Abstract
We surveyed benthic macroinvertebrates in the Upper Mississippi River near Minneapolis and St. Paul, Minnesota, USA (1350–1380 river km) using Hester-Dendy multiplate samplers as artificial substrates. Two study sites were sampled in multiple seasons at two depths over two years during the period from 2007 to 2009. We examined the patterns of seven community metrics summarizing richness levels and/or relative abundances of major community subsets. Richness metrics were driven by rare (occurring in less than 10% of the samples) taxa but overall densities and relative abundances of major groups reflected the presence of a few dominant taxa. Using analysis of variance, community metrics differed between study years or sampling depths only sporadically, but consistently differed among survey periods. Non-metric multidimensional scaling analysis indicated samples primarily clustered by survey period (within years) and sampling depth at one of our sites and by study year and survey period at the other site. A relatively small number of taxa were consistently abundant in our study, many of which have been reported as community dominants in other large river studies. Annual variability in assemblage composition generally reflected atypically high numbers of one or more of these persistent dominants, and was likely related to differences in hydrological conditions between years. Seasonal variability, however, was relatively consistent between years at both study sites.
Introduction
Biological assessments of wadeable streams have revealed the structures of their macroinvertebrate communities across much of the eastern United States. There is also interest in evaluating macroinvertebrate community structure to facilitate bioassessment in great (ninth–tenth order) rivers (Poulton et al. Citation2003; Applegate et al. Citation2007; Angradi, Pearson, et al. Citation2009). However, the difficulty in collecting samples from these systems, due to their greater width and depth, and their complex hydrology, has limited the information available (Johnson et al. Citation2006). Battle et al. (Citation2007) observed that published investigations of lotic systems were dominated by studies of streams smaller than the seventh order, and that studies on great rivers were generally of short duration and in small geographic areas. Two recent exceptions to the trend were the basin-wide sampling of the Ohio River initiated in 1991 by the Ohio River Valley Water Sanitation Commission (Applegate et al. Citation2007) and the US Environmental Protection Agency (EPA) surveys covering most of the Upper Mississippi, Missouri, and Ohio Rivers from 2004 to 2006 (Angradi, Bolgrien, et al. Citation2009). These studies had numerous objectives, including descriptions of habitat associations, demonstrating longitudinal – and in the US EPA study, inter-basin – trends in assemblage composition and evaluating macroinvertebrate community characteristics with regard to their suitability for multimetric biological assessments.
Published studies of seasonal dynamics in great river macroinvertebrate communities are not as numerous as those of smaller lotic systems (but, see Modde and Schmulbach Citation1973; Barnum and Bachmann Citation1988). In wadeable streams, substantial temporal variability in macroinvertebrate community composition has been documented (Minshall Citation1981; Robinson et al. Citation1990; Armitage et al. Citation1995) sometimes between periods as short as 2 weeks (McCord and Lambrecht Citation2006). Short-term fluctuations in the abundances of macroinvertebrate taxa in streams are likely attributable to life history dynamics or the consequences of disturbance (Robinson et al. Citation1990). Seasonal or other short-term temporal variation has been examined indirectly, if at all, in the recent large river studies we reviewed. Hay (Citation2006) reported that total drift density in the Missouri River was positively correlated with degree-days. Also in the Missouri River, Grohs (Citation2008) noted higher total richness in the summer and fall, but discriminatory power was limited by family-level identifications. And, although temporal patterns were not a focal point of their study, Angradi, Pearson, et al. (Citation2009) found that community index scores occasionally changed between great river samples taken at the same location several weeks apart.
In this study, we examined the macroinvertebrate community structure and its short-term temporal patterns at two depths across two study years at two locations in the Upper Mississippi River. Our objective was to determine whether community characteristics such as richness and the relative contributions of major subgroups (Ephemeroptera, Plecoptera, and Trichoptera (EPT), Chironomidae, non-insects) were consistently similar within seasons but differed substantially between seasons. We also compared the macroinvertebrate communities at these locations to those reported in other great river studies.
Methods
Our study reach was in the impounded section of the Upper Mississippi River approximately 1350–1380 river km upstream of the entrance of the Ohio River into the Mississippi River (). Two 3-km study areas, located near the High Bridge (HB; 44°56′16″N, −93°5′59″W) and Riverside (RIV; 45°0′55″N, −93°16′30″W) electric power plants in the Minneapolis/St. Paul metropolitan area, were surveyed for benthic macroinvertebrates in 2008–2009 (HB) and 2007 and 2009 (RIV). Samples were not collected at RIV in 2008 due to nearby construction activities in the river.
