Abstract
Habitat associations of sharpnose shiner (Notropis oxyrhynchus) in the upper Brazos River, USA, were studied over a two-year period. We used generalized linear models with a logit link to assess the influence of individual physical and chemical habitat variables on the presence of sharpnose shiner at the microhabitat scale. Our models indicated that depth was the only habitat variable that significantly predicted sharpnose shiner presence during the first year of the study and that current velocity was the only variable that significantly predicted sharpnose shiner presence during the second year. Monthly plots of fitted regression estimates superimposed over frequency distributions for both significant variables revealed a distinct pattern of habitat use related to habitat availability. When only a narrow range of depths and current velocities was available, sharpnose shiner occurred more frequently in deeper and swifter microhabitats. In contrast, during periods with the greatest depths and current velocities, sharpnose shiner presence decreased with increasing depth and current velocity. Our results suggest that sharpnose shiner in the Brazos River exhibits habitat preferences but these preferences are conditional, based on the range of habitats available on a given date.
Introduction
The need to secure water supplies for municipal, agricultural, and industrial purposes has led to the anthropogenic modification of most of the world's major river systems (Stanford and Ward Citation1979; Dynesius and Nilsson Citation1994). Historically free-flowing from source to outlet, most rivers now comprise a series of fragmented reaches bounded by reservoirs or other water diversion structures (Kingsford Citation2000; Nilsson et al. Citation2005; Perkin and Gido Citation2011; CitationWilde and Urbanczyk Forthcoming). The alterations of stream and river environments frequently have resulted in changes in fish community structure and the imperilment of obligate riverine species (Cross and Moss Citation1987; Winston et al. Citation1991; Aparicio et al. Citation2000; Bonner and Wilde Citation2000; Jelks et al. Citation2008). These changes have been most profound in semi-arid and arid regions of the world, where water resources are naturally scarce and availability is variable (Agnew and Anderson Citation1992; Kingsford Citation2000).
In the past century, modification of the Great Plains rivers resulted in changes in fish assemblages that have been ascribed to changes in turbidity, predation, reproductive failure, and loss of specific microhabitats (Cross and Moss Citation1987; Pflieger and Grace Citation1987). The importance of microhabitat availability and use in determining the structure and maintenance of stream fish communities in highly variable environments has been a topic of much debate (Baker and Ross Citation1981; Grossman et al. Citation1982; Paine et al. Citation1982; Grossman et al. Citation1985), but there is a general recognition of the importance of fish-habitat associations in the maintenance of historical patterns of species distribution and abundance (Rosenfeld Citation2003). Nevertheless, specific patterns in habitat use have been difficult to identify for some species and systems because of inconsistencies in results among studies. For example, based on the univariate and bivariate habitat characteristics, Matthews and Hill (Citation1980) and Polivka (Citation1999) found evidence of nonrandom selection of microhabitats by the dominant cyprinid species in the South Canadian River, Oklahoma, but observed little consistency in the types of habitats used by these species. In contrast, Kehmeier et al. (Citation2007) reported that small-bodied cyprinids were nonrandom in their use of various habitat types in the Pecos River, New Mexico, but that there was substantial overlap among these habitats in physical and chemical characteristics. It remains unclear if fishes that inhabit the Great Plains streams and rivers have specific habitat requirements or preferences that, as yet, have not been definitively identified or if the inconsistent evidence for specific habitat associations reflects a generalist strategy among these species for coping with life in extremely variable environments.
Modification of the Brazos River began in 1941 with the construction of Possum Kingdom reservoir (Anderson et al. Citation1983). Since then, the Brazos River basin has been extensively developed. Currently, there are 3 mainstem impoundments, nearly 40 tributary impoundments in the Brazos River drainage, and an additional 8 impoundments are permitted or are under construction. These modifications have been associated with decreases in distribution and abundance of several native fish species (Anderson et al. Citation1983; Moss and Mayes Citation1993; Anderson et al. Citation1995; Wilde and Ostrand Citation1999) including the sharpnose shiner, Notropis oxyrhynchus, which was listed as a candidate species under the Endangered Species Act (Federal Register Citation2002) and is now being considered for listing as a threatened or endangered species. Little is known of the ecology of sharpnose shiner, particularly of its habitat preferences and requirements. Herein, we assess the relationship between the presence of sharpnose shiner and several habitat variables in the Brazos River. Our results contribute to an understanding of the biology of this imperiled species and provide an insight into local patterns of fish-habitat associations in an arid-land river.
