
Abstract
Recreational fishing can alter the demography and life history characteristics of fish populations. Unexploited populations are useful references for fisheries managers to place into context the extent of changes in exploited populations and the outcomes of management actions. Our objective is to characterize the largemouth bass Micropterus salmoides populations in two unexploited reservoirs in Connecticut and compare them with an exploited reservoir. Unexploited Hemlock Reservoir was characterized by large proportions of large fish in good condition with relatively low population mortality rates. Unexploited Easton Reservoir's bass proportional size distributions were more similar to exploited Mansfield Hollow Reservoir bass population. However, both unexploited reservoirs had higher catch per effort of stock-size and greater fish than Mansfield Hollow Reservoir indicating greater overall density of large fish. The study of long-term unexploited populations may become more important in the future as managers consider the evolutionary consequences of recreational exploitation.
Introduction
Recreational fisheries can alter previously unexploited population characteristics quickly, including size and age structures, and mortality rates. For example, Goedde and Coble (Citation1981) documented rapid changes (within three years) in population characteristics of five recreationally targeted species in previously unexploited Wisconsin lakes after recreational angling began. Likewise, Jacobs and O’Donnell (Citation1996) reported declines of between 65% and 80% of electrofishing catch per unit effort of quality-sized (30 cm) largemouth bass Micropterus salmoides in three previously unexploited Connecticut drinking water reservoirs within a year of being of opening to angling. Management agencies employ numerous techniques and regulations, including seasonal closures, minimum, maximum and/or slot length limits, creel limits, and supplemental stocking to mitigate changes resulting from exploitation in an attempt to maintain recreational fisheries. The study of unexploited populations can provide managers with benchmarks to evaluate the status of fisheries and the effect of management regimes and actions (Hilborn & Walters Citation1992; Paukert & Willis Citation2001).
Largemouth bass are among the most popular freshwater sport fish in North America (USFWS 2011). They occur in 49 US states, in seven Canadian provinces, throughout Mexico (Noble Citation2002), and have been introduced worldwide. As a result of this popularity, the economic impact (USFWS Citation2011) and the ecological importance (e.g., Carpenter et al. Citation1987; Mittelbach et al. Citation1995) of largemouth bass, substantial management efforts have been expended to maintain or increase the abundance and improve the size structure of largemouth bass populations (Noble Citation2002). However, few unexploited populations exist and fewer yet have been studied (e.g., Clady Citation1975; Goedde & Coble Citation1981; Schindler et al. Citation1997; Mueller et al. Citation2005). Our objective was to define the population characteristics of interest to managers (e.g., size and age structure) of two unexploited largemouth bass populations in southern New England drinking water reservoirs and contrast them with an exploited population of largemouth bass.
Methods
Night boat electrofishing was used to sample three Connecticut largemouth bass populations. Hemlock Reservoir (176.8 ha; N 41° 13′ 2.27″, W 73° 17′ 19.78″) and Easton Reservoir (197.5 ha; N 41° 15′ 55.25″, W 73° 16′ 1.10″) are mesotrophic drinking water impoundments in southwest Connecticut, USA. Hemlock Reservoir and Easton Reservoir were impounded in 1914 and 1926, respectively. Both reservoirs have been closed to angling since their construction and while some illegal shoreline angling likely occurs, the lack of public boat access coupled with water company law enforcement patrols have left both lakes virtually unaffected by angling. Mansfield Hollow Reservoir (186.2 ha; N 41° 46′ 5.77″, W 72° 10′ 31.33″) is a mesotrophic flood control reservoir constructed in the early 1950s that supports a popular largemouth bass fishery (Edwards Jr. et al. Citation2004). Largemouth bass are not native to Connecticut; however, they were heavily stocked throughout the state in the 1800s. The bass populations in all three impoundments likely originated from the inundation of ponds containing these naturalized fish and perhaps from downstream transport from other ponds in the watershed.
