 fish, 0.71 fish/ha) and Middle Lynn Lake (117 fish, 0.42 fish/ha). In 2009, a total of 175 fish were collected and stocked into East Krause Lake (50 fish, 0.71 fish/ha) and Middle Lynn Lake (125 fish, 0.45 fish/ha). Stocking densities were selected to be higher than those that resulted in successful establishment of gizzard shad populations in other SD waterbodies (see Wuellner et al. Citation2008 for details).
Assessment of reproduction and recruitment
To assess spawning success (reproduction), the abundances of larval gizzard shad (≤30 mm TL) were estimated in East Krause Lake and Middle Lynn Lake. In each lake, six sites were selected using a stratified random approach to encompass both nearshore and offshore habitats within each lake. These six sites were trawled at the water surface using a 1-m diameter ichthyoplankton trawl with 1000-µm mesh. Trawling occurred every 7–12 days starting in mid-June and continuing until at least mid-July resulting in a total of seven, five, and four trawl dates in 2008, 2009, and 2010, respectively. Trawls lasted for 5 min, and water volume sampled was determined using a flowmeter (Model 20762; General Oceanics Inc., Miami, Florida) to estimate larval density (larvae/100 m3). All trawl samples were fixed using a 95% ethanol, and gizzard shad were enumerated and measured (TL).
Survival and recruitment of larval gizzard shad to fall age-0 were estimated from fish collected by nighttime, pulsed-DC boat electrofishing in September 2008, 2009, and 2010. Catch rates (fish/h) of age-0 gizzard shad were determined from four randomly selected 10-min nearshore transects within each lake. These same transects were used in all 3 years (2008–2010). Additionally, as the gizzard shad is known to have patchy distributions (Downey and Toetz Citation1983; Johnson et al. Citation1988), each lake was electrofished for an additional 40 min in both littoral and pelagic habitats (depths of 0.3 to 4.0 m). To obtain a more accurate depiction of length distributions for fall age-0 gizzard shad, the standardized electrofishing samples were supplemented using graded mesh gill nets (n = 3/lake). Length-frequency histograms were then developed for all age-0 gizzard shad collected in both lakes and all years during which age-0 shad were collected.
Overwinter survival
To determine whether any adult or juvenile gizzard shad survived winter, gill nets were deployed in May, July, and September, and both daytime and nighttime electrofishing were conducted during July and September of 2009 and September of 2010. In July, both nearshore and offshore areas were electrofished for 40 min in both lakes. September sampling occurred at the four standardized transects plus an additional 40 min of electrofishing. Due to the low overall adult densities, trawling for larval shad, the progeny of any potential overwintering adults occurred in the spring and summer of 2010. If any larval gizzard shad were collected during 2010, this would indicate that adults had survived over winter and subsequently reproduced. All gizzard shad captured in 2009 were checked for pelvic fin clips, and saggital otoliths were removed to determine fish age. Otoliths were cracked through the nucleus and aged double-blind by experienced readers. Any gizzard shad captured in 2009 that did not have a pelvic fin clip and greater than 1 year of age was a fish from the 2008 stocking that survived one winter. Additionally, any fish determined to be 1 year of age in either 2009 or 2010 or a fish determined to be 2 years of age in 2010 would have survived at least one winter period.
Among water body comparisons
Peak mean larval abundance estimates (number/100 m3) were obtained from the SDGFP larval trawling database (unpublished data). Larval gizzard shad in these systems were sampled during annual standardized monitoring. Sampling occurred every 10–14 days from early May to mid-August (four to eight sampling events per lake) using a 1 m-diamater ichthyoplankton surface trawl with 500–1000 µm mesh. A flowmeter was mounted in the mouth of the trawl to estimate the volume of water filtered. Previous research found no significant differences in catch rates or peak larval abundance estimates between 500 and 1000 µm mesh ichthyoplankton trawls making these data directly comparable to those from Middle Lynn Lake, East Krause Lake, and Lake Byron (Isermann et al. Citation2002). The systems selected for comparison represented a wide range of waterbody sizes, trophic states, and watersheds and had climatological differences (; ). Mean annual air temperatures varied by 1°C across the systems, resulting in differences of 500–1000 growing degree-days (National Climatic Data Center). The reproduction obtained from stocked adults was assessed by comparing peak mean larval abundance from both study lakes with those from other waterbodies in SD, including naturally recruiting populations, supplementally stocked populations, and a system with an adult population known to have overwintered after a natural range expansion (Lake Byron).
