Abstract
The initial, rapid expansion of the invasive round goby (Neogobius melanostomus) throughout the Great Lakes drainage was largely confined to lentic systems. We recently observed round gobies ascending two tributaries of the St. Lawrence River. The expansion of gobies into small lotic environments may place ecologically similar species at risk. Fantail darter (Etheostoma flabellare) is one of the several benthic species of the New York Great Lakes drainages that are threatened by round goby invasion. We examined the habitat use and diet composition of fantail darters and round gobies in Mullet Creek, a third-order tributary of the St. Lawrence River, NY, USA. The objectives of this study were to determine the degree of habitat and diet overlap between fantail darters and round gobies in a tributary of the St. Lawrence River. Gobies and darters co-occurred at 22% of capture sites. Of the four habitat variables examined (cover, depth, substrate and velocity), only depth use was significantly different with gobies using deeper habitats than darters. Among the two species and size classes sampled (large vs. small), large darters had the most restricted habitat use requirements. There was variation in round goby and darter diet composition, but only moderate diet overlap occurred between fantail darters and round gobies (Cλ = 0.43). Conditions in Mullet Creek were appropriate for the evaluation of possible spatial and dietary competition between round goby and native darters. Early detection and management of round goby invasions is critical to maintaining ecological integrity of lotic ecosystems in the St. Lawrence Valley.
Introduction
The round goby (Neogobius melanostomus) colonized all five of the Great Lakes within five years of their North American introduction into the St. Clair River in 1990 (French and Jude Citation2001). Current observations and estimates suggest that gobies have become a major component of fish assemblages throughout much of the Great Lakes basin (Cooper et al. Citation2007; Johnson et al. Citation2008). Despite the rapid proliferation of the species in lentic habitat, gobies had not populated lotic habitats until recently. Current research indicates that 20 years after their initial introduction, round gobies have colonized the St. Lawrence River and are now ascending and colonizing Great Lakes tributaries (French and Jude Citation2001; Phillips et al. Citation2003; Krakowiak and Pennuto Citation2008). In addition, gobies in drowned river mouths, lakes and wetlands tend to be immigrants from other habitats (i.e. young of year and age 1+ individuals seeking uncolonized environments; Cooper et al. Citation2007). The round goby is a successful colonizer, in part, because of its wide tolerance of various environmental factors, high site fidelity on rocky habitats (Ray and Corkum Citation2001) and opportunistic foraging ability (French and Jude Citation2001; Phillips et al. Citation2003; Carman et al. Citation2006; Krakowiak and Pennuto Citation2008).
Dreissenid mussels are a primary component of the round goby diet. However, most Great Lakes tributaries lack dreissenid mussels and decline in the richness and abundance of native benthic macroinvertebrates (with greatest effects on Ephemeroptera, Plecoptera and Trichoptera species) in streams invaded by round goby have been documented (Krakowiak and Pennuto Citation2008). French and Jude (Citation2001) noted that native darter species (Etheostoma spp. and Percina spp.) were especially susceptible to competition from round goby due to great overlap in diet and habitat use; at lengths <75 mm, gobies, rainbow darter (E. caeruleum) and logperch (Percina caprodes) consumed common macroinvertebrate resources. During the day, round goby chose prey that are abundant, exposed and active (Carman et al. Citation2006). Similarly, Etheostoma darters are visual, diurnal feeders that forage on a variety of benthic invertebrates. Thus, where food resources are limiting, round goby are likely to adversely affect darters and other benthic species where they co-occur. Laboratory studies confirm that St. Lawrence River gobies readily consume non-Dreissenid prey items in northern New York tributaries (Baldwin et al. Citation2011).