Figure 1. Map of the study area in relation to the Upper Mississippi River, and illustrating sample stations at the HB and RIV Generating Stations, 2007–2009.
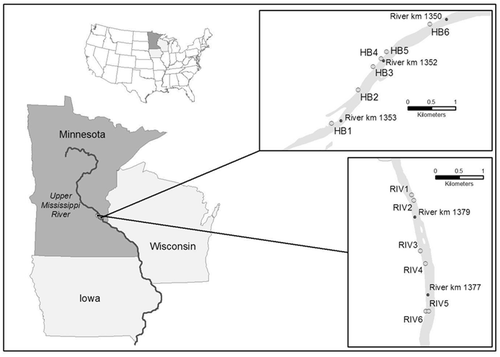
At each study area, samples (one sample equaled six replicate sampling units) were collected from six sites in the river mainstem using Hester-Dendy artificial substrates. Each Hester-Dendy unit consisted of 14 round plates 75 mm in diameter, with variable (3–12 mm) spacing between plates; the total surface area of each unit was 0.1277 m2. Sampling units were attached to a 3-mm-coated steel cable at mid-water and just above the river bottom. At HB, the mid-water samples ranged from 1.5 to 3 m deep (mean = 2 m) and bottom samples were taken at depths of 3–6 m (mean = 4 m). At RIV, mid-water samples ranged from 1.3 to 2.7 m (mean = 1.5 m) and bottom samples were 2.3 to 5 m deep (mean = 3.2 m).
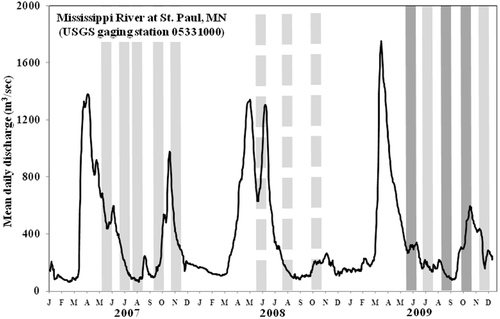
Samplers were retrieved after 30-day colonization periods. There were three colonization periods per year at HB and five per year at RIV (). This resulted in six samples from both mid-water and bottom habitats for each survey period. Upon retrieval, samplers were placed into #30 (500 µm mesh) sieve buckets, disassembled, and the plates rinsed with river water. Organisms and debris were placed in plastic sample jars and preserved with 95% ethanol. In the laboratory, target numbers for samples were 250–500 organisms. In most cases (70% at HB, 75% at RIV), this required processing of the entire sample. When large numbers of organisms were encountered, samples were split once, or in rare circumstances, twice. In HB samples, macroinvertebrates were identified to the lowest practical taxon, which was typically genus or species group for insects and crustaceans, genus or family for oligochaetes and mollusks, and class for turbellarians. For RIV samples, insects were only identified to genus; thus, the taxonomic endpoints for this study area were generally somewhat conservative.
Figure 2. Mean daily discharge in the Mississippi River at St. Paul, MN, 2007–2009. Shaded bars indicate periods of sample collection. Light gray (solid line) indicates RIV samples; light gray (dashed line) indicates HB samples; and dark gray (solid line) indicates both RIV and HB samples.
Seven community composition metrics commonly used in bioassessments (DeShon Citation1995; Wessell et al. Citation2008; Weigel and Dimick Citation2011) were calculated for each sample: abundance (per 0.1277 m2 sampling unit), total richness, richness within the orders EPT, richness within the dipteran family Chironomidae, and the relative abundances of EPT organisms, chironomids, and non-insects. We also identified the major taxa from each study area as those that occurred in at least 35% of samples and accounted for >1% of the total collection. Community variables and major taxa abundances were entered into a one-way ANOVA to test for differences between years, depths, and survey periods, followed by a Tukey's test when there were more than two treatment groups. When assumptions of either normality (Kolmogorov–Smirnov test) or equal variance (Levene's test) were violated, a non-parametric Kruskal–Wallis test was substituted for the ANOVA and a Mann–Whitney test with Bonferroni correction for the α-level was substituted for the Tukey's test. The non-parametric tests were typically used for the tests of major taxa differences, as one or both assumptions were nearly always violated.
