
Abstract
To investigate the effect of summer monsoon-derived inputs of particulate organic matter (POM) on patterns of stable isotopes in the food web in a reservoir, we traced dietary sources of carbon and nitrogen in pelagic zooplankton and herbivorous littoral zoobenthos following a monsoonal disturbance in Lake Paldang, a large regulated reservoir in Korea. Carbon and nitrogen stable isotopes were analyzed in POM, zooplankton, sedimentary organic matter, zoobenthos, and piscivorous fishes in pre- and post-summer monsoon periods. Sharp depletion of 13C and enrichment of 15N of POM along with elevated POC/Chl a ratios occurred following the summer monsoon period, reflecting the rapid reduction of phytoplankton biomass and a pulsed supply of terrestrial organic detritus. Herbivorous zooplankton δ13C values, which were initially similar to POM δ13C values in the pre-summer monsoon period, decreased to nearly that of terrestrial organic matter, providing isotopic evidence of allochthonous contribution to zooplankton in the post-summer monsoon period. In contrast, herbivorous zoobenthos in the littoral area and higher fishes appeared to have a similar stable isotope composition between the pre- and post-summer monsoon period. Our findings indicate that although monsoon storms dramatically decrease phytoplankton biomass, the augmented allochthonous material after the summer monsoon offsets this effect temporarily by subsidizing pelagic zooplankton production. Allochthonous subsidies to herbivorous littoral zoobenthos and piscivorous fishes are relatively limited.
Introduction
As an undefined mixture of allochthonous and autochthonous sources (Hedges et al. Citation1986; Ock & Takemon Citation2014), the quantity and composition of particulate organic matter (POM) in lakes can provide an integrated record of natural and anthropogenic activity within a basin (Kendall et al. Citation2001) and play an important role in influencing secondary production in freshwater ecosystems (Thorp & Delong Citation1994; Rosi-Marshall & Wallace Citation2002; Bunn et al. Citation2003; Lau et al. Citation2014). The composition of lentic POM (i.e., the proportions of autochthonous and allochthonous organic matter) is known to change seasonally along gradients of temperature, light, and precipitation, including storm events, throughout a year. Typically, phytoplankton in temperate lakes are more productive in summer, resulting in increasing autochthony of POM (Gaedke Citation1992; Grey et al. Citation2001). Meanwhile, a large fraction of allochthonous carbon is usually added to lakes through increased runoff waters during rainy season, which is winter in Mediterranean regions (Gu et al. Citation2006) and summer in monsoon regions (Kim et al. Citation2000). In addition to temporal variation, sources of carbon fueling lake food webs can be spatially heterogeneous within a reservoir system along environmental and trophic gradients (Doi et al. Citation2013).
Recent studies have argued that increased allochthony can subsidize zooplankton production in pelagic food webs. Grey et al. (Citation2001) found that herbivorous zooplankton seasonally shift their diet sources between autochthonous and allochthonous matter. Rautio et al. (Citation2011) demonstrated that phytoplankton was not the sole production source for zooplankton in lakes particularly where primary production was restricted. Carpenter et al. (Citation2005) emphasized the role of allochthony for zooplankton production in unproductive lakes by comparing it to eutrophic lakes. Conversely, Brett et al. (Citation2012) suggested that lake herbivore consumption is supported largely by autochthonous production in a lake where hydraulic advection is strong, despite a high flux of allochthonous resources.
In the monsoon climate regions of East Asian countries, including South Korea, more than 70% of annual rainfall occurs intensively in summer and a long dry period follows in winter, both of which cause great fluctuations in storage and discharge of reservoirs. Previous studies conducted in monsoon reservoirs observed that the balance of allochthonous and autochthonous resources in reservoirs is subjected to temporal disturbance by summer monsoon storms; during summer, phytoplankton production drops drastically whereas large amounts of terrestrial materials are temporarily imported to reservoirs from upstream rivers and tributaries (Kim et al. Citation2000; Park et al. Citation2009). However, there have been few studies to assess the effect of summer monsoon-derived inputs on higher consumers in a reservoir food web (i.e., zooplankton in a pelagic area, zoobenthos in a littoral area, and fishes) because of the complexity of effective tracers linking the ecological measurements with biochemical analysis, as well as multiple environmental factors which significantly obscure the effect of summer monsoon.
Natural stable isotope techniques can be used to overcome the limitation by understanding the dynamics of food webs and detecting responses to environmental change. In particular, carbon isotopes are effective tracers for identifying POM sources and tracing food sources of consumers in aquatic ecosystems (Finlay Citation2001; Post Citation2002). Nitrogen isotopes are useful for estimating trophic position in food webs (Vander Zanden et al. Citation1997; Post Citation2002; Boecklen et al. Citation2011; Perkins et al. Citation2014). Therefore, increasing isotopic evidence associated with allochthonous material subsidies in lake ecosystems can be applicable to monsoon reservoirs where trophic status changes considerably following summer rainfalls. In this study, we attempt to clarify if augmented allochthony could subsidize secondary production in a reservoir food web (i.e., pelagic zooplankton and littoral zoobenthos) and if the effect would be reflected in the upper components of the food web by assessing the responses of reservoir pelagic and littoral food webs to summer monsoon-derived inputs of POM in a monsoon reservoir system. We measured carbon and nitrogen stable isotope composition of POM, pelagic zooplankton, littoral sedimentary organic matter (SOM), zoobenthos, and piscivorous fishes in Lake Paldang on the Han River system, South Korea.