The St. Lawrence River and its contiguous tributaries provide unique aquatic habitats within the Great Lakes drainage that are inhabited by some of the rarest darters in New York state, channel darter (P. copelandi), Iowa darter (E. exile) and eastern sand darter (Ammocrypta pellucida). St. Lawrence River goby populations are now plentiful, reproductively active and capable of dispersing during all life stages. We report on some of the earliest observations of round goby invading lotic habitats of St. Lawrence River tributaries also occupied by darters. One would predict that round gobies and fantail darters would utilize similar habitats and food sources within these tributaries (Poos et al. Citation2009). Fantail darters (E. flabellare) were used in this study due to their high abundance in our sample area. Our objectives were to (1) examine the habitat use and diet preference of different size classes of darters and gobies, (2) test if darters and gobies were selecting distinct habitats and food items and (3) measure the degree of habitat and diet overlap occurring between the two species.
Methods
Habitat
Mullet Creek (44.2756° N, 76.0063° W) is a long (16.75 km), third-order tributary in New York State that drains a 73,625 km2, agriculturally rich watershed into the St. Lawrence River. The creek has a low (0.25 m) concrete barrier 225 m upstream of the tributary mouth. We collected fish and habitat data during the day (1000–1300 h) on two separate sampling events in the summer (mid-August to early September) of 2010. Subsets of captured fishes were preserved for diet analysis. Sampling was carried out 11 days apart to allow for darters and gobies to recolonize preferred habitats. We used a spot-sample electrofishing technique to describe fish habitat use (Bovee Citation1986; Johnson Citation2008). A numbered, weighted bobber was placed at each location where a darter or goby was observed. At each bobber the number of each fish species present within each size category (small: ≤49 mm or large: ≥50 mm) was recorded. Depth, velocity, substrate and cover were recorded at each bobber location after shocking was completed. Depth was recorded to the closest 0.5 cm. Velocity was measured using a Marsh–McBirney model 201D digital flow meter at 60% of the depth and recorded to 0.1 m s−1 (Buchanan and Somers Citation1969). Visual estimation using a modified Wentworth particle size scale ranging from detritus (1) to bedrock (7) was used to quantify substrate size. The amount of cover within a radius of approximately 0.50 m for small, and 1.00 m for large fishes was estimated visually. Available habitat was quantified from transects located every 10 m along the 150-m stream reach sampled. Habitat variables were recorded at 10 stations along each transect.
Diet
In the laboratory, round gobies and darters were measured (mm TL ± SE) and weighed (nearest 0.001 g). Stomachs were removed and weighed (nearest 0.001 g) prior to the identification of their contents. Prey items were identified to the lowest possible taxon and enumerated. Diet items were dried to constant weight in an oven at 105 °C; diet composition was quantified based on dry weight for each taxon. Feeding patterns were examined by dividing the wet weight of the full stomach by the wet weight of the entire fish to provide an estimate of feeding intensity.
Statistical analysis
Diet overlap among the species and size classes (small, large) was determined by calculating overlap coefficients (Cλ) using the equation of Morisita (Citation1959), later modified by Horn (Citation1966). The Cλ values range from 0, in which there are no samples in common, to 1 when samples are identical. Because the Shapiro–Wilks test showed that the data were not normally distributed, we used the non-parametric Kruskal–Wallis one-way analysis of variance test to examine the frequency distribution of habitat variables between size categories and species for darters, gobies and available habitat. The Kruskal–Wallis all-pairwize multiple-comparison test was applied when significance was detected. A significance level α = 0.05 was used for all statistical comparisons. Principal component analysis (PCA) was used to examine the similarity of habitat use by darters and gobies, with reference to available habitat.
