Abstract
We used logistic regression to model the persistence of two imperiled pelagic-spawning cyprinids, sharpnose shiner Notropis oxyrhynchus and smalleye shiner N. buccula, endemic to the Brazos and Wichita Rivers, Texas, USA. There was a significant positive relationship between persistence and river fragment length. Mean length of river fragments in which sharpnose shiner or smalleye shiner still occur was significantly greater (599 ± 27 km) than the mean length of fragments from which these species were absent (187 ± 20 km). Persistence of both species appears to depend on the presence of river fragments long enough to allow spawned ova and swim-up fry time to develop and move out of the current into the backwater areas.
Introduction
Over half of the world's rivers have been fragmented as a result of human activities, particularly the construction of impoundments (Nilsson et al. Citation2005). The proportion of streams and rivers impacted by impoundments within the contiguous United States is substantially greater (85%, Hughes et al. Citation2005) and has contributed to the imperilment of a number of North American freshwater and diadromous fishes (Jelks et al. Citation2008), including several species of the Great Plains fishes (e.g., Winston et al. Citation1991; Luttrell et al. Citation1999; Wilde and Ostrand Citation1999; Bonner and Wilde Citation2000; Hoagstrom et al. Citation2011). Perkin and Gido (Citation2011) modeled the relationship between persistence of eight species of Great Plains pelagic-spawning cyprinids and the length of river fragments and concluded that river fragment length explained 71% of cumulative extirpations among these species.
The sharpnose shiner Notropis oxyrhynchus and smalleye shiner N. buccula are endemic to the Brazos River drainage in Texas, USA. Historically, both species ranged widely in the Brazos River main stem and its upper tributaries, from just off the escarpment of the Llano Estacado, downstream to within 100 km of the Gulf Coast. Notropis oxyrhynchus also occurred in the North and South Wichita rivers (Cross et al. Citation1986; Moss and Mayes Citation1993) but it is now generally believed to be extirpated from the Wichita River drainage. Both the Brazos and Wichita River drainages have been extensively modified by impoundments. The Brazos River is the most fragmented river basin in Texas. Currently, there are 3 main stem impoundments and nearly 40 tributary impoundments in the Brazos River drainage and an additional 8 impoundments are permitted or are under construction. Although the Wichita River drainage is smaller than the Brazos River drainage, it is fragmented by two main stem impoundments and at least 8 tributary impoundments.
As a result of the modifications of the Brazos and Wichita rivers, the distributions of both N. oxyrhynchus and N. buccula have decreased substantially. Both species were listed as Candidate Species under the Endangered Species Act (Federal Register Citation2002) and are now being considered for listing as threatened or endangered species. Because of the extensive fragmentation of the Brazos River drainage basin, and the likelihood of continued fragmentation as proposed and ongoing water development projects are completed (e.g., Figure ), the conservation of the N. oxyrhynchus and N. buccula necessitates an understanding of the length of the river fragment that is necessary to maintain populations of these species. Herein we present an analysis of the persistence of N. oxyrhynchus and N. buccula as a function of river fragment length following the general methods of Perkin and Gido (Citation2011).
Figure 1 Map showing the location of Wichita River (fragments 1 and 2) and Brazos River fragments (3–7) studied herein. The arrow above fragment 3 indicates the location of the one dam currently permitted for construction on the Brazos River and its upper tributaries
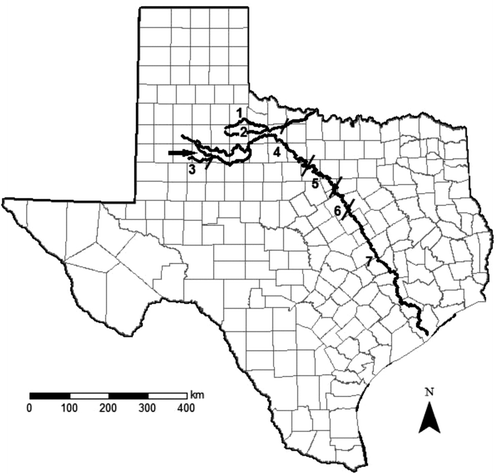
Methods
We obtained river fragment lengths (km) for the Brazos River using the US Geological Survey National Hydrography Database and ArcGIS® 10 software (Table ). We determined fragment lengths downstream from Lake Whitney, the most downstream impoundment on the Brazos River main stem, as the distances from Lake Whitney to the site of the most downstream record for N. buccula (Mussel Shoals, near Bryan, Texas) and N. oxyrhynchus (at US Hwy 90A, Richmond, Texas). N. oxyrhynchus historically occurred in the North and South Wichita rivers (e.g., Moss and Mayes Citation1993), presumably as the result of the capture of a Brazos River headwater stream (Cross et al. Citation1986). Therefore, we assumed these populations represented natural occurrences of the species. Fragment lengths for the North and South forks of the Wichita River were determined as the distances upstream from Lake Kemp Dam to the headwaters of both rivers (North Wichita River = 192 km; South Wichita River = 168 km) and were obtained from The Handbook of Texas Online (Citation2012). Introduced populations of N. oxyrhynchus and N. buccula in the Colorado River, both now presumed to be extirpated, were not included in the analyses presented herein.
