Abstract
The relationships between temporal patterns of macroinvertebrate community and water physicochemical parameters were investigated in the Xiangxi River, which is the largest tributary to the Three Gorges Reservoir (TGR) in Hubei province, China. The construction of the TGR has strongly impacted the Xiangxi River watershed, but inter-annual patterns in stream macroinvertebrate communities have not been examined. We sampled water and macroinvertebrates monthly from 2004 to 2006. We assessed the persistence (similarity between years of species assemblage) and stability (similarity between years with respect to relative abundance of species in the community) of the macroinvertebrate community by the Bray–Curtis distance. The relationship between changes in water physicochemical conditions and changes in community structure were assessed with linear regression. The results showed that both the physical and chemical conditions of the water differed significantly among years. Similarly, the macroinvertebrate community structure and composition were significantly different among years. The compositional persistence was positively and significantly related to the physical condition, supporting the Habitat Templet Theory and indicating the importance of physical habitat protection. This study provides information for environmental assessment, which can be used to develop sustainable management strategies.
Introduction
In response to increasing population growth and consumption, anthropogenic demands placed on freshwater systems continue to escalate (Mcintosh et al. Citation2008). Recently, anthropogenic river modification has become both widespread and intensive (Bona et al. Citation2008). Aquatic ecologists have been interested in the effects of disturbances on macroinvertebrate communities (Bradt et al. Citation1999) because of their importance for assessment and management of disturbances. The Habitat Templet Theory (Southwood 1977, 1988) suggests that the habitat can act as a filter, determining the make-up of the community. Nevertheless, it is difficult for stream ecologists to quantify and explain the temporal patterns of macroinvertebrate communities in the face of environmental change (Hildrew and Giller Citation1994).
It is well documented that communities are spatially and temporally variable (Dutilleul and Legendre Citation1993; Hastings et al. Citation1993), and this concept was the central assumption within the Habitat Templet Theory. Investigations of temporal variability in complex natural systems have been limited due to the lack of adequate statistics to quantify and compare variability across communities, each comprising many species (Micheli et al. Citation1999). However, it is often critical to quantify temporal variation in the relative abundance of species in a community (Collins et al. Citation2000). It is a challenge for stream ecologists to determine the extent to which biological communities are structured by temporal variability (Palmer and Poff Citation1997).
Temporal patterns displayed by communities are often characterized using the concepts of stability and persistence (Collier Citation2008). Stability is defined as the degree of constancy in numbers, or relative abundance of organisms, whereas persistence refers to the continued presence of particular species in a community (Holling Citation1973; Connell and Sousa Citation1983; Scarsbrook Citation2002; Milner et al. Citation2006; Collier Citation2008). Construction of the Three Gorges Reservoir (TGR) led to anthropogenic perturbations in local streams and a changed the stream environment. Therefore, we examined the impact of the changes to the physicochemical conditions on inter-annual patterns in the stability and persistence of stream macroinvertebrate communities. Our hypothesis was that water physicochemical parameters and macroinvertebrate communities would differ among years.
Previous studies have shown that, in stream macroinvertebrate communities the persistence and stability are typically highest where environmental conditions are relatively constant (Robinson et al. Citation2000) or where conditions change slowly over long periods of time (Woodward et al. Citation2002). However, relationships between changes in environmental conditions and changes in community stability and persistence are relatively complex. Scarsbrook et al. (Citation2002) studied macroinvertebrate communities in New Zealand over a nine-year period and showed that persistence and stability were significantly related to the flow conditions, while there was no significant response to water quality changes. Brown et al. (Citation2006) showed that in alpine streams water quality variables (water temperature and suspended sediment concentration) were more important to stability than water quantity because of taxa possessing adaptations to flow variability.
It is now generally recognized that environmental changes can affect the community structure (relative abundance), but more extreme disturbance events are often necessary to cause changes to their presence/absence (Milner et al. Citation2006). Different types of disturbances selectively decrease certain taxa (Swanson et al. Citation1998). If disturbances selectively decrease rare species, then changes in the composition will be greater than that in the structure because measures of community persistence are greatly influenced by rare species (Robinson et al. Citation2000). Other studies showed that disturbance, a key factor in determining the community structure, may have little impact on taxa richness, but may shift the dominant taxa (Reice Citation1994). Immediately after a disturbance event, an opportunistic, rapidly reproducing community may dominate, while during recovery and until the next disturbance better interspecific competitors may dominate (Wootton et al. Citation1996). Additionally, harsh disturbance conditions may provide a habitat template that favors low-diversity macroinvertebrate communities that are highly persistent over time (Winterbourn Citation1997; Collier Citation2008).
