
ABSTRACT
Many outdoor education centers, camps, nature centers, and parks nationwide bring thousands of students to designated stream sites each year. Organizations that repeatedly use the same site for stream ecology classes have the potential to negatively impact the aquatic macroinvertebrate community via frequent rearrangement of stream substrate by students. We placed 15 rocks of three size classes within a riffle subjected to outdoor education stream classes and another set of 15 rocks within an unused riffle and measured rock movement weekly from April 2014 to May 2014 within an agricultural stream in central Ohio. We also documented the number and type of aquatic macroinvertebrates colonizing each rock at the end of the six week study. Upstream rock movement was greater in the class site than the unused site. Total rock movement, vertical movement, and lateral movement were greater in the class site than the unused site only during baseflow conditions. Macroinvertebrate abundance, taxa richness, EPT (Ephemeroptera, Plecoptera, Trichoptera) abundance, and clinger abundance were greater in the unused site than the class site. Our results indicate that outdoor education centers can avoid negatively impacting the aquatic macroinvertebrate community and improve student educational experiences by alternating the stream sites at which stream classes are conducted.
Introduction
Substrate plays a pivotal role in the physical structuring and biotic composition of streams. It has been referred to as the stage on which aquatic insects play out their ecological roles (Minshall Citation1984). Erman and Erman (Citation1984) maintained that changes in substrate lead directly to changes in community composition, and the importance of quantifying substrate movement has since been acknowledged (Cobb et al. Citation1992; Townsend et al. Citation1997) and studied (Death and Winterbourn Citation1994; Downes et al. Citation1998; Death Citation2002). Negative effects of substrate movement on aquatic macroinvertebrates have been documented for multiple disturbance types (Boulton et al. Citation1992; Cobb et al. Citation1992; Matthaei and Townsend Citation2000; Death Citation2002; Gjerlov et al. Citation2003). Spates have received the greatest attention as agents of substrate movement, and the magnitude of a spate has been shown to influence the biological response to a spate, although a consistent relationship between spate magnitude and biological response has not been demonstrated (Boulton et al. Citation1992; Matthaei et al. Citation2000). The patchy arrangement of substrate on the streambed and disturbance history of different patches yield areas of streambed that are more prone to substrate disturbance than others (Cobb et al. Citation1992; Downes et al. Citation1998; Matthaei et al. Citation2000). The amount of sunlight and the consequent reliance on algal productivity or availability of leaf litter in a stream may also influence effects of substrate disturbance on stream biota (Death Citation2003).
In addition to natural disturbances, anthropogenic disturbance caused by logging, construction, and nutrient input has also impacted streams and been the focus of much study (Elliott et al. Citation1997; Stanley et al. Citation2010). Water pollution problems that plagued many streams and rivers in the United States throughout the 1960s led to development in the 1970s of laws regulating water quality and point source pollution and launched the modern environmental movement promoting environmental awareness and education. While the environmental movement has helped solve some human-induced environmental problems, it has initiated others. Ecotourism is one outgrowth of the environmental movement and has been defined by the International Union for Conservation of Nature as responsible travel to relatively undisturbed natural areas of the world to enjoy nature, promote conservation, and provide economic benefit for the host community. Several studies have demonstrated negative impact on the environment from recreational activities associated with ecotourism such as the effects of: 1) diving on coral reefs (Santos et al. Citation2015; Giglio et al. Citation2016; Roche et al. Citation2016); 2) hiking on trailside plants (Guo et al. Citation2015); and 3) beachside driving on shorebirds (Weston et al. Citation2014). Such studies are insightful and important because proponents of ecotourism are often unaware about the possibility of environmental harm from ecotourism activities or they proceed under the impression that their activities are environmentally harmless (Wall Citation1997; Priskin Citation2003; Weston et al. Citation2014; Giglio et al. Citation2016). The extent of ecotourism that is sustainable for a given environment remains a debated topic (Lee and Jan Citation2015), but it is a debate that benefits from considerable empirical research into the issue (Priskin Citation2003).
Another development stemming from the environmental movement has been the growth of outdoor education (OE), which is a popular, hands-on educational method conducted at outdoor sites and aimed at fostering appreciation for nature among school children. In the aftermath of the aforementioned water pollution problems in the United States and the subsequent public emphasis on protecting water quality, stream ecology courses became an emphasized component of the OE curriculum. Camps, nature centers, and parks offering OE stream ecology courses frequently include a focus on stream biomonitoring as a method for assessing water quality based on the abundance and diversity of pollution sensitive aquatic macroinvertebrates (i.e. Ephemeroptera, Plecoptera, Trichoptera (EPT), Neotaenioglossa: Pleuroceridae, Coleoptera: Psephenidae, and Neuroptera: Corydalidae) (Firehock et al. Citation1995). Since biomonitoring necessitates stream exploration, students participating in OE stream ecology courses are encouraged to move and flip rocks within the streambed in search of aquatic macroinvertebrates. Due to familiarity, ease of access, and safety considerations, OE organizations typically use the same stream and often even the same site for their stream ecology courses. Over the course of a year the same stretch of stream may consequently incur substrate disruption by thousands of students in search of macroinvertebrates, which may impose a cumulative negative impact on the local macroinvertebrate community. Disturbance of an entire stream reach has been categorized as a Level 3 disturbance, and disruption to discrete patches of streambed or individual rocks qualifies as a Level 4 disturbance (Gore and Milner Citation1990). Thus, the regular use of the same stream reach by OE stream classes may represent Level 3 and/or Level 4 disturbances.