Results
Habitat
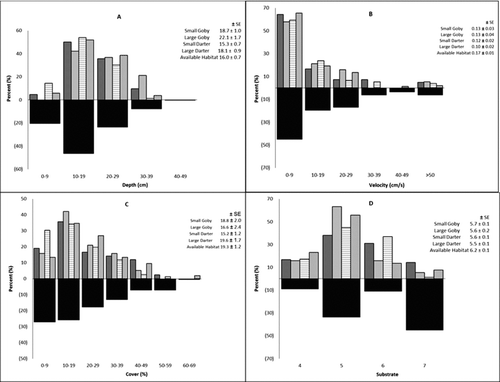
Fish and habitat data were recorded from 132 locations during summer 2010. We collected 128 and 61 habitat points for fantail darters and round gobies, respectively, within our study area (Table ). Two other Etheostoma species were captured in our study area; six tessellated darters (E. olmstedi) and two Johnny darters (E. nigrum). Small and large fantail darters were observed at 59% and 41% of the sites, while small and large gobies were observed at 69% and 31% of the sites. Fantail darters used habitat unaccompanied by round gobies at 75 sites, while round gobies used habitat unoccupied by darters at 28 sites; the two species co-existed at 29 sites (22% overlap in habitat use). Analysis of variance showed large darters occupied regions of slower current velocity compared to available velocities, large gobies occupied deeper areas compared to available depths, and both size classes of the two species were associated with significantly smaller substrate size (or less hardness) than was available within the study reach (Table , Figure ). No statistical significance was found among the means of the available cover variable and fish habitat use. Of the four habitat variables examined (cover, depth, substrate and velocity), only depth was significantly different between round gobies and fantail darters, with gobies using deeper habitats than darters.
Table 1 Number of fantail darters (E. flabellare) and round gobies (N. melanostomus) by size class used to quantify habitat during late summer in Mullet Creek, NY, USA
Table 2 Average habitat values associated with available habitat conditions and fantail darter and round goby collection sites by size category collected during late summer 2010 in Mullet Creek, NY, USA
Figure 1 Utilisation of habitat based on habitat parameter, available habitat parameters, species (round goby and fantail darter) and size category (small and large), during September 2010 in Mullet Creek, Fisher's Landing, New York. (A = depth, B = velocity, C = cover, D = substrate). Small goby ![]()
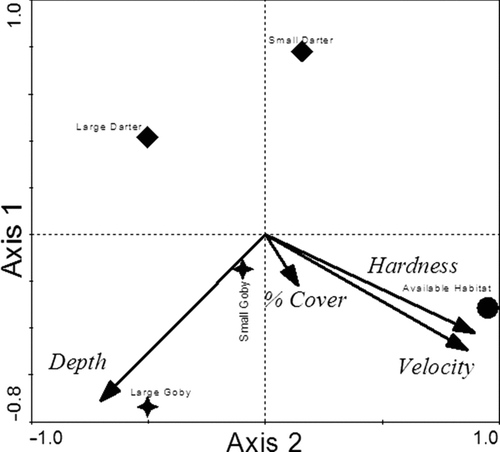
The first two PCA axes accounted for 80.6% of variation in habitat use by the size classes of the two study species (Figure ). The PCA showed that habitat centroids for large darters were most divergent from available habitat centroids, suggesting strong habitat selectivity. Small gobies exhibited the least habitat specificity. Large goby and small darter centroids were equidistant from the available habitat centroid, suggesting an equal degree of habitat specificity.
Figure 2 Principal components analysis ordination gradients and hyperspace centroid locations of species by size class in Mullet Creek, NY
Diet
A total of 49 round goby (17 small, 32 large) and 43 fantail darter (13 small, 30 large) stomachs were examined for diet composition (Table ). Decapods (i.e. crayfish), annelids and fish were the major prey of round goby, comprizing 11.7%, 11.3% and 11.2% of their diet, respectively (Figure ). Small gobies consumed decapods (15.3%), fish (15.3%) and trichopterans (12.6%); and large gobies fed on decapods (13.8%), annelids (13.6%) and fish (13.3%). Species of Limnephilidae and Gammaridae were the major prey of fantail darter, comprizing 11.7% and 11.6% of their diet, respectively. Small fantail darters consumed Gammarus spp. (20.4%), other amphipods (14.5%) and Limnephilidae spp. (12.5%), while large fantail darters consumed Limnephilidae spp. (16.2%), Gammarus spp. (11.5%) and Heptageniidae spp. (11.2%).