Largemouth bass were sampled in October and November 2011 (Hemlock Reservoir), October and November 2012 (Mansfield Hollow Reservoir and Easton Reservoir). Each lake was sampled with one complete circuit of the shoreline. Individual fish were measured to the nearest centimeter, a subset of five fish per centimeter group (when possible) were weighed to the nearest gram and scales (five individuals per centimeter group when possible) were collected for age estimation. Additional samples collected during fall 2012 were used to supplement the number of age samples for Mansfield Hollow Reservoir. Scales were independently read by two of the authors (J. Hessenauer first read and E. O’Donnell or J. Davis second read), and discrepancies in age estimates were discussed among them and an agreed upon age estimate was assigned.
An age–length key was created for each lake using the proportional assignment method (Isermann & Knight Citation2005) based on 1-cm size bins and was used to assign ages to all individuals that had not been directly age-estimated. A log weight vs. log length regression was fit to the subset of individuals for which weight at capture was collected. Age distributions were compared using a chi-square test. Overall experiment-wise type-I error rates were maintained by applying a Bonferroni correction (data considered significant when p < 0.017). Given that all three populations are located in a relatively small geographic area and are genetically similar (Hessenauer, unpublished data), multiple Kolmogorov–Smirnov (KS) tests were used to compare length–frequency distributions among the three populations (corrected for multiple comparisons). Proportional size distributions were calculated (PSD; Guy et al. Citation2007), proportional size distribution of preferred sized fish (PSD-P) and proportional size distribution of memorable-sized fish (PSD-M), with 20 cm used as stock size, 30 cm used as quality size, 38 cm used as preferred size, and 51 cm used as memorable size (Anderson Citation1980). Catch per effort (CPE; individuals captured/hour) was estimated during electrofishing of stock, quality, preferred and memorable-sized fish in each population and compared those values using a chi-square test, with alpha corrected for multiple comparisons. Fish condition was estimated as relative weight (Wege & Anderson Citation1978) for fish over 15 cm using the following equation:
where Wr is the relative weight, W is the log weight of an individual fish and Ws is the length-specific standard weight of largemouth bass across its geographical range (from Blackwell et al. Citation2000). A von Bertalanffy (Citation1938) growth curve was fit to log length-at-age data for each population using the following equation:
where Lt is the length at age t, L∞ is the theoretical asymptotic length, K is the Brody growth coefficient, t is the age in years, and t0 is the theoretical age at which length was zero. An iterative approach was implemented, with start values determined by visually fitting a curve to the data. Standard errors for each parameter were estimated by bootstrapping 1000 iterations.
Total annual mortality rates were calculated using the Chapman and Robson (Citation1960) estimator and weighted regression on the descending limb of the catch curve. The ‘peak plus one’ approach advocated by Smith et al. (Citation2012) was implemented, such that analysis began one year after the ‘peak’ in the catch curve. Mortality rates were compared by testing for different slopes on the descending limb of the catch curve. Because data were only available from a single sampling year for each population, mortality rates might have been heavily influenced by a particularly strong year class (e.g., Miranda & Bettoli Citation2007). Therefore, for comparison, we also calculated total mortality using von Bertalanffy growth parameters, and the mean size of individuals vulnerable to the gear (assumed recruited to the gear at 20 cm) using the following equation:
where K is the Brody growth coefficient, L∞ is the asymptotic growth rate, Lmean is the mean size of individuals recruited to the gear, and Lx is the minimum size vulnerable to the gear (Miranda Citation2002). All statistical analyses were conducted using program R version 2.13.1 (R Development Core Team Citation2009, http://www.R-project.org).
Results
A total of 356, 836, and 418 individuals were sampled from Hemlock Reservoir, Easton Reservoir, and Mansfield Hollow Reservoir, respectively. Age estimates were made of 102, 160, and 221 individuals from Hemlock Reservoir, Easton Reservoir, and Mansfield Hollow, respectively.