Methods
Study area
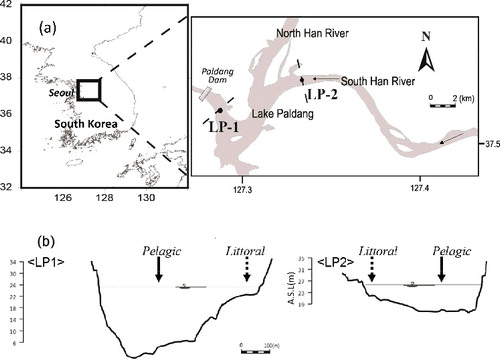
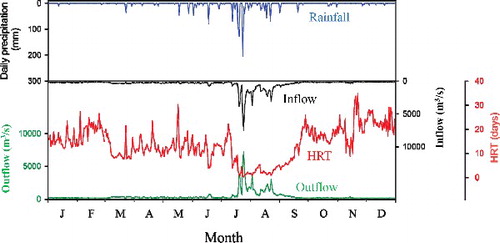
Lake Paldang (37°31'33" N, 127°16'44" E) is a large reservoir (250 million m3 of storage capacity) covering 23,800 km2 of basin area that was built in 1973 for water supply and hydropower generation in the downstream area of the Han River system. This lake is the largest drinking water source for the downstream Seoul metropolitan area with approximately 24 million people. This shallow reservoir (6.5 m average depth) has a fast turnover rate (5 d average hydraulic retention time [HRT]; Park et al. Citation2013) due to flow regulation for multi-purpose dam operation, and thus reservoir water is mixed vertically and rarely stratified in the summer. The catchment basin exhibits a typical Asian monsoon climate characterized by a wet summer and dry winter. The basin area is surrounded mostly by hardwood deciduous forest, followed by agricultural fields (20%) and urban areas (6%) which cause large anthropogenic loads of nutrients from upstream rivers.
Field sampling and laboratory analyses
We established two study sites along the lake, one near the dam (LP1) with deep and wide waters and the other near an inflowing river (LP2) with relatively shallow and narrow waters (). At each site cross-section, we spatially separated a pelagic area located at the deepest open zone from a littoral area close to the lake shore (<1.0 m depth). We collected POM samples at the surface layer (0.1 m depth) by boat near the center of each site using a Van Dorn sampler monthly, from March to October 2008 (winter period was excluded due to lake freezing) at the LP1 site. Sampling at the LP2 site was done before and after the monsoon rainfall (May and September, respectively). The samples were filtered using a 100 µm mesh to remove zooplankton, and were filtered again using pre-combusted GF/F filters. We also separately collected zooplankton, SOM, zoobenthos, and fish on the two occasions: zooplankton were collected using a vertically towed plankton sampler (23 cm mouth diameter, 50 cm length, and 60 µm mesh) from surface water in the pelagic area. Zoobenthos and SOM were sampled using a grab sampler at 0.5–1.0 m depth in the littoral area, where a few taxa of macroinvertebrate species were collected, such as non-biting midges (Chironomidae sp.), two freshwater snails (Semisulcospira gottschei and Physa acuta), an oriental river prawn (Macrobrachium nipponense), and a common burrowing mayfly (Ephemera orientalis). For piscivorous fish samples, skygager (Erythroculter erythropterus), steed barbel (Hemibarbus labeo), largemouth bass (Micropterus salmoides), and bluegill (Lepomis macrochirus) were captured using a triangular net (5 mm mesh) in the pelagic area. Only adult samples more than 20 cm in length (except bluegill which was more than 15 cm) were selectively taken, and transported on ice to the laboratory and stored frozen until analysis. For analysis of stable carbon and nitrogen isotope ratios, most samples were ground into a powder, but small ones (less than 1 mg in dry mass) were analyzed in bulk. For the carbon stable isotope analysis, inorganic carbon was removed with 1N HCl treatment, whereas untreated samples were used for the nitrogen analysis (Carabel et al. Citation2006). For dominant zooplankton, more than 50 individuals of cladoceran Daphnia and copepod Cyclops were manually extracted for measurement. For fish samples, white muscle tissue anterior to the dorsal fin was prepared and dichloromethane-methanol treatment was added for lipid extraction. Stable carbon and nitrogen isotope ratios of the samples were measured separately using a mass spectrometer (EA-IRMS: EuroEA-Isoprime IRMS, GV Instruments, UK) at Hanyang University, South Korea. Carbon and nitrogen isotopic compositions are reported in the standard delta (δ) notation relative to the Vienna Peedee Belemnite and to atmospheric nitrogen, respectively.
Figure 1. Study site map (a) and cross sections (b), LP1 near dam is deep and wide, whereas LP2 near an inflowing river is relatively shallow and narrow (b).