Table 3 Number of round goby and fantail darter examined for diet composition, average size and size range by size class in Mullet Creek, September 2010
Figure 3 Taxon of prey items and percentage of diet for round gobies and fantail darters during September 2010 in Mullet Creek, Fisher's Landing, NY. Round goby ![]()
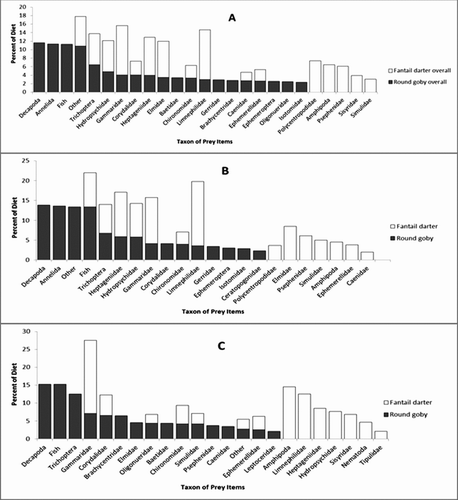
The frequency of specific prey items was noteworthy. Fifty-eight percent of fantail darters analysed for diet content consumed chironomids and heptageniids. Eighty-four percent of round gobies consumed ostracoda and 78% of gobies consumed chironomids. Seven of 49 gobies (6 large, 1 small) consumed larval fishes (of undetermined species due to the degradation of their remains); however, fantail darters and round gobies dominated the stream reach studied and fantail darters were the likely species consumed by gobies if the larval community structure was similar to the adult community.
Moderate diet overlap occurred between fantail darters and round gobies (Cλ = 0.43). There was a strong relationship in diet overlap among different size classes of the same species. Diet overlap between small and large fantail darters was (Cλ = 0.75) and between small and large round gobies was (Cλ = 0.72).
Discussion
The secondary invasion of small lotic habitats by round goby is a recent occurrence in the Great Lakes–St. Lawrence River system. Previous studies indicate native darters (Etheostoma spp., Percina spp. and logperch, P. caprodes) and other benthic, stream-dwelling fishes (e.g. Cottus spp.) are most affected by competition with round gobies (French and Jude Citation2001; Lauer et al. Citation2004; Poos et al. Citation2009), which are aggressive space competitors (Balshine et al. Citation2005) that can reach larger body size than similar species (Charlebois et al. Citation2001). In some systems, Johnny darter and rainbow darter (E. caeruleum) occur only in streams without gobies (Krakowiak and Pennuto Citation2008), and Johnny and channel darters have been eliminated from Lake Erie near-shore fish communities (Reid and Mandrak Citation2008). A study of St. Lawrence River tributaries related to this work showed that round goby was absent from the US tributaries, except for the overlap in Mullet and Tibbetts Creek (McKenna et al. Citation2012).
Researchers have documented that secondary invasions by round goby have been restricted to areas of low species diversity, particularly those lacking a rich benthic fish assemblage (Carman et al. Citation2006). While many lower Great Lakes assemblages are relatively diverse (Poos et al. Citation2009), the section of Mullet Creek that we sampled (200 m upstream of a drowned river mouth) supported a low-diversity assemblage. Fish species richness and habitat diversity were greater historically (Greeley and Greene Citation1935) and upstream of our sample site (McKenna et al. Citation2012); richness may have been greater in the study area before round goby invasion. Fantail darters and round gobies used similar habitats in Mullet Creek and exhibited a nearly identical degree of selectivity. Round gobies prefer riffles (French and Jude Citation2001; Phillips et al. Citation2003; Carman et al. Citation2006), which is also the preferred habitat of most darter species (Mundahl and Ingersoll Citation1983; Ingersoll and Claussen Citation1984). In Mullet Creek, round gobies occupied deeper habitats than darters, and darters used slower current areas than gobies. This could be the result of displacement of darters by gobies. Appropriate goby and darter substrate in Mullet Creek is a limiting habitat feature, because much of the bottom is composed of bedrock. Benthic fishes were restricted to regions of less consolidated substrate and smaller particle size than generally available habitat. Thus, darters and gobies in Mullet Creek overlap within the restricted amount of appropriate habitat. The abundance of round gobies decreased, and the abundance of darters increased upstream, indicating an inverse relationship of abundance. Shallow bedrock areas may be acting as a physical barrier to the upstream progression of gobies as observed by Phillips et al. (Citation2003), and corroborated in our study; in Mullet Creek, gobies were not captured upstream of a series of bedrock shelves that were downstream of a small (6″ above water level) concrete barrier. We document a period of active invasion of Mullet Creek by round goby and displacement of native darters may become more severe as colonization proceeds.