Methods
We conducted our field sampling within an 800 m reach of the Brazos River located in the City of Seymour, Baylor County, TX (33°34′09″ N, 99°16′1″ W). Three characteristics of this stream reach made it ideal for this study. First, it is within the historic and current range of sharpnose shiner (Moss and Mayes Citation1993). Second, although discharge in this reach of the river is variable, this reach seldom becomes intermittent as do reaches further upstream. The periods of intermittency restrict fishes to isolated pools, preventing them from moving freely and precluding any meaningful study on habitat use. Third, the fish assemblage within this reach has been affected little by upstream impoundments (Wilde and Ostrand Citation1999; Ostrand and Wilde Citation2002).
We sampled sharpnose shiner monthly from April 2003 through March 2004 and April 2004 through March 2005, herein referred to as 2003 and 2004 samples, respectively. On each sampling date, we made 50 seine hauls along a series of transects that were set perpendicular to the main channel. The transects were spaced 20 m apart. Within each transect, nonoverlapping seine hauls covering a distance of 5 m were made over homogenous habitat using a 3.7 × 1.8 m seine (5 mm mesh). We identified, enumerated, and released the collected fishes. During the summer months, May through September, we separately enumerated adult and juvenile fish. We measured water depth (cm), current velocity (m/s), temperature (°C), conductivity (μS/cm), dissolved oxygen (mg/L), and turbidity (nephelometric turbidity units, NTU) at the midpoint of the area covered by each seine haul.
We used a generalized linear model with a logit link to model the relationship between the presence of sharpnose shiner (>20 mm in total length) and the physical and chemical habitat variables. We included month in each model as a categorical variable to account for predictable seasonal variation in habitat variables. The importance of individual habitat variables within sampling dates was then assessed, given that month was already included in the model. If the variation explained by a habitat variable was not significant (α = 0.05), that variable was omitted from further analysis. For habitat variables that explained a significant proportion of variation in sharpnose shiner presence, the model was expanded to include a quadratic term and an interaction term between the habitat variable and month. We used Akaike information criterion (AIC) to select the most appropriate model for variables with multiple significant terms (Akaike Citation1973). To assess patterns of habitat use, plots of the resulting fitted models were superimposed over monthly plots of frequency distributions for each significant habitat variable.
Results
The generalized linear models that related sharpnose shiner presence to depth, current velocity, temperature, conductivity, dissolved oxygen, and turbidity, given that month was included in the model, indicated that depth (p = 0.007) was the only habitat variable that significantly predicted sharpnose shiner presence in 2003 (Table ). Similarly, current velocity (p = 0.036) was the only habitat variable that significantly predicted sharpnose shiner presence in 2004. In both cases, there was a positive relationship between these variables and sharpnose shiner presence.
Table 1 Chi-square and significance values for generalized linear models in 2003 for six physical and chemical habitat variables used to predict sharpnose shiner presence in the Brazos River, TX, USA in 2003 (n = 425) and 2004 (n = 549)
The expanded (full) models for sharpnose shiner presence and depth in 2003 showed there was a significant interaction between depth and month. Likewise, the full model for current velocity in 2004 showed there was a significant interaction between current velocity and month (Table ). Thus, the specific relationship between sharpnose shiner presence and depth (2003) and current velocity (2004) differed among months. This suggests that habitat use by sharpnose shiner was dynamic: sharpnose shiner presence did not depend on specific depths or current velocities, but was contingent on conditions that existed on a given date. Model selection based on AIC indicated that, in the case of depth (2003) and current velocity (2004), the full model was the best model for predicting presence of sharpnose shiner (Table ). The full model for depth in 2003 explained 43% of the total variation in sharpnose shiner presence. The full model for current velocity in 2004 explained 42% of the total variation in sharpnose shiner presence.
Table 2 Model selection for nested generalized linear models for significant habitat variables for sharpnose shiner in the Brazos River, TX, USA
The plots of the fitted model predictions superimposed on frequency histograms for depth (2003) and current velocity (2004) showed temporal variation in sharpnose shiner presence as related to these habitat variables. In 2003, when habitats <35 cm in depth were available, the generalized linear model had a positive slope, indicating that sharpnose shiner preferred deeper areas. However, when a broader range of depths was available, models had a negative slope, indicating that fish avoided the deepest areas on these dates (Figure ). The changes in available current velocities from month to month during 2004 also appeared to affect habitat use by sharpnose shiner in a similar way. In 2004, sharpnose shiner exhibited an apparent preference for current velocities of approximately 0.35 m/s in months where the majority of available current velocities were less than 0.50 m/s (Figure ). In months with the greatest current velocities, sharpnose shiner presence was negatively related to current velocity.