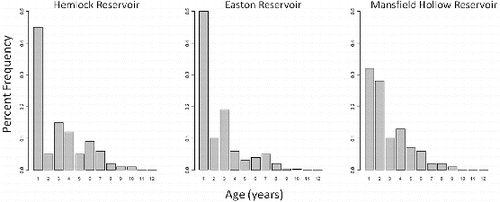
Unexploited Hemlock Reservoir had a similar age structure () as exploited Mansfield Hollow Reservoir (chi-square = 0.42, p = 0.51), but unexploited Easton Reservoir was significantly different from Hemlock Reservoir (chi-square = 10.95, p < 0.001) and Mansfield Hollow Reservoir (chi-square = 7.06, p = 0.007). Twenty-four percent (n = 74) of Hemlock individuals (excluding age-0) were estimated to have been age-5 and older. Fifteen percent (n = 61) of Easton individuals (excluding age-0) were estimated to be age-5 or older. Twenty-two percent (n = 72) of Mansfield Hollow individuals (excluding age-0) were estimated to have been age-5 and older.
Figure 1. Age frequency histograms (excluding age-0) of unexploited Hemlock Reservoir and Easton reservoir, and exploited Mansfield Hollow largemouth bass populations.
Hemlock Reservoir was characterized by greater PSD, PSD-P, and PSD-M than either Mansfield Hollow Reservoir or Easton Reservoir (). The mean length of sampled Hemlock Reservoir individuals was 26.7 ± 13.9 cm (±SD) and ranged from 5 to 52 cm. The mean length of sampled Easton Reservoir individuals was 19.0 ± 10.7 cm and ranged from 5 to 51 cm. The mean length of sampled Mansfield Hollow individuals was 17.1 ± 10.9 cm and ranged from 6 to 49 cm. Length–frequencies () were significantly different among all three populations (Hemlock and Mansfield: D = 0.41, p < 0.001; Hemlock and Easton: D = 0.34, p < 0.001; Mansfield and Easton: D = 0.24, p < 0.001).
Table 1. Population characteristics and 95% confidence intervals (where possible) for Hemlock Reservoir, Easton Reservoir, and Mansfield Hollow Reservoir in Connecticut, USA.
Figure 2. Length frequency histograms of unexploited Hemlock Reservoir and Easton Reservoir, and exploited Mansfield Hollow Reservoir largemouth bass populations.
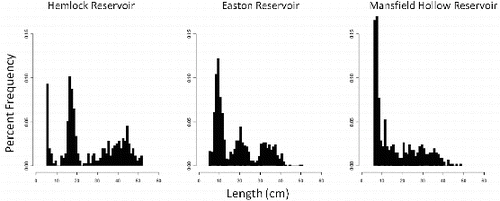
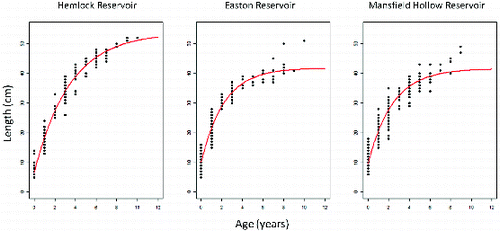
Figure 3. von Bertalanffy growth equation fit to length-at-age data of unexploited Hemlock Reservoir and Easton Reservoir, and exploited Mansfield Hollow largemouth bass populations.
While PSD values for Easton Reservoir were similar to Mansfield Hollow Reservoir, CPE data for Easton Reservoir more closely resembled Hemlock Reservoir, indicating higher densities of stock-sized and greater fish in the unexploited reservoirs (). CPE of stock, quality and memorable sized fish did not significantly differ between Hemlock Reservoir and Easton Reservoir (stock size: chi-square = 1.1, p = 0.29; quality size: chi-square: 0.57, p = 0.45); however, there was a significant difference between Hemlock Reservoir and Easton Reservoir for preferred size fish (chi-square = 12.3, p < 0.001). Mansfield Hollow had significantly lower CPE for stock and quality-sized fish when compared to Hemlock and Easton Reservoirs, respectively (stock size compared to Hemlock Reservoir, chi-square = 10.4, p < 0.001, compared to Easton Reservoir, chi-square = 18.4, p < 0.001; quality size: compared to Hemlock Reservoir, chi-square = 18.0, p < 0.001, compared to Easton Reservoir, chi-square = 12.6, p < 0.001). Mansfield Hollow Reservoir had a significantly lower CPE for preferred size when compared to Hemlock Reservoir (chi-square = 25.4, p < 0.001), but not Easton Reservoir (chi-square = 3.7, p = 0.05).
