Abstract
American eel (Anguilla rostrata), a once common species, is now in decline throughout much of its native range in North America. There is little information on the role of American eel in river food webs. A better understanding of the diet and ecological role of American eel will help in the conservation of this important species. During autumn 2009, eel and aquatic invertebrate samples were collected from Hannacroix Creek, a tributary of the Hudson River, in Albany and Greene counties, New York, USA. Eel diet was analyzed by the eel size and time period (day or night). A high proportion of eel stomachs were empty (73%). Eel diets varied among size classes and day and night feeding periods (p = 0.001). Diet overlap was significant between small and medium eels caught both during the day (α = 0.71) and at night (α = 0.84). Nocturnal diet and nocturnal invertebrate samples were similar (α = 0.65), indicating a preference for bottom feeding during the night. Mayfly nymphs were the major prey consumed in each period by all size classes. Among eels that fed, night-feeding eels had the greatest stomach weight (as a percent of total body weight). The swim-bladder parasite, Anguillicoloides crassus, was also observed in eels of all size classes with nearly 50% afflicted.
Introduction
Eel are in decline throughout their range. However, the Hudson River and its contiguous tributaries (those not impeded by impassable fish barriers) still harbor American eels (Anguilla rostrata) at some of the highest densities and range of ages (Machut et al. Citation2007). Consequently, this area affords the opportunity to examine eel ecology, including diet, on a relatively unimpaired water body. Few studies have been conducted on diel feeding patterns of American eel. Studies on the European eel (Anguilla anguilla) in northwestern Spain have reported seasonal variability in feeding and a close association with the ambient water temperature, but observed no relationship between the feeding intensity and time of day (Costas-Dias and Lobon-Cervia 2008). Other studies on European eel, American eel, and short-finned eel noted size-specific differences in feeding patterns (Ezzat and El-Seraffy Citation1977; Lookabaugh and Angermeier Citation1992; Sagar and Glova Citation1998). Feeding ecology of American and European eels has been documented as highest just after sunset and then decreasing steadily until sunrise (Sorensen et al. Citation1986; Schulze et al. Citation2004). Studies on American and European eels have noted that small eels (<40 cm length) rely more heavily on benthic invertebrates, while larger eels (>60 cm length) feed on other, larger prey items such as fish (Lookabaugh and Angermeier Citation1992; Schulze et al. Citation2004). Size-specific variations in diet and ontogeny of American and European eels have been shown to vary by geographic regions (Mancini and Able Citation2005, Costas-Dias and Lobon-Cervia 2008).
The primary objective of this study was to examine ontogenetic and diel differences in American eel feeding ecology. Specifically, we examined (1) the differences in the diet composition of three size classes of American eel, (2) whether there was a difference in the diet composition of day- versus night-feeding eels, and (3) whether the feeding intensity varied between the nocturnal and diurnal time periods. This study also afforded an opportunity to examine the incidence of swim-bladder parasite (A. crassus) infestation. Little is known about the effects of A. crassus on eel feeding ecology.
Methods
Hannacroix Creek is a third-order tributary to the Hudson River . Our study site was located 1600 m upstream from the tributary confluence and was approximately 200 m in length. Small (0–175 mm total length), medium (176–300 mm total length), and large (>300 mm total length) American eels were collected during autumn 2009. Collections were made during day and night periods approximately 1 week apart. The daytime collections were made between 11:00 am and 2:00 pm and night-time collections were made between 9:30 pm and 12:00 am. Eels were captured by electrofishing and immediately euthanized in a solution of 13.2 mL clove oil to 1 L ethanol. Upon completion of electrofishing the study area, eels were transferred to 10% buffered formalin for the preservation of stomach contents. The eels were stored in formalin for approximately 6 months until the stomachs were extracted. In order to assess prey availability, five drift nets (91 × 61 × 30 cm3 frame, 1000 µm Nitex mesh) were deployed just upstream of the sampling segment prior to electrofishing. Five Surber samples (bottom fauna) were also collected at the downstream end of the sampling segment prior to electrofishing. Drift nets and Surber samples were taken during both day and night sampling periods.
Figure 1. Sample location in Hannacroix Creek, Albany and Greene counties, NY, USA.