Figure 1 Fitted probabilities of logistic regression models for sharpnose shiner presence and frequency of depths for samples taken monthly from April 2003 through March 2004. Black dots represent the presence or absence of sharpnose shiner in each of the 50 seine hauls made during each month. Sharpnose shiner were absent in all samples during the months (August, December 2003 to February 2004) for which results are not presented
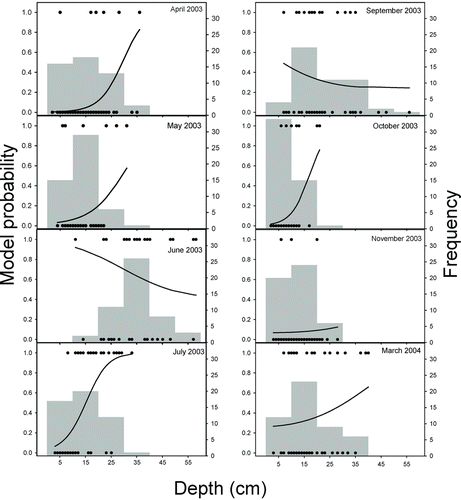
Figure 2 Fitted probabilities of logistic regression models for sharpnose shiner presence and frequency of current velocities for samples taken monthly from April 2004 through March 2005. Black dots represent the presence or absence of sharpnose shiner in each of the 50 seine hauls made during each month. Flood conditions prevented sampling during November 2004
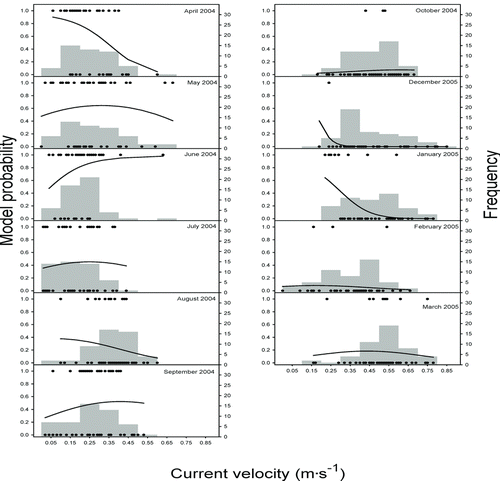
In general, sharpnose shiner was more likely to be absent from our collections during the months in which the greatest depths and current velocities were observed (Figures and ). The scarcity of fish in samples collected at these times is biologically relevant and presumably reflects the ability of sharpnose shiner to seek more suitable habitats outside the study area during periods of high flow.
Discussion
The presence of sharpnose shiner, at the microhabitat scale, was best predicted by depth or current velocity in different years of our study. The previous studies on habitat use by fish similarly have shown that depth and current velocity are the primary predictors of habitat use at the microhabitat scale. For example, Bain (Citation1995) studied over 100 different riverine fish species and found depth to be the single most influential variable defining habitat use. Current velocity, alone or in combination with depth, also has been found to be an important predictor of habitat use in a variety of stream fishes (Grossman and Freeman Citation1987; Bain et al. Citation1988; Harvey and Stewart Citation1991; Zweimüller Citation1995). The changes in the range of depths and current velocities observed through time in the upper Brazos River are primarily a reflection of seasonal and interannual patterns in regional precipitation, which varied substantially across the two years of our study. Precipitation was extremely low throughout the upper Brazos River region during 2003, whereas near record precipitation occurred during 2004. Consequently, we observed few extremes in current velocity during 2003, which may explain why depth, and not current velocity, was the only significant predictor of sharpnose shiner presence in that year. In contrast, current velocities were much greater on average during 2004; the year in which it was the only significant variable explaining sharpnose shiner presence.
Sharpnose shiner was absent from our collections during four months in 2003, even though the species was present at upstream and downstream sites during these months based on samples taken for other purposes. This exposes one limitation of our generalized linear models: they predict the presence of sharpnose shiner within one sampling reach, at the microhabitat scale, but do not necessarily allow predictions of sharpnose shiner presence among river reaches. The ability to make such predictions would be useful as fishes in the upper Brazos River, including sharpnose shiner, are known to move into and out of various reaches in response to changes in physical and chemical conditions (Ostrand and Wilde 2004) and because many Great Plains minnows, including the sharpnose shiner, are known to require extensive river reaches for population maintenance (Dieterman and Galat Citation2004; Perkin and Gido Citation2011; CitationWilde and Urbanczyk Forthcoming).
