Abstract
Otolith microstructure was analyzed to determine the early growth of Neosalanx taihuensis juveniles from Three-Gorges Reservoir (TGR) and Tian-e-zhou Oxbow (TEO, below the dam). The early growth of the N. taihuensis juveniles differed significantly between the two populations. Higher relative condition index and wider otolith increments of the fish in the TGR indicated the faster early growth and larger otolith nucleus radius indicated the larger size of larvae at hatch for the TGR population, which may result in higher early survival rate. We suggest that increased primary production and food availability following the impoundment of the TGR contributed importantly to the increased early growth and survival of the N. taihuensis population. Fishery management and conservation in the TGR should consider the potential impacts to other fish species and the ecosystem of the increasing abundance of N. taihuensis.
Introduction
Neosalanx taihuensis is a freshwater-resident icefish mainly inhabiting the middle and lower reaches of the Yangtze River, including its tributaries and affiliated lakes (Xie and Xie 1997). Because of its high commercial value, N. taihuensis has been introduced into lakes and reservoirs in over 20 provinces and has become the most commercially exploited fish in China (Hu et al. Citation1998). Its life span is reported to be 1 year (Gong et al. Citation2009a) and it has two spawning stocks; one spawning in autumn from late September through early November and the other in spring from early January through late April (Gong et al. Citation2009a). Similar to many other short-lived fish species, populations of N. taihuensis often show considerable fluctuation, rapidly expanding in some years and drastically declining in others (Huang and Chang Citation2001). Knowledge about the fundamental biology of N. taihuensis is essential both to understand the population dynamics and to effectively manage the fisheries for this species.
N. taihuensis was one of the most abundant fishes in Tian-e-zhou Oxbow (TEO), a side channel system on the Yangtze River about 200 km below the Three-Gorges Dam (TGD) (). Annual commercial catch of N. taihuensis in TEO peaked in 2003 at 50 tons, but abundance declined dramatically in 2004 and annual catch has been less than one ton since 2006 (unpublished data from the Tian-e-zhou Oxbow National Baiji Reserve). N. taihuensis was not a commercially targeted species in the Three-Gorges Reservoir (TGR) area of the Yangtze River before impoundment in 2003. Abundance of the species increased following impoundment and it has become a major commercial species in the TGR.
Figure 1. Sampling locations for Neosalanx taihuensis in Three-Gorges Reservoir (TGR) at the Xiangxi Bay and in Tian-e-zhou Oxbow (TEO), showing the position of the bay (•) and the oxbow (▴), a flood-regulating weir (□) and Three-Gorges Dam (+++) in the middle and lower reaches of the Yangtze River.
Reproductive investment and early recruitment are two critical factors affecting population dynamics (Wright and Trippel Citation2009; Parkos and Wahl Citation2010). Our previous studies found that fecundity and egg size of the spring spawning stock of N. taihuensis in the TGR was higher than in the TEO (Gong et al. Citation2009b). Growth and development during early life stages are of major importance for survival. In general, faster growing larvae have a higher probability of survival (Houde Citation1987; Meekan and Fortier Citation1996; Takahashi and Watanabe Citation2004; Jenkins and King Citation2006). Maternal investment is another important factor determining individual performance and early survival of fish (Einum and Fleming Citation1999; Keckeis et al. Citation2000; Vallin and Nissling Citation2000). A larva hatched from a larger egg is usually larger in size at hatch and has a higher survival probability than one from a smaller egg (McCormick Citation2003). Knowledge about the early growth and development of N. taihuensis is very limited.
Otoliths record life-history events of fishes during early life stages (Radhakrishnan Citation2009). Otolith growth increments have been shown to occur daily for many species (Xie and Watanable Citation2005, Citation2007; Perter and Elise Citation2008). The otolith growth is a function of somatic growth, so the size and growth of fishes can be back-calculated from otolith size and increment width (Campana Citation1990). In addition, the nucleus radius of otolith is related to egg size in some fish and reflects the status of maternal investment (Ceriola and Jackson Citation2010). In this study, early growth and development of N. taihuensis juveniles were investigated in the TGR and TEO. The results will provide essential information for understanding the mechanisms regulating fluctuations of N. taihuensis populations that are important for resource forecasts and successful management of this species.