A subset of 91, 155, and 179 individuals from Hemlock Reservoir, Easton Reservoir, and Mansfield Hollow Reservoir, respectively were weighed. A significant (p < 0.05) log weight-log length relationship existed for all three populations (Hemlock: Weight = 3.21 × Length − 4.97, r2-adj = 0.99, F1,89 = 12,260, p < 0.001; Easton: Weight = 3.02 × Length − 4.34, r2-adj = 0.99, F1,153 = 18,750, p < 0.001; Mansfield: Weight = 3.20 × Length − 4.94, R2-adj = 0.99, F1,177 = 28,804, p < 0.001). All three populations were characterized by high relative weights: Hemlock Reservoir: 107.5 ± 3.4 (mean ± SD), Easton Reservoir: 107.3 ± 3.6, Mansfield Hollow Reservoir: 107.9 ± 3.5. The 95% confidence intervals overlapped () for all three estimates suggesting no difference among the three populations. A significant (p < 0.05) linear decline in relative weight-by-length existed for all three populations (Hemlock: Wr = −Length × 0.19 + 113.3, r2-adj = 0.47, F1,89 = 81.1, p << 0.001; Easton: Wr = −Length × 0.37 + 117.7, R2-adj = 0.69, F1,153 = 343, p < 0.001; Mansfield: Wr = −Length × 0.27 + 116.1, R2-adj = 0.55, F1,177 = 222.6, p < 0.001).
Individuals from Hemlock Reservoir reached larger asymptotic lengths than either Easton Reservoir or Mansfield Hollow Reservoir (). The von Bertalanffy growth equation for Hemlock Reservoir was: lt = 53.7[1 − e−0.29(t + 0.44)]. The growth equation for Easton Reservoir was lt = 41.7[1 − e−0.45(t + 0.60)]. For Mansfield Hollow Reservoir, the growth equation was lt = 41.5[1 − e−0.42(t + 0.61)].
Total annual mortality rates were similar (overlap of 95% confidence intervals) among the three populations. The descending limb of the age–frequency histograms () suggests full recruitment to the sampling gear at age-1 for all three populations, which corresponds to a mean size of 17.8 cm, 21.0 cm, and 22.0 cm, for Hemlock Reservoir, Easton Reservoir, and Mansfield Hollow Reservoir, respectively (). Therefore, as recommended by Smith et al. (Citation2012), we estimated mortality using the Chapman–Robson method for ages 2 to 10 for Hemlock Reservoir, Easton Reservoir, and Mansfield Hollow Reservoir, respectively. This yielded instantaneous total mortality (Z) estimates of 0.31 ± 0.035 (estimate ± SE) for Hemlock Reservoir, 0.41 ± 0.029 for Easton Reservoir, and 0.49 ± 0.045 for Mansfield Hollow Reservoir. No significant difference in slope (F2,20 = 1.73, p = 0.20) was detected for the descending limbs of the catch curve for each lake, suggesting that mortality rates were similar among the three lakes. Estimation of instantaneous total mortality (Z) based on von Bertalanffy growth parameters, recruitment to the gear at 20 cm and mean size of individuals recruited to the gear (37.9 cm, 29.9 cm, and 30.2 cm for Hemlock Reservoir, Easton Reservoir, and Mansfield Hollow Reservoir, respectively) yielded similar results (Hemlock: 0.25, Easton: 0.53, and Mansfield: 0.46) to those estimated from catch curve analysis. The similarity in results between these methods suggests that the catch curve analysis provided a reasonable approximation of total mortality in these populations.
Table 2. Observed mean length-at-age (cm) of largemouth bass in unexploited Hemlock Reservoir, unexploited Easton Reservoir, and exploited Mansfield Hollow Reservoir.