Individual American eels were measured (mm) and weighed (g) in the laboratory. Eel stomachs (including esophagus, stomach, mid-, and hind-guts) were removed and weighed separately (with contents) to account for an individual eel stomach weight, then stored in labeled vials of ethanol until the contents could be processed. Stomach contents were extracted and classified to the lowest practical taxonomic level, usually family (McCafferty Citation1983; Peckarsky et al. Citation1990; Merritt et al. Citation2008). Feeding intensity was calculated as the ratio of the amount of food in the stomach to the total weight of the eel. The content of the drift nets and Surber samples were stored in 70% ethanol upon collection and then classified in a similar manner as the stomach contents. Emptied stomachs were again weighed to determine the weight of stomach content by difference. Swim bladders were also inspected and the number of A. crassus parasites was tallied for each eel.
A diet overlap analysis using the equation of Horn (1966) was applied to diet composition to compare feeding patterns among diel periods and eel sizes. Overlap values of ≥0.60 are considered biologically significant (Zaret and Rand Citation1971). We used a one-way analysis of variance (ANOVA) to test the differences in feeding intensity. When differences were significant, we used Tukey's pairwise comparison test to determine which size classes differed significantly in feeding intensity (Statistix 8.0, 2003, Analytical software, Tallahassee, Florida). Strauss’ (Citation1979) Linear Food Selection Index was used to quantify diel prey selection by eels in each size category. A significance level of α = 0.05 was used for all comparisons. We estimated the probability (P) of eel feeding at night (as opposed to during daylight) with respect to the stomach weight as a percentage of body weight (S) using binary logistic regression (Ott and Longnecker Citation2001) according to the following formula:
Binary logistic regression coefficients for the log odds ratio intercept and regression parameters were computed using Statistix 8.0. Cohen's κ value was calculated as an index of inter-rater reliability.
Results
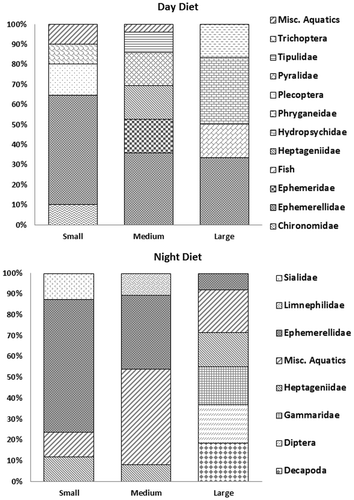
A total of 189 eels were analyzed during the study: 52 small eels (25 night, 27 day), 64 medium eels (35 night, 29 day), and 73 large eels (37 night, 36 day). Small eels measured 68–166 mm in length, with 28 individuals having food in their stomach; medium eels measured 176–300 mm, with 23 individuals having food in their stomachs; and the large eels measured 301–591 mm, with 14 individuals having food in their stomach. Ephemerellidae comprised 39% (p ≤ 0.01) of the night samples and 44% (p < 0.01) of the day samples. Ephemerellid mayflies were the most widely consumed prey at any time, for any size eel, occurring in 63% of the eel stomachs that contained food and contributing 7.7–55.8% of biomass to the diet.
Diet composition differed among different size classes of eel between day and night samples . The contribution of ephemerellids in the diet declined with the eel size and exceeded 50% for small eels during both day and night samples. Fish and decapods were only found in the stomachs of large eels; only one eel had consumed a fish . American eels with empty stomachs were observed during the day (71.7%) and night (67%). Small eels had the highest incidence of food in their stomach (53.8%).
Figure 2. Percent diet composition (based on dry weights) of American eels during the day and night in Hannacroix Creek, New York. (Only items with 8% or greater composition are represented, the remaining items are summed to represent miscellaneous aquatics.)
The diets of the three size classes of American eel were more similar to the composition of the Surber samples than the drift samples . The diets of all size classes of eels were more similar to the composition of benthic fauna samples at night than during the day. Feeding intensity was significantly greater for both small (p = 0.046) and large eels (p = 0.024) at night than during the day. Medium eels showed no significant difference in diel feeding intensity (p = 0.073). Diet overlap was greatest between small and medium eels during both the day (α = 0.71) and night (α = 0.84; ). The predicted probability of night-time feeding computed from binary logistic regression showed that night-feeding American eels had the greatest stomach weight (as a percent of total body weight; ). The regression coefficient β 1 = −40.3692 was significant (p = 0.0006). Cohen's κ for feeding intensity was 0.20. Prey selection indices were generated from Surber samples only and showed no strong selection (+0.01) toward a particular organism. Moderate avoidance was exhibited toward trichopterans (−0.23).
Figure 3. Probability that feeding (total stomach weight as a % of total body weight) occurs during the night.