Methods
Study area
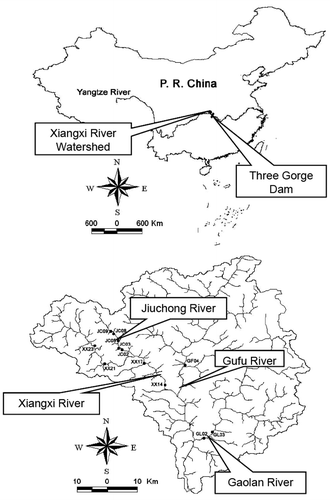
The study was conducted in the Xiangxi River, which is the largest tributary of the TGR in Hubei province, China. The TGR construction has strongly affected the Xiangxi River watershed (Zhou et al. Citation2008; Wu et al. Citation2009). The Xiangxi River is a sixth-order stream that originates from Mount Shennongjia (3, 150 m a.s.l.) and discharges into the Yangtze River. It has three main tributaries: the Jiuchong River, the Gufu River, and the Gaolan River (). Its watershed has an area of 3099 km2 and a natural fall of 1540 m from its headwaters to its confluence with the Yangtze River (Tang et al. Citation2002; Jiang et al. Citation2005; Qu et al. Citation2005). The average annual precipitation within this watershed is 988 mm (Li et al. Citation2010). Forests are mainly distributed on the upper regions of hillsides in this watershed. Urban areas are mainly distributed in the middle and lower regions of the basin near the river bank. We selected 12 sites in this watershed for monthly sampling from 2004 to 2006 ().
Figure 1. Location of the Xiangxi River watershed in China and the distribution of sampling sites.
Macroinvertebrate sampling
One to three random sampling locations were established (30 × 30 cm2 quadrats) at each site for benthic macroinvertebrate measurements. All stones within the Surber sampler frame (mesh = 420 µm) were scrubbed with a soft brush to remove the attached organisms. The river bed was sampled to a depth of approximately 10 cm in areas of unconsolidated substrata. Macroinvertebrates were separated from the sand and mud by hand under a magnifying lens, and preserved in 10% formalin. The biological samples were identified to the lowest feasible taxonomic level according to taxonomic references (Morse et al. Citation1994; Epler Citation2001). Most of the insect, gastropod, oligochaeta, and crustacea were identified to the genus or species level. The nematomorpha, arachnida, and hirudinea were identified to the class level.
Stream physicochemical variables
We used a Hydrolab Minisonde™ (Hach Co., Loveland, Colorado, USA) to measurein situ variables that included pH, conductivity, salinity, total dissolved solids (TDS), and water temperature at each site where benthic macroinvertebrates were captured. We also measured water depth, water width, and current velocity (LJD-10 water current meter, Chongqing Hydrological).
Surface water samples also were collected in two 380 mL, cleaned plastic containers to measure chemical variables according to the standard methods in the laboratory (Huang Citation1999; Cai Citation2007), including total nitrogen (TN), ammonium (NH4–N), nitrate (NO3–N), total phosphorus (TP), orthophosphate (PO4–P), hardness, calcium (Ca2+), chloride (Cl−), alkalinity (Alk), silicon (SiO2), and chemical oxygen demand (COD).
Data analysis
Assessments of stability and persistence must be carried out at an appropriate temporal scale, with the minimum time period being at least one complete turnover of individuals in population (Connell and Sousa Citation1983); hence, an annual time scale has been widely used for determining the community stability. Changes in water physical conditions (water temperature, salinity, TDS, conductivity, water width, water depth, velocity) and chemical conditions (TN, NH4–N, NO3–N, TP, PO4–P, Ca2+, Cl−, hardness, Alk, COD, SiO2, pH) from year to year were estimated for each site by calculating a Euclidean distance. Prior to calculation, the data were normalized by the maximum value of each variable. This resulted in each variable having values between 0 and 1. Then physicochemical data were analyzed using non-metric multidimensional scaling (NMS) carried out in PC-ORD (McCune and Mefford Citation1997). We also used multi-response permutation procedures (MRPP) to test the significance of differences among years of each year group.