Discussion
Unexploited populations are typically characterized by high densities of large individuals (e.g., Clady et al. Citation1975; Goedde & Coble Citation1981; Reed & Rabeni Citation1989; Kocovsky & Carline Citation2001) relative to exploited populations. Unexploited Hemlock and Easton reservoirs had higher CPE of stock-sized and quality-sized individuals than exploited Mansfield Hollow Reservoir, consistent with these expectations. We also observed higher catch rates of preferred-sized individuals in Hemlock Reservoir relative to Easton and Mansfield Hollow reservoirs. We believe that this is related to the population of landlocked Alewives Alosa pseudoharengus, a prey source not overly common in impoundments in the Northeast (and not present in either Easton Reservoir or Mansfield Hollow Lake). The relationship between largemouth bass growth and clupeid prey is well documented, and seems particularly beneficial to largemouth bass once they reach a size capable of exploiting them (Michaletz Citation1997; Yako et al. Citation2000).
Despite the significant difference in CPE between Easton and Mansfield Hollow reservoirs, PSD were similar among the two populations. While heavily utilized by recreational anglers, creel surveys indicate harvest rates at Mansfield Hollow have been very low (∼1% or fewer of bass captured are harvested by anglers) during the past decade (Connecticut Department of Energy and Environmental Protection Inland Fisheries Division. unpublished data). The similarity in PSD and concurrent reduction in CPE, may reflect the fact that contemporary mortality associated with heavy recreational utilization is not overly size specific because few fish are harvested, rather fishing mortality is dominated by unintentional catch-and-release hooking mortality acting on all catchable size groups (likely stock-size and greater) equally. It is also possible that watershed characteristics and lake morphology may have affected CPE and basal differences in trophic state and transfer among the fish communities might be explanatory factors that we did not quantify.
The size structure () of Easton Reservoir more closely resembled Hemlock Reservoir (three clear modes representing ages-0, age-1, and age-2+) and both were markedly different from exploited Mansfield Hollow Reservoir. Easton Reservoir largemouth bass initially grew quickly, reaching 75% of L∞ by age 2, and ‘stockpiling’ at sizes just below L∞ (41.7 cm). Therefore, it is possible that in the absence of an abundant forage population (e.g., alewife), density-dependent processes shift the size structure of largemouth bass in Easton Reservoir to smaller sizes relative to Hemlock Reservoir. Paukert and Willis (Citation2004) documented similar density-dependent effects on largemouth bass size structure, but not condition, in shallow Nebraska lakes. Few memorable-sized individuals were located in either of the unexploited populations that we surveyed, suggesting that in the absence of angling and, in the case of Hemlock Reservoir, a source of preferred forage, few largemouth bass reach these sizes in the Northeast. Therefore, management efforts to increase the frequency of memorable-sized individuals in exploited populations in this region may be unlikely to elicit a measurable response.
Unexploited populations typically contain more relatively older individuals (e.g., Clady et al. Citation1975; Goedde & Coble Citation1981; Reed & Rabeni Citation1989; Kocovsky & Carline Citation2001) than exploited populations. This pattern was not observed in either Hemlock Reservoir or Easton Reservoir as only 24% and 15% of individuals we sampled were estimated over age 5 (not including sampled age-0 individuals), and no individuals older than age 10 were sampled in either population. Individuals of age 10 (and older) are not uncommon in the majority of regional exploited populations (Connecticut Department of Energy and Environmental Protection Inland Fisheries Division unpublished data). It is possible that the fast growth observed early in life reduces the average longevity of the population, as evidence among multiple taxa (including fish) suggests a tradeoff between the rate of growth and life span (Reznick et al. Citation2002; Metcalfe & Monaghan Citation2003). Alternatively, it is possible that ages estimated from scales inaccurately measured actual fish age, particularly of older individuals. Maceina and Sammons (Citation2006) reported that age estimates derived from largemouth bass scales from the Hudson River, NY, typically overestimated fish ages relative to otoliths for ages 1–9, and then underestimated ages relative to otoliths for age-10+ individuals. We observed relatively few individuals age-10 and older (3, 1, and 3 for Hemlock Reservoir, Easton Reservoir, and Mansfield Hollow Reservoir, respectively); however, the extent to which underestimation of fish ages may have affected our results is unknown. The sacrificing of large numbers of individuals (particularly large and presumably old individuals) from unexploited reservoirs and popular recreational fisheries to obtain otolith estimates was not possible. Despite the documented shortcomings of age estimation from scales, they remain the structure most commonly utilized by fisheries managers in the northeast for age estimation, facilitating comparison between the results we report here and data collected by fisheries managers.