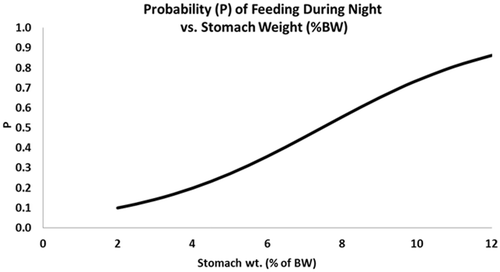
Table 1. Diet overlap of available prey items (drift and Surber) and actual stomach contents. (Diet overlap is a measure of affinity over periods of time, 0 = least, 1 = greatest.)
Table 2. Diet overlap comparisons between each eel size category and day and night time sampling periods.
The parasitic nematode, A. crassus, was observed in eels of all size classes with an infestation rate of 49.7%. The average intensity of parasite infection was 3.2 parasites per infected eel (range: 1–17). Large eels exhibited a significantly higher (p < 0.001) incidence of infection (68.5%) than medium eels (40.6%) or small eels (34.6%). Two or more swim-bladder parasites were present in 30.2% of the infected eels. Feeding intensity was significantly less (p = 0.011) in eels with two or more parasites present than those with zero or one parasite present. American eel length was not correlated with the intensity of parasite infection in this study (p < 0.05).
Discussion
We found that small and large American eels in Hannacroix creek had a greater overall stomach weight and fed most actively at night on the bottom, which is consistent with studies that have regarded eels as nocturnal predators (Sorenson et al. 1986) feeding on a variety of vertebrates and macroinvertebrates (Tesch Citation1977). Medium eels did not show a strong tendency toward day or night feeding, but the comparison was nearly significant (p = 0.073) and a larger sample size may reveal a difference. Eel diet consisted mainly of ephemeropterans, primarily ephemerellids that were present in the diet during both day and night. Previous investigations also found ephemerellids to be a major component of American eel diet (Lookabaugh and Angermeier Citation1992; Denoncourt and Stauffer Citation1993).
Large eels showed a preference for larger, harder bodied prey items including decapods (20%) and gammarid amphipods (20%), but fish were rare in the diet. Although not inventoried, several species of fish were caught during sampling and would be available for consumption, including longnose dace (Rhinichthys cataractae), creek chub (Semotilus atromaculatus), cutlips minnow (Exoglossum maxillingua), and fallfish (Semotilus corporalis). The relative absence of vertebrates in the diet of large eels in Hannacroix Creek was unexpected because larger eels are capable of consuming vertebrates and gaining a bioenergetic advantage by consuming larger prey items (Lookabaugh and Angermeier Citation1992). The drift and Surber samples allowed us to determine the relative importance of benthic versus drift feeding by American eel. Past investigations on short-finned and long-finned eel have shown that eel diets are more closely associated with the benthos (Jellyman Citation1989), but there are no studies documenting the role of drift feeding in American eel diet. In Hannacroix Creek, the diet of all sizes of American eel was more closely associated with the benthos than with the drift, and overlap was higher at night.
We recorded a slightly higher prevalence of A. crassus (49.7%) in American eel swim bladders than the 35.7% reported by Machut and Limburg (2008). The highest incidence of empty stomachs was observed in large eels caught during the night, which also had the highest incidence of parasite infestation. Previous studies on other species of eel have shown similar incidences of empty stomachs (Jellyman Citation1989; Schulze et al. Citation2004) and a 2003 study of American eels in the Hudson River estuary found that the average intensity of A. crassus infection in eels increased over a 3-year time period (Morrison and Secor Citation2003). There may be a threshold at which parasite infection affects feeding, but it is unclear whether size plays a significant role.
Our findings expand on other studies of American eel diet and emphasize the importance of understanding ontogenetic and diel differences in eel feeding ecology in small streams. However, further studies on the effect of eel size in conjunction with day and night feeding periods on diet composition may provide useful information. Possible options for future studies include incorporating a seasonal component to the diet study. This study on juvenile eels in Hannacroix Creek, New York, contributes some new insight on feeding preferences and early life history of American eels, which may be useful for developing conservation plans in areas where eel population is declining and restoration efforts are under consideration.
Acknowledgments
We thank Mike Slattery for his assistance with GIS maps, and J. Ruggirello, M. Chalupnicki, and C. Nack for their assistance with field sampling. H.G. Ketola and M. Chalupnicki provided assistance with statistical analysis. Any use of trade, product, or firm names is for descriptive purpose only and does not imply endorsement of the U.S. Government. This article is contribution 1701 of the US Geological Survey Great Lakes Science Center.