The changes in relative importance of depth and current velocity observed during the two years of our study demonstrate the difficulty associated with predicting abundance from specific values of individual variables. Nevertheless, predictions of population abundance are routinely made based on habitat suitability curves within the framework of instream flow models. Habitat suitability curves predict population size based on changes in habitat availability at different magnitudes of discharge (Orth and Maughan Citation1982; Rosenfeld Citation2003). The use of habitat suitability curves to predict abundance of fishes has been widely criticized because there is often little relationship between suitable habitat and fish abundance and these relationships have been rarely validated for individual species prior to their incorporation into instream flow models (Mathur et al. Citation1985; Scott and Shrivell Citation1987; Jowett Citation1992). Accurate predictions of abundance require mechanistic models based on factors directly related to population dynamics such as survival and fecundity (Wilde and Durham Citation2008; Durham and Wilde Citation2009), or models that incorporate comprehensive, species-specific information on both biotic and abiotic aspects of life history and ecology (Jowett Citation1992).
Fish that persist in extremely variable environments, such as occur in many Great Plains rivers, characteristically have broad tolerances to physical and chemical conditions (Matthews and Maness Citation1979; Matthews Citation1998; Ostrand and Wilde Citation2001; Higgins and Wilde Citation2005). However, the ability of fish to survive in these environments does not preclude them from actively selecting preferred or optimal habitats within the suite of habitats available at any given time. Thus, for sharpnose shiner, and other Great Plains fishes, it might be particularly important to distinguish between habitat requirements and habitat preferences (Rosenfeld Citation2003). As dissolved oxygen, salinity, and temperature during the two years of this study were within the range of tolerance for sharpnose shiner (Ostrand and Wilde Citation2001), the patterns of habitat use documented in this study likely reflect habitat preferences rather than specific habitat requirements related to physiological tolerances. Our results suggest that the relative importance of any one abiotic variable to sharpnose shiner habitat use at the microhabitat and local scales may increase or decrease as abiotic conditions vary through time.
Acknowledgements
We thank P. Bean, C. Chizinski, B. Durham, B. Gaines, C. Higgins, E. Holting, C. Huber, C. Hunt, R. Hunt, D. Knabe, M. Laman, C. Sanchez, J. Shuck, C. Stennett, and A. Urbanczyk for assistance in field collections. R. Patiño, K. Pope, L. Smith, and R. Strauss provided helpful comments on earlier drafts of this manuscript. Financial support for this study was provided by the Texas Parks & Wildlife Department and the United States Fish and Wildlife Service.
References
- Agnew , C and Anderson , E . 1992 . Water resources in the arid realm , London , , UK : Routledge Publishers .
- Akaike , H. 1973 . “ Information theory and an extension of the maximum likelihood principle ” . In Second international symposium on information theory , 268 – 281 . Budapest , , Hungary : Akademiai Kiado .
- Anderson , A A , Hubbs , C , Winemiller , K O and Edwards , R J . 1995 . Texas freshwater fish assemblages following three decades of environmental change . Southwestern Naturalist. , 40 : 314 – 321 .
- Anderson , K A , Beitinger , T L and Zimmerman , E G . 1983 . Forage fish assemblages in the Brazos River upstream and downstream from Possum Kingdom Reservoir, Texas . Journal of Freshwater Ecology , 2 : 81 – 88 . doi: 10.1080/02705060.1983.9664578
- Aparicio , E M , Vargas , J , Olmo , J M and de Sostoa , A . 2000 . Decline of native freshwater fishes in a Mediterranean watershed on the Iberian Peninsula: a quantitative assessment . Environmental Biology of Fishes , 59 : 11 – 19 . doi: 10.1023/A:1007618517557
- Bain , M B . 1995 . Habitat at the local scale: multivariate patterns for stream fishes . Bulletin Français de la Pêche et de la Pisciculture , 337–339 : 165 – 177 . doi: 10.1051/kmae:1995019
- Bain , M B , Finn , J T and Booke , H E . 1988 . Stream regulation and fish community structure . Ecology , 69 : 382 – 392 . doi: 10.2307/1940436
- Baker , J A and Ross , S T . 1981 . Spatial and temporal resource utilization by southeastern cyprinids . Copeia. , 1981 : 178 – 189 . doi: 10.2307/1444052
- Bonner , T H and Wilde , G R . 2000 . Changes in the fish assemblage of the Canadian River, Texas associated with reservoir construction . Journal of Freshwater Ecology , 15 : 189 – 198 . doi: 10.1080/02705060.2000.9663736
- Cross , F B and Moss , R E . 1987 . “ Historic changes in fish communities and aquatic habitats in plains streams of Kansas ” . In Community and evolutionary ecology of North American stream fishes , Edited by: Matthews , W J and Heins , D C . 155 – 165 . Norman , OK : University of Oklahoma Press .
