Abstract
We investigated the carbon and nitrogen stable isotope composition of particulate organic matter (POM) and zooplankton to understand the effects of highly variable hydrologic inputs with the onset of the summer monsoon season in Lake Soyang, Korea. The main characteristic separating pre-monsoon and post-monsoon seasons in Lake Soyang is the dramatic difference in hydrologic inflow. Pre-monsoon δ13C-POM (range −32.4 to −21.8‰) was lower than post-monsoon δ13C-POM (range −25.5 to −19.9‰). Epilimnetic δ13C-POM was higher post-monsoon compared to pre-monsoon, except in 2007. In the metalimnion, pre-monsoon δ13C-POM varied among years, while the variation in δ13C-POM post-monsoon was small and was related to the inflow of organic matter during summer monsoon storms. δ13C-zooplankton was lower post-monsoon (range −31.9 to −24.4‰) compared to pre-monsoon (range −19.4 to −17.7 ‰), while δ15N-zooplankton showed the opposite pattern (pre- and post-monsoon average 6.6 ± 2.0‰ and 4.3 ± 1.5‰, respectively). In Lake Soyang, the seasonal variation in δ13C-zooplankton was closely related to annual succession in the phytoplankton community and δ13C-phytoplankton. Thus, the results of this study provide some insights into the use of δ13C and δ15N measurements for understanding linkages between varying hydrologic characteristics and seasonal variation in the stable isotope composition of zooplankton. The dramatic seasonal change in hydrologic inputs in the monsoonal climate and consequent changes in turbid surface water inflow is linked with changes in the phytoplankton and zooplankton communities in Lake Soyang.
Introduction
The monsoon climate in East Asia is characterized by a rainy season and a much longer dry season. In Korea, this precipitation pattern has a major effect on water quality in reservoirs due to increased turbidity resulting from dramatic changes in flow during the monsoon season. In regions undergoing land use and demographic changes, there is a need for improving our understanding of the relationships between land use change, climate change, soil erosion and surface water quality.
The monsoon season in Korea (nominally July through September) is characterized by the inflow of turbid water resulting from the intense rainfall. The turbid storm runoff flowing into reservoirs inserts itself at metalimnetic depths because its density is greater than that of the surface water (Heo et al. Citation1998; Park et al. Citation2005). Turbid runoff to the metalimnion is the source of massive quantities of allochthonous organic matter (especially particulate organic matter (POM)) and nutrients to reservoirs during summer storms and the post-monsoon season (Heo et al. Citation1998; Choi et al. Citation2004; Park et al. Citation2011). Studies are needed to identify the sources and fates of POM, quantify organic matter and nutrient budgets and understand linkages to biological processes as well as the implications of the highly turbid water to trophic relationships in reservoirs. The largest reservoir in Korea, Lake Soyang, is the major source of water for the Seoul metropolitan area (with more than 24 million residents); therefore, protecting and enhancing water quality in Lake Soyang are critical needs for the future.
Namkung et al. (Citation2001) reported that allochthonous organic matter from the watershed accounted for about 43% of the total organic load to Lake Soyang. Nevertheless, the allochthonous contribution to the POM in the epilimnion would be expected to be small in Lake Soyang because most terrestrial organic matter is discharged from the dam outlet without affecting the epilimnion (Namkung et al. Citation2001; Choi et al. Citation2004). Nutrients in the turbid layer (metalimnion) are supplied to the epilimnion with deepening of the thermocline in late summer, which may stimulate cyanobacterial blooms (Heo et al. Citation1998).
Carbon and nitrogen stable isotopes can be used to identify the origin of organic matter and energy sources in food webs (Minagawa and Wada Citation1984; Vander Zanden and Rasmussen Citation2001). Additionally, stable isotopes can be used as a tool to study the relationships between surface waters and the surrounding watershed. Recent studies have used δ15N measurements in lakes to link increases in nitrogen loading from anthropogenic sources to eutrophication (Anderson and Cabana Citation2006; Risk et al. Citation2009).
Del Giorgio and France (Citation1996) suggested that the difference in carbon isotope signatures between zooplankton and POM reflected the relative contribution of allochthonous and autochthonous organic matter to bulk POM and varied with the trophic state. In lakes, seasonal variation in δ13C-zooplankton can be related to seasonal succession of phytoplankton and is directly related to δ13C-phytoplankton (Zohary et al. Citation1994; Gu et al. Citation1999; Syväranta et al. Citation2006). Grey et al. (Citation2000) reported few differences in carbon isotope composition between POM and phytoplankton in lakes with higher trophic states, showing that most of the POM consisted of phytoplankton in eutrophic lakes.
In this study, we aimed to study seasonal variation in POM and zooplankton in Lake Soyang using carbon and nitrogen stable isotopes. We focused on the linkages between hydrological characteristics (i.e., the seasonal influx of highly turbid runoff) and the stable isotope composition of POM and zooplankton in Lake Soyang. Based on previous studies, we hypothesized that autochthonous organic matter in the epilimnion would be expected to contribute a significant fraction of the total pool of carbon available to consumers such as herbivorous zooplankton in Lake Soyang (Kim et al. Citation1999; Namkung et al. Citation2001). Our study approach was to investigate the seasonal differences in carbon and nitrogen stable isotope composition of POM and zooplankton in order to compare patterns in stable isotopes between pre- and post-monsoon and to assess the relationship to varying hydrologic conditions.