Macroinvertebrate data were expressed both in terms of relative abundance and presence/absence. The Bray–Curtis (BC) distance (Bray and Curtis Citation1957) was calculated for each pairing of successive years at each site. This measurement is commonly used to compare the faunal similarity in stream communities and is particularly useful in multivariate analyses (Scarsbrook Citation2002). Macroinvertebrate data were also summarized using NMS and then MRPP was used to test the significance of between-year differences of each year group. We also calculated the macroinvertebrate community diversity indexes.
The indicator value method (IndVal; Dufrêne and Legendre Citation1997) was used to detect how strongly each species discriminated among the NMS groups. The indicator values range from 0 (no indication) to 100 (perfect indication) and the indicator value attained its maximum value when all members of a taxon occurred at all sites within a single group. The Monte Carlo randomization procedure with 1000 permutations was used to test the significance of the indicator value for each species. The relationship between changes in environmental conditions (Euclidean distance for both physical and chemical variables) and changes in community structure (bray-Curtis distance) were assessed with simple linear regression.
Results
Variability in environmental variables
The physical and chemical conditions significantly changed over the study period as we predicted. The physical Euclidean distance at the 12 study sites covered a wide range, both for the years 2004–2005 (average distance = 1.000) and 2005–2006 (average distance = 0.585). Inter-annual NMS ordinations of 2004–2005 and 2005–2006 showed that sites were well separated on axis 1 (a). MRPP analysis indicated significant differences in physical parameters among years (p < 0.05). Axis 1 of the NMS was strongly correlated (p < 0.05) with conductivity (r 2 = 0.84) and salinity (r 2 = 0.79), and Axis 2 was strongly correlated (p < 0.05) with the water width (r 2 = 0.81).
Figure 2. NMS ordination of the physical condition (a) and chemical condition (b) at 12 sites over three years. Ordination stress, final instability, and number of iterations were (respectively): (a) 15.23, 0.00162, 43; (b) 16.82, 0.00461, 39. Legends in the figure are in the format: year (last 2 digits only)-site.
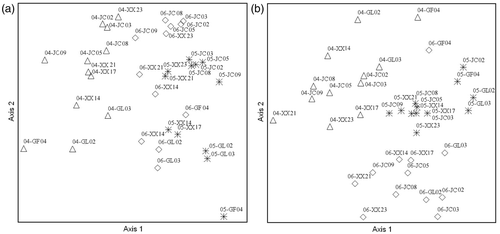
The chemical Euclidean distance of the 12 study sites also covered a wide range for both 2004–2005 (average distance = 1.242) and 2005–2006 (average distance = 1.099). Inter-annual NMS ordinations for 2004–2005 sites were well separated on axis 1 and well separated on axis 2 for 2005–2006 sites (b). MRPP analysis indicated significant differences in chemical conditions among years (p < 0.05). Axis 1 of the NMS was strongly correlated with NO3–N (r 2 = 0.73; p < 0.05) and TN (r 2 = 0.72; p < 0.05), whereas Axis 2 was strongly correlated with pH (r 2 = 0.70; p < 0.05).
Macroinvertebrate community stability and persistence
The BC distance is a measure of dissimilarity with values ranging from 0 to 1; 0 denotes identical samples and higher values denote lower compositional stability (Milner et al. Citation2006). Our average BC distance calculated by relative abundance at 12 sites was 0.657 from 2004 to 2005, and 0.596 from 2005 to 2006. Inter-annual NMS ordinations for 2004–2005 and 2005–2006 showed sites were well separated on axis 1 (a). MRPP analysis indicated significant differences in the macroinvertebrate community structure among years (p < 0.05). Indicator value analysis showed that EPT species accounted for 38.5% and Diptera accounted for 40.2% of the significant indicator species. We also used presence/absence data for NMS ordination. The inter-annual result for 2004–2005 and 2005–2006 showed that sites were well separated on axis 1 (b). MRPP analysis indicated significant differences in macroinvertebrate community composition among years (p < 0.05). The macroinvertebrate community diversity indices are presented in .