Table 1 Persistence of N. oxyrhynchus and N. buccula in fragments of the Wichita and Brazos rivers. Persistence of each species is indicated by a plus (+). Absence of a date of last occurrence indicates the species did not occur in that river fragment
Because the number of river fragments in which N. oxyrhynchus (n = 6) and N. buccula (n = 5) historically occurred is small, we pooled data for both species and included an indicator variable (0 = N. oxyrhynchus, 1 = N. buccula) in all initial statistical analyses. We used a t-test to determine whether river fragment length differed between reaches in which either N. oxyrhynchus or N. buccula was present versus those in which these species were absent. The indicator variable for species was not significant (p = 0.802) and was not included in our final model. We used logistic regression to model persistence (continued presence) of N. oxyrhynchus and N. buccula in each river fragment as a function of fragment length. The indicator variable was not significant (p = 0.966) and was not included in our final model. Because there was no heterogeneity in any fragment in the occurrence of N. oxyrhynchus and N. buccula (i.e., both were either present or absent), we used a penalized likelihood (Firth Citation1993) to assess model adequacy. A nominal value of α = 0.10 was used in all analyses. We conducted all statistical analyses using SAS® version 9.3 software (SAS Institute, Inc., http://www.sas.com/).
Results
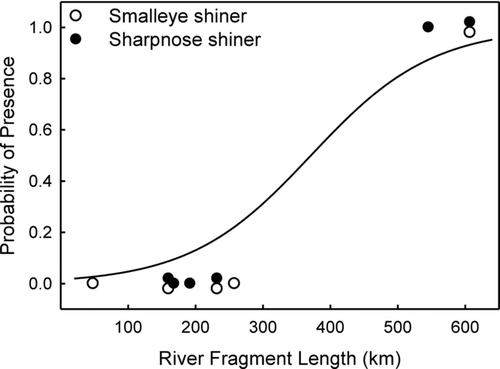
Neither N. oxyrhynchus nor N. buccula occurred in river fragments less than 300 km in length, whereas neither was absent from river fragments greater than 545 km in length. There was a significant difference (t = 11.71; df = 9; p < 0.0001) in the mean length (±SE) of river fragments in which N. oxyrhynchus or N. buccula occurred (599 ± 27 km) compared with the mean length (187 ± 20 km) of river fragments from which these species were absent.
Logistic regression showed persistence of N. oxyrhynchus and N. buccula was significantly related to river fragment length (p < 0.004, n = 11, r 2 = 0.532) according to the equation:
Figure 2 Probability of persistence of sharpnose shiner and smalleye shiner based on river fragment length in the Brazos and Wichita rivers. For three river reaches in which both species are either present or absent, symbols are jiggered vertically to facilitate visualization
Discussion
N. oxyrhynchus and N. buccula are members of a reproductive guild of pelagic-spawning cyprinids common in Great Plains rivers that broadcast spawn semi-buoyant ova into the current (Platania and Altenbach Citation1998). Ova and newly hatched swim-up fry are kept afloat by the current, which prevents them from settling to the river bottom where they may be covered and suffocated by silt and sand sediments (Moore 1944). Platania and Altenbach (Citation1998) calculated that the developing ova could be displaced as far as 72 to 144 km downstream before hatching and that fry could be displaced an additional 216 km downstream before they could move out of the current into backwater areas. Thus, the early life history of members of this guild appears to require long reaches of un-fragmented river. Perkin and Gido (Citation2011) found that stream fragment length was a predictor of the conservation status of several known, or suspected, Great Plains pelagic-spawning cyprinids and observed that mean fragment lengths were least for extirpated (140 ± 55 km) and declining (205 ± 65 km) populations and greatest for stable (425 ± 185 km) populations. Similarly, Dieterman and Galat (Citation2004) determined that the persistence of sicklefin chub Macrhybopsis meeki, a likely member of this reproductive guild, required river fragments of at least 301 km in length. In comparison, N. oxyrhynchus and N. buccula appear to require longer mean river fragments (599 km) for persistence. We note, however, that the Brazos River main stem and upper tributaries consist exclusively of short (<260 km) and long (>620 km) river fragments so it is possible that, given the absence of river reaches of intermediate length, our model might overestimate the minimum fragment needed for persistence.