For samples used to measure phytoplankton and zooplankton composition and density, the same methods of collecting POM and zooplankton described above were used. Then plankton particles were sorted and counted using an optical microscope with 100–400× magnification (National Institute of Environmental Research [NIER] Citation2009). At each site, water samples for biochemical analyses were separately collected at the pelagic area and filtered by Whatman GF/F filter to separate POM from dissolved. Chlorophyll a concentration (Chl a) was measured spectrophotometrically and particulate organic carbon (POC) concentration was determined using an elemental analyzer (Euro EA3028, http://www.eurovector.it). Elemental concentration analysis of POM samples was not conducted. Regression analysis was used to evaluate the relationship among hydrological parameters, and between POM isotopic and environmental parameters. Student's t-test was used to compare temporal and spatial isotopic differences of basal food sources and consumers. All statistical analyses were performed with SPSS software (ver. 21. IBM Inc., http://www.ibm.com).
Results
Reservoir hydrology
Typical of meteorological characteristics in monsoon regions, more than 70% of annual precipitation in 2008 occurred between July and September followed by a dry period (). The most intensive rainfalls occurred during typhoons: in July, 610 mm of precipitation was recorded and there were occasional heavy rains exceeding 50 mm/d until August. A significant positive linear relationship was observed between precipitation and inflow (r > 0.9, p < 0.001), and between inflow and outflow (r > 0.9, p < 0.001) with distinct seasonal variations, where all hydrological parameters were higher in summer than in autumn or spring seasons. Thus, monthly inflow to the lake depended strongly on precipitation, and the volume of downstream release from the dam was almost equally as large as the inflow throughout the year. As a result, precipitation in the reservoir tends to determine HRT of the reservoir (r = 0.8, p < 0.01; ). The short HRT (<5 d) were observed in July when heavy rainfalls occurred. Annual mean HRT in the lake was 14 d in 2008, indicating a rapid hydrological turnover occurring 26 times a year.
Figure 2. Annual distribution of rainfall, reservoir hydrology, and HRT in 2008.
Composition and biomass changes in phytoplankton and zooplankton
Phytoplankton composition in Lake Paldang varied seasonally and spatially. Diatoms Stephanodiscus hantzschii, Asterionella forma, and Aulacoseria granulate dominated in the reservoir throughout the year, although the proportion of diatoms declined gradually during the period from spring to autumn as the fractions of green algae (e.g., Hormidium spp.) and cyanobacteria (e.g., Anabaena spp. and Microcystis spp.) increased (Figure 3(a) and 3(b)). Chl a, an indicator for phytoplankton biomass (Flemer Citation1969), did not follow the typical seasonal trend of temperate lakes. At the LP1 site, high Chl a values in the spring (18–26 µg C/L) reached a peak of 30 µg C/L in July before the rainy season started. However, Chl a dropped abruptly to 11 µg C/L (a 63% reduction) in response to monsoon rains in August. It then increased gradually to 25 µg C/L for autumn ((a)). At the LP2 site, overall high Chl a (ranging between 17–58 µg C/L) was observed before summer monsoon period, but showed the much sharper decline in August from 27 to 5 µg C/L (an 81% reduction, (b)). POC concentration was temporally stable in the LP1 site (1.2 ± 0.2 mg C/L) relative to that in the LP2 site (1.1 ± 0.43 mg C/L), where the highest value (1.7 mg C/L) was observed in August ().
Figure 3. Monthly variation in phytoplankton composition and chlorophyll a concentration (a and b), and zooplankton composition and biomass (c and d).
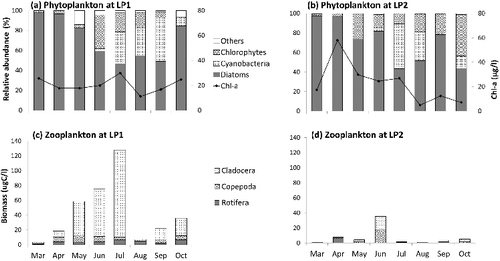
The composition of zooplankton community varied seasonally in response to summer rainfall, similarly to phytoplankton changes in the same year. Over the course of the year, the dominant zooplankton at LP1 were cladoceran species, including Daphnia galeata, Bosmina longirostris, and Diaphnanosoma brachyurum (33.6 ± 37.6 µg C/L), followed by copepods Cyclops vicinus, Eodiptomus sp., and copepodids (4.7 ± 3.8 µg C/L), and rotifers Polyarthra vulgaris and Keltella cochlearis (3.3 ± 2.3 µg C/L). The rotifers became dominant in August as the biomasses of both cladocerans and copepods rapidly dropped during and immediately after the summer rainfall periods. Total biomass of zooplankton grew rapidly in the spring, reaching its peak in July (128 µg C/L) but dropped sharply by 95% in August (7 µg C/L) and then increased steadily until late autumn ((c)). Zooplankton composition of the LP2 site appeared to have a similar seasonal trend as the LP1 site but the overall total biomass was 73% lower (7.6 ± 11.6 µg C/L). The peak of zooplankton biomass at the LP2 site reached 35.7 µg C/L in June and then declined to 1 µg C/L in July ((d)).