Figure 3. NMS ordination of macroinvertebrate relative abundance (a) and presence/absence (b) at 12 sites over three years. Ordination stress, final instability, and number of iterations were (respectively): (a) 18.54, 0.00332, 28; (b) 15.46, 0.00401, 37. Legends in the figure are in the format: year (last 2 digits only)-site.
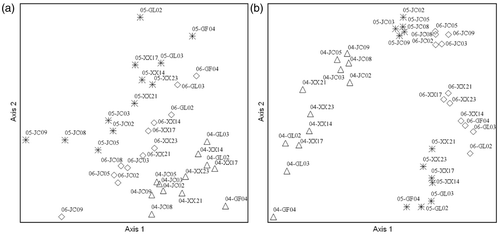
Table 1. Means (standard deviation) of macroinvertebrate community diversity at different sites on the Xiangxi River during 2002–2006.
Regression analysis
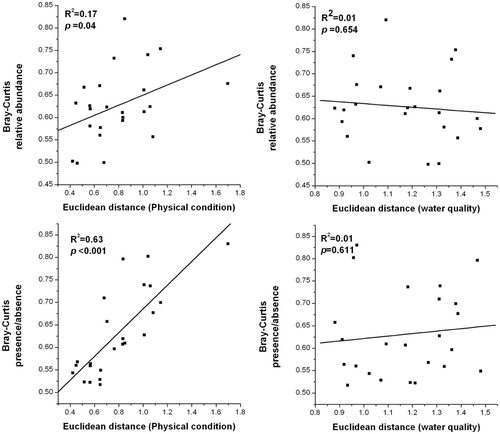
The relationship of community stability and persistence with physicochemical habitat variables showed that changes in the physical condition had a strong, positive and significant effect on community persistence (p < 0.05; R 2 = 0.63; ). Community stability and persistence had no significant relationship with chemical conditions of the water (p > 0.05; ). We can generate a prediction that community species composition differs along with the physical environment change over time.
Figure 4. Relationship between the inter-annual change in a community composition (relative abundance and presence/absence data) and changes in both water quality and physical condition. p values are given. Refer to individual F-tests for the significance of the slope of best-fit line.
Discussion
The stability of a biotic community is the tendency of a community's composition to remain constant (Pielou Citation1974). Coefficients of variation (CVs) have been used in numerous previous studies, examining spatial and temporal variation of the stream macroinvertebrate community (e.g., Minshall and Robinson Citation1998; Robinson et al. Citation2000). Along with developing multivariate statistical techniques, numerous investigations have used the BC distance to study the changes of macroinvertebrate community spatially and temporally (e.g., Scarsbrook Citation2002; Milner et al. Citation2006). Therefore, we used the BC distances of relative abundance and presence/absence data to assess the changes of community structure and composition.
Community persistence has a significant relationship with stability (Milner et al. Citation2006), but communities showed greater change in the community structure (relative abundance) than in the composition (presence/absence) under the varied physical and chemical conditions over three years in our study. Previous studies also showed that the community structure had changed more than the composition in the face of disturbance (e.g., Boulton et al. Citation1992; Scarsbrook Citation2002). Species persistence and assemblage stability might vary considerably depending on habitat diversity (Liefferinge et al. Citation2010). In streams of the Denali National Park, Alaska, USA, community persistence showed a significant relationship with winter snowfall; stability was significantly related to the Pfankuch Index of channel stability (Milner et al. Citation2006). Differences in the assemblage structure among wilderness streams were attributed to the stream size (shift in shredder assemblages) and the temperature (shift in plecopteran taxa) in Idaho, USA (Robinson et al. Citation2000). In our research, chemical and physical conditions changed significantly from 2004 to 2006, affecting the macroinvertebrate community. Significantly different community structures were mainly attributed to the changes in Ephemeroptera, Plecoptera, and Trichoptera taxa, and Diptera species. Robinson et al. (Citation2000) also suggested that the abundance of macroinvertebrate can change quickly in response to short-term fluctuations in environmental conditions and season, whereas the assemblage composition may be maintained.