Methods
Study site
Lake Soyang is the deepest and largest reservoir in Korea (37°56′4 8.45″N, 127°49′13.01″E, elevation 203 m). The reservoir was constructed on the North Han River in 1973 for the purpose of providing flood control and electricity. The reservoir has one main inflow, which contributes 90% of the inflow water volume. Lake Soyang is oligo-mesotrophic, dendritic and warm monomictic; water column mixing occurs in this lake throughout the winter. The Lake Soyang watershed area is about 2700 km2 and land use in the basin is approximately 87% forest, 6% agricultural and 7% urban. There is little influence from industrial or sewage wastes in the watershed. The hydraulic residence time averages 0.7 years. Annual precipitation in the region is about 1100 mm yr−1. More than half of total precipitation occurs during the summer monsoon season (July to September; , ). The Lake Soyang watershed is primarily undeveloped, but concerns about declining water quality and soil erosion resulting from expanding agricultural land use are intensifying.
Figure 1. Daily precipitation (mm), discharge (m3 s−1) and water level (m) of Lake Soyang from January 2007 to December 2009. Source: Korea Water Resources Corporation (KOWACO) and the Chuncheon Meteorological Service.
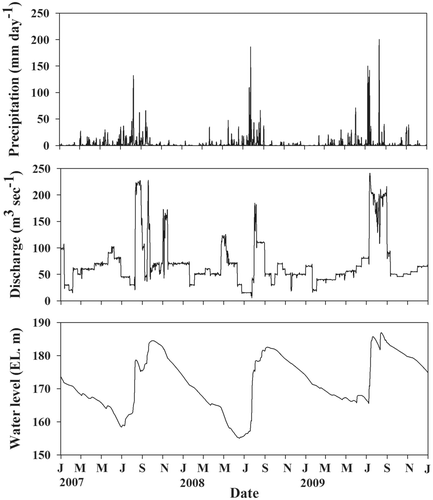
Table 1. Hydrological characteristics of Lake Soyang.
Biological and chemical analysis
Water quality parameters such as temperature (Cole Parmer Instrument Co., 93909-62), pH (Denver model 15), dissolved oxygen (YSI 55) and turbidity (HACH 2100P) were measured in situ. A sampling site located near the deepest area of the reservoir was used to characterize conditions in Lake Soyang. A long-term water quality monitoring installation is located at the sampling site. Samples for nutrient analysis were preserved by addition of 0.25 mL H2SO4(18 N) and kept frozen prior to analysis. Total phosphorus (TP) was measured according to American Public Health Association (APHA) (Citation1998) using the ascorbic acid method after sample digestion with K2S2O8. Total nitrogen (TN) was determined with an autoanalyzer (Skalar 5100; Skalar, Breda, The Netherlands) using the cadmium reduction method after digestion with K2S2O8. Chlorophyll-a concentrations were measured according to APHA (Citation1998).
Samples for zooplankton (63 µm mesh) were vertically hauled from 20 m depth. In the laboratory, zooplankton samples were filtered through a 200 µm mesh net. Samples were freeze dried and pulverized to a fine powder using a mill. To remove inorganic carbon, samples for carbon isotope analysis were fumigated in a desiccator with HCl (1 N) for 24 h (Carabel et al. Citation2006).
POM samples were collected at depths of 0, 2, 5, 10, 20, 40 and 70 or 80 m using a horizontal Van-Dorn sampler. Samples were pre-filtered through a 45 µm mesh net to remove zooplankton and these samples were filtered using precombusted (450°C) GF/F filters.
Phytoplankton samples for stable isotope analysis were collected by towing a weighted plankton net (35 µm mesh) to the surface from a depth of 20 m. Phytoplankton taxa were separated using gravitational separation in a 3 L cylinder. Buoyant cyanobacteria gathered in the upper layer, while other phytoplankton species were gravity settled. Floating cyanobacteria were obtained using a pipette.
Potential sources of terrestrial organic matter were collected in and around the inflowing stream (Inbuk stream). Periphyton samples were collected by scraping with a brush (area 25 cm2). Organic fragments such as invertebrates and particulate debris were separated using a net (45 µm mesh) in the laboratory. Periphyton samples were examined under a microscope in the laboratory to confirm that no debris or other organic matter was included in each sample. Samples were filtered using precombusted GF/F filters (450°C)
Representative surface soil samples were collected using a shovel in the area adjacent to an inflowing stream (Mandae stream) in the upper part of the watershed (38°17′ 17.14″N, 128°09′ 54.35″E) identified as having erodible soils (Kim et al. Citation2007). Leaf litter at this site was collected from 13 randomly selected points on the streambed. Samples of the most common riparian plant, Phragmites spp., were also collected by cutting the leaves from several reeds growing near the stream bank. After being transported to the laboratory, these samples were processed by drying (45°C) and pulverizing with a mill.
Isotope analysis
All samples for carbon stable isotope analysis were fumigated (HCl, 1 N) for 24 h and dried at 45°C. Samples for nitrogen stable isotope analysis were not acidified. Carbon and nitrogen stable isotope composition was measured using an isotope ratio mass spectrometer (EuroEA-IsoPrime). All stable isotope data are presented using the conventional per mil (‰) notation:
Statistical analyses
Comparisons between pre and post-monsoon were conducted using paired-sample t-tests with a significance level of 0.05 with SigmaPlot 11 software. In this study, we refer to post-monsoon as the time period for samples collected after the inflow of turbid water has started (August to October). Pre-monsoon includes the samples collected prior to the onset of the inflow of turbid water (May to July).
Results
Limnological characteristics of Lake Soyang
The water level and discharge from Lake Soyang varied considerably with changes in precipitation (). The water level during the pre-monsoon season (elevation 159 ± 5 m (average ± range)) was much lower compared to post-monsoon season (185 ± 2 m, ). Monitoring results for selected water quality parameters are shown in . Some changes between pre- and post-monsoon conditions in the water column were apparent for turbidity, epilimnetic pH and metalimnetic total phosphorus concentration ().