Our findings of the diversity of round goby diet composition and habitat preference in third-order streams demonstrate their opportunistic feeding nature and habitat use in a St. Lawrence River tributary. Round gobies prefer to feed on molluscs (Andraso et al. Citation2011), but will adjust their diet based on depth, substrate and prey availability (French and Jude Citation2001; Phillips et al. Citation2003; Carman et al. Citation2006; Johnson et al. Citation2008). The moderate diet overlap between fantail darters and round gobies (Cλ = 0.43) in this study shows the potential for competition where these species are sympatric. There was also a strong relationship in diet overlap among different size classes of species. We did not measure food resource limitation, so competition is unknown, but diet overlap provides a measure of the potential for competition. Round gobies had a wider diet breadth than fantail darter, with larval fish, decapods, annelids and several families of aquatic insects unique to the goby diet. The wider diet breadth of round gobies is likely due to their larger mouth gape (Ray and Corkum Citation1997; Hayden and Miner Citation2009), allowing round gobies to consume larger prey than darters. This diet flexibility has aided the round goby in its invasion of Great Lakes tributary habitats. Predation by gobies has also been observed on numerous species of benthic fishes, including greenside darter (E. blennoides) and rainbow darter; fish <30 mm were most susceptible (Poos et al. Citation2009). In Mullet Creek, fish remains in gobies stomachs were likely young of year and 1+ fantail darters based on their relative abundance compared to other species.
This study reports the first documented invasion of lotic tributaries of the St. Lawrence River by round goby and potential competition for habitat and food with native species in tributary habitat. Invasive species are a significant threat to freshwater biodiversity (Dopazo et al. Citation2008), and these results emphasize the importance of protecting unique tributary habitats that contain native species from invasion by round goby. Early detection and prevention of species invasion are paramount for maintaining the ecological integrity of uninvaded habitats (Poulos et al. Citation2012). Long-distance dispersal (over 50 km) of goby from founder populations in Lakes Erie and St. Clair, suggests that dispersal and upstream colonization of inland tributaries is likely to continue (Poos et al. Citation2009). Our findings show that conditions are appropriate for possible spatial and dietary competition between round goby and darters in Mullet Creek, which is similar to many tributaries of the St. Lawrence River due to geology, gradient and land use within their watersheds. If competitive exclusion occurs, we would expect darter abundance to decline. As round goby expands its range within the St. Lawrence Valley, this situation could be repeated in any of the accessible tributaries. Round goby movement was limited by small obstructions and such low-head barriers might help protect tributary habitats from invasion.
Acknowledgements
We thank Marc Chalupnicki for his assistance with statistical analysis, and Mike Slattery for his expert assistance with geographical information systems (GIS). Any use of trade, product, or firm names is for descriptive purpose only and does not imply endorsement of the U.S. Government. This article is contribution 1756 of the US Geological Survey Great Lakes Science Center.
The work of James H. Johnson, James E. McKenna, Jr. and Dawn E. Dittman was authored as part of their official duties as an Employee of the United States Government and is therefore a work of the United States Government. In accordance with 17 USC. 105, no copyright protection is available for such works under US Law.