- Dieterman , D J and Galat , D L . 2004 . Large-scale factors associated with sicklefin chub distribution in the Missouri and Lower Yellowstone rivers . Transactions of the American Fisheries Society , 133 : 577 – 587 . doi: 10.1577/T03-002.1
- Durham , B W and Wilde , G R . 2009 . Population dynamics of smalleye shiner an imperiled cyprinid fish endemic to the Brazos River, Texas . Transactions of the American Fisheries Society , 138 : 666 – 674 . doi: 10.1577/T07-234.1
- Dynesius , M and Nilsson , C . 1994 . Fragmentation and flow regulation of river systems in the northern third of the world . Science. , 266 : 753 – 762 . doi: 10.1126/science.266.5186.753
- Federal Register . 2002 . Endangered and threatened wildlife and plants; review of species that are candidates or proposed for listing as endangered or threatened; annual notice of findings on recycled petitions; annual description of progress on listing actions , Vol. 67 , 40657 – 40679 . Washington , DC : Office of the Federal Register, National Archives and Records Administration . No. 114
- Grossman , G D and Freeman , M C . 1987 . Microhabitat use in a stream fish assemblage . Journal of Zoology , 212 : 51 – 176 . doi: 10.1111/j.1469-7998.1987.tb05121.x
- Grossman , G D , Freeman , M C , Moyle , P B and Whitaker , J O Jr. 1985 . Stochasticity and assemblage organization in an Indiana stream fish assemblage . American Naturalist , 126 : 275 – 285 . doi: 10.1086/284415
- Grossman , G D , Moyle , P B and Whitaker , J O Jr. 1982 . Stochasticity in structural and functional characteristics of an Indiana stream fish assemblage: a test of community theory . American Naturalist. , 120 : 423 – 454 . doi: 10.1086/284004
- Harvey , B C and Stewart , A J . 1991 . Fish size and habitat depth relationships in headwater streams . Oecologia. , 87 : 336 – 342 . doi: 10.1007/BF00634588
- Higgins , C L and Wilde , G R . 2005 . The role of salinity in structuring fish assemblages in a prairie stream system . Hydrobiologia. , 549 : 197 – 203 . doi: 10.1007/s10750-005-0844-7
- Jelks , H L , Walsh , S J , Burkhead , N M , Conteras-Balderas , S , Diaz-Pardo , E , Hendrickson , D A , Lyons , J , Mandrak , N E , McCormick , F Nelson , J S . 2008 . Conservation status of imperiled North American freshwater and diadromous fishes . Fisheries. , 33 : 372 – 407 . doi: 10.1577/1548-8446-33.8.372
- Jowett , I G . 1992 . Models of the abundance of large brown trout in New Zealand rivers . North American Journal of Fisheries Management , 12 : 417 – 432 . doi: 10.1577/1548-8675(1992)012<0417:MOTAOL>2.3.CO;2
- Kehmeier , J W , Valdez , R A , Medley , C N and Myers , O B . 2007 . Relationship of fish mesohabitat to flow in a sand-bed southwestern river . North American Journal of Fisheries Management , 27 : 750 – 764 . doi: 10.1577/M06-016.1
- Kingsford , R T . 2000 . Protecting rivers in arid regions or pumping them dry? . Hydrobiologia. , 427 : 1 – 11 . doi: 10.1023/A:1004033915662
- Mathur , D , Bason , W H , Purdy , E J and Silver , C A . 1985 . A critique of the instream flow incremental methodology . Canadian Journal of Fisheries and Aquatic Sciences , 42 : 825 – 831 . doi: 10.1139/f85-105
- Matthews , W J . 1998 . Patterns in freshwater fish ecology , Norwell , MA : Kluwer Academic Publishers .