Temperature effects
Because the gizzard shad is thermally limited in the northern portion of its range (Eichner and Ellison Citation1983; Porath Citation2006), water temperature data were collected at 3-h intervals during the winter of 2009–2010 in both lakes using a Hobo data logger (Onset Computer Corporation, Bourne, Massachusetts). Temperature was recorded from the sediment–water interface at a depth of approximately 4 m to prevent ice from destroying the loggers. As water temperature data were not available from the 2008–2009 winter and air temperature and water temperature are often correlated (McCombie Citation1959; Kothandaraman Citation1972; Livingstone and Lotter Citation1998), we tested whether we could use air temperature data as a surrogate for water temperature by performing correlation analysis on mean weekly water temperatures and air temperatures during the 2009–2010 winter. Additionally, to further assess the potential for overwinter survival and range expansion in the northeastern glacial lakes, mean weekly air temperature data (November–April) during the 2007–2008 winter were obtained for Lake Byron, where overwinter survival was documented, and compared to air temperatures at Middle Lynn Lake and East Krause Lake during both winters of the study (Weather Underground).
Results
Reproduction and recruitment
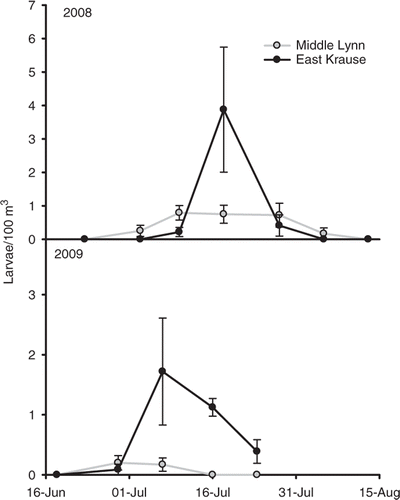
Gizzard shad successfully spawned in both lakes in 2008 and 2009. Mean (±SE) peak larval abundances (number/100 m3) were 3.87 (±1.87) and 1.72 (±0.89) in East Krause Lake and 0.79 (±0.22) and 0.17 (±0.11) in Middle Lynn Lake, respectively (). No larval gizzard shad were collected from either lake during trawling in 2010. Mean TLs of larval gizzard shad across sampling dates ranged from 13.0 to 18.9 mm in 2008 and from 13.5 to 18.0 mm in 2009 for both lakes.
Figure 2. Larval gizzard shad density estimates (number/100 m3 ± SE) for East Krause Lake and Middle Lynn Lake.
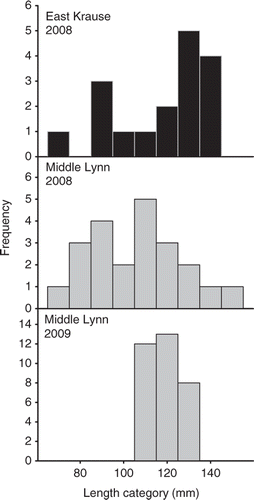
Mean (±SE) catch rates from nighttime electrofishing for autumn age-0 gizzard shad were 11 fish/h (±6) in East Krause Lake and eight fish/h (±8) in Middle Lynn Lake in 2008. TL of age-0 gizzard shad ranged from 77 to 144 mm in East Krause Lake and from 73 to 150 mm in Middle Lynn Lake in 2008 (). In 2009, fall age-0 gizzard shad were only documented in Middle Lynn Lake with a mean catch per unit effort (CPUE) of five fish/h (±5); the length-frequency distribution of fall age-0 gizzard shad in Middle Lynn Lake was more truncated ranging from 113 to 136 mm ().