The relationships between inter-annual patterns in the stability and persistence and physicochemical habitat variables were complex. In simple regression, community stability had no significant relationship with the chemical and physical conditions of the water. This is to be expected, as macroinvertebrate abundances can change quickly with response to food availability (Richards and Minshall Citation1988) or life history dynamics (Robinson et al. Citation1992). Fortunately, we found that compositional persistence was positively and significantly related to the physical condition. This was similar to other studies (e.g., Weatherly and Omerod Citation1990; Scarsbrook Citation2002) supporting the Habitat Templet Theory. Modifications in the environment result in the movement of species that use different stream habitats to complete their life cycles (Silva et al. Citation2010). In our study, changes in the physical condition might cause the macroinvertebrate drift to increase, leading to the observed differences in community composition
Persistence represents the ability of a community to return to some previous state following perturbation. Changes in the physical condition might make it difficult for the community persistence to recover, indicating the importance of physical habitat protection. A longer time record may provide a different view of persistence, especially in the context of low-frequency environmental events (Watson and Estes Citation2011). Hence, further research is necessary to address the potential relationship between the community structure and environmental change over the long term, which could help develop sustainable management strategies.
Acknowledgments
We thank Daofeng Li, Tao Tang, Naicheng Wu, Fengqin Li, Xinghuan Jia, and Shugui Duan for their assistance with the field sampling. We also thank Ruiqiu Liu for her assistance in the measurement of water quality. Special thanks to professor Kyle D. Hoagland and Mr Mathias Kuemmerlen for their constructive comments on this manuscript and their help with English. This work was funded by the National Natural Science Foundation of China (30330140), the Major Science and Technology Program for Water Pollution Control and Treatment (2008ZX07526-002-07) and Special Funds of Ministry of Water Resources of China for Scientific Research on Public Interest (No. 201101001).
Related Research Data
References
- Bona , F , Falasco , E , Fenoglio , S , Iorio , L and Badino , G . 2008 . Response of macroinvertebrate and diatom communities to human-induced physical alteration in mountain streams . River Research and Applications , 24 : 1068 – 1081 . doi: 10.1002/rra.1110
- Boulton , A J , Petersen , G G , Grimm , N B and Fisher , S G . 1992 . Stability of an aquatic macroinvertebrate community in a multiyear hydrologic disturbance regime . Ecology , 73 : 2192 – 2207 . doi: 10.2307/1941467
- Bradt , P , Urban , M , Goodman , N , Bissell , S and Spiegel , I . 1999 . Stability and resilience in benthic macroinvertebrate assemblages . Hydrobiologia , 403 : 123 – 133 . doi: 10.1023/A:1003711928980
- Bray , J R and Curtis , J T . 1957 . An ordination of the upland forest communities of southern Wisconsin . Ecological Monographs , 27 : 325 – 349 . doi: 10.2307/1942268
- Brown , L E , Milner , A M and Hannah , D M . 2006 . Stability and persistence of alpine stream macroinvertebrate communities and the role of physicochemical habitat variables . Hydrobiologia , 560 : 159 – 173 . doi: 10.1007/s10750-005-1260-8
- Cai , Q (ed) . 2007 . Protocols for standard observation and measurement in aquatic ecosystems , Beijing : Chinese Environmental Science Press (in Chinese) .
- Collier , K J . 2008 . Temporal patterns in the stability, persistence and condition of stream macroinvertebrate communities: relationships with catchment land-use and regional climate . Freshwater Biology , 53 : 603 – 616 . doi: 10.1111/j.1365-2427.2007.01923.x
- Collins , S L , Micheli , F and Hartt , L . 2000 . A method to determine rates and patterns of variability in ecological communities . Oikos , 91 : 285 – 293 . doi: 10.1034/j.1600-0706.2000.910209.x
- Connell , J H and Sousa , W P . 1983 . On the evidence needed to judge ecological stability or persistence . American Naturalist , 121 : 789 – 824 . doi: 10.1086/284105
- Dufrêne , M and Legendre , P . 1997 . Species assemblages and indicator species: the need for a flexible asymmetrical approach . Ecological Monographs , 67 : 345 – 366 .