To our knowledge no previous studies have quantified the effect of OE stream classes on substrate movement and aquatic macroinvertebrates within streams, and only a few studies have examined related issues such as impacts of research (Falk Citation1974) and recreational activities (Wright and Li Citation1998; Laing Citation2008; Caires et al. Citation2010) on aquatic macroinvertebrates within steams. Falk (Citation1974) discussed the bias that researchers may introduce into field studies by collecting and vouchering specimens and suggested that researchers in effect become a new top predator in the ecosystem through their collecting efforts. Wright and Li (Citation1998) studied the impact of various outdoor recreational activities (i.e. goldmining, streamside camping, swimming, rafting, and fishing) on Dicosmoecus gilvipes (family: Trichoptera) and found reduced densities of this caddisfly species at disturbed sites. Two studies (Laing Citation2008; Caires et al. Citation2010) examined the effects of humans hiking through streams on stream macroinvertebrate communities. Laing (Citation2008) found that hikers negatively impacted macroinvertebrate abundance and taxa richness in a study of ten tributaries of the Niobrara River in north central Nebraska. Conversely, Caires et al. (Citation2010) found that hikers wading through the stream did not lead to a change in macroinvertebrate abundance or taxa richness in a cobble-bottomed stream in southwestern Utah.
We examined the extent to which stream classes at an OE center in central Ohio cause substrate movement and how such activity may affect the resident aquatic macroinvertebrates. Our hypotheses for this study were:
| (1) | student activity at a riffle used by stream classes will result in greater movement of marked rocks than at an unused riffle and | ||||
| (2) | the class riffle exhibiting greater substrate movement will exhibit reduced macroinvertebrate colonization compared to macroinvertebrate colonization at an unused riffle. | ||||
Methods
Our study was conducted at two sites within the upper Alum Creek, which is a fourth-order stream located in Morrow County in central Ohio, USA. The class site (40° 23′ 01.014″ N, 82° 52′ 37.870″ W) is a 7-meter long riffle that is regularly used as part of stream classes taught by Heartland Outdoor School (Heartland), which is an OE center that provides overnight/multi-day hands-on nature and environmental science programs for K–12 students. Heartland's hands-on educational activities at the class riffle include collection of aquatic macroinvertebrates by use of a dipnet, kick seine, or by turning over rocks, identification of aquatic macroinvertebrates, and measurements of select water chemistry variables. These activities are for educational purposes only and are not part of data collection efforts for a volunteer stream assessment program. The class site is characterized by forested riparian habitat with riparian widths > 30.5 m and bordered by an agricultural field on the right bank. Alum Creek at the class site is a C-4 riffle-pool stream (Rosgen Citation1996) with moderate sinuosity (= 1.3), no constrictions, minimal to no canopy cover, and substrate composed primarily of coarse gravel (D50 = 38% coarse gravel substrate). The unused site (40° 23′ 23.608″ N, 82° 52′ 27.751″ W) is a 21-meter long riffle that served as the control for our study and has never been used for OE stream classes by Heartland or other OE organizations. The unused site is located approximately 1.2 km upstream of the class site. The unused site is also characterized by forested riparian habitat with riparian widths > 30.5 m and an agricultural field bordering the riparian habitat on the right bank. Alum Creek at the unused site is a C-4 riffle-pool stream (Rosgen Citation1996) with moderate sinuosity (= 1.3), no constrictions, minimal to no canopy cover, and substrate composed primarily of coarse gravel (D50 = 36% coarse gravel substrate) with some large boulders.
We selected a set of 15 rocks from the dry streambed at each of the two sites. To ensure that all rocks were free of algae and aquatic macroinvertebrates, each rock was cleaned with a scrub brush and placed on the dry streambank for one week prior to the start of our study. To aid in locating the rocks throughout the study, the rocks were marked with an earth-tone paint to distinguish them from naturally-occurring streambed stones and still maintain an inconspicuous appearance to students participating in OE stream classes. Both sets of 15 rocks were further divided via use of a gravelometer into three size classes consisting of: (1) small = < 91 mm (n = 5); (2) medium = 91–128 mm (n = 5); and (3) large = 129–180 mm (n = 5). Each marked rock was also labeled with a letter and number with a marking pen to identify its site and size class. Rocks were placed with their labels facing down along a transect at the head of the riffle at both sites. Rocks were placed equidistant (0.61 m) from each other along the transects at both sites because the wetted width was the same at both sites at the onset of the study. Placement order for the rocks along the transect was determined randomly using a random number table and the same random pattern was used at both sites.