Methods
Fish sampling
N. taihuensis juveniles were sampled from commercial catch by square lift net (17 m × 17 m, 2-mm mesh) in the TGR (N 30°57′–31°34′′, E 110°25′–111°06′). Fish were attracted to the net with light and were collected in the evening on 21 May 2009. Fish in the TEO (N 29°47′–51′, E 112°33–37′) were sampled using a seine (350-m length, 12-m height, and 2-mm mesh) on 23 May 2009. The seine was drawn parallel to the shore where water depth ranged up to 1.5 m. The specimens were put on ice immediately after sampling and brought to the laboratory for further analyses. A total of 137 juveniles were collected with 69 from TEO and 68 from TGD. Fish were individually measured for standard length (SL, to 0.1 mm), weighed (BW, to 0.01 g). SL–BW relationship of the juveniles was fitted to BW = 2.96 × 10−7 × SL3.68 (n = 137, r 2 = 0.97, p < 0.05). Standard body weight (SW) of each juvenile was then calculated by entering its SL into the SL–BW function and relative condition factor (Kn , Pope and Kruse Citation2007) of each juvenile was calculated by:
Otolith analysis
Both the right and left sagittal otoliths were extracted from each fish, cleaned, and dried. Usually the right otolith was used for analysis. The otolith was mounted on a glass slide using nail polish, ground with 1000–2000 grit wet sandpaper and then polished with 3 µm lapping film until the core and the increments were visible under a light microscope (Wu et al. Citation2011). Sagittal otoliths of N. taihuensis have a single primordium. There were about 40 to 60 increments discernible by microscope surrounding the otolith core. After that, increments could usually not be accurately counted (Wu et al. Citation2011). The increments were suggested to be deposited daily, similar to those of Protosalanx hyalocranins, a related Salangid species (Fu et al. Citation1997; Wu et al. Citation2011). The nuclear radius of each otolith was measured. Increments were measured for the first 40 increments out of the core along the longest axis in the anterior area. Measurements were made using an image analysis system (Ratoc System Engineering, Tokyo, Japan) with a direct data feed between the light microscope and the computer.
Data analysis
The Kn , nucleus radius and increment width were compared between the two populations using a t-test. The level of significance was at α = 0.05. Data are presented as means ± standard deviations (SD). The analyses were conducted using Statistica 6.0.
Results and discussion
The SL of the analyzed juveniles ranged from 27.6 to 48.6 mm. The Kn of the TGR juveniles was 101.5 ± 13.9, and was significantly higher than that of the TEO juveniles (96.9 ± 9.2). The nuclear radius of the TGR juveniles was 6.46 ± 0.57 µm and was significantly larger than that of the TEO juveniles (6.00 ± 0.57 µm; ).
Table 1. Relative condition factor (Kn ) and otoliths nuclear radius for Neosalanx taihuensis juveniles in Three-Gorges Reservoir (TGR) and Tian-e-zhou Oxbow (TEO) in 2008.
The increment width profiles in the sagittal otolith of N. taihuensis juveniles varied between the two populations (). The increment width of the TEO population tended to be narrower than those of the TGR population, and was significantly different from the 15th through the 37th increments (t-tests, p < 0.05; ).
Figure 2. Variations in increment width in sagittae otolith of N. taihuensis juveniles between the Three-Gorges Reservoir (TGR) population and the Tian-e-zhou Oxbow (TEO) population in 2008. (•), TGR and (▴), TEO. Error bars represent one SD from the mean.
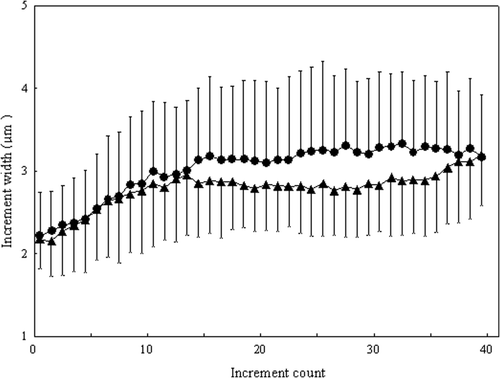
Results in this study revealed apparent variations in early growth and development of the N. taihuensis juveniles between the TGR and TEO populations. Higher relative condition factor and larger otolith increment width of the N. taihuensis juveniles in TGR indicated faster early growth than in TEO. As faster growing larvae often have a higher probability of survival (Oshima et al. Citation2010), early survival rate of the TGR population should be higher than that of the TEO population. Our research also demonstrated larger otolith nuclear radius of the N. taihuensis juveniles in TGR than in TEO. Thus, the size of the larvae at hatch should be larger and, consequently, the survival rate during the larval stage should be higher in TGR.