- Dutilleul , P and Legendre , P . 1993 . Spatial heterogeneity against heteroscedasticity: an ecological paradigm versus a statistical concept . Oikos , 66 : 152 – 171 . doi: 10.2307/3545210
- Epler , J H . 2001 . Identification manual for the larval chironomidae (Diptera) of North and South Carolina , Special Publication SJ2001-SP13. Raleigh (NC) : North Carolina Department of Environment and Natural Resource; alatka (FL): St. . Johns River Water Management District. p. 70–97
- Hastings , A , Hom , C L , Ellner , S , Turchin , P and Godfray , H CJ . 1993 . Chaos in ecology: is mother nature a strange attractor? . Annual Review Ecology Systematics , 24 : 1 – 33 .
- Hildrew , A G and Giller , P S . 1994 . “ Patchiness, species interactions and disturbance in the stream benthos ” . In Aquatic ecology: scale, pattern and process , Edited by: Giller , P S , Hildrew , A G and Raffaelli , D G . 21 – 62 . London : Blackwell .
- Holling , C S . 1973 . Resilience and stability of ecological systems . Annual Review of Ecology and Systematics , 4 : 1 – 23 . doi: 10.1146/annurev.es.04.110173.000245
- Huang , X . 1999 . Survey, observation and analysis of lake ecology , Beijing : China Standards Press (in Chinese) .
- Jiang , M , Deng , H , Cai , Q and Wu , G . 2005 . Species richness in a riparian plant community along the banks of the Xiangxi River, the Three Gorges region . The International Journal of Sustainable Development and World Ecology , 12 ( 1 ) : 60 – 67 . doi: 10.1080/13504500509469619
- Li , F , Cai , Q and Ye , L . 2010 . Developing a benthic index of biological integrity and some relationship to environmental factors in the subtropical Xiangxi River, China . International Review of Hydrobiology , 95 ( 2 ) : 171 – 189 . doi: 10.1002/iroh.200911212
- Liefferinge , C H , Simoens , I , Vogt , C , Cox , T JS , Breine , J , Ercken , D , Goethals , P , Belpaire , C and Meire , P . 2010 . Impact of habitat diversity on the sampling effort required for the assessment of river fish communities and IBI . Hydrobiologia , 644 : 169 – 183 . doi: 10.1007/s10750-010-0110-5
- McCune , B and Mefford , M J . 1997 . PC-ORD. Multivariate analyses of ecological data Gleneden Beach (OR): MjM Software
- Mcintosh , M D , Schmitz , J A , Benbow , M E and Burky , A J . 2008 . Structural and functional changes of tropical riffle macroinvertebrate communities associated with stream flow withdrawal . River Research and Applications , 24 : 1045 – 1055 . doi: 10.1002/rra.1104
- Micheli , F , Cottingham , K L , Bascompte , J , Bjornstad , O N , Eckert , G L , Fischer , J M , Keitt , T H , Kendall , B E , Klug , J L and Rusak , J A . 1999 . The dual nature of community variability . Oikos , 85 : 161 – 169 . doi: 10.2307/3546802
- Milner , A M , Conn , S C and Brown , L E . 2006 . Persistence and stability of macroinvertebrate communities in streams of Denali National Park, Alaska: implications for biological monitoring . Freshwater Biology , 51 : 373 – 387 . doi: 10.1111/j.1365-2427.2005.01488.x
- Minshall , G W and Robinson , C T . 1998 . Macroinvertebrate community structure in relation to measures of habitat heterogeneity . Archiv fur Hydrobiologie , 141 : 129 – 151 .
- Morse , J C , Yang , L and Tian , L . 1994 . Aquatic insect of china useful for monitoring water quantity , Nanjing : Hehai University Press .
- Palmer , M A and Poff , N L . 1997 . The influence of environmental heterogeneity on patterns and processes in streams . Journal of the North American Benthological Society , 16 : 169 – 173 . doi: 10.2307/1468249
- Pielou , E C . 1974 . Population and community ecology: principles and methods , New York : Gordon and Breach .
- Qu , X , Tang , T , Xie , Z , Liu , R , Li , D and Cai , Q . 2005 . Distribution of macroinvertebrates in the Xiangxi River System and their relationship with environmental factors . Journal of Freshwater Ecology , 20 ( 2 ) : 233 – 238 . doi: 10.1080/02705060.2005.9664962
- Reice , S R . 1994 . Nonequilibrium determinants of biological community structure . American Scientist , 82 : 424 – 435 .