Stable isotope compositions of POM and zooplankton
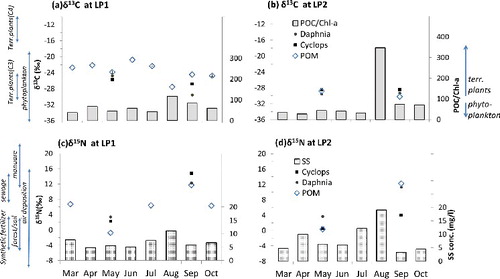
At the LP1 site, the POM δ13C values varied widely over time from −27.6 to −20.8‰ (with a mean of −23.8 ± 2.0‰), showing a rapid drop to the lowest values in August during monsoon rainfalls ((a)). The POM δ13C values taken from March to July were significantly higher (−22.4 ± 1.1‰) than those collected after the summer rainfalls in August to October (−25.6 ± 1.4‰; t = 3.27, df = 6, p < 0.01), indicating possible temporal change in the POM composition following the summer monsoon period. In May, the δ13C values of zooplankton at the LP1 were very close to that of POM (0.8–1.8‰ difference), while in September, because of more 13C depleted zooplankton, the difference in δ13C values between zooplankton and POM became more than two times greater (as much as 2.3–5.1‰; (a)). The temporal variation means that a heavy trophic reliance of zooplankton on the bulk POM composition in the dry season became weaker following the rainy season. On the other hand, POM δ15N in the LP1 site showed a wide range of variation (6.4 ± 4.5‰, n = 5), recording the lowest value (−0.7‰) in May and the highest (11.8‰) in September. The δ15N of Daphnia and Cyclops in September became enriched compared with those collected in May ((b)). At site LP2, POM δ13C values were more 13C-depleted in both May and September, as compared to the values taken at LP1. A similar pattern of 13C-depletion and 15N-enrichment in POM samples was observed after the rainy season, despite a limited data-set (Figure 4(c) and 4(d), and ).
Figure 4. Temporal variations in carbon and nitrogen stable isotope compositions of POM and zooplankton, POC/Chl a values (a and b), and SS concentration (c and d). Typical ranges for potential sources are added in the vertical axis. The data are modified from Meybeck et al. (Citation1988); Riemann et al. (Citation1989); Kendall et al. (Citation2001); Dawson et al. (Citation2002); Finlay & Kendall (Citation2007).
Stable isotope compositions of SOM, zoobenthos and fish
In contrast to POM and zooplankton's isotopic variations, there were few differences between May and September for the stable carbon isotope compositions of SOM, littoral zoobenthos, and fish in the LP1 site (p > 0.05 for each) (). Littoral zoobenthos and SOM δ13C values were almost within the range of typical trophic fractionation values in May, and a similar pattern was observed in the stable nitrogen isotopes, showing a reliance on SOM for energy resources in pre-summer monsoon period. However, the differences increased slightly in September. Piscivorous fishes H. labeo, L. macrochirus, and M. salmoides showed the highest signatures of δ15N (14.1 ± 0.64‰) among all animal samples collected in the dry and wet season, implying that they are top level predators in the reservoir food webs. On the other hand, for stable isotope ratios observed at LP2, overall carbon isotope ratios of SOM, zoobenthos, and fish samples were significantly 13C-depleted on both occasions when compared with those at the LP1 site (by 1.5‰, t = 7.64, df = 2, p < 0.001 for SOM, and by 3.6‰, t = 2.26, df = 4, p < 0.05 for zoobenthos, and by 2.1‰ t = 2.50, df = 3, p < 0.05 for fish), implying spatial heterogeneity between LP1 and LP2 sites.
Discussion
The effect of summer monsoon-derived inputs of POM on reservoir POM
Despite a rapid decrease in Chl a concentration in August, POC concentration was observed to be high in both study sites ( and ). It can be related to the temporal changes in balance of autochthonous and allochthonous materials of reservoir POM. Isotopic evidence demonstrates that allochthonous materials were introduced more rapidly to the lake during the rainy period while phytoplankton biomass was reduced in the water column immediately after monsoonal rainfall. POC/Chl a ratios can trace the temporal variation in POM source composition, because POC in lake waters includes terrestrial components whereas Chl a is specific to autotrophs (Marra Citation2002). Based on the POC/Chl a values, Meybeck et al. (Citation1988) and Riemann et al. (Citation1989) distinguished terrestrial detritus (POC/Chl a greater than 100 (mg/L)/ (mg/L)) from phytoplankton (POC/Chl a between 30 and 100). In our study, in the pre-summer monsoon period, the POC/Chl a values were relatively constant from spring to early summer ranging between 39 and 68 at LP1 and between 28 and 44 at LP2. However, the values increased abruptly in August to 116 at LP1 and 347 at LP2 (Figure 4(a) and 4(b)). Thus, there was a notable change in POM composition from phytoplankton-derived organic matter in the pre-summer monsoon period to a relatively large amount of terrestrial organic matter during the summer monsoon period. Previous studies reported these drastic monsoonal changes between autochthonous and allochthonous organic materials. Park et al. (Citation2009) found a similar drop in autochthonous contribution when a wet season began with the lowest value in the rainy period reaching nearly 0%, meaning virtually all allochthonous organic matter when higher monsoon rainfall occurred in the reservoir. Kim et al. (Citation2000) estimated the lowest phytoplankton fraction during the summer rainy season. They suggested that a rapid turnover rate (HRT < 5 d) for the rainy season along with a much higher turbidity suppressed primary production and more phytoplankton exported through dam outflow, resulting in a reduction in total phytoplankton biomass. Also, such large amounts of terrestrial material increase the importance of allochthonous contributions to lentic ecosystems (Kim et al. Citation2000; Wang et al. Citation2011; Park et al. Citation2013).