Figure 2. Water temperature (°C), turbidity (NTU), pH, dissolved oxygen concentration (DO; mg O2 L−1), total phosphorus (TP; µg P L−1), total nitrogen (TN; mg N L−1) and chlorophyll-a concentration (Chl a; µg L−1) in Lake Soyang. Upper panels show pre-monsoon (May to July) and lower panels show post-monsoon (August to October). Dashed line indicates the upper and lower position of the outflow (metalimnetic discharge).
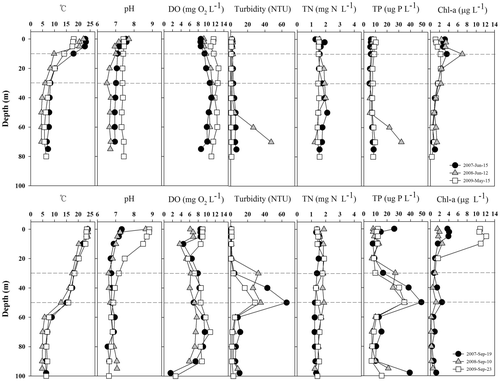
A thermocline gradually developed in Lake Soyang pre-monsoon and stratification intensified during the summer. Post-monsoon input of ambient temperature water from upstream resulted in the lowering of the thermocline by 50 m (). Pre-monsoon dissolved oxygen (DO) concentrations ranged from 7.6 to 11.2 mg O2 L−1 and post-monsoon DO concentrations ranged from 3.7 to 10.8 mg O2 L−1 (). Some post-monsoon decreases in DO concentrations were indicated at 10–20 m depth (in the epilimnion) and in the hypolimnion.
Table 2. Average ± standard deviation, minimum (min), and maximum (max) for selected water quality parameters in Lake Soyang.
In the epilimnion, the post-monsoon water column pH showed a wider range (6.6–8.8) than the pre-monsoon one (6.5–7.7, ). Pre- and post-monsoon turbidity ranged from 0.5 to 47.4 NTU and from 0.4 to 65.5 NTU, respectively. The maximum turbidity was observed only during post-monsoon when the density current developed ().
Pre-monsoon TP concentrations did not change markedly with depth and ranged from 5 to 32 µg P L−1 while post-monsoon TP concentrations ranged from 7 to 49 µg P L−1. The highest TP concentrations occurred in the metalimnion following summer storms every year (, ). TN concentrations varied only slightly with depth in both pre- and post-monsoon (, ). Pre-monsoon TN concentrations ranged from 1.3 to 2.1 mg N L−1. Post-monsoon TN concentrations ranged from 1.2 to 1.9 mg N L−1(, ).
Chlorophyll-a concentration during pre- and post-monsoon ranged from 0.5 to 7.0 µg L−1 and from 0.2 to 12.2 µg L−1, respectively (). The maximum chlorophyll-a concentration was near the depth of the thermocline (10 m) during pre-monsoon, while post-monsoon, the maximum chlorophyll-a concentration occurred near the surface ().
δ 13C-POM and other sources
δ13C-POM showed a considerable difference between pre- and post-monsoon (p < 0.001; ). For 2008 and 2009, post-monsoon δ13C-POM was approximately 6‰ higher than pre-monsoon δ13C-POM.
Figure 3. Vertical profile of δ13C-POM in Lake Soyang during pre-monsoon (May to July) and post-monsoon (August to October). Dashed line indicates the upper and lower position of the outflow.
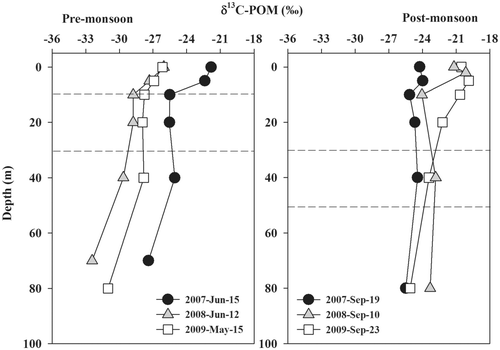
The pre-monsoon δ13C-POM decreased with depth but little variation with depth was indicated during the post-monsoon season. The average pre-monsoon δ13C-POM was −25.9 ± 2.3‰ in the epilimnion, −27.5 ± 1.8‰ in the metalimnion, and −30.3 ± 2.6‰ in the hypolimnion (average ± SD). Post-monsoon, the average δ13C-POM was −22.2 ± 2.1‰ in the epilimnion, −23.5 ± 1.0‰ in the metalimnion, and −24.2 ± 1.3‰ in the hypolimnion.
δ13C-phytoplankton ranged from −32.0 ± 2.5‰ pre-monsoon to −22.6 ± 1.7‰ post-monsoon (). δ13C-periphyton ranged from −24.8 to −17.3‰ (−21.7 ± 3.1‰) pre-monsoon, and from −25.0 to −22.8‰ (−24.1 ± 0.9‰) post-monsoon (). δ13C-soil collected adjacent to the inlet stream ranged from −23.9 to −22.2 (−23.4 ± 0.8‰). The average δ13C for stream bed leaf litter and riparian vegetation (Phragmites spp.) was −24.9 ± 1.5‰, −23.8 ± 0.5‰, respectively.
Figure 4. Seasonal variation in δ13C and δ15N of phytoplankton (inverted triangle) and zooplankton (circle) in Lake Soyang from January 2007 to December 2009. Solid arrow indicate the spring clear water phase. The species name indicates the dominant phytoplankton species at the time of sampling. Sampling dates are shown.