Figure 3. Length-frequency histograms (10-mm length groups) for age-0 gizzard shad collected from East Krause and Middle Lynn lakes.
Overwinter survival
Sampling with gill nets and electrofishing collected only four adult gizzard shad in Middle Lynn Lake and one in East Krause Lake. All adult gizzard shad collected in Middle Lynn Lake were 5–9 years old. The one adult gizzard shad collected in East Krause Lake was 11 years old. All adult gizzard shad collected in 2009 were fin-clipped indicating that they were stocked in 2009. No adult gizzard shad were captured during 2010. Additionally, we did not capture any larval gizzard shad in 2010, and no age-1 shad were captured in either lake in 2009 or 2010. It is possible that gizzard shad were present in 2010 but not captured due to low densities and patchy distribution (Downey and Toetz Citation1983; Johnson et al. Citation1988). However, failure during the 2009 sampling season to capture any adult or juvenile gizzard shad that were not stocked that year and the failure to capture any larval, juvenile, or adult shad in 2010 most likely suggests that no overwinter survival occurred.
Among water body comparison
Peak larval abundances in Middle Lynn Lake and East Krause Lake in 2008 and 2009 were lower than other SD waters with gizzard shad populations except Lake Byron in 2008 (). In 2009, peak larval abundances were lower in all lakes than in 2008, with Middle Lynn Lake having the lowest larval density (0.20/100 m3) and Lake Sharpe having the highest (108.53/100 m3).
Table 2. Mean peak larval density (number/100 m3) estimates for gizzard shad from various South Dakota water bodies during 2008 and 2009.
Autumn CPUE of age-0 gizzard shad was at least an order of magnitude lower during both 2008 and 2009 in both northeastern glacial lakes than in other SD systems during corresponding years where electrofishing was the standardized collection protocol (Angostura, Belle Fourche and Shadehill reservoirs; SDGFP Citation2009b, Citation2010). For example, during 2008, mean CPUEs of autumn age-0 gizzard shad were 633, 185 and 47 fish/hour in Angostura Reservoir, Belle Fourche Reservoir and Shadehill Reservoir, respectively, compared to 11 and 5 fish/hour in East Krause Lake and Middle Lynn Lake.
Temperature effects
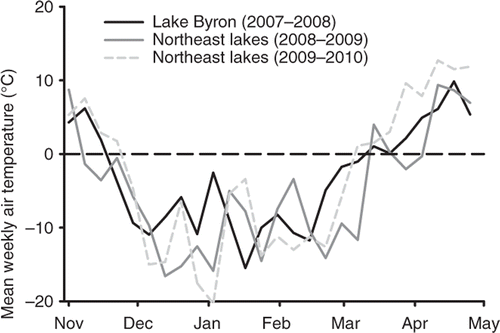
Water temperature at the sediment-water interface during the 2009–2010 winter was below 2.2°C for 114 and 62 consecutive days in East Krause Lake and Middle Lynn Lake, respectively (). Mean weekly air temperature was positively correlated with water temperature during the 2009–2010 winter in East Krause Lake (r = 0.80) and Middle Lynn Lake (r = 0.77). Mean weekly air temperature at Lake Byron ranged from −15.5°C to 9.8°C, with a mean of −3.2°C during the 2007–2008 winter. Mean weekly air temperature in the Middle Lynn Lake and East Krause Lake area ranged from −16.5°C to 9.4°C (mean = −5.0°C) during the 2008–2009 winter and −20.3°C to 12.7°C (mean = −3.0°C) during the 2009–2010 winter (Weather Underground; ).