We observed a significant decline in relative weight as total length increased for both unexploited populations, which is consistent with other unexploited fish populations (Reed & Rabeni Citation1989). Unexploited populations reportedly often contain old individuals in relatively poor body condition (Van Den Avyle & Hayward Citation1999). The suggested hypothesis is that a density-dependent process such as competition among the numerous large fish creates the decline in condition of these individuals (Reed & Rabeni Citation1989; Schindler et al. Citation1997). However, despite the overall negative trend, the average relative weight of individuals over 38 cm (preferred size) in Hemlock and Easton reservoirs still indicated good condition (Hemlock : 105.0 ± 1.9; Easton mean: 103.8 ± 1.9) and was close to each population's total mean, possibly indicating intraspecific competition was not strong enough to substantially affect condition.
The unexploited largemouth bass populations that we studied were markedly different than an unexploited largemouth bass population studied by Mueller et al. (Citation2005) in Washington, DC, USA. Unexploited Spencer Lake (28 ha) is located in the San Juan Archipelago, and contained a largemouth bass population which grew slowly (von Bertalanffy growth coefficient = 0.13 compared to 0.29 in Hemlock Reservoir and 0.45 in Easton Reservoir), reached slightly older ages (max observed age = 13 years), but remained relatively small, with no individuals sampled over 420 mm, and few over 300 mm. The authors suggested that this was a result of inadequate food supply (the lake contains only largemouth bass, smallmouth bass M. dolomieu, and Prickly Sculpin Cottus asper) and intense interspecific and intraspecific competition. The drastic contrast between the population studied by Mueller et al. (Citation2005) and the populations of Hemlock and Easton reservoirs suggests utility in locating and studying unexploited populations of popular sportfish in different water body types over broad geographic locations and biological conditions. Water supply reservoirs in Connecticut are perhaps unique since they provide household drinking water and pre-date modern water treatment procedures, so they have been protected and left largely undisturbed (and unfished) for approximately a century. The comparison value of unexploited populations increases when the unexploited system is of a similar size, species composition and climate as exploited populations of interest.
The utility of unexploited populations may expand as our understanding of the effects of recreational fishing on populations extends into the evolutionary arena. The potential for fisheries-induced evolution (Law Citation2007) was highlighted by research documenting that angling vulnerability in largemouth bass is heritable. Philipp et al. (Citation2009) developed two strains of largemouth bass with significantly different angling vulnerability by separating individuals caught frequently and infrequently by anglers into different ponds over multiple generations. Significant differences in metabolic rate and exercise recovery time were observed for the low and high vulnerability strains of largemouth bass (Cooke et al. Citation2007; Redpath et al. Citation2010). Likewise, high vulnerability individuals had greater reproductive fitness (Sutter et al. Citation2012). More diverse research techniques for characterizing unexploited populations, particularly locations with long-term closure, are needed to fully develop our understanding of the consequences of exploitation and the conservation potential of unexploited populations.
Acknowledgements
Electrofishing was conducted by DEEP Inland Fisheries Staff: P. Aarrestad, T. Barry, S. Gephard, N. Hagstrom, M. Humphrey, E. Machowski, C. McDowell and B. Orciari, and the following DEEP Seasonal Resource Assistants: K. Abbott, M. Cruz, C. Finch, H. Fitzpatrick, J. Geenty, M. Goklowski, M. Hecklinger, K. Job, K. Kleza, E. Lindquist, N. McMahon, J. Pioli, A. Rice, M. Smith and P. Vogt. We would also like to thank the Aquarion water company. Finally we would like to thank the anonymous reviewers whose comments improved earlier drafts of this manuscript.