Table 1. Monthly measurements of POC, chlorophyll a concentrations, and their ratio in the study sites in 2008.
Table 2. Carbon and nitrogen stable isotope composition (average ± standard deviation) of POM, SOM, zooplankton, zoobenthos, and fish during pre- and post-monsoon periods.
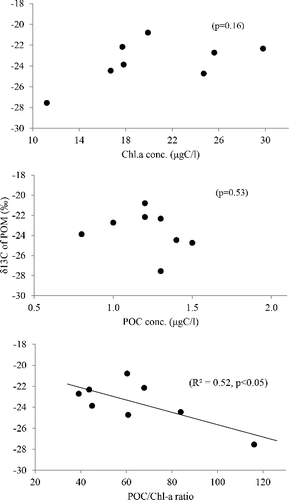
Because of a frequent overlap in carbon stable isotope value between freshwater phytoplankton (with a wide range variation from −42 to −24‰ depending on the dissolved inorganic carbon (DIC) sources and taxonomical groups; Vuorio et al. Citation2006) and the C3 vascular tree litter of temperate forests (which are relatively constant around −29 to −24‰ due to well-mixed atmosphere carbon pool; Balesdent et al. Citation1993; Dawson et al. Citation2002), there has been a limitation to the use of a single carbon isotope to distinguish the two major sources of POM. The POC/Chl a value in combination with carbon isotope signatures can be a useful tracer for identifying the summer monsoon effect on POM source composition in the reservoir. The POM 13C values were significantly depleted in the post-summer monsoon (−25.6 ± 1.7‰) compared with those in the pre-summer monsoon (−22.4 ± 1.1‰) at the LP1site (t = 3.83, df = 7, p < 0.01). In particular, there was a significant negative correlation of δ13C POM with POC/Chl a ratio (r = −0.73, F1,6 = 6.56, p < 0.05) but with neither Chl a nor POC (). The results indicate that the terrestrial-derived POM following the monsoon rainfall has a relatively lighter signature in 13C value (which corresponds to that generally found in temperate forests; Balesdent et al. Citation1993) than phytoplankton-dominated POM (which ranged between −24 and −20‰) in the pre-summer monsoon. This is isotopic evidence reflecting the monsoonal change in POM source composition shifting from autochthonous to allochthonous organic materials in the reservoir trophic system.
Figure 5. Relationship between chlorophyll a concentration and POM δ15C values.
On the other hand, the highest range of δ15N (around 12‰) in all POM samples throughout the year was found after monsoon rainfall period (September). The enriched 15N values in the post-summer monsoon are distinctly out of range of typical temperate forests (−3 to 5‰; Kendall et al. Citation2001; Finlay & Kendall Citation2007) but within the ranges of agricultural manure soil (7 to 20‰; Cravotta Citation1997; ) and is likely derived from allochthonous inputs from upstream farm land drainage (Lake et al. Citation2001; Kohzu et al. Citation2009; Nguyen & Hur Citation2011). The highest total nitrogen concentration was observed in August at the LP1 site when suspended solids concentration was at the highest level, suggesting an allochthonous nitrogen input to the lake by the rainfall runoff. However, the carbon isotope differences between LP1 and LP2 exhibited spatial heterogeneity of allochthony within the lake. The relatively lower δ13C values observed at LP2 in September indicate that a shallow water site near an inflowing river can be more sensitive than a deep water site near a dam in its response to summer monsoon influence on POM composition. With flow reduced in a lacustrine zone (typically deep and still waters), the relative importance of allochthony can decrease due to the sedimentation process of allochthonous particles crossing through a relatively shallow and fast-flowing transitional zone (Fisher & Likens Citation1972; Cushing et al. Citation1993; Webster & Meyer Citation1997; Doi Citation2009).
The effect of monsoon on primary consumers and top predators
The zooplankton 13C values were significantly depleted in September following monsoon storms (−28.6 ± 2.5‰) compared with the dry period of May (−25.1 ± 0.6‰, t = 3.52, df = 3, p < 0.05), suggesting the zooplankton dietary source likely shifted from phytoplankton to terrestrial-derived organic matter (). In addition, it was found that the narrow range of δ13C values between zooplankton and POM in May (<2‰ in difference) broadened in September (2.4–5.1‰ in difference), especially in Daphnia relative to Cyclops. A broader gap which is more than the trophic fractionation ratios (Vander Zanden & Rasmussen Citation2001) indicates a weaker reliance on bulk POM for dietary availability after the monsoon rainfall. Grey et al. (Citation2000) found that zooplankton δ13C values are closer to those of bulk POM in eutrophic lakes where phytoplankton is available for diets than in oligotrophic lakes where phytoplankton biomass is limited. According to France & Peters (Citation1997), zooplankton rely on allochthonous material under oligotrophic conditions but the extent of reliance varies depending on specific feeding types of zooplankton species. Our results reveal that in the pre-monsoon season, when phytoplankton-dominated POM in the reservoir, both Daphnia and Cyclops fed mainly on phytoplankton, whereas they changed their diet sources when the lake POM composition became temporarily terrestrial dominated following summer monsoon rainfall. Cladocerans which filter whole suspended materials appeared more 13C-depleted than the large Cyclops which can selectively take chlorophytes and rotifers (Zohary et al. Citation1994; Pulido-Villena et al. Citation2005). Thus, our study also shows that the filter-feeding Daphnia assimilated more allochthonous sources following the summer monsoon period than the large copepods did. Pace et al. (Citation2004) supports our finding showing a significant subsidy of terrestrial-derived carbon to Daphnia accounting for 20%–50% contribution. Although Daphnia can shift their basal resource depending on POM composition condition in the post-summer monsoon period by allochthonous subsidy, total biomass of zooplankton declined to the lowest level at this time (Figure 3(c) and 3(d)). Its effect on the higher trophic level consumers feeding on zooplankton could be limited because of deficits in diet resources.