We used the non-metric multidimensional scaling (NMS) ordination technique in PC-ORD, Version 5 software to examine compositional similarity among samples (McCune and Grace Citation2002). Analyses were run on log (x + 1) transformed data for taxa that were present, on average, at least once per sampling event (resulting in 80 taxa for HB and 59 for RIV). We ran separate analyses rather than combining the data sets because of the differences in taxonomic endpoints and differing survey periods between sites. NMS settings were Sorensen (Bray–Curtis) distance measure and a step length of 0.20. Monte Carlo tests recommended a 3-dimensional solution (p = 0.02) for both the HB and RIV datasets. The HB solution had a final stress of 11.5 and instability of 0.00001 after 75 iterations; the RIV solution had a final stress = 14.0 and instability = 0.00001 after 84 iterations. Both NMS ordinations employed a second matrix of macroinvertebrate data and hydrologic data – mean flow for the 30 days the sampling units were in the river (US Geological Survey gage 05331000 at St. Paul, Minnesota). The cut-off r 2 was 0.37 for both ordinations. We expected the results to serve as confirmation of the analysis of variance, i.e., general patterns of temporal variation and/or variation between depths would be evident in both the ANOVA and the ordination.
Results
The overall taxonomic richness levels were comparable between the study sites with 120 taxa found at the HB site and 108 at the RIV site (). The lower number of taxa at RIV despite the greater number of samples (120, as compared to 71 at HB) reflected the more conservative identification endpoints. At HB, EPT taxa and chironomids each contributed over 40% of the total collection, and non-insects added another 10%. At RIV, chironomids were more prevalent, comprising 65% of the total, with EPT taxa and non-insects adding 27% and 7%, respectively. Nineteen major taxa were identified from HB samples and 16 were identified from RIV samples. Collectively, these major taxa comprised 85% of the total number collected at HB and 91% at RIV. There was considerable similarity in composition at the two study sites. The amphipod Gammarus and turbellarians were the most abundant non-insect taxa at both HB and RIV. Numerically dominant EPT taxa consistently included heptageniid mayflies (Maccaffertium spp.), and hydropsychid (Cheumatopsyche and P. flava) and polycentropodid (Cyrnellus fraternus) caddisflies. Major chironomid taxa included Thienemannimyia group, Dicrotendipes spp., Glyptotendipes, Polypedilum convictum group, Stenochironomus, and Rheotanytarsus in both study areas. Total and relative abundance variables largely reflected the presence of these major taxa, but richness variables were more driven by uncommon and rare taxa.
Table 1. Mean macroinvertebrate community characteristics and abundances of major taxa in Hester-Dendy samples from the Upper Mississippi River near HB Generating Station.
Community variables differed between study years or between sampling depths only sporadically but consistently differed among survey periods. At HB, no community variable differed significantly between years (). Total abundance was similar in both years, and the only major taxa whose numbers differed between years were Turbellaria, Dicrotendipes modestus/neomodestus, and Glyptotendipes. EPT richness and Percent EPT were greater in mid-water samples, reflecting increased abundance of mayflies and hydropsychid caddisflies, particularly Hydropsyche bidens/orris. Percent non-insects was greater in bottom samples, where Gammarus and turbellarians were more numerous. In terms of differences among survey periods (pooled years), total abundance did not differ but all richness variables were highest in June and lowest in October. Percent EPT was greater in June and August, in part reflecting greater abundances of Caenis, Tricorythodes, and Cheumatopsyche in June and M. integrum, Pseudocloeon, and C. fraternus in August. Although not differing significantly among periods, H. bidens/orris and P. flava also comprised much of the EPT abundance in June and/or August. In October, the assemblage was dominated by chironomids, due to greater abundances of Glyptotendipes and to a lesser extent, D. lucifer/simpsoni. At RIV, chironomid richness was higher in 2007 (). Percent non-insects was greater in 2009, when greater numbers of Gammarus and Oligochaeta were collected. The relative presences of EPT organisms and chironomids were similar between years, but abundances of major taxa within these groups differed considerably. Significant differences were evident for Cricotopus, Polypedilum, and Rheotanytarsus (greater in 2007), and for Polycentropus, Thienemannimyia gp., Dicrotendipes, Glyptotendipes, and Stenochironomus (greater in 2009). No community characteristics or abundances of major taxa differed between depths at RIV. With regard to seasonal variation, total abundance was greatest in October and least in November. Conversely, all richness variables were lowest in October. Percent EPT was greater in May and November than in the late summer/early fall months, whereas percent Chironomidae was greatest in October and August. Differences in the abundances of major taxa between survey periods at RIV were more ambiguous but could be categorized into one of two groups. The first included taxa that were generally most numerous in the summer and/or early fall (July through October), such as Turbellaria, C. fraternus, Polycentropus, Dicrotendipes, Glyptotendipes, and Stenochironomus. The second group included taxa that were more abundant in spring (May) and/or late fall (November), e.g., Cheumatopsyche, P. flava, Cricotopus, and Polypedilum.