Ross Abbett and Emily M. Waldt hereby waive their right to assert copyright, but not their right to be named as co-authors in the article.
References
- Andraso , G , Cowles , J , Colt , R , Patel , J and Campbell , M . 2011 . Ontogenetic changes in pharyngeal morphology correlate with a diet shift from arthropods to dreissenid mussels in round gobies (Neogobius melanostomus) . J Great Lakes Res. , 37 : 738 – 743 . doi: 10.1016/j.jglr.2011.07.011
- Baldwin , B S , Carpenter , M , Rury , K and Woodward , E . 2011 . Low dissolved ions may limit secondary invasion of inland waters by exotic round gobies and Dreissenid mussels in North America . Biol Invasions. , 14 ( 6 ) : 1157 – 1175 . doi: 10.1007/s10530-011-0146-0
- Balshine , S , Verma , A , Chant , V and Theysmeyer , T . 2005 . Competitive interactions between round gobies and logperch . J Great Lakes Res. , 31 : 68 – 77 . doi: 10.1016/S0380-1330(05)70238-0
- Bovee , K D . 1986 . “ Development and evaluation of habitat suitability criteria for use in instream flow incremental methodology ” . In Instream Flow Information Paper No. 21 , Washington , DC : U.S. Fish and Wildlife Service . Biology Report 86 (7)
- Buchanan , T J and Somers , W P . 1969 . Discharge measurements at gauging stations. Techniques of Water Resources Investigations, Book 3 , Washington, , DC : US Geological Survey .
- Carman , S M , Janssen , J , Jude , D J and Berg , M B . 2006 . Diel interactions between prey behavior and feeding in an invasive fish, the round goby, in a North American river . Freshwater Biol. , 51 : 742 – 755 . doi: 10.1111/j.1365-2427.2006.01527.x
- Charlebois , P M , Corkum , L D , Jude , D J and Knight , C . 2001 . The round goby (Neogobius melanostomus) invasion: current research and future needs . J Great Lakes Res. , 27 : 263 – 266 . doi: 10.1016/S0380-1330(01)70641-7
- Cooper , M J , Ruetz , C R , Uzarski , D G and Burton , T M . 2007 . Distribution of round gobies in coastal areas of Lake Michigan: are wetlands resistant to invasion . J Great Lakes Res. , 33 : 303 – 313 . doi: 10.3394/0380-1330(2007)33[303:DORGIC]2.0.CO;2
- Dopazo , S N , Corkum , L D and Mandrak , N E . 2008 . Fish assemblages and environmental variables associated with Gobiids in nearshore areas of the lower Great Lakes . J Great Lakes Res. , 34 : 450 – 460 . doi: 10.3394/0380-1330(2008)34[450:FAAEVA]2.0.CO;2
- French , J R and Jude , D J . 2001 . Diets and diet overlap of nonindigenous gobies and small benthic native fishes co-inhabiting the St. Clair River, Michigan . J Great Lakes Res. , 27 : 300 – 311 . doi: 10.1016/S0380-1330(01)70645-4
- Greeley , John R and Greene , C W . 1935 . “ Fishes of the watershed with annotated list ” . In A biological survey of the Oswegatchie and Black River systems , 77 – 92 . New York State Conservation Dept. . Suppl. 21st Annual Report 1931.