There is an interaction between river fragment length and the presence and magnitude of river discharge that is important to the persistence of N. oxyrhynchus and N. buccula and, presumably, other Great Plains pelagic-spawning cyprinids. Durham (Citation2007) and Durham and Wilde (Citation2009a) developed population dynamics models for N. oxyrhynchus and N. buccula and determined that an average summer discharge of 2.61 m3s−1 (N. oxyrhynchus) to 5.52 m3s−1 (N. buccula) was necessary to maintain stable populations. Both species spawn throughout a protracted spawning season, May through September, during which at least some fish spawn on any given day; additionally, more synchronized, population-wide spawning events occur in response to pulses in discharge (Durham Citation2007; Durham and Wilde Citation2008). Although spawning occurs daily, or almost so, Durham and Wilde (Citation2009b) found no evidence that N. oxyrhynchus and N. buccula successfully produced young during periods of drought, when the river ceased to flow. Dudley and Platania (Citation2007) have also suggested that, given a river fragment of some length, an upper threshold to discharge may limit populations of broadcast-spawning cyprinids: excessive discharge may displace ova and early life stages into downstream reservoirs, where ova may settle to the lake bottom and suffocate and fry are susceptible to predation by piscivorous fishes. Thus, there is a minimum threshold for reproductive success and, apparently, a maximum threshold above which ova and fry are displaced from the system.
There may be an important interrelationship between river fragment length and habitat suitability (or integrity) that contributed to the persistence, or absence, of N. oxyrhynchus and N. buccula in portions of the Brazos River. Each successive modification of the river, as a result of changes in habitat, flow regime, or the fish assemblage, likely affects the length of river fragment needed to sustain these species. Our model does not account for these potential changes and it would be difficult to assess them retrospectively, but we do acknowledge their possible effects on the persistence of N. oxyrhynchus and N. buccula in various river fragments. Thus, our estimates of river fragment length necessary for persistence of N. oxyrhynchus and N. buccula may be conservative (longer than required).
Dudley and Platania (Citation2007) concluded that neither natural flow regimes nor habitat restoration efforts would be likely to result in long-term recovery of pelagic-spawning cyprinids in the Rio Grande basin, unless fragmented river reaches were reconnected. This is consistent with Perkin and Gido's (Citation2011) results, which showed that persistence of populations of eight species of Great Plains pelagic-spawning cyprinids was negatively correlated with minimum thresholds in fragment length. Perkin and Gido argued this was evidence that fragmentation had played a role in the observed declines in abundance and distribution among species they studied. Our results for N. oxyrhynchus and N. bucculasuggest that the conservation of extant populations of these species in the Brazos River depends on the maintenance of the two remaining long river fragments. Efforts to repatriate the species to other portions of their historic distributions may be unsuccessful unless smaller fragments are reconnected (i.e., dams decommissioned and removed) to form larger, continuous river segments or river discharge within various segments is modified, relative to river fragment length, to support the life-history needs of N. oxyrhynchus and N. buccula.
Notes
aSee Figure for number codes.
bNot recorded from this reach, but the species undoubtedly occurred here. It was presumably extirpated at the same time as was the N. oxyrhynchus.
References
- Bonner , T H and Wilde , G R . 2000 . Changes in the fish assemblage of the Canadian River, Texas associated with reservoir construction . Journal of Freshwater Ecology , 15 : 189 – 198 . doi: 10.1080/02705060.2000.9663736
- Cross , F B , Mayden , R L and Stewart , J D . 1986 . “ Fishes in the western Mississippi Basin (Missouri, Arkansas, and Red Rivers) ” . In The zoogeography of North American freshwater fishes , Edited by: Hocutt , C H and Wiley , E O . 363 – 412 . New York , NY : John Wiley .
- Dieterman , D J and Galat , D L . 2004 . Large-scale factors associated with sicklefin chub distribution in the Missouri and Lower Yellowstone rivers. . Transactions of the American Fisheries Society , 133 : 577 – 587 . doi: 10.1577/T03-002.1
- Dudley , R K and Platania , S P . 2007 . Flow regulation and fragmentation imperil pelagic-spawning riverine fishes . Ecological Applications , 17 : 2074 – 2086 . doi: 10.1890/06-1252.1
- Durham , B W . 2007 . Reproductive ecology, habitat associations, and population dynamics of two imperiled cyprinids in a Great Plains river [dissertation] , Lubbock , TX : Texas Tech University .