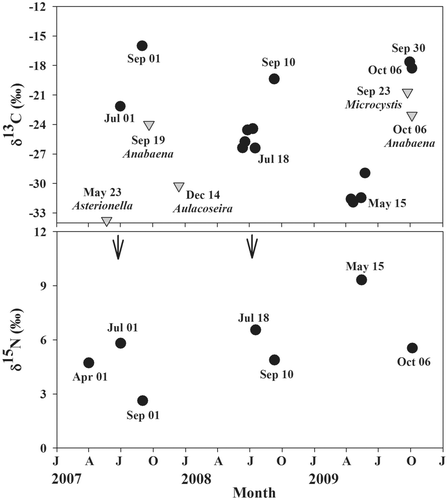
Table 3. δ13C of potential sources of organic matter to Lake Soyang (average ± SD).
Seasonal variation in δ 13C- and δ 15N-zooplankton
The zooplankton community in Lake Soyang was comprised mostly cladoceran species. Zooplankton δ13C and δ15N showed consistent seasonal changes between pre-monsoon and post-monsoon during the study (). Similar to the pattern for phytoplankton, δ13C-zooplankton was generally higher post-monsoon compared to pre-monsoon, while δ15N-zooplankton showed the opposite pattern, with lower δ15N in the pre-monsoon. For example, pre-monsoon δ13C- and δ15N-zooplankton was −26.9 ± 3.3‰ and 7.2 ± 1.8‰, respectively. Post-monsoon δ13C- and δ15N-zooplankton was −17.8 ± 1.4‰ and 4.3 ± 1.5‰, respectively ().
Discussion
Pre- and post-monsoon δ 13C-sources for zooplankton
We found a large difference in epilimnetic δ13C-POM between pre- and post-monsoon, which is likely due to a change in the availability of bicarbonate to the dominant phytoplankton (Microcystis spp.) relative to other phytoplankton taxa and variation in pH (Doi et al. Citation2006).
The variation in δ13C-POM, which is mostly phytoplankton, depends on the seasonal succession of phytoplankton (). This implies that δ13C-POM, when cyanobacteria are dominant, should be higher than the δ13C-POM of a diatom-dominated population due to a change in the availability of bicarbonate, as shown in other studies (Gu and Schelske Citation1996; Vuorio et al. Citation2006). Although we did not measure dissolved inorganic carbon (DIC) concentrations and δ13C-DIC, this pattern in POM should coincide with seasonal patterns in δ13C-phytoplankton in Lake Soyang. Most of the POM should be composed of phytoplankton in the euphotic zone, except during the mixing season (from December to March). In Lake Soyang, the pre-monsoon phytoplankton community was dominated by diatoms such as Asterionella formosa and Aulacoseira granulata, but during the post-monsoon period, a dense population of cyanobacteria such as Microcystis spp. and Anabaena spp. had developed (Heo and Kim Citation1997; Kim, personal observation). Thus seasonal differences in epilimnetic δ13C-POM may be influenced by seasonal variation in the phytoplankton community and differences in nutrient concentrations from the metalimnion between pre- and post-monsoon periods.
On the other hand, we found a different trend in δ13C-POM for 2007, which showed higher δ13C-POM relative to 2008 and 2009 for the post-monsoon period (). This may be due to reduced light penetration caused by high turbidity. With increasing turbidity in aquatic ecosystems, δ13C of primary producers can become lower due to the reduction in light penetration (Grice et al. Citation1996).
The post-monsoon pH was lower in 2007 compared to 2008 and 2009. Seasonal differences between δ13C-POM may be explained by the difference in pH, where a high rate of CO2 uptake by primary producers leads to increases in pH and a change in speciation from CO2(aq) to . The uptake of
by phytoplankton can result in higher δ13C-phytoplankton (and δ13C-POM) because δ13C-
is greater than δ13C-CO2(aq) (Yoshioka Citation1997; Vuorio et al. Citation2006; Rautio and Vincent Citation2007). Thus, variation in pH and turbidity, which are related to changes in hydrological characteristics (inflow of turbid runoff) are the most plausible explanations for our results showing lower δ13C-POM post-monsoon compared to pre-monsoon in 2007.
Changes in δ 13C-POM with depth
In Lake Soyang, pre-monsoon δ13C-POM decreased with depth but post-monsoon δ13C-POM showed less variation with depth (). This difference in the vertical profile of δ13C-POM is most likely related to different sources of organic carbon with varying hydrologic input between pre- and post-monsoon. We suspect that autochthonous organic matter (mostly phytoplankton) represents the major portion of POM in the metalimnion during the pre-monsoon period, while post-monsoon POM mostly comprises allochthonous organic matter due to terrestrial organic matter inputs through the metalimnion.
Gu et al. (Citation2006) and Gu et al. (Citation2011) showed that low primary production and dominance of non-living organic matter result in small seasonal changes in δ13C-POM. The isotope signature of non-living organic matter is not sensitive to the changes in physicochemical environments in the water column. Grey and Jones (Citation2001) showed that δ13C-POM, which was consistent with terrestrial plant origin from the catchment, exhibited little seasonal variation unlike δ13C-phytoplankton in oligotrophic lakes. The difference in δ13C-POM (−23.5 ± 1.0‰, average ± SD) in the post-monsoon metalimnion among the study years was small. δ13C-POM was similar to δ13C values for terrestrial leaf litter (−24.9 ± 1.5‰) and soil (−23.4 ± 0.8‰) in the watershed, probably resulting from the inflow of the turbid density current and the descending thermocline in Lake Soyang (Choi et al. Citation2004; ). Kim (Citation2007) also measured δ13C-organic matter in streams from forest and paddy fields located in the Han River watershed and found averages values of −25.1 ± 1.2‰ and −24.7 ± 0.8‰, respectively. These values are similar to the range of δ13C in this study for δ13C-POM in the metalimnion, supporting the view that δ13C-POM in the metalimnion is influenced by the input of terrestrial organic matter from the watershed.