Related Research Data
References
- Costas-Dias , S and Lobon-Cervia , J . 2008 . Diel feeding activity and intensity in the European eel Anguilla anguilla (L.) during an annual cycle in a Cantabrian stream . Knowledge and Management of Aquatic Ecosystems , 1 : 390 – 391 .
- Denoncourt , C E and Stauffer Jr , J R . 1993 . Feeding selectivity of the American eel Anguilla rostrata (LeSueur) in the Upper Delaware River . American Midland Naturalist , 129 : 301 – 308 . doi: 10.2307/2426511
- Ezzat , A E and El-Seraffy , S S . 1977 . Food of Anguilla anguilla in Lake Manzalah, Egypt . Marine Biology , 41 : 287 – 291 . doi: 10.1007/BF00394917
- Horn , H S . 1966 . Measurement of ‘overlap’ in comparative ecological studies . The American Naturalist , 100 : 419 – 424 . doi: 10.1086/282436
- Jellyman , D J . 1989 . Diet of two species of freshwater eel (Anguilla spp.) in Lake Pounui, New Zealand . New Zealand Journal of Marine and Freshwater Research , 23 : 1 – 10 . doi: 10.1080/00288330.1989.9516334
- Lookabaugh , P S and Angermeier , P L . 1992 . Diet patterns of American eel, Anguilla rostrata, in the James River drainage, Virginia . Journal of Freshwater Ecology , 7 ( 4 ) : 425 – 431 . doi: 10.1080/02705060.1992.9664712
- Machut , L S and Limburg , K E . 2008 . Anguillicola crassus infection in Anguilla rostrata from small tributaries of the Hudson River watershed, New York, USA . Diseases of Aquatic Organisms , 79 : 37 – 45 . doi: 10.3354/dao01901
- Machut , L S , Limburg , K E , Schmidt , R E and Dittman , D . 2007 . Anthropogenic impacts on American eel demographics in Hudson River tributaries, New York . Transactions of the American Fisheries Society , 136 : 1699 – 1713 . doi: 10.1577/T06-140.1
- Mancini , F T and Able , K W . 2005. Food habits of young-of-the-year estuarine fishes in the middle Atlantic bight: a synthesis. Technical Report No. 2005-15, Rutgers University
- McCafferty , W P . 1983 . Aquatic entomology: the fisherman's and ecologists’ illustrated guide to insects and their relatives , 1st , Sudbury (MA) : Jones and Bartlett .
- Merritt , R W , Cummins , K W and Berg , M B . 2008 . An introduction to the aquatic insects of North America , 4th , Dubuque (IA) : Kendall Hunt Publishing .
- Morrison , W E and Secor , D H . 2003 . Demographic attributes of yellow-phase American eels (Anguilla rostrata) in the Hudson River estuary . Canadian Journal of Fisheries and Aquatic Sciences , 60 : 1487 – 1501 . doi: 10.1139/f03-129
- Ott , R L and Longnecker , M . 2001 . An introduction to statistical methods and data analysis , 5th , Pacific Grove (CA) : Duxbury Press .
- Peckarsky , B L , Fraissinet , P R , Penton , M A and Conklin Jr , D J . 1990 . Freshwater macroinvertebrates of northeastern North America , Ithaca (NY) : Cornell University Press .
- Sagar , P M and Glova , G J . 1998 . Diel feeding and prey selection of three size classes of shortfinned eel (Anguilla australis) in New Zealand . Marine and Freshwater Research , 49 ( 5 ) : 421 – 428 . doi: 10.1071/MF97154
- Schulze , T , Kahl , U , Radke , R J and Benndorf , J . 2004 . Consumption, abundance and habitat use of Anguilla anguilla in a mesotrophic reservoir . Journal of Fish Biology , 65 : 1543 – 1562 . doi: 10.1111/j.0022-1112.2004.00565.x
- Sorensen , P W , Bianchini , M L and Winn , H W . 1986 . Diel foraging activity of American eels Anguilla rostrata (Lesueur), in a Rhode Island estuary . Fishery Bulletin , 84 : 3
- Strauss , R E . 1979 . Reliability estimates for Ivlev's electivity index, the forage ratio, and a proposed linear index of food selection . Transactions of the American Fisheries Society , 108 : 344 – 352 .
- Tesch , F W . 1977 . The eel: biology and management of anguillid eels , London (UK) : Chapman and Hall Limited .
- Zaret , T M and Rand , A S . 1971 . Competition in tropical stream fishes: supports for competitive exclusion principle . Ecology , 52 : 336 – 342 . doi: 10.2307/1934593