Table 2. Mean macroinvertebrate community characteristics and abundances of major taxa in Hester-Dendy samples from the Upper Mississippi River near RIV Generating Station.
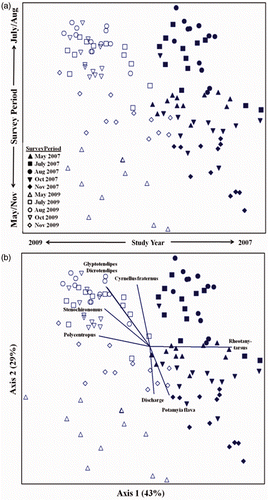
At HB, the three axes of the NMS ordination accounted for 91% of the variance in the dataset, with axes 1 and 2 accounting for 70% of the total. Samples were generally clustered by survey period and sample depth; axis 1 differentiated samples from different periods based on greater abundances of D. lucifer/simpsoni (r = 0.63) and Glyptotendipes (r = 0.69) in October versus high numbers of M. integrum, Pseudocloeon, P. convictum gp, and Rheotanytarsus in June (r = −0.64 to − 0.72; ). Axis 2 differentiated samples between depths based on higher abundances of Cheumatopsyche, H. bidens/orris, and P. flava, in mid-water samples (r = 0.61–0.73) versus greater numbers of Tubificidae without hair chaetae (r = −0.64) in bottom samples. At RIV, the three ordination axes cumulatively accounted for 86% of the variance, and axes 1 and 2 accounted for 72% of the total. Samples were clustered primarily by study year and secondarily by survey period; axis 1 differentiated samples from years based on greater abundance of Rheotanytarsus in 2007 (r = 0.84) versus higher abundances of Polycentropus, Dicrotendipes, Glyptotendipes, and Stenochironomus in 2009 (r = −0.60 to −0.66; ). Axis 2 differentiated samples between survey periods based on greater abundances of C. fraternus, Dicrotendipes, and Glyptotendipes in August and October samples (r = 0.69–0.70) versus P. flava in May and November samples (r = −0.62). Axis 2 was also inversely correlated (r = −0.62) with mean discharge during the colonization period.
Figure 3. NMS ordination of macroinvertebrate assemblages from the Upper Mississippi River at the HB Generating Station in St. Paul, MN. (a) Illustrates seasonal and annual groupings among 3 survey periods in 2008 and 2009. Vectors indicate taxa (r 2 > 0.37) associated with axes. ‘T w/o HC’ indicates Tubificidae without hair chaetae. (b) Same ordination but symbols indicate sample depth.
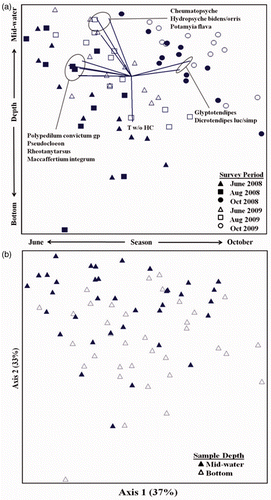
Figure 4. NMS ordination of macroinvertebrate assemblages from the Upper Mississippi River at the RIV Generating Station in Minneapolis, MN. (a) Illustrates seasonal and annual groupings among five survey periods in 2007 and 2009. (b) Same ordination but includes vectors for taxa (r 2>0.37) and mean river discharge during surveys.
Discussion
Assemblage structure
This study revealed a macroinvertebrate assemblage in the Upper Mississippi River comparable in taxonomic composition to that reported in other great river studies. Major groups included amphipods (Gammarus), turbellarians, heptageniid mayflies (primarily Maccaffertium spp.), polycentropodid (C. fraternus), and hydropsychid (Cheumatopsyche, Hydropsyche spp., and P. flava) caddisflies, and the chironomids Thienemannimyia gp., Dicrotendipes spp., Glyptotendipes, Polypedilum spp., and Rheotanytarsus. Assemblages also included large numbers of taxa that occurred more sporadically and in lower numbers. Indeed, total richness estimates were largely reflective of these rarer taxa. In this study, taxa other than the major groups comprised 10–15% of the total abundance but 75–80% of the total richness in samples. This pattern was similar to that reported for a third order Rocky Mountain stream (Minshall Citation1981), and probably reflects a characteristic of macroinvertebrate communities over a large size range of lotic systems.