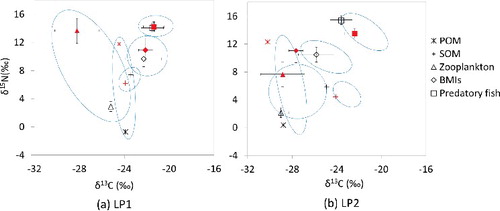
When compared to a large range of temporal changes in zooplankton stable isotope ratios (a decrease in δ13C by 3.0‰ and an increase in δ15N by 10.8‰ at LP1 site), only a minor variation was found in zoobenthos (almost no change in δ13C and an increase in δ15N of 1.2‰, (a)), implying that in spite of monsoonal POM disturbance diet sources, littoral zoobenthos were stable relative to pelagic zooplankton. Similarly, the fish δ13C and δ15N values were stable following the monsoonal disturbance in basal resources. Considering trophic isotopic enrichment of nitrogen between prey and predator, the elevated δ15N of zooplankton (near 13.7‰) was too high to be assimilated by the top predatory fish (14.2‰ on average), whereas it implied a closer reliance on zoobenthos ((a)). It indicates that the allochthonous subsidies to zooplankton following the summer monsoon storms have little influence on top predator fishes. It is likely that a slow biological turnover in fishes as long as 116–173 d (Trudel et al. Citation2011) prevents seeing differences, buffering the effect of allochthonous subsidizes to zooplankton, but also a littoral area can provide an important feeding habitat for the fishes during post-summer monsoon. Fish are known to have an important trophic position in lake food webs, integrating the pelagic and littoral food webs by their flexible feeding habits (Post et al. Citation2000; Vander Zanden & Vadeboncoeur Citation2002; Doi Citation2009). However, the isotopic differences of consumers between LP1 and LP2 (Figure 6(a) and 6(b)) show that the food web responses to the augmented allochthony in basal resources are spatially heterogeneous within a lake system. The zoobenthos and fishes near the inflowing river are found to be relatively sensitive to the POM composition changes.
Figure 6. Comparisons of carbon and nitrogen stable isotope composition for POM, zooplankton, SOM, herbivorous zoobenthos, and top predatory fish between pre- and post-monsoon time periods at the sites LP1 (a) and LP2 (b); open and shaded figures represent the samples collected in May and September, respectively. Lined and dotted circles indicate the boundary of isotopic composition in the samples of pelagic and littoral area, respectively.
For a regulated reservoir in the monsoon climate region, lake food webs are not simply based on autochthonous phytoplankton production but are also linked with terrestrial inputs of organic matter. However, the magnitude of the allochthony effect can differ between pelagic and littoral areas as well as the trophic level of consumers. Although summer monsoon storms decrease phytoplankton biomass in a regulated reservoir, the augmented allochthony after the summer monsoon offsets this effect temporarily by subsidizing pelagic zooplankton production. Allochthonous subsidies to herbivorous littoral zoobenthos and piscivorous fishes are, however, relatively limited.
Conclusions
In many reservoirs in a monsoon climate region, basal energy resources fueling lake ecosystems are influenced by the combination of monsoon climate (extremely wet in summer) and dam operations. However, subsequent effects on pelagic and littoral reservoir food web ecosystems remain unclear, because of lack of useful indicators and multiple environmental factors which dilute the impact of summer monsoon. This study overcame the limitations by the combination of natural stable isotopes as tracers to detect ecosystem responses to environmental change and the suitable location, where the summer monsoon impact is apparently significant but other environmental factors are likely negligible.
We assessed responses of pelagic and littoral food webs to the summer monsoon-derived disturbance in primary energy resources in a large reservoir regulated by a large dam. Our results showed that monsoon rains in summer changed the balance of allochthonous and autochthonous carbon sources in the regulated reservoir system, reducing phytoplankton biomass and increasing a pulsed supply of allochthonous materials. Also, the augmented allochthony to basal resources after the summer monsoon offsets this effect temporarily by subsidizing pelagic zooplankton production. Allochthonous subsidies to herbivorous littoral zoobenthos and piscivorous fishes are, however, relatively limited. Our findings may be limited in application to other freshwater reservoirs because of a limited data-set collected in a given year and the site. Further studies on the effect of reservoir hydrology and patterns of stable isotope composition of phytoplankton, zooplankton, and zoobenthos are required together with long-term monitoring efforts.
Acknowledgements
We are grateful to Drs YJ Lee, SY Ha, SK Park, and DH Won for collecting samples of POM, zooplankton, and macroinvertebrates. We thank Prof. Mary Power for insightful comments on the early manuscript and Jeanney McSloy for proofreading.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Balesdent J, Girardin C, Mariotti A.1993. Site-related δ13C of tree leaves and soil organic matter in a temperate forest. Ecology. 74: 1713–1721.