Figure 4. Mean daily water temperature taken at the sediment-water interface at a depth of approximately 4 m in East Krause Lake (black line) and Middle Lynn Lake (grey line) during the 2009–2010 winter. The grey, dashed horizontal line represents the threshold at which mortality has been reported for gizzard shad.
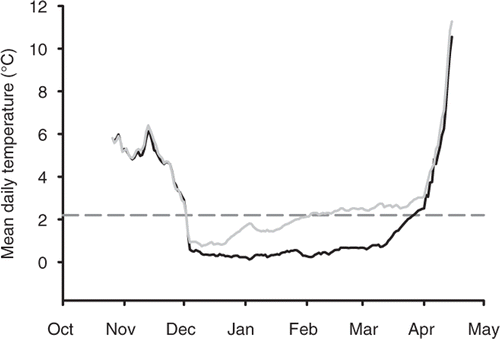
Figure 5. Mean weekly air temperatures (weather underground) during the 2007–2008 winter (November–April) near Lake Byron and the 2008–2009 and 2009–2010 winters near the northeastern South Dakota glacial lakes (Middle Lynn Lake and East Krause Lake).
Discussion
Gizzard shad successfully spawned in both lakes during both years, and the progeny subsequently recruited to fall age-0 in Middle Lynn Lake in 2008 and 2009 and in East Krause Lake in 2008. However, no overwinter survival was documented in either lake in either year, likely due to water temperature below the thermal tolerance of gizzard shad for a prolonged period of time. Wehr (Citation1976) suggested that gizzard shad mortality increases when temperature drops below 4°C. Research from Lake Rathbun, Iowa indicated that substantial mortality occurred when water temperature in nearshore areas was between 2.2°C and 3.3°C (Mayhew Citation1975). Further, gizzard shad experienced die-offs as far south as New Mexico when water temperature dropped below 3.3°C (Jester and Jensen Citation1972). Finally, Walburg (Citation1964) reported no survival of age-0 gizzard shad when ice cover exceeded 103 days, a duration exceeded during our study. Although we did not measure temperature at the deepest portion of the lake, likely where the water was warmest (4°C), the fact that water temperature fell below 2.2°C for at least 62 consecutive days in both our study lakes suggests that winter temperature precluded overwinter survival of gizzard shad.
Another factor potentially influencing overwinter survival is the rate at which water temperature declines at the onset of winter. Small, shallow lakes in the Great Plains region are susceptible to harsh winter conditions, rapid changes in water temperature, and sustained periods of cold water potentially causing lethal or sub-lethal effects for gizzard shad. During laboratory experiments, Griffith (Citation1978) found that a high proportion of threadfin shad (Dorosoma petenense) lost equilibrium in treatments where water temperature was decreased more rapidly. Rapid decreases in water temperature have been suggested to cause mortality in gizzard shad (Agersborg Citation1930; Miller Citation1960) and have also been shown to decrease swimming ability in clupeids making them more vulnerable to predation (Colby Citation1973; Griffith and Tomljanovich Citation1976; Griffith Citation1978) and decreasing their ability to osmoregulate (Wehr Citation1976). While research has suggested that adult gizzard shad are likely less vulnerable to overwinter mortality than juveniles, mortality rates commonly differ among waterbodies (Porath Citation2006; Ward et al. Citation2006). For example, during the warmer winter of 2007–2008 (Weather Underground), gizzard shad successfully overwintered in Lake Byron and subsequently reproduced the following spring (SDGFP Citation2009a). Air temperature during this winter did not decline as fast or go as low as it did during winters in which gizzard shad winter mortality was recorded in the northeastern lakes. Unfortunately, water temperature data from Lake Byron were unavailable during the 2007–2008, but air temperature during both winters of this study were colder than the 10-year average for this geographic location (Weather Underground), which likely led to decreased water temperature and subsequent direct or indirect mortality of gizzard shad.