In addition, periphyton brought to the reservoir from the inlet stream are sometimes observed in the metalimnion during the post-monsoon season due to removal of periphyton from upstream substrates (Lee et al. Citation2010; Kim, personal observation). We measured δ13C for several periphyton samples and found that δ13C-periphyton averaged −24.1 ± 0.9‰ (). This indicates that periphyton is also a possible source of terrestrial inputs during the monsoon season, though we expect that the magnitude of periphyton-derived C inputs would be lower compared to C inputs from suspended sediments due to the huge input of C in suspended particles during the monsoon.
The vertical distribution in δ13C-POM in Lake Soyang differs from the patterns in δ13C-POM with greater depth found in some other lakes. Studies of δ13C-POM in deep lakes have suggested that as POM settles through the water column and decomposes, 12C is preferentially released, and the residual POM becomes progressively 13C-enriched as it sinks (Ostrom et al. Citation1998). On the other hand, Del Giorgio and France (Citation1996) showed that respiration within the water column and sediments of lakes tends to produce CO2 (aq) which is depleted in 13C, and phytoplankton and POM at depths influenced by heterotrophic activity would be expected to have lower δ13C.
In this study, the descending thermocline during the post-monsoon period has an important role in sustaining higher epilimnetic δ13C-POM. The lowering of the thermocline into the dam's discharge layer results in increased concentrations of nutrients available for phytoplankton in the epilimnion (Heo et al. Citation1998). Although most of the allochthonous organic matter is discharged during the post-monsoon period (July to September), nutrients from the input of the sediment-laden density current can diffuse into the epilimnion (Heo et al. Citation1998, Choi et al. Citation2004). Therefore, chlorophyll-a concentrations may increase from this resupply of nutrients, as post-monsoon epilimnetic δ13C-POM was found to increase compared to pre-monsoon δ13C-POM. To better understand the mechanisms for the decrease in δ13C-POM with depth, more detailed studies on carbon cycling in Lake Soyang are needed.
Seasonal variation of δ 13C- and δ 15N-zooplankton
The inflow of large pulses of nutrients during the monsoon period is likely one of the important factors determining the stable isotope composition of zooplankton. In this study, δ13C-zooplankton differed between pre- and post-monsoon periods (). Other studies have suggested that seasonal variation in δ13C-zooplankton is dependent on seasonal shifts in the phytoplankton community and δ13C-phytoplankton (Zohary et al. Citation1994; Gu and Schelske Citation1996; Syväranta et al. Citation2006; Vuorio et al. Citation2006). Zooplankton directly suppresses phytoplankton abundance during the pre-monsoon season which results in the clear water phase in Lake Soyang (Kim et al. Citation1999).
The seasonal variation in δ13C-zooplankton in Lake Soyang shows a different pattern from the seasonal trends shown in some previous studies (Yoshioka et al. Citation1989; Bouillon et al. Citation2000; Perga and Gerdeaux Citation2006; Syväranta et al. Citation2006). Yoshioka et al. (Citation1989) showed that δ13C-zooplankton had higher values pre-monsoon compared to post-monsoon in Lake Kizaki, Japan. In Lake Jyväsjärvi, an urban lake in Finland, δ13C-zooplankton showed a clear seasonal trend of increase in δ13C in July and a decrease in August (Syväranta et al. Citation2006). Bouillon et al. (Citation2000) also showed that, in an estuarine system, δ13C-zooplankton was higher during pre-monsoon compared to post-monsoon, which indicates that zooplankton utilized prey of lower δ13C during post-monsoon compared to that during pre-monsoon. In Lake Soyang, post-monsoon δ13C-zooplankton was higher than pre-monsoon probably due to an increase in phytoplankton biomass and resupply of nutrients from the metalimnion (Perga and Gerdeaux Citation2006).
In our study, δ13C-zooplankton was similar to epilimnetic δ13C-POM when POM is presumably mainly composed of phytoplankton (). Even if zooplankton migrate into the metalimnion in search of prey (Del Giorgio and France Citation1996), zooplankton mainly exploit epilimnetic resources in Lake Soyang. Although highly detailed studies of δ13C-zooplankton, δ13C-POM and patterns in zooplankton migration in Lake Soyang are not available, our data suggest that δ13C-zooplankton does not reflect the type of prey items in the metalimnetic zone because zooplankton migrating into metalimnion would be lost with the discharge at the dam outlet. Feeding strategies of zooplankton can change when zooplankton are exposed to a high proportion of inedible POM (Greyand Jones Citation2001; Pulido-Villena et al. Citation2005). Microcystis spp. are unsuitable as a food source for zooplankton because Microcystis can inhibit not only feeding ability but also zooplankton growth (Chen and Xie Citation2003; Liu et al. Citation2006). Choi et al. (Citation1996) showed that bacterial growth was tightly coupled with phytoplankton growth during a bloom of Anabaena spp. in Lake Soyang. In Lake Soyang, Sim and Ahn (Citation1992) showed that macrozooplankton and some rotifers directly consume bacteria during the monsoon season in addition to phytoplankon. Hwang (Citation1997) suggested that zooplankton may preferentially use the heterotrophic pathway when phytoplankton populations are too large or include a high proportion of undesirable food source. In oligotrophic Loch Ness, Grey and Jones (Citation2001) used δ13C measurements to show that Eudiaptomus gracilis utilized allochthonous carbon even during the time of peak phytoplankton production.
Unlike patterns in δ13C in POM and zooplankton, no well-defined patterns were detected for δ15N-POM in some studies (Yoshioka et al. Citation1989; Grey and Jones Citation2001). In Lake Soyang, we found evidence of a consistent temporal pattern in δ15N-zooplankton during each year of our study. Pre-monsoon δ15N-zooplankton was always higher than post-monsoon δ15N-zooplankton (). In general, N-fixation will result in phytoplankton and POM having lower δ15N relative to anthropogenic inputs such as sewage, which has higher δ15N (Gu and Alexander Citation1993; Wang et al. Citation2005; Kaown et al. Citation2009). For example, Jones et al. (Citation2004) found that the primary producers had higher δ15N in nitrogen-limited lakes and in areas less affected by atmospheric nitrogen inputs.