The survival rate during the early life stage is a critical factor in determining year class strength in fish population (Watanable et al. Citation1995; Poizat et al. Citation2002; Nakazawa et al. Citation2009). We suggest that this potential higher early survival rate due to the faster early growth and the larger size of larvae at hatch as demonstrated in this study contributed strongly to the higher and increasing abundance of the N. taihuensis in TGR. Our previous study demonstrated that fecundity and egg size were higher in the TGR population than the TEO population (Gong et al. Citation2009b). By monitoring early growth and development together with reproductive traits, future fluctuations of icefish population may be predicted.
Food availability is one of the primary determinants of early growth and survival of fish (McCormick Citation2003). Primary productivity in the TGR region apparently increased following its impoundment in 2003 due to increased nutrient loading and the changes from lentic to lotic habitat (Ye et al. Citation2007). At the same time, retention of sediment and nutrients in TGR lowers productivity in downstream areas (Li et al. Citation2004). Measured primary productivity in TGR in summer ranged from 4.65 to 6.95 g O2 m−2 d−1 (Ma et al. Citation2011), which was much higher than in TEO (0.37 to 2.67 g O2 m−2 d−1) (Shen Citation2007). N. taihuensis is primarily a zooplantivore (Ni and Zhu Citation2005; Qin et al. Citation2007). Increased food availability may be a major factor enhancing growth rate of N. taihuensis in TGR. Increased primary production following impoundment benefits fishery production typically during the early stages of reservoir life (Fernando et al. Citation1998). Together with the increase of N. taihuensis, production of the major carps (e.g., silver carp (Hypophthalmichthys molitrix) and bighead carp (Aristichthys nobilis)) has also increased in TGR since the impoundment (Wu et al. Citation2007). With the decline of primary productivity, downstream fishery harvests have declined dramatically since the impoundment of TGR (Xie et al. Citation2007). The annual commercial catch of N. taihuensis in TEO declined by 98% from 2003 to 2006 (Gong et al. Citation2009b). Growth and survival rates of fish larvae and juveniles downstream of TGR were demonstrated to be very low (Zhang et al. Citation2012). Declined primary productivity could be a major reason contributing to the lower early growth of N. taihuensis in the TEO. Further studies should be conducted investigating the impacts of impoundment of TGR on early growth and survival and, consequently, population dynamics of fishes below and above the dam. It is essential for fishery management and conservation to investigate the interaction of N. taihuensis with other species to evaluate its impacts on those species in TGR.
Acknowledgments
This research was financially supported by National Basic Research Program of China (973 Program, No. 2009CB119205), the National Natural Science Foundation of China (Nos 30771642 and 30972257), the One-hundred Talents Program of The Chinese Academy of Sciences issued to S. Xie, and the Network of Aquaculture Centres in Asia-Pacific (ICE/SL/FIS/2007/02-NACA).
References
- Campana , S E . 1990 . How reliable are growth back-calculations based on otoliths? . Canadian Journal of Fisheries and Aquatic Science , 47 : 2219 – 2227 . doi: 10.1139/f90-246
- Ceriola , L and Jackson , G D . 2010 . Growth, hatch size and maturation in a southern population of the loliginid squid Loliolus noctiluca . Journal of the Marine Biological Association of the United Kingdom , 90 : 755 – 767 . doi: 10.1017/S0025315409991445
- Einum , S and Fleming , I A . 1999 . Maternal effects of egg size in brown trout (Salmo trutta): norms of reaction to environmental quality . Proceedings of the Royal Society of London Series B-Biological Sciences. , 266 : 2095 – 2100 . doi: 10.1098/rspb.1999.0893
- Fernando , C H , Gurgel , J JS and Moyou , N AG . 1998. A global view of reservoir fisheries. International Review of Hydrobiology. 83 (special issue):31–42
- Fu , L J , Xie , Y H , Li , B and Zhao , H C . 1997 . Study on daily- growth increment of otolith and the growth of larvae icefish . Journal of Fishery Sciences of China , 4 : 21 – 27 .