- Matthews , W J and Hill , L G . 1980 . Habitat partitioning in the fish community of a southwestern river . Southwestern Naturalist. , 25 : 51 – 60 . doi: 10.2307/3671211
- Matthews , W J and Maness , J . 1979 . Critical thermal maxima, oxygen tolerances, and population fluctuations in southwestern stream fishes . American Midland Naturalist , 102 : 374 – 377 . doi: 10.2307/2424665
- Moss , R E and Mayes , K B . 1993 . Current status of Notropis buccula and Notropis oxyrhynchus in Texas , Austin , TX) : Texas Parks and Wildlife Department . River Studies Report 8
- Nilsson , C , Reidy , C A , Dynesius , M and Revenga , C . 2005 . Fragmentation and flow regulation of the world's large river systems . Science. , 308 : 405 – 408 . doi: 10.1126/science.1107887
- Orth , D J and Maughan , O E . 1982 . Evaluation of the incremental methodology for recommending instream flows for fishes . Transactions of the American Fisheries Society , 111 : 413 – 445 . doi: 10.1577/1548-8659(1982)111<413:EOTIMF>2.0.CO;2
- Ostrand , K G and Wilde , G R . 2001 . Temperature, dissolved oxygen, and salinity tolerances of five prairie stream fishes and their role in explaining fish assemblages . Transactions of the American Fisheries Society , 130 : 742 – 749 . doi: 10.1577/1548-8659(2001)130<0742:TDOAST>2.0.CO;2
- Ostrand , K G and Wilde , G R . 2002 . Seasonal and spatial variation in a prairie stream-fish assemblage . Ecology of Freshwater Fish , 11 : 137 – 149 . doi: 10.1034/j.1600-0633.2002.00005.x
- Ostrand , K G and Wilde , G R . 2004 . Changes in prairie stream fish assemblages restricted to isolated streambed pools . Transactions of the American Fisheries Society , 133 : 1329 – 1338 . doi: 10.1577/T03-193.1
- Paine , M D , Dodson , J J and Power , G . 1982 . Habitat and food resource partitioning among four species of darters (Percidae: Etheostoma) in a southern Ontario stream . Canadian Journal of Zoology , 60 : 1635 – 1641 . doi: 10.1139/z82-214
- Perkin , J S and Gido , K B . 2011 . Stream fragmentation thresholds for a reproductive guild of Great Plains fishes . Fisheries , 36 : 371 – 383 . doi: 10.1080/03632415.2011.597666
- Pflieger , W L and Grace , T B . 1987 . “ Changes in the fish fauna of the lower Missouri River, 1940-1983 ” . In Community and evolutionary ecology of North American stream fishes , Edited by: Matthews , W J and Heins , D C . 166 – 177 . Norman , OK : University of Oklahoma Press .
- Polivka , K M . 1999 . The microhabitat distribution of the Arkansas River shiner, Notropis girardi: a habitat-mosaic approach . Environmental Biology of Fishes , 55 : 265 – 278 . doi: 10.1023/A:1007577411279
- Rosenfeld , J. 2003 . Assessing the habitat requirements of stream fishes: an overview and evaluation of different approaches . Transactions of the American Fisheries Society , 132 : 953 – 968 . doi: 10.1577/T01-126
- Scott , D and Shrivell , C S . 1987 . “ A critique of the instream flow incremental methodology with observations on flow determination from New Zealand ” . In Regulated streams: advances in ecology , Edited by: Kemper , B and Craig , J F . 27 – 43 . New York , NY : Plenum Press .
- Stanford , J A and Ward , J V . 1979 . The ecology of regulated streams , New York , NY : Plenum Press .
- Wilde , G R and Durham , B W . 2008 . A life-history model for peppered chub, a broadcast spawning cyprinid . Transactions of the American Fisheries Society , 137 : 1657 – 1666 . doi: 10.1577/T07-075.1
- Wilde , G R and Ostrand , K G . 1999 . Changes in the fish assemblage of an intermittent prairie stream upstream from a Texas impoundment . Texas Journal of Science , 51 : 203 – 210 .
- Wilde , G R and Urbanczyk , A C . 2013 . Relationship between river fragment length and persistence of two imperiled Great Plains cyprinids . Journal of Freshwater Ecology , doi: 10.1080/02705060.2013.785984
- Winston , M R , Taylor , C M and Pigg , J . 1991 . Upstream extirpation of four minnow species due to damming of a prairie stream . Transactions of the American Fisheries Society , 120 : 98 – 105 . doi: 10.1577/1548-8659(1991)120<0098:UEOFMS>2.3.CO;2
- Zweimüller , I. 1995 . Microhabitat use by two small benthic stream fish in a 2nd order stream . Hydrobiologia. , 303 : 125 – 137 . doi: 10.1007/BF00034050