In Lake Soyang, variations in δ15N-zooplankton depend on seasonal succession of phytoplankton, implying that δ15N-zooplankton, during periods when cyanobacteria dominate, would be lower than during the periods when diatoms are dominant (Vuorio et al. Citation2006; Wu et al. Citation2006). It is plausible that seasonality in δ15N-zooplankton in Lake Soyang is caused by succession of the phytoplankton species because different phytoplankton taxa may fractionate 15N differently under similar environmental conditions. Similar results have been reported in other lakes where indirect feeding on N-fixing cyanobacteria as a main food source was responsible for low δ15N-zooplankton. In Lake Soyang, the decrease in δ15N-zooplankton after monsoonal inflow of sediments and nutrients indicates that nitrogen fixation by cyanobacteria becomes more important because biologically fixed nitrogen has a δ15N of only −2 to 0‰ (Gu and Alexander Citation1993; Gu et al. 2006; Wu et al. Citation2006). Vuorio et al. (Citation2006), in four lakes in southwest Finland, showed that diatoms had higher δ15N than cyanobacteria and in particular Anabaena spp. had lower δ15N than other phytoplankton taxa. Although nitrogen fixation in Lake Soyang was positively correlated to Anabaena sp. heterocyst numbers (Heo and Kim Citation1997), heterocysts in Anabaena have not been commonly observed recently (Kim, personal observation). This is consistent with the typical N concentrations, which are almost always abundant relative to P concentrations for supporting phytoplankton growth. Therefore, we expect that cyanobacteria in Lake Soyang utilize dissolved nitrogen species originating from the watershed rather than atmospheric nitrogen. The variation in δ15N-zooplankton may be affected by an increase in fertilizer use, resulting from the increase in agricultural land use (Wang et al. Citation2005; Anderson and Cabana Citation2006; Choi et al. Citation2007). Anderson and Cabana (Citation2006) reported that the relationship between aquatic biota δ15N and anthropogenic nitrogen loads was highly correlated according to the origin of the N source.
Although bacteria and detritus might be important components in the diet of zooplankton, post-monsoon after the inflow of large amounts of sediment and nutrients to Lake Soyang, the major food sources for zooplankton are phytoplankton (Sim and Ahn Citation1992; Kim et al. Citation1999). The results of this study suggest that the difference in stable isotope composition of zooplankton between pre- and post-monsoon is controlled by the difference in food source and particularly, varies with hydrologic conditions in the reservoir and intensity of rainfall.
The purpose of this study was to investigate the carbon and nitrogen isotope composition (δ13C and δ15N) of POM and zooplankton in Lake Soyang, where the monsoonal climate leads to a characteristic seasonal pattern of a dramatic pulse of turbid runoff to the reservoir during the summer monsoon. Our study shows that δ13C- and δ15N-zooplankton clearly differed between pre-monsoon and post-monsoon periods in Lake Soyang. The difference in zooplankton δ13C and δ15N between pre-monsoon and post-monsoon periods showed seasonal patterns different from some previous studies on lakes in different climatic and hydrological regimes. The results suggest that seasonal changes in zooplankton δ13C and δ15N in Lake Soyang are closely related to the succession of phytoplankton (diatoms to cyanobacteria) and vary according to the re-supply of nutrients from the dense turbid inflowing layer of water during monsoon storms. The autochthonous organic matter component of POM in the epilimnion is probably the most important food source for zooplankton because most of the terrestrial-derived organic matter is discharged through the metalimnion at the reservoir outlet. Our results suggest that POM and zooplankton δ13C and δ15N measurements are important indicators for understanding the influence of turbidity and hydrologic inflow characteristics during the monsoon season and structuring the base of the food web in this deep reservoir.
Acknowledgements
We thank the students in the Environmental Research Laboratory for assisting with fieldwork. We appreciate the suggestions from three anonymous reviewers, which improved the paper. This study was supported by the CAER (Center for Aquatic Ecosystem Restoration) of Eco-STAR project (EW 42-08-10) from MOE (Ministry of Environment, Republic of Korea). This research was conducted for the Survey on the Environment and Ecosystem of Lakes in the Han River System in Fundamental Investigation on the Environment of the Han River project funded by the Han River Watershed Management Fund. Jae-Yong Lee was supported by Brain Korea 21 Program.
Related Research Data
References
- Anderson , C and Cabana , G . 2006 . Does δ15N in river food webs reflect the intensity and origin of N loads from the watershed? . Science of the Total Environment , 367 : 968 – 978 . doi: 10.1016/j.scitotenv.2006.01.029
- APHA, AWWA and WEF . 1998 . Standard methods for the examination of water and wastewater , 20th , 4 – 139 . Washington , DC : American Public Health Association .
- Bouillon , S C , Sreenivas , M N and Dehairs , F . 2000 . Sources of suspended organic matter and selective feeding by zooplankton in an estuarine mangrove ecosystem as traced by stable isotopes . Marine Ecology Progress Series , 208 : 79 – 92 . doi: 10.3354/meps208079
- Carabel , S , Godínez-Domínguez , E , Verísimo , P , Fernández , L and Freire , J . 2006 . An assessment of sample processing methods for stable isotope analyses of marine food webs . Journal of Experimental Marine Biology and Ecology , 336 : 254 – 261 . doi: 10.1016/j.jembe.2006.06.001
- Chen , F and Xie , P . 2003 . The effects of fresh and decomposed Microcystis aeruginosa on cladocerans from a subtropic Chinese Lake Taihu . Journal of Freshwater Ecology , 18 : 97 – 104 . doi: 10.1080/02705060.2003.9663955
- Choi , S I , Ahn , T S and Kato , K . 1996 . Image analysis of bacterial cell size by diurnal changes in Lake Soyang, Korea . Journal of Microbiology , 34 : 300 – 304 .