- Gong , W B , Li , H T , Xie , S Q , Liu , J S , Murphy , B R and Xie , S G . 2009a . Two spawning stocks of icefish Neosalanx taihuensis revealed from annual reproductive cycle analyses . Fisheries Science , 75 : 1157 – 1165 . doi: 10.1007/s12562-009-0145-6
- Gong , W B , Wu , L , Liu , J S , Xie , S Q , Li , Z J , Murphy , B R and Xie , S G . 2009b . Variation in reproductive traits between two populations of Neosalanx taihuensis Chen above and below the Three Gorges Dam . Journal of Freshwater Ecology , 24 : 529 – 533 . doi: 10.1080/02705060.2009.9664329
- Houde , E D . 1987 . Fish early life dynamics and recruitment variability . American Fisheries Society Symposium , 2 : 17 – 29 .
- Hu , C L , Chen , W X and Liu , J S . 1998 . Status of transplantation and enhancement of icefishes in China and their development strategies . Reservoir Fisheries , 2 : 3 – 7 .
- Huang , Z L and Chang , J B . 2001 . Can icefish (Salangidae) production be predicted? . Acta Ecological Sinica , 21 : 86 – 93 .
- Jenkins , G P and King , D . 2006 . Variation in larval growth can predict the recruitment of a temperate, seagrass-associated fish . Oecologia , 147 : 641 – 649 . doi: 10.1007/s00442-005-0336-5
- Keckeis , H , Bauer-Nemeschkal , E , Menshutkin , V V , Nemeschkal , H L and Kamler , E . 2000 . Effects of female attributes and egg properties on offspring viability in a rheophilic cyprinid, Chondrostoma nasus . Canadian Journal of Fisheries and Aquatic Science , 57 : 789 – 796 . doi: 10.1139/f00-006
- Li , C X , Yang , S Y , Fan , D D and Zhao , J . 2004 . The change in Changjiang suspended load and its impact on the delta after completion of Three Gorges Dam . Quaternary Sciences , 24 : 495 – 500 .
- Ma , L N , Bi , Y H and Hu , Z Y . 2011 . Phytoplankton primary production during summer bloom in the Xiangxi bay, branch of the Yangtze River . Resources and Environment in the Yangtze Basin , 20 : 123 – 128 .
- McCormick , M I . 2003 . Consumption of coral propagules after mass spawning enhances larval quality of damselfish through maternal effects . Oecologia , 136 : 37 – 45 . doi: 10.1007/s00442-003-1247-y
- Meekan , M G and Fortier , L . 1996 . Selection for fast growth during the larval life of Atlantic cod Gadus morhua on the Scotian shelf . Marine Ecology-Progress Series , 137 : 25 – 37 . doi: 10.3354/meps137025
- Nakazawa , T , Ohgushi , T and Yamamura , N . 2009 . Resource-dependent reproductive adjustment and the stability of consumer-resource dynamics . Population Ecology , 51 : 105 – 113 . doi: 10.1007/s10144-008-0101-9
- Ni , Y and Zhu , C D . 2005 . Fishes of the Taihu Lake , 217 – 221 . Shanghai, China : Shanghai Scientific and Technical Publishers .
- Oshima , M , Robert , D , Kurita , Y , Yoneda , M , Tominaga , O , Tomiyama , T , Yamashita , Y and Uehara , S . 2010 . Do early growth dynamic explain recruitment success in Japanese flounder Paralichthys olivaceus off the Pacific coast of northern Japan? . Journal of Sea Research , 64 : 94 – 101 . doi: 10.1016/j.seares.2009.12.002
- Parkos , J J and Wahl , D H . 2010 . Intra- and intersystem variation in largemouth bass recruitment: Reproduction, prey availability, and the timing of year-class establishment . Transactions of the American Fisheries Society , 139 : 1372 – 1385 . doi: 10.1577/T09-097.1
- Perter , C S and Elise , R I . 2008 . Validation of daily ring deposition in the otoliths of age-0 channel catfish . North American Journal of Fisheries Management , 28 : 212 – 218 . doi: 10.1577/M06-176.1
- Pope , K L and Kruse , C G . 2007 . Condition , In : Guy CS, Brown ML. . editors. Analysis and interpretation of freshwater fisheries data. Bethesda (MD): American Fisheries Society. pp 423–471
- Poizat , G , Rosecchi , E and Crivelli , A J . 2002 . Life-history variation within a three-spined stickleback population in the Camargue . Journal of Fish Biology , 60 : 1296 – 1307 . doi: 10.1111/j.1095-8649.2002.tb01721.x
- Qin , J H , Xu , J and Xie , P . 2007 . Diet overlap between the endemic fish Anablariflus grahami (Cyprinidae) and the exotic noodlefish Neosalanx taihuensis (Salangidae) in lake Fuxian, China . Journal of Freshwater Ecology , 22 : 365 – 370 . doi: 10.1080/02705060.2007.9664165
- Radhakrishnan , K V , He , W P , Liu , M and Xie , S G . 2009 . Otoliths—the biological CD-ROMs of fish . Current Science , 8 : 1121 – 1122 .