Additional information
Funding
References
- Anderson RO. 1980. Proportional stock density (PSD) and relative weight (Wr): interpretive indices for fish populations and communities. In: Gloss S, Shupp B, editors. Practical fisheries management: more with less in the 1980's. Proceedings of the 1st Annual Workshop of the New York Chapter American Fisheries Society; p. 27–33. Ithaca (NY).
- Blackwell BG, Brown ML, Willis DW. 2000. Relative weight (Wr) status and current use in fisheries assessment and management. Rev Fish Sci. 8:1–44.
- Carpenter SR, Kitchell JF, Hodgson JR, Cochran PA, Elser JJ, Elser MM, Lodge DM, Kretchmer D, He X, von Ende CN. 1987. Regulation of lake primary productivity by food web structure. Ecology. 68:1863–1876.
- Chapman DG, Robson DS. 1960. The analysis of a catch curve. Biometrics. 16:354–368.
- Clady MD. 1975. The effects of a simulated angler harvest on biomass and production in lightly exploited populations of smallmouth and largemouth bass. Trans Am Fish Soc. 104:270–276.
- Clady MD, Campbell DE, Cooper GP. 1975. Effects of trophy angling on unexploited populations of smallmouth bass. In: Stoud R, Clepper H, editors. Washington (DC): Black Bass Biology and Management, Sport Fishing Institute; p. 425–429.
- Cooke SJ, Suski CD, Ostrand KG, Wahl DH, Philipp DP. 2007. Physiological and behavioral consequences of long-term artificial selection for vulnerability to recreational angling in a Teleost fish. Physiol Biochem Zool. 80:480–490.
- Edwards Jr. GP, Neumann RM, Jacobs RP, O’Donnell EB. 2004. Impacts of small club tournaments on black bass populations in Connecticut and the effects of regulation exemptions. North Am J Fish Manag. 24:811–821.
- Goedde LE, Coble DW. 1981. Effects of angling on a previously fished and unfished warmwater fish community in two Wisconsin lakes. Trans Am Fish Soc. 110:594–603.
- Guy CS, Neumann RM, Willis DW, Anderson RO. 2007. Proportional size distribution (PSD): a further refinement of population size structure index terminology. Fisheries. 32:348.
- Hilborn R, Walters CJ. 1992. Quantitative fisheries stock assessment: choice, dynamics, and uncertainty. New York (NY): Chapman and Hall.
- Isermann DA, Knight CT. 2005. A computer program for age-length keys incorporating age assignment to individual fish. North Am J Fish Manag. 25:1153–1160.
- Jacobs RP, O’Donnell EB. 1996. An electrofishing survey of selected Connecticut Largemouth Bass Lakes. Final Report 1996. Federal Aid to Sport Fish Restoration F57-R14. Hartford (CT): Connecticut Department of Environmental Protection.
- Kocovsky PM, Carline RF. 2001. Dynamics of the unexploited walleye population of Pymatuning Sanctuary, Pennsylvania, 1997–1998. North Am J Fish Manag. 211:178–187.
- Law R. 2007. Fisheries-induced evolution: present status and future directions. Mar Ecol Prog Ser. 355:271–277.
- Maceina MJ, Sammons RF. 2006. An evaluation of different structures to age freshwater fish from a northeastern US river. Fish Manag Ecol. 13:237–242.
- Metcalfe NB, Monaghan P. 2003. Growth versus lifespan: perspectives from evolutionary ecology. Exp Gerontol. 38:935–940.
- Michaletz PH. 1997. Influence of abundance and size of age-0 Gizzard Shad on predator diets, diet overlap, and growth. Trans Am Fish Soc. 126:101–111.