- Durham , B W and Wilde , G R . 2008 . Synchronous and asynchronous spawning in a Great Plains cyprinid fish . Ecology of Freshwater Fish , 17 : 528 – 541 . doi: 10.1111/j.1600-0633.2008.00303.x
- Durham , B W and Wilde , G R . 2009a . Population dynamics of smalleye shiner an imperiled cyprinid fish endemic to the Brazos River, Texas . Transactions of the American Fisheries Society , 138 : 666 – 674 . doi: 10.1577/T07-234.1
- Durham , B W and Wilde , G R . 2009b . Effects of streamflow and intermittancy on the reproductive success of two broadcast-spawning cyprinid fishes . Copeia , 2009 : 21 – 28 . doi: 10.1643/CE-07-166
- Federal Register . 2002 . Endangered and threatened wildlife and plants; review of species that are candidates or proposed for listing as endangered or threatened; annual notice of findings on recycled petitions; annual description of progress on listing actions , Vol. 67 , 40657 – 40679 . Washington , DC : Office of the Federal Register, National Archives and Records Administration . 114
- Firth , D . 1993 . Bias reduction of maximum likelihood estimates . Biometrika , 80 : 27 – 38 . doi: 10.1093/biomet/80.1.27
- Hoagstrom , C W , Brooks , J E and Davenport , S R . 2011 . A large-scale conservation perspective considering endemic fishes of the North American plains . Biological Conservation , 144 : 21 – 34 . doi: 10.1016/j.biocon.2010.07.015
- Hughes , R M , Rinne , J N and Calamusso , B . 2005 . “ Introduction to historical changes in large river fish assemblages of the Americas ” . In Historical changes in large river fish assemblages of the Americas , Edited by: Rinne , J N , Hughes , R M and Calamusso , B . 1 – 12 . Bethesda , MD : American Fisheries Society Symposium . 45
- Jelks , H L , Walsh , S J , Burkhead , N M , Conteras-Balderas , S , Diaz-Pardo , E , Hendrickson , D A , Lyons , J , Mandrak , N E , McCormick , F Nelson , J S . 2008 . Conservation status of imperiled North American freshwater and diadromous fishes . Fisheries , 33 : 372 – 407 . doi: 10.1577/1548-8446-33.8.372
- Luttrell , G R , Echelle , A A , Fisher , W L and Eisenhour , D J . 1999 . Declining status of two species of the Macrhybopsis aestivalis complex (Teleostei: Cyprinidae) in the Arkansas River basin and related effects of reservoirs as barriers to dispersal . Copeia , 1999 : 981 – 989 . doi: 10.2307/1447973
- Moore , G A. 1944 . Notes on the early life history of Notropis girardi . Copeia , : 209 – 214 . doi: 10.2307/1438675
- Moss , R E and Mayes , K B . 1993 . Current status of Notropis buccula and Notropis oxyrhynchus in Texas. River Studies Report 8 , Austin , TX : Texas Parks and Wildlife Department .
- Nilsson , C. , Reidy , C A , Dynesius , M and Revenga , C . 2005 . Fragmentation and flow regulation of the world's large river systems . Science , 308 : 405 – 408 . doi: 10.1126/science.1107887
- Perkin , J S and Gido , K B . 2011 . Stream fragmentation thresholds for a reproductive guild of Great Plains fishes . Fisheries , 36 : 371 – 383 . doi: 10.1080/03632415.2011.597666
- Platania , S P and Altenbach , C S . 1998 . Reproductive strategies and egg types of seven Rio Grande Basin cyprinids . Copeia , 1998 : 559 – 569 . doi: 10.2307/1447786
- The Handbook of Texas Online [Internet] . c1999-2012 . Texas Historical Association [cited 2012 Oct 10] Available from http://www.tshaonline.org/handbook/
- Wilde , G R and Ostrand , K G . 1999 . Changes in the fish assemblage of an intermittent prairie stream upstream from a Texas impoundment . Texas Journal of Science , 51 : 203 – 210 .
- Winston , M R , Taylor , C M and Pigg , J . 1991 . Upstream extirpation of four minnow species due to damming of a prairie stream . Transactions of the American Fisheries Society , 120 : 98 – 105. . doi: 10.1577/1548-8659(1991)120<0098:UEOFMS>2.3.CO;2