- Boecklen WJ, Yarnes CT, Cook BA, James AC. 2011. On the use of stable isotopes in trophic ecology. Annu Rev Ecol Evol Syst. 42: 411–440.
- Brett MT, Arhonditsis GB, Chandra S, Kainz MJ. 2012. Mass flux calculations show strong allochthonous support of freshwater zooplankton production is unlikely. PLOS One. 7: e39508.
- Bunn SE, Davies PM, Winning M. 2003. Sources of organic carbon supporting the food web of an arid zone floodplain river. Freshw Biol. 48: 619–635.
- Carabel SE, Godínez-Domínguez E, Verísimo P, Fernández L, Freire J. 2006. An assessment of sample processing methods for stable isotope analyses of marine food webs. J Exp Mar Biol Ecol. 336:254–261.
- Carpenter SR, Cole JJ, Pace ML, Van de Bogert M, Bade DL, Bastviken D, Gille CM, Hodgson JR, Kitchell JF, Kritzberg ES. 2005. Ecosystem subsidies: terrestrial support of aquatic food webs from 13C addition to contrasting lakes. Ecology. 86: 2737–2750.
- Cravotta CA. 1997. Use of stable isotopes of carbon, nitrogen, and sulfur to identify sources of nitrogen in surface waters in the lower Susquehanna River Basin, Pennsylvania. (USGS Report, vol. 2497), Denver: USGS.
- Cushing CE, Minshall GW, Newbold JD. 1993. Transport dynamics of fine particulate organic matter in two Idaho streams. Limnol Oceanogr. 38: 1101–1115.
- Dawson TE, Mambelli S, Plamboeck AH, Templer PH, Tu KP. 2002. Stable isotopes in plant ecology. Annu Rev Ecol Syst. 33: 507–559.
- Doi H. 2009. Spatial patterns of autochthonous and allochthonous resources in aquatic food webs. Popul Ecol. 51: 57–64.
- Doi H, Zuykova EI, Shikano S, Kikuchi E, Ota H, Yurlova NI, Yadrenkina E. 2013. Isotopic evidence for the spatial heterogeneity of the planktonic food webs in the transition zone between river and lake ecosystems. Peer J. 1: e222.
- Finlay JC. 2001. Stable-carbon-isotope ratios of river biota: implications for energy flow in lotic food webs. Ecology. 82: 1052–1064.
- Finlay JC, Kendall C. 2007. Stable isotope tracing of temporal and spatial variability in organic matter sources to freshwater ecosystems. In: Michener R, Lajtha K, editors. Stable isotopes in ecology and environmental science (2nd ed). Malden (MA): Blackwell; p. 283–333.
- Fisher SG, Likens GE. 1972. Stream ecosystem: organic energy budget. Bioscience. 22: 33–35.
- Flemer DA. 1969. Chlorophyll analysis as a method of evaluating the standing crop phytoplankton and primary productivity. Chesapeake Sci. 10: 301–306.
- France R, Peters R. 1997. Ecosystem differences in the trophic enrichment of 13C in aquatic food webs. Can J Fish Aquat Sci. 54: 1255–1258.
- Gaedke U. 1992. The size distribution of plankton biomass in a large lake and its seasonal variability. Limnol Oceanogr. 37: 1202–1220.
- Grey J, Jones RI, Sleep D. 2000. Stable isotope analysis of the origins of zooplankton carbon in lakes of differing trophic state. Oecologia. 123: 232–240.
- Grey J, Jones RI, Sleep D. 2001. Seasonal changes in the importance of the source of organic matter to the diet of zooplankton in Loch Ness, as indicated by stable isotope analysis. Limnol Oceanogr. 46: 505–513.
- Gu B, Chapman AD, Schelske CL. 2006. Factors controlling seasonal variations in stable isotope composition of particulate organic matter in a soft water eutrophic lake. Limnol Oceanogr. 51: 2837–2848.
- Hedges JI, Clack WA, Quay PD, Richey JE, Devol AH, Santos UDM. 1986. Composition and fluxes of organic matter in the Amazon River. Limnol Oceanogr. 31: 717–738.
- Kendall C, Silva SR, Kelly VJ. 2001. Carbon and nitrogen isotopic compositions of particulate organic matter in four large river systems across the United States. Hydrol Process. 15: 1301–1346.
- Kim B, Choi K, Kim C, Lee U-H, Kim Y-H. 2000. Effects of the summer monsoon on the distribution and loading of organic carbon in a deep reservoir, Lake Soyang, Korea. Water Res. 34: 3495–3504.
- Kohzu A, Tayasu I, Yoshimizu C, Maruyama A, Kohmatsu Y, Hyodo F, Onoda Y, Igeta A, Matsui K, Nakano T, et al. 2009. Nitrogen-stable isotopic signatures of basal food items, primary consumers and omnivores in rivers with different levels of human impact. Ecol Res. 24: 127–136.
- Lake JL, McKinney RA, Osterman FA, Pruell RJ, Kiddon J, Ryba SA, Libby AD. 2001. Stable nitrogen isotopes as indicators of anthropogenic activities in small freshwater systems. Can J Fish Aquat Sci. 58: 870–878.