- Choi , K S , Kim , B C , Park , J H , Kim , Y H and Jun , M S . 2004 . Temporal and vertical variability in the relationship among organic matter indices in a deep reservoir ecosystem . Lake and Reservoir Management , 20 : 130 – 140 . doi: 10.1080/07438140409354357
- Choi , W J , Han , G H , Lee , S M , Lee , G T , Yoon , K S , Choi , S M and Ro , H M . 2007 . Impact of land-use types on nitrate concentration and δ15N in unconfined groundwater in rural areas of Korea . Agriculture Ecosystems and Environment , 120 : 259 – 268 . doi: 10.1016/j.agee.2006.10.002
- Del Giorgio , P A and France , R L . 1996 . Ecosystem-specific patterns in the relationship between zooplankton and POM or microplankton δ13C . Limnology and Oceanography , 41 : 359 – 365 . doi: 10.4319/lo.1996.41.2.0359
- Doi , H , Zuykova , E I , Kikuchi , E , Shikano , S , Kanou , K , Yurlova , N and Yadrenkina , E . 2006 . Spatial changes in carbon and nitrogen stable isotopes of the plankton food web in a saline lake ecosystem . Hydrobiologia , 571 : 395 – 400 . doi: 10.1007/s10750-006-0205-1
- Grey , J , Jones , R I and Sleep , D . 2000 . Stable isotope analysis of the origins of zooplankton carbon in lakes of differing trophic state . Oecologia , 123 : 232 – 240 . doi: 10.1007/s004420051010
- Grey , J and Jones , R I . 2001 . Seasonal changes in the importance of the source of organic matter to the diet of zooplankton in Loch Ness, as indicated by stable isotope analysis . Limnology and Oceanography , 46 : 505 – 513 . doi: 10.4319/lo.2001.46.3.0505
- Grice , A M , Loneragan , N R and Dennison , W C . 1996 . Light intensity and the interactions between physiology, morphology and stable isotope ratios in five species of seagrass . Journal of Experimental Marine Biology and Ecology , 195 : 91 – 110 . doi: 10.1016/0022-0981(95)00096-8
- Gu , B and Alexander , V . 1993 . Estimation of N2 fixation based on differences in the natural abundance of 15N among freshwater N2 fixing and non-N2 fixing algae . Oecologia , 96 : 43 – 48 . doi: 10.1007/BF00318029
- Gu , B and Schelske , C L . 1996 . Temporal and spatial variations in phytoplankton carbon isotopes in a polymictic subtrophical lake . Journal of Plankton Research , 18 : 2081 – 2092 . doi: 10.1093/plankt/18.11.2081
- Gu , B , Alexander , V and Schell , D M . 1999 . Seasonal and interannual variability of plankton carbon isotope ratios in a subarctic lake . Freshwater Biology , 42 : 417 – 426 . doi: 10.1046/j.1365-2427.1999.00472.x
- Gu , B , Chapman , A D and Schelske , C L . 2006 . Factors controlling seasonal variations in stable isotope composition of particulate organic matter in soft water eutrophic lake . Limnology and Oceanography , 51 : 2837 – 2848 . doi: 10.4319/lo.2006.51.6.2837
- Gu , B , Schelske , C L and Waters , M N . 2011 . Patterns and controls of seasonal variability of carbon stable isotopes of particulate organic matter in lakes . Oecologia , 165 : 1083 – 1094 . doi: 10.1007/s00442-010-1888-6
- Heo , W M and Kim , B C . 1997 . The change in N/P ratio with eutrophication and cyanobacterial blooms in Lake Soyang, Korea . Verhandlungen des Internationalen Verein Limnologie , 26 : 491 – 495 .
- Heo , W M , Kim , B C , Kim , Y H and Choi , K S . 1998 . Storm runoff of phosphorus from nonpoint sources into Lake Soyang and transportation of turbid water mass within the lake . Korean Journal of Limnology , 31 : 1 – 8 [in Korean with English abstract] .
- Hwang , S J . 1997 . Impact of eutrophication on the microbial food web dynamics in Lake Erie ecosystem . Korean Journal of Limnology , 30 ( Suppl ) : 496 – 505 .