- Shen , Z . W. 2007. Study on the primary productivity of the Tian'e-zhou and Hei-wa-wu Oxbows of the Yangtze River and its relationships with several ecological factors [Master Thesis] [Chongqing]: Southwest University. pp 20, 30–32
- Takahashi , M and Watanabe , Y . 2004 . Growth-rate dependent recruitment of Japanese anchovy Engraulis japonicus in the Kuroshio–Oyashio transitional waters . Marine Ecology-Progress Series , 266 : 227 – 238 . doi: 10.3354/meps266227
- Vallin , L and Nissling , A . 2000 . Maternal effects on egg size and egg buoyancy of Baltic cod, Gadus morhua implications for stock structure effects on recruitment . Fisheries Research , 49 : 21 – 37 . doi: 10.1016/S0165-7836(00)00194-6
- Watanabe , Y , Zenitani , H and Kimura , R . 1995 . Population decline of the Japanese sardine Sardinops melanostictus owing to recruitment failures . Canadian Journal of Fisheries and Aquatic Science , 52 : 1609 – 1616 . doi: 10.1139/f95-154
- Wright , P J and Trippel , E A . 2009 . Fishery-induced demographic changes in the timing of spawning: consequences for reproductive success . Fish and Fishery , 10 : 283 – 304 . doi: 10.1111/j.1467-2979.2008.00322.x
- Wu , L , Liu , J S , Wang , X L , Zhang , G , Zhang , Z Y , Murphy , B R and Xie , S G . 2011 . Identification of individuals born in different spawning seasons using otolith microstructure to reveal life history of Neosalanx taihuensis . Fisheries Science , 77 : 321 – 327 . doi: 10.1007/s12562-011-0333-z
- Wu , Q , Duan , X B , Xu , S Y , Xiong , C X and Chen , D Q . 2007 . Studies on fishery resources in the Three Gorges Reservoir of the Yangtze River . Freshwater Fisheries , 37 : 70 – 75 .
- Xie , S G , Li , Z J , Liu , J S , Xie , S Q , Wang , H Z and Murphy , B R . 2007 . Fisheries of the Yangtze River show immediate impacts of the Three Gorges Dam . Fisheries , 32 : 343 – 344 .
- Xie , S G and Watanabe , Y . 2005 . Hatch date-dependent differences in early growth and development recorded in the otolith microstructure of Trachurus japonicus . Journal of Fish Biology , 66 : 1720 – 173 . doi: 10.1111/j.0022-1112.2005.00764.x
- Xie , Y H and Xie , H . 1997 . Classification, distribution, and population ecology of Salangidae fishes . Chinese Journal of Fisheries , 2 : 11 – 19 .
- Xie , S G and Watanabe , Y . 2007 . Transport-determined early growth and development of jack mackerel Trachurus japonicus juvenile immigrating into Sagami Bay, Japan . Marine and Freshwater Research , 58 : 1048 – 1055 . doi: 10.1071/MF06165
- Ye , L , Han , X Q and Cai , Q H . 2007 . Spatial analysis for spring bloom and nutrient limitation in Xiangxi bay of Three Gorges Reservoir . Environmental Monitoring and Assessment , 127 : 135 – 145 . doi: 10.1007/s10661-006-9267-9
- Zhang , G , Wu , L , Li , H T , Liu , M , Cheng , F , Murphy , B R and Xie , S G . 2012 . Preliminary evidence of delayed spawning and suppressed larval growth and condition of the major carps in the Yangtze River below the Three Gorges Dam . Environmental Biology of Fishes , 93 : 439 – 447 . doi: 10.1007/s10641-011-9934-8