We compared macroinvertebrate community characteristics encountered at HB in this study to those reported from other large river studies that used artificial substrates. We did not include RIV in the comparison because of the differences in taxonomic endpoints relative to HB and the other studies. In terms of densities and relative abundances of major groups, our samples were similar to Hester-Dendy samples from the Ohio River (R. Argo, Ohio River Valley Sanitation Commission (ORSANCO), pers. comm.) and dissimilar to rock basket samples from the Upper Mississippi River (Battle et al. Citation2007) and Lower Missouri River (Poulton et al. Citation2003; ). Generally, the rock basket samples contained much higher densities, which reflected great numbers of hydropsychid caddisflies. As a result, percent EPT was substantially greater than either percent Chironomidae or percent non-insects. Total richness and chironomid richness levels were lower in the rock basket data sets (Poulton et al. Citation2003; Battle et al. Citation2007) but this likely reflected the smaller amount of sampling effort. EPT richness was highest in the rock basket samples of the Upper Mississippi River (Battle et al. Citation2007). Richness variation may also have reflected habitat differences of the particular sampling locations; rock baskets were set on the immersed substrates of wing dikes whereas the ORSANCO Hester-Dendy samplers were in the water column near shore. In the two Hester-Dendy studies, macroinvertebrate community composition primarily differed with regard to the relative abundances of EPT organisms (greater in the study reported here) and non-insects (greater in the Ohio River datasets).
Table 3. Comparison of the present study to elements of selected other macroinvertebrate studies on the Upper Mississippi, Missouri, and Ohio Rivers.
The major taxa in all of these studies were similar to each other. In the Upper Mississippi River approximately 1200 km downstream from our study area, Battle et al. (Citation2007) reported an assemblage dominated by H. bidens/orris and P. flava, and to a lesser extent Polypedilum and Rheotanytarsus. Poulton et al. (Citation2003) likewise reported samples from the Lower Missouri River dominated by H. orris, H. scalaris, P. flava, and M. integrum. Their dominant chironomid taxa included Dicrotendipes and Polypedilum. In the Ohio River, Gammarus fasciatus, C. fraternus, D. lucifer/simpsoni, and D. modestus/neomodestus were numerically dominant for the length of the river (R. Argo, ORSANCO, pers. comm.). However, there was also a longitudinal shift of decreasing abundance of the mayfly Stenacron interpunctatum and the gastropod Hydrobiidae combined with increasing abundance of the zebra mussel Dreissena polymorpha and the chironomid Glyptotendipes moving from the upper to the lower river. The large-scale variation noted among these datasets may have been associated with physical differences between the more natural substrates of the rock baskets as opposed to the tempered masonite substrate of the Hester-Dendy units. Alternatively, they may have reflected the different flow regimes; that is, greater current velocity in the Missouri River and ‘open’ portion of the Upper Mississippi River as opposed to the impounded Ohio River and upper segments of the Upper Mississippi River (Delong Citation2005; Galat et al. Citation2005). Fremling (Citation1960) noted current velocity preferences for H. orris (faster) as compared to Cheumatopsyche and P. flava (slower) in the Upper Mississippi River.
Annual variability
In their five-year study on an open river segment of the Upper Mississippi River, Battle et al. (Citation2007) found that macroinvertebrate densities in rock basket samples varied greatly between years and inferred that annual patterns were probably associated with the dynamics of hydrologic conditions. We observed a similar trend, but of smaller magnitude, at our RIV site. With regard to community-scale characteristics, the difference between study years was slight. But, abundances of individual taxa did vary considerably between years at both HB and RIV. Although only three major taxa at HB and 10 at RIV differed by significant margins using ANOVA, the ordination for RIV suggested that sample composition clustered primarily by survey year. The chironomids Dicrotendipes, Glyptotendipes, Stenochironomus, and Rheotanytarsus were among the taxa whose abundances were most associated with the difference. The first three of those taxa were also consistently abundant in samples from the impounded Ohio River (R. Argo, ORSANCO, pers. comm.), whereas Rheotanytarsus was the most numerous chironomid taxa in the Battle et al. (Citation2007) study as well as in the Lower Missouri (Poulton et al. 2005) and Lower Mississippi Rivers (Way et al. Citation1995) where fast-flowing conditions predominated. We infer that the pattern in the present study is additional evidence of the effect of hydrological conditions on macroinvertebrate assemblage composition hypothesized by Battle et al. (Citation2007). Flow conditions (as expressed by mean daily discharge) in the Upper Mississippi River section containing our study area generally ranged between 85 and 1400 m3 s−1 from 2007 to 2009 (USGS Citation2010). In contrast, Upper Mississippi River discharge near Chester, Illinois, which is approximately 90 river km upstream of the Battle et al. (Citation2007) study area, ranged from 2800 to 17,000 m3 s−1 during the same period. So, while the hydrological variability was much less in our study area, the effect on macroinvertebrate community composition was still discernable.