Acknowledgements
We thank Nick Peterson, Madeline Wedge, Jason Harris, Nate Stukel, Tom Bacula, Andrew Jansen, and Melissa Wuellner from South Dakota State University and Todd Kaufman, Kyle Potter, Kris Edwards, Bob Hanten, Bill Miller and Gene Galinat from the South Dakota Department of Game, Fish and Parks for assistance with sample collection, for aid in fish stockings, and for providing additional data. Mark Kaemingk provided valuable feedback to improve the manuscript, and Josh White provided assistance with the graphics. Funding for this project was provided by Federal Aid in Sport Fish Restoration Project F-15-R, Study 1509, administered through South Dakota Department of Game, Fish and Parks.
References
- Agersborg , HPK . 1930 . The influence of temperature on fish . Ecology , 11 : 136 – 144 .
- Carlson , RE . 1977 . A trophic state index for lakes . Limnology and Oceanography , 22 : 361 – 369 .
- Colby , PJ . 1973 . “ Responses of alewives, Alosa pseudoharengus, to environmental change ” . In Responses of fish to environmental change , Edited by: Chavin , W . 163 – 198 . Springfield , IL : Charles C. Thomas Publishers .
- Downey , P and Toetz , D . 1983 . Distribution of larval gizzard shad (Dorosoma cepedianum) in Lake Carl Blackwell, Oklahoma . American Midland Naturalist , 109 : 23 – 33 .
- Eichner D, Ellison DG. 1983. Lake McConaughy Fishery Investigations. Federal Aid in Sportfish Restoration, Project F-51-R-5, Study 6. Nebraska Game and Parks Commission. Lincoln, Nebraska
- Griffith , JS and Tomljanovich , DA . 1976 . Susceptibility of threadfin shad to impingement . Proceedings of the Annual Conference of Southeastern Association of Game and Fish Commissions , 29 : 223 – 234 .
- Griffith , JS . 1978 . Effects of low temperature on the survival and behavior of threadfin shad, Dorosoma petenense . Transactions of the American Fisheries Society , 107 : 63 – 70 .
- Hanten R. 2006. Seasonal food habits, condition, growth and distribution of Lake Oahe, South Dakota walleye during depressed prey fish conditions spring 2001 to spring 2002. Pierre (SD): South Dakota Department of Game, Fish and Parks. Fisheries Division Report 07-02
- Heidinger RC. 1983. Life history of gizzard shad and threadfin shad as it relates to the ecology of small lake fisheries. In Proceedings of small lakes management workshop ‘pros and cons of shad’ Iowa Conservation Commission and Sport Fishing Institute, 23 and 24 March, Des Moines (IA)
- Isermann , DA , Hanchin , PA and Willis , DW . 2002 . Comparison of two mesh sizes for collecting larval yellow perch in surface trawls . North American Journal of Fisheries Management , 22 : 585 – 589 .
- Jester DB, Jensen BL. 1972. Life history and ecology of the gizzard shad, Dorosoma cepedianum (LeSueur) with reference to Elephant Butte Lake. Las Cruces (NM): New Mexico State University. New Mexico Agricultural Experiment Station Research Report 218
- Johnson , BM , Stein , RA and Carline , RF . 1988 . Use of a quadrat rotenone technique and bioenergetics modeling to evaluate prey availability to stocked piscivores . Transactions of the American Fisheries Society , 117 : 127 – 141 .
- Kothandaraman , V . 1972 . Air–water temperature relationship in Illinois River . Water Resources Bulletin , 8 : 38 – 45 .
- Livingstone , DM and Lotter , AF . 1998 . The relationship between air and water temperatures in lakes of the Swiss Plateau: a case study with paleolimnological implications . Journal of Paleolimnology , 19 : 181 – 198 .
- Mayhew J. 1975. Abundance, distribution, mortality, and production of 0-age fish. Federal Aid in Fish Restoration, Project F-88-R-1. Des Moines (IO): Iowa Conservation Commission. Performance Report Jobs 1, 3
- McCombie , AM . 1959 . Some relations between air temperatures and surface water temperatures of lakes . Limnology and Oceanography , 4 : 252 – 258 .