- Lau DCP, Sundh I, Vrede T, Pickova J, Goedkoop W. 2014. Autochthonous resources are the main driver of consumer production in dystrophic boreal lakes. Ecology. 95: 1506–1519.
- Marra J. 2002. Approaches to the measurement of plankton production. In: Williams PJLB, Tomas DN, Reynolds CS, editors. Phytoplankton productivity: carbon assimilation in marine and freshwater ecosystems. Abingdon: Wiley; p. 78–108.
- Meybeck M, Cauwet G, Dessery S, Somville M, Gouleau D, Billen G. 1988. Nutrients (organic C, P, N, Si) in the eutrophic river Loire (France) and its estuary. Estuar Coast Shelf Sci. 27: 595–624.
- Nguyen HV-M, Hur J. 2011. Tracing the sources of refractory dissolved organic matter in a large artificial lake using multiple analytical tools. Chemosphere. 85: 782–789.
- [NIER] National Institute of Environmental Research. 2009. Survey on the environment and ecosystem of lakes in the Han River system. Seoul: National Institute of Environmental Research.
- Ock G, Takemon Y. 2014. Effect of reservoir-derived plankton released from dams on particulate organic matter composition in a tailwater river (Uji River, Japan): source partitioning using stable isotopes of carbon and nitrogen. Ecohydrology. 7: 1172–1186.
- Pace ML, Cole JJ, Carpenter SR, Kitchell JF, Hodgson JR, Van de Bogert MC, Bade DL, Kritzberg ES, Bastviken D. 2004. Whole-lake carbon-13 additions reveal terrestrial support of aquatic food webs. Nature. 427: 240–243.
- Park H, Byeon M, Shin Y, Jung D. 2009. Sources and spatial and temporal characteristics of organic carbon in two large reservoirs with contrasting hydrologic characteristics. Water Resour Res. 45: W11418.
- Park H, Cho K, Won D, Lee J, Kong D, Jung D. 2013. Ecosystem responses to climate change in a large on-river reservoir, Lake Paldang, Korea. Clim Change. 120: 477–489.
- Perkins MJ, McDonald RA, Van Veen FJF, Kelly SD, Rees G, Bearhop S. 2014. Application of nitrogen and carbon stable isotopes (δ15N and δ13C) to quantify food chain length and trophic structure. PLOS One. 9: e93281.
- Post DM. 2002. Using stable isotopes to estimate trophic position: models, methods, and assumptions. Ecology. 83: 703–718.
- Post DM, Conners ME, Goldberg DS. 2000. Prey preference by a top predator and the stability of linked food chains. Ecology. 81: 8–14.
- Pulido-Villena E, Reche I, Morales-Baquero R. 2005. Food web reliance on allochthonous carbon in two high mountain lakes with contrasting catchments: a stable isotope approach. Can J Fish Aquat Sci. 62: 2640–2648.
- Rautio M, Mariash H, Forsstrom L. 2011. Seasonal shifts between autochthonous and allochthonous carbon contributions to zooplankton diets in a subarctic lake. Limnol Oceanogr. 56: 1513–1524.
- Riemann B, Simonsen P, Stensgaard L. 1989. The carbon and chlorophyll content of phytoplankton from various nutrient regimes. J Plankton Res. 11: 1037–1045.
- Rosi-Marshall EJ, Wallace JB. 2002. Invertebrate food webs along a stream resource gradient. Freshw Biol. 47: 129–141.
- Thorp JH, Delong MD. 1994. The riverine productivity model: an heuristic view of carbon sources and organic processing in large river ecosystems. Oikos. 70: 305–308.
- Trudel M, Weidel BC, Carpenter SR, Kitchell JF, Vander Zanden MJ. 2011. Rates and components of carbon turnover in fish muscle: insights from bioenergetics models and a whole-lake 13C addition. Can J Fish Aquat Sci. 68: 387–399.
- Vander Zanden MJ, Cabana G, Rasmussen JB. 1997. Comparing trophic position of freshwater fish calculated using stable nitrogen isotope ratios (δ15N) and literature dietary data. Can J Fish Aquat Sci. 54: 1142–1158.
- Vander Zanden MJ, Rasmussen JB. 2001. Variation in δ15 N and δ13 C trophic fractionation: implications for aquatic food web studies. Limnol Oceanogr. 46: 2061–2066.
- Vander Zanden MJ, Vadeboncoeur Y. 2002. Fishes as integrators of benthic and pelagic food webs in lakes. Ecology. 83: 2152–2161.
- Vuorio K, Meili M, Sarvala J. 2006. Taxon-specific variation in the stable isotopic signatures (δ13C and δ15N) of lake phytoplankton. Freshw Biol. 51: 807–822.
- Wang Y, Yu X, Li W, Xu J, Chen Y, Fan N. 2011. Potential influence of water level changes on energy flows in a lake food web. Chin Sci Bull. 56: 2794–2802.
- Webster JR, Meyer JL. 1997. Organic matter budget for streams: a synthesis. J North Am Benthol Soc. 16: 141–161.
- Zohary T, Erez J, Gophen M, Berman-Frank I, Stiller M. 1994. Seasonality of stable carbon isotopes within the pelagic food web of Lake Kinneret. Limnolo Oceanogr. 39: 1030–1043.