Differences between depths and seasons
Delong and Payne (Citation1985) found differences in macroinvertebrate richness and abundance between depths and hypothesized that they may have been related to varying food availability and/or the effect of negative phototaxis. In the present study, assemblage characteristics and the abundances of certain taxa varied between depths at one study area (HB) but not the other. The depth difference at HB was most associated with increased abundance of hydropsychid caddisflies in mid-water samples, but these taxa did not differ between depths at RIV. Sample locations were nearly 1 m deeper, on average, at HB than at RIV, and this difference may have been sufficient for a pattern of depth preference to emerge. Sampling depths in this study were greater than in the Ohio River studies (R. Argo, ORSANCO, pers. comm.). This difference may also have been responsible for the compositional differences noted.
Studies of large river macroinvertebrate communities frequently have the goal of developing metrics for biological criteria to be applied to these systems (Poulton et al. Citation2003; Applegate et al. Citation2007; Angradi, Pearson, et al. Citation2009). By design, the studies collect samples within a 1 - or 2-month period to standardize data for comparison with later collections. In bioassessment studies, metrics such as the richness levels and percent compositions of various groups are used as estimates of community quality. Specifically, as the values of total richness, EPT richness, and relative abundance of EPT organisms increase, the community is considered to be of higher quality (Barbour et al. Citation1999). Reduced richness and/or increased relative abundances of chironomids and non-insects are generally interpreted as an evidence of impairment. But these community characteristics may also vary for natural reasons, such as stream size or life cycle-related seasonal differences (Lenat Citation1983). Studies of smaller streams have demonstrated substantial variation in assemblage composition between seasons (Williams Citation1980; Minshall Citation1981; Robinson et al. Citation1990; Armitage et al. Citation1995). In the Upper Mississippi River, we found that richness and the relative abundance of EPT organisms decreased and the relative abundance of chironomids increased from spring/early summer to late summer/fall. There was evidence at one of our locations (RIV) that the assemblage may return to its earlier characteristics in the late fall, a pattern similar to that noted in the upper Missouri River by Modde and Schmulbach (Citation1973). Nearly all of the major taxa in our study were present throughout 5–6-month survey periods, but their numbers varied dramatically on a short-term temporal basis that likely reflected their life-history patterns, rather than any impairment effect. Seasonal flow patterns during the three-year period of this study were comparable, i.e., highest in April/May and generally low in the summer and winter ().
Future large river studies
A challenge for future bioassessments of large river macroinvertebrate communities is the numerical dominance of a relatively small number of taxa (Battle et al. Citation2007). The fact that several of the major taxa (e.g., M. integrum, C. fraternus, H. bidens/orris, and D. modestus/neomodestus) are present across large geographic distances contrasts the compositional patterns of small streams, in which assemblage dominants often differ between streams of adjacent watersheds (MDNR Citation2010; OEPA Citation2010). Thus, the pool of community dominants – with varying habitat requirements, feeding modes, and pollution tolerances (Johnson et al. Citation2006) – on which stressors can have an observable effect is much reduced. An additional challenge involves the ‘fourth dimension’ of the macroinvertebrate community structure – temporal variation (Ward Citation1989). Although Ward's emphasis was on response times to disturbances or evolutionary changes, his point that lotic ecosystems are temporally dynamic also applies to interpretation of bioassessment data. If sampled assemblages are in a different successional state from the reference condition to which they are compared, another source of variation is added to those of interest. In the present study, an example is the difference in assemblage composition between mid-October and late November. Between these sampling periods at RIV, the emergence of a few major taxa resulted in a community shift from a high abundance, low richness assemblage dominated by chironomids to a lower abundance, higher richness assemblage with approximately equal presence of EPT organisms and chironomids. Multi-metric indices are designed to be robust in spite of this variation, but consideration of temporal differences as well as adjustments to account for dominant taxa (Jackson et al. Citation2009) can improve their interpretive strength.