- Miranda, LE. 2002. Establishing sized-based mortality caps. North Am J Fish Manag. 22:433–440.
- Miranda, LE, Bettoli, PW. 2007. Mortality. In: Guy CS, Brown ML, editors. Analysis and interpretation of freshwater fisheries data. Bethesda (MD): American Fisheries Society; p. 229–277.
- Mittelbach GG, Turner AM, Hall DJ, Rettig JE, Osenberg CW. 1995. Perturbation and resilience: a long-term, whole-lake study of predator extinction and reintroduction. Ecology. 76:2347–2360.
- Mueller KW, Brouwer NL, Congdon BD. 2005. Characteristics of unexploited black bass populations from a remote lake in the San Juan Archipelago, Washington. Northwest Sci. 79:131–140.
- Noble RL. 2002. Reflections on 25 years of progress in black bass management. In: Philipp DP, Ridgway MS, editors. Black bass: ecology, conservation, and management. Bethesda (MD): American Fisheries Society, Symposium 31; p. 419–431.
- Paukert CP, Willis DW. 2001. Comparison of exploited and unexploited yellow perch Perca flavescens (Mitchill) populations in Nebraska Sandhill lakes. Fish Manag Ecol. 8:533–542.
- Paukert CP, Willis DW. 2004. Environmental influences on largemouth bass Micropterus salmoides populations in shallow Nebraska lakes. Fish Manag Ecol. 11:345–352.
- Philipp DP, Cooke SJ, Claussen JE, Koppelman JB, Suski CD, Burkett DP. 2009. Selection for vulnerability to angling in largemouth bass. Trans Am Fish Soc. 138:189–199.
- R Development Core Team. 2009. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available from: http://www.R-project.org.
- Redpath TD, Cooke SJ, Suski CD, Arlinghaus R, Couture P, Wahl DH, Philipp DP. 2010. The metabolic and biochemical basis of vulnerability to recreational angling after three generations of angling-induced selection in a Teleost fish. Can J Fish Aquat Sci. 67:1983–1992.
- Reed MS, Rabeni CF. 1989. Characteristics of an unexploited smallmouth bass population in a Missouri Ozark stream. North Am J Fish Manag. 9:420–426.
- Reznick D, Ghalambor C, Nunney L. 2002. The evolution of senescence in fish. Mech Ageing Dev. 123:773–789.
- Schindler DE, Hodgson JR, Kitchell JF. 1997. Density-dependent changes in individual foraging specialization of largemouth bass. Oecologia. 110:592–600.
- Smith MW, Then AY, Wor C, Ralph G, Pollock KH, Hoenig JM. 2012. Recommendations for catch-curve analysis. North Am J Fish Manag. 32:956–967.
- Sutter DAH, Suski CD, Philipp DP, Klefoth T, Wahl DH, Kersten P, Cooke SJ, Arlinghaus R. 2012. Recreational fishing selectively captures individuals with the highest reproductive fitness potential. Proc Natl Acad Sci USA. 109:20960–20965.
- [USFWS] U.S. Department of the Interior, Fish and Wildlife Service, and U.S. Department of Commerce, U.S. Census Bureau. 2011. Washington (DC): National Survey of Fishing, Hunting and Wildlife-Associated Recreation.
- Van Den Avyle MJ, Hayward RS. 1999. Dynamics of exploited fish populations. In: Kohler CC, Hubert WA, editors. Inland fisheries management in North America, 2nd ed. Bethesda (MD): American Fisheries Society; p. 127–166.
- von Bertalanffy L. 1938. A quantitative theory of organic growth. Human Biology. 10:181–213.
- Wege GW, Anderson RO. 1978. Relative weight (Wr): a new index of condition for largemouth bass. In: Novinger GD, Dillard JG, editors. New approaches to the management of small impoundments. Bethesda (MD): North Central Division, American Fisheries Society, Special Publication No. 5; p. 79–71.
- Yako LA, Mather ME, Juanes F. 2000. Assessing the contribution of anadromous herring to largemouth bass growth. Trans Am Fish Soc. 129:77–88.