- Richards , C and Minshall , G W . 1988 . The influence of periphyton abundance on Baetis bicaudatus distribution and colonization in a small stream . Journal of the North American Benthological Society , 7 : 77 – 86 . doi: 10.2307/1467913
- Robinson , C T , Minshall , G W and Royer , T V . 2000 . Inter-annual patterns in macroinvertebrate communities of wilderness streams in Idaho, USA . Hydrobiologia , 421 : 187 – 198 . doi: 10.1023/A:1003956114073
- Robinson , C T , Reed , L R and Minshall , G W . 1992 . Influence of flow regime on life history, production, and genetic structure of Baetis bicaudatus (Ephemeroptera) and Hesperoperla pacifica (Plecoptera) . Journal of the North American Benthological Society , 11 : 278 – 289 . doi: 10.2307/1467648
- Scarsbrook , M R . 2002 . Persistence and stability of lotic invertebrate communities in New Zealand . Freshwater Biology , 47 : 417 – 431 . doi: 10.1046/j.1365-2427.2002.00810.x
- Silva , F R , Ferreira , E JG and Deus , C P . 2010 . Structure and dynamics of stream fish communities in the flood zone of the lower Purus River, Amazonas State, Brazil . Hydrobiologia , 651 : 279 – 289 . doi: 10.1007/s10750-010-0307-7
- Southwood , T RE . 1977 . Habitat, the templet for ecological strategies . Journal of Animal Ecology , 46 : 337 – 365 . doi: 10.2307/3817
- Southwood , T RE . 1988 . Tactics, strategies and templets . Oikos , 52 : 3 – 18 . doi: 10.2307/3565974
- Swanson , F J , Johnson , S L , Gregory , S V and Acker , S A . 1998 . Flood disturbance in a forested mountain landscape . BioScience , 48 ( 9 ) : 681 – 689 . doi: 10.2307/1313331
- Tang , T , Cai , Q , Liu , R , Li , D and Xie , Z . 2002 . Distribution of epilithic algae in the Xiangxi River system and their relationships with environmental factors . Journal of Freshwater Ecology , 17 ( 3 ) : 345 – 352 . doi: 10.1080/02705060.2002.9663907
- Watson , J and Estes , J A . 2011 . Stability, resilience, and phase shifts in rocky subtidal communities along the west coast of Vancouver Island, Canada . Ecological Monographs , 81 ( 2 ) : 215 – 239 . doi: 10.1890/10-0262.1
- Weatherly , N S and Omerod , S J . 1990 . The constancy of invertebrate assemblages in soft-water streams: implications for the prediction and detection of environmental change . Journal of Applied Ecology , 27 : 952 – 964 . doi: 10.2307/2404389
- Winterbourn , M J . 1997 . “ New Zealand mountain stream communities: stable yet disturbed? ” . In Evolutionary ecology of freshwater animals , Edited by: Streit , B , Stadler , T and Lively , C M . 32 – 54 . Basel (Switzerland) : Birkhauser Verlag .
- Woodward , G , Jones , J I and Hildrew , A G . 2002 . Community persistence in Broadstone Stream (UK) over three decades . Freshwater Biology , 47 : 1419 – 1435 . doi: 10.1046/j.1365-2427.2002.00872.x
- Wooten , J T , Parker , M S and Power , M E . 1996 . Effects of disturbance on river food webs . Science , 273 : 1558 – 1561 . doi: 10.1126/science.273.5281.1558
- Wu , N , Tang , T , Zhou , S , Jia , X , Li , D , Liu , R and Cai , Q . 2009 . Changes in benthic algal communities following construction of a run-of-river dam . Journal of the North American Benthological Society , 28 ( 1 ) : 69 – 79 . doi: 10.1899/08-047.1
- Zhou , S , Tang , T , Wu , N , Fu , X and Cai , Q . 2008 . Impacts of a small dam on riverine zooplankton . International Review of Hydrobiology , 93 ( 3 ) : 297 – 311 . doi: 10.1002/iroh.200711038