Acknowledgments
All information that was not publically available is acknowledged. We are very grateful to Brad Giese, Brian Schmidt, Seth Carlson, and Luke Kusilek for assistance in the field, and to Len Ferrington and his students for help with sample processing and taxonomic identifications at RIV. We also thank Ryan Argo for providing data from the Ohio River. Jan Battle provided invaluable guidance with data analysis. Comments by Ted Angradi, Jan Battle, Todd Hubbard, Casey Scott, and two anonymous reviewers greatly improved the manuscript. We also thank Jeff Barrington, Jim Bodensteiner and Dan Orr for their support of this project. The research was supported by grants from Xcel Energy.
References
- Angradi , T R , Bolgrien , D W , Jicha , T M , Pearson , M S , Taylor , D L and Hill , B H . 2009 . Multispatial-scale variation in benthic and snag-surface macroinvertebrate assemblages in mid-continent US great rivers . Journal of the North American Benthological Society , 28 : 122 – 141 . doi: 10.1899/07-149.1
- Angradi , T R , Pearson , M S , Bolgrien , D W , Jicha , T M , Taylor , D L and Hill , B H . 2009 . Multimetric macroinvertebrate indices for mid-continent US great rivers . Journal of the North American Benthological Society , 28 : 785 – 804 . doi: 10.1899/09-003.1
- Applegate , J M , Baumann , P C , Emery , E B and Wooten , M S . 2007 . First steps in developing a multimetric macroinvertebrate index for the Ohio River . River Research and Applications , 23 : 683 – 697 . doi: 10.1002/rra.1011
- Armitage , P D , Pardo , I and Brown , A . 1995 . Temporal constancy of faunal assemblages in “mesohabitats” – application to management? . Archives of Hydrobiology , 133 : 367 – 387 .
- Barbour , M T , Gerritsen , J , Snyder , B D and Stribling , J B . 1999 . Rapid bioassessment protocols for use in wadeable streams and rivers: periphyton, benthic macroinvertebrates, and fish , 2nd , Washington (DC) : US Environmental Protection Agency; Office of Water . (EPA report 841-B-99-002)
- Barnum , J B and Bachmann , R W . 1988 . Benthic macroinvertebrate habitat associations of the channelized middle Missouri River . Journal of the Iowa Academy of Science , 95 : 60 – 65 .
- Battle , J M , Jackson , J K and Sweeney , B W . 2007 . Annual and spatial variation for macroinvertebrates in the Upper Mississippi River near Cape Girardeau, Missouri . Archives of Hydrobiology , 168 : 39 – 54 .
- Delong , M D . 2005 . “ Upper Mississippi River basin ” . In Rivers of North America , Edited by: Benke , A C and Cushing , C E . 327 – 373 . Burlington (MA) : Elsevier .
- Delong , M D and Payne , J F . 1985 . Patterns of colonization by macroinvertebrates on artificial substrate samplers: the effects of depth . Freshwat Invert Biology , 4 : 194 – 200 . doi: 10.2307/1467161
- DeShon , J E . 1995 . “ Development and application of the invertebrate community index (ICI) ” . In Biological assessment and criteria: tools for water resource planning and decision making , Edited by: Davis , W S and Simon , T P . 217 – 243 . Boca Raton (FL) : Lewis Publishers .
- Fremling , C R . 1960 . Biology and possible control of nuisance caddisflies of the Upper Mississippi River . Iowa State University of Science and Technology, Research Bulletin , 483 : 856 – 879 .
- Galat , D L , Berry , C S , Peters , E J and White , R G . 2005 . “ Missouri River basin ” . In Rivers of North America , Edited by: Benke , A C and Cushing , C E . 427 – 480 . Burlington (MA) : Elsevier .
- Grohs , K L . 2008. Macroinvertebrate composition and patterns of prey use by juvenile pallid sturgeon (Scaphirhynchus albus) in the Missouri River, South Dakota and Nebraska [master's thesis]. [Brookings (SD)]: South Dakota State University
- Hay , C H . 2006. Fish and invertebrate abundance in relation to abiotic factors in the Missouri River [dissertation]. [Lincoln (NE)]: University of Nebraska
- Jackson , J K , Battle , J M and Sweeney , B W . 2009 . Monitoring the health of large rivers with macroinvertebrates: do dominant taxa help or hinder the assessment? . River Research and Applications , 26 : 931 – 947 .