- Jones , R I , King , L , Dent , M M , Maberly , S C and Gibson , C E . 2004 . Nitrogen stable isotope ratios in surface sediments, epilithon and macrophytes from upland lakes with differing nutrient status . Freshwater Biology , 49 : 382 – 391 . doi: 10.1111/j.1365-2427.2004.01194.x
- Kaown , D , Koh , D C , Mayer , B and Lee , K K . 2009 . Identification of nitrate and sulfate sources in groundwater using dual stable isotope approaches for an agricultural area with different land use (Chuncheon, mid-eastern Korea) . Agriculture, Ecosystems and Environment , 132 : 223 – 231 . doi: 10.1016/j.agee.2009.04.004
- Kim , B C , Kim , J O , Jun , M S and Hwang , S J . 1999 . Seasonal dynamics of phytoplankton and zooplankton community in Lake Soyang . Korean Journal of Limnology , 32 : 127 – 134 . [in Korean with English abstract]
- Kim , J K , Kim , B C , Jung , S M , Jang , C W , Shin , M-S and Lee , Y K . 2007 . The distribution of DOM and POM and the composition of stable carbon isotopes in streams of agricultural and forest watershed located in the Han River system, Korea . Korean Journal of Limnology , 40 : 93 – 102 . [in Korean with English abstract]
- Kim , J K . 2007. The distribution of DOC and POC and stable carbon isotope ratios in streams and reservoirs with different land use. PhD Dissertation. Kangwon National University
- Lee , J Y , Islam , J M , Shin , M S , Jung , S M and Kim , B C . 2010 . The spatial and temporal variation of periphyton in the Inbuk stream . Journal of Korean Society on Water Quality , 26 : 104 – 110 . [in Korean with English abstrac]
- Liu , Y , Xie , P and Wu , X P . 2006 . Effects of toxic and nontoxic Microcysis aeruginosa on survival, population-increase, and feeding of two small cladocerans . Bulletin of Environmental Contamination and Toxicology , 77 : 566 – 573 . doi: 10.1007/s00128-006-1101-0
- Minagawa , M and Wada , E . 1984 . Stepwise enrichment of 15N along food chains: further evidence and the relation between δ15N and animal age . Geochimica et Cosmochimica Acta , 48 : 1135 – 1140 . doi: 10.1016/0016-7037(84)90204-7
- Namkung , H , Kim , B C , Hwang , G S , Choi , K S and Kim , C G . 2001 . Organic matter sources in a reservoir (Lake Soyang); primary production of phytoplankton and DOC, and external loading . Korean Journal of Limnology , 34 : 166 – 174 . [in Korean with English abstract]
- Ostrom , N E , Long , D T , Bell , E M and Beals , T . 1998 . The origin and cycling of particulate and sedimentary organic matter and nitrate in Lake Superior . Chemical Geology , 152 : 13 – 28 . doi: 10.1016/S0009-2541(98)00093-X
- Park , H K , Kwon , O Y and Jung , D I . 2011 . Characteristics of allochthonous organic matter in large dam reservoir, Lake Soyang . Journal of Korean Society on Water Quality , 27 : 88 – 97 [in Korean with English abstract] .
- Park , J C , Park , J W , Shin , J K and Lee , H M . 2005 . Dynamics of high turbid water caused by heavy rain of monsoon and typhoon in a large Korean reservoir (Andong Reservoir) . Korean Journal of Limnology , 38 : 105 – 117 . [in Korean with English abstract]
- Perga , M E and Gerdeaux , D . 2006 . Seasonal variability in the δ13C and δ15N values of the zooplankton taxa in two alpine lakes . Acta Oecologica , 30 : 69 – 77 . doi: 10.1016/j.actao.2006.01.007
- Pulido-Villena , E , Reche , I and Morales-Baquero , R . 2005 . Food web reliance on allochthonous carbon in two high mountain lakes with contrasting catchments: a stable isotope approach . Canadian Journal of Fisheries Aquatic Sciences , 62 : 2640 – 2648 . doi: 10.1139/f05-169
- Rautio , M and Vincent , W F . 2007 . Isotopic analysis of the sources of organic carbon for zooplankton in shallow subartic and arctic waters . Ecography , 30 : 7 – 87 .
- Risk , M J , Lapointe , B E , Sherwood , O A and Bedford , B J . 2009 . The use of δ15N in assessing sewage stress on coral reefs . Marine Pollution Bulletin , 58 : 793 – 802 . doi: 10.1016/j.marpolbul.2009.02.008
- Sim , D S and Ahn , T S . 1992 . On the feeding behavior of zooplankton in Lake Soyang . The Korean Journal of Microbiology , 30 : 129 – 133 . [in Korean with English abstract]
- Syväranta , J , Hämäläinen , H and Jones , R . 2006 . Within-lake variability in carbon and nitrogen stable isotope signatures . Freshwater Biology , 51 : 1090 – 1102 . doi: 10.1111/j.1365-2427.2006.01557.x
- Vander Zanden , M J and Rasmussen , J B . 2001 . Variation in δ15N and δ13C trophic fractionation: implications for aquatic food web . Limnology and Oceanography , 46 : 2061 – 2066 . doi: 10.4319/lo.2001.46.8.2061
- Vuorio , K , Meili , M and Sarvala , J . 2006 . Taxon-specific variation in the stable isotopic signatures (δ13C and δ15N) of lake phytoplankton . Freshwater Biology , 51 : 807 – 822 . doi: 10.1111/j.1365-2427.2006.01529.x
- Wang , H , Magesan , G N , Clinton , P W and Lavery , J M . 2005 . Using natural 15N abundances to trace the fate of waste-derived nitrogen in forest ecosystems: New Zealand case studies . Isotopes in Environmental and Health Studies , 41 : 31 – 38 . doi: 10.1080/10256010500053250
- Wu , J , Lin , L , Gagan , M K , Schleser , G H and Wang , S . 2006 . Organic matter stable isotope (δ13C and δ15N) response to historical eutrophication of Lake Taihu, China . Hydrobiologia , 563 : 19 – 29 . doi: 10.1007/s10750-005-9133-8
- Yoshioka , T , Hayashi , H and Wada , E . 1989 . Seasonal variations of carbon and nitrogen isotope ratios of plankton and sinking particles in Lake Kizaki . Japanese Journal of Limnology , 50 : 313 – 320 . doi: 10.3739/rikusui.50.313
- Yoshioka , T . 1997 . Phytoplanktonic carbon isotope fractionation: equations accounting for CO2-concentrating mechanisms . Journal of Plankton Research , 19 : 1455 – 1476 . doi: 10.1093/plankt/19.10.1455
- Zohary , T , Erez , J , Gophen , M , Berman-Frank , I and Stiller , M . 1994 . Seasonality of stable carbon isotopes within the pelagic food web of Lake Kinneret . Limnology and Oceanography , 39 : 1030 – 1043 . doi: 10.4319/lo.1994.39.5.1030