- Michaletz PH. 1998. Effects on sport fish growth of spatial and temporal variation in age-O shad availability North American Journal of Fishers Management. 18:616–624
- Miller , RR . 1960 . Systematics and biology of the gizzard shad (Dorosoma cepedianum) and related fishes , 173 Washington , DC : United States Fish and Wildlife Service. Fishery Bulletin .
- Miller B, James D, Galinat G, Shearer J. 2007. Statewide fisheries surveys, 2006 surveys of public waters Part 1 Lakes Region I. Pierre, SD: South Dakota Game, Fish and Parks. Fisheries Division Report No. 07–11
- National Climatic Data Center 2010. Available at http://climate.sdstate.edu/normals/annual-gdd/index.asp
- Noble , RL . 1981 . Management of forage fishes in impoundments of the southern United States . Transactions of the American Fisheries Society , 110 : 738 – 750 .
- Pflieger , WL . 1997 . The fishes of Missouri , Jefferson City : Missouri Department of Conservation .
- Porath , MT . 2006 . Climate and habitat factors related to a localized extirpation of gizzard shad (Dorosoma cepedianum) . Great Plains Research , 16 : 127 – 135 .
- SDGFP 2009a. Lake Byron. Pierre, Statewide Fisheries Survey No. 2102 F21-R-42
- SDGFP 2009b. Survey of public waters, Part 1 lakes region 1. Pierre, Annual Report 09–10
- SDGFP 2010. Survey of public waters, Part 1 lakes region 1. Pierre, Annual Report 10–12
- Smith Jr WA. 1959. Shad management in reservoirs. Proceedings of the Annual Conference of the Southeastern Association of Game and Fish Commissions 12:143–147
- Swingle , HS . 1950 . Relationships and dynamics of balanced and unbalanced fish populations. Auburn: Alabama Polytechnic Institute . Agricultural Experiment Station Bulletin , 274
- US Department of Agriculture, Natural Resources Conservation Service 2010. Processed annual average temperatures. Fort Worth (TX): National Cartography and Geospatial Center
- US Environmental Protection Agency 2010. Western Ecology Division. pp. 1. Available at ftp://ftp.epa.gov/wed/ecoregions/us/Eco_Level_III_US_pg.pdf
- Walburg CH. 1964. Fish population studies, Lewis and Clark Lake, Missouri River, 1956 to 1962. Washington (DC): US Fish and Wildlife Services. Special Scientific Report-Fisheries No. 482
- Ward , MJ , Willis , DW and Galinat , GF . 2006 . Gizzard shad recruitment patterns in a Western South Dakota irrigation reservoir . Journal of Freshwater Ecology , 21 : 201 – 207 .
- Ward , MJ , Willis , DW , Miller , BH and Chipps , SR . 2007 . Walleye consumption and long-term population trends following gizzard shad introduction into a western South Dakota reservoir . Journal of Freshwater Ecology , 22 : 339 – 345 .
- Weather underground. Available at http://www.wunderground.com/history/ (various dates)
- Wehr LW. 1976. Osmotic variation of serum of gizzard shad Dorosoma cepedianum, in relation to seasonal temperature changes [doctoral dissertation]. [Carbondale (IL)]: Southern Illinois University
- Willis , DW . 1987 . Reproduction and recruitment of gizzard shad in Kansas reservoirs . North American Journal of Fisheries Management , 7 : 71 – 80 .
- Wuellner , MR , Graeb , BDS , Ward , MJ and Willis , DW . 2008 . “ Review of gizzard shad dynamics at the northwestern edge of its range ” . In Balancing fisheries management and water uses for impounded river systems , Edited by: Allen , MS , Sammons , S and Maceina , MJ . 637 – 653 . Bethesda , MD : American Fisheries Society .