- Johnson , B R , Stribling , J B , Flotemersch , J E and Paul , M J . 2006 . “ Benthic macro-invertebrates ” . In Concepts and approaches for the bioassessment of non-wadeable streams and rivers. Cincinnati (OH): US Environmental Protection Agency [Report 600-R-06-127]. Edited by: Flotemersch , J E , Stribling , J B and Paul , M J . 6-1 – 6-26 .
- Lenat , D R . 1983 . Chironomid taxa richness: natural variation and use in pollution assessment . Freshwater Invertebrate Biology , 2 : 192 – 198 . doi: 10.2307/1467151
- McCord , S B and Lambrecht , P R . 2006 . Seasonal succession in the aquatic insect community of an Ozark stream . Journal of Freshwater Ecology , 21 : 323 – 329 . doi: 10.1080/02705060.2006.9665002
- McCune , B and Grace , J B . 2002 . Analysis of ecological communities: PC-ORD, Version 5 , Gleneden Beach (OR) : MJM Software Design .
- MDNR 2010 . Results of wadeable stream macroinvertebrate surveys [Internet]. Jefferson City (MO): Missouri Department of Natural Resources
- Minshall , G W . 1981 . Structure and temporal variations of the benthic macroinvertebrate community inhabiting Mink Creek, Idaho, USA, a 3rd order Rocky Mountain stream . Journal of Freshwater Ecology , 1 : 13 – 26 . doi: 10.1080/02705060.1981.9664013
- Modde , T C and Schmulbach , J C . 1973 . Seasonal changes in the drift and benthic macroinvertebrates in the unchannelized Missouri River in South Dakota . Proceedings of the South Dakota Academy of Science , 52 : 118 – 126 .
- OEPA 2010 . Reports of wadeable stream studies [Internet]. Columbus (OH): Ohio Environmental Protection Agency. Available from: http://www.epa.state.oh.us/ dsw/document_index/psdindx.aspx
- Poulton , B C , Allert , A L , Echols , K R and Brumbaugh , W G . 2005. Validation of aquatic macroinvertebrate endpoint for assessment of biological condition in the channelized Lower Missouri River [technical report]. [Columbia (MO)]: U.S. Geological Survey
- Poulton , B C , Wildhaber , M L , Charbonneau , C S , Fairchild , J F , Mueller , B G and Schmitt , C J . 2003 . A longitudinal assessment of the aquatic macroinvertebrate community in the channelized Lower Missouri River . Environmental Monitoring and Assessment , 85 : 25 – 53 . doi: 10.1023/A:1023301016001
- Robinson , C T , Minshall , G W and Rushforth , S R . 1990 . Seasonal colonization dynamics of macroinvertebrates in an Idaho stream . Journal of the North American Benthological Society , 9 : 240 – 248 . doi: 10.2307/1467587
- USGS 2010 . River stage and flow data [Internet]. Reston (VA): U.S. Geological Survey. Available from: http://waterdata.usgs.gov/mo/nwis/dv
- Ward , J V . 1989 . The four-dimensional nature of lotic ecosystems . Journal of the North American Benthological Society , 8 : 2 – 8 . doi: 10.2307/1467397
- Way , C M , Burky , A J , Bingham , C R and Miller , A C . 1995 . Substrate roughness, velocity refuges, and macroinvertebrate abundance on artificial substrates in the lower Mississippi River . Journal of the North American Benthological Society , 14 : 510 – 518 . doi: 10.2307/1467537
- Weigel , B M and Dimick , J J . 2011 . Development, validation, and application of a macroinvertebrate-based index of biotic integrity for nonwadeable rivers of Wisconsin . Journal of the North American Benthological Society , 30 : 665 – 679 . doi: 10.1899/10-161.1
- Wessell , K J , Merritt , R W , Wilhelm , J GO , Allan , J D , Cummins , K W and Uzarski , D G . 2008 . Biological evaluation of Michigan's non-wadeable rivers using macroinvertebrates . Aquatic Ecosystem Health , 11 : 335 – 351 . doi: 10.1080/14634980802297729
- Williams , D D . 1980 . Temporal patterns in recolonization of stream benthos . Archives of Hydrobiology , 90 : 56 – 74 .