- Hayden , T A and Miner , J G . 2009 . Rapid dispersal and establishment of a benthic Ponto-Caspian goby in Lake Erie: diel vertical migration of early juvenile round goby . Biol Invasions. , 11 : 1767 – 1776 . doi: 10.1007/s10530-008-9356-5
- Horn , H S . 1966 . Measurement of “overlap” in comparative ecological studies . Am Nat , 100 : 419 – 424 . doi: 10.1086/282436
- Ingersoll , C G and Claussen , D L . 1984 . Temperature selection and critical thermal maxima of the fantail darter, Etheostoma flabellare, and Johnny darter, E. nigrum, related to habitat and season . Environ Biol Fishes , 2 ( 2 ) : 131 – 138 . doi: 10.1007/BF00002262
- Johnson , J H . 2008 . Seasonal habitat use of brook trout and juvenile Atlantic salmon in a tributary of Lake Ontario . Northeast Nat. , 15 : 363 – 374 . doi: 10.1656/1092-6194-15.3.363
- Johnson , J H , McKenna , J E Jr. , Nack , C C and Chalupnicki , M A . 2008 . Diel diet composition and feeding activity of round goby in the nearshore region of Lake Ontario . J Fresh Ecol. , 23 : 607 – 612 . doi: 10.1080/02705060.2008.9664248
- Lauer , T J , Allen , P J and McComish , T S . 2004 . Changes in mottled sculpin and Johnny darter trawl catches after the appearance of round gobies in the Indiana waters of Lake Michigan . Trans Am Fish Soc. , 133 : 185 – 189 . doi: 10.1577/T02-123
- Krakowiak , P J and Pennuto , C M . 2008 . Fish and macroinvertebrate communities in tributary streams of eastern Lake Erie with and without round gobies (Neogobius melanostomus, Pallas 1814) . J Great Lakes Res , 34 : 675 – 689 .
- McKenna , J E Jr , David , A , Johnson , J H and Dittman , D E . 2012 . “ Evaluation of threatened, endangered, and declining species of the St. Lawrence River and its tributaries ” . In Final report to the fish enhancement, mitigation and research fund projects # 2005-0129-010 and #2005-0129-009 13045 New York , , USA Available from: US Fish and Wildlife Service, Ecological Services Office, 3817 Luker Road, Cortland
- Morisita , M. 1959 . Measuring of interspecific association and similarity between communities . Mem Fac Sci Kyushu Univ Ser E , 3 : 65 – 80 .
- Mundahl , N D and Ingersoll , C G . 1983 . Early autumn movements and densities of Johnny (Etheostoma nigrum) and fantail (E. flabellare) darters in a southwestern Ohio stream . Ohio J Sci , 83 ( 3 ) : 103 – 108 .
- Phillips , E C , Washek , M E , Hertel , A W and Niebel , B N . 2003 . The round goby (Neogobius melanostomus) in Pennsylvania tributary streams of Lake Erie . J Great Lakes Res. , 29 : 34 – 40 . doi: 10.1016/S0380-1330(03)70413-4
- Poos , M , Dextrase , A J , Schwalb , A N and Ackerman , J D . 2009 . Secondary invasion of the round goby into high diversity Great Lakes tributaries and species at risk hotspots: potential new concerns for endangered freshwater species . Biol Invasions , 12 : 1269 – 1284 . doi: 10.1007/s10530-009-9545-x
- Poulos , H M , Chernoff , B , Fuller , P L and Butman , D . 2012 . Ensemble forecasting of potential habitat for three invasive fishes . Aquat Invasions , 7 : 59 – 72 . doi: 10.3391/ai.2012.7.1.007
- Ray , W J and Corkum , L D . 1997 . Predation of zebra mussels by round gobies (Neogobius melanostomus) . Environ Biol Fish. , 50 : 267 – 273 . doi: 10.1023/A:1007379220052
- Ray , W J and Corkum , L D . 2001 . Habitat and site affinity of the round goby . J Great Lakes Res. , 27 : 329 – 334 . doi: 10.1016/S0380-1330(01)70648-X
- Reid , S M and Mandrak , N E . 2008 . Historical changes in the distribution of threatened channel darter (Percina copelandi) in Lake Erie with general observations on the beach fish assemblage . J Great Lakes Res. , 34 ( 2 ) : 324 – 333 . doi: 10.3394/0380-1330(2008)34[324:HCITDO]2.0.CO;2