The study was conducted for six weeks from 19 April 2014 to 31 May 2014 during the busy spring teaching season at Heartland when the largest number of students and greatest number of classes occurs (). A total of 1418 students participated in 71 individual stream classes at the class site during the study period, while the unused site experienced no student activity and no stream classes during the same time period. To avoid biasing student behavior, weekly rock movement measurements were obtained on Saturday or Sunday when stream classes were not in session and students were not present. Rock movement measurements were conducted at the downstream class site first followed by the upstream unused site. To determine direction and distance of movement at each site for each response variable, a tape measure was stretched taut between a fixed point on each bank across the transect to serve as a reference point for distance measurements. A second tape measure was then used to measure movement distances downstream or upstream and left or right from the original starting position for each rock. We also noted whether the rocks were flipped over from their original position, which was evident by face-up labels. Rock flipping was regarded as evidence of vertical movement and measured as one half the maximum planar width of the rock, since this corresponds to the minimum height a rock would have to be lifted off the streambed to be flipped. After all rock movement related measurements were completed all rocks were restored to their original positions along the transect at each site.
Table 1. Heartland Outdoor School schedule for stream classes within Alum Creek, OH, during the spring (mid-April–early June) outdoor education field trip season showing daily class times and a continuous count of classes for a typical week in 2014.
Prior to placing the rocks into the stream at the start of the study, each rock was visually inspected to confirm that no aquatic macroinvertebrates were present. At the end of the six week study each rock was removed from the stream and examined for aquatic macroinvertebrates. All aquatic macroinvertebrates found on each rock were counted, identified on-site to family level, and released.
Statistical analysis
We calculated six response variables to describe rock movement that included total movement (sum of distance moved in all directions), downstream movement (distance moved downstream), upstream movement (distance moved upstream), lateral movement (distance moved toward the right or left bank), and vertical movement (flipping). We also calculated five macroinvertebrate community response variables that included: (1) macroinvertebrate abundance (number of macroinvertebrates found on each rock); (2) taxa richness (number of families found on each rock); (3) EPT abundance (number of Ephemeroptera, Plecoptera, and Trichoptera found on each rock); (4) clinger abundance (number of macroinvertebrates in the clinger mobility guild found on each rock); and (5) dorsoventrally flattened (DVF) clinger abundance (number of macroinvertebrates with DVF body morphology found on each rock). Insect families for which all taxa were reliably considered to be clingers consisted of Heptageniidae (nymphs), Baetidae (nymphs), Hydropsychidae (larvae, pupae), and Psephenidae (larvae, adults) (Voshell Citation2002). The DVF clingers guild consisted of individuals from the families Heptageniidae (nymphs) and Psephenidae (larvae, adults) because of the DVF morphology exhibited by these two families.
Two spate events occurred during this study and provided the opportunity to evaluate the influence of spates on rock movement and macroinvertebrate colonization. Elevated discharge levels during Week 2 (26 April–2 May) and Week 4 (10 May–16 May) resulted in a cancellation of OE stream classes due to the unsafe increase in water velocity and depth. Consequently, no student activity took place at the class site during spate weeks. The average mean daily discharge measured at the USGS gauge located 5.6 km downstream of the class site during spate weeks was 11.7 m3/s with a minimum of 1.4 m3/s and a maximum of 32.1 m3/s. Alum Creek was at base flow conditions during Weeks 1, 3, 5, and 6 and the average mean daily discharge at the USGS gauge during baseflow weeks was 2.4 m3/s with a minimum of 1.0 m3/s and a maximum of 6.9 m3/s. Heartland's stream classes proceeded as usual at the class site during base flow weeks.
Rock movement at the class site during Week 1 was excluded from the statistical analyses of rock movement because the students had discovered the experimental rocks. To correct for this bias in the remaining weeks of the study, we removed the letter labels from the rocks and replaced the letters with a series of less conspicuous coded dots.
We used generalized linear model analyses (Zuur et al. Citation2009) because all rock movement and aquatic macroinvertebrate response variables did not meet the assumptions of normality and/or equal variance. Specifically, we used a three factor (site, rock size class, and hydrologic regime) generalized linear model analysis with the gamma distribution using the glm function (R Core Team Citation2015) to determine the effect of OE classes, rock size class, and hydrologic regime) on rock movement. All rock movement response variables were transformed (X + 0.001) prior to generalized linear model analyses to meet the requirements of the gamma distribution. We report on the effects of site, rock size class, and the interaction effects of site x rock size class, site × hydrologic regime, rock size class × hydrologic regime, and site × rock size class × hydrologic regime on our rock movement response variables. Selective reporting of our results enables us to focus on the most important results related to our research hypotheses and enables us to account for the potential effect of hydrologic regime. Additionally, we used a two factor (site and rock size class) generalized linear model analysis with the glm function and the quasi-Poisson distribution to determine the effect of OE classes and rock size class on macroinvertebrate colonization. A Tukey Test (glht function and multcomp package, Bretz et al. Citation2010) was used to determine differences among means if the generalized linear model analyses indicated that a significant effect occurred. We used R statistical software (R Core Team Citation2015) and a significance level of 0.05 for all statistical analyses.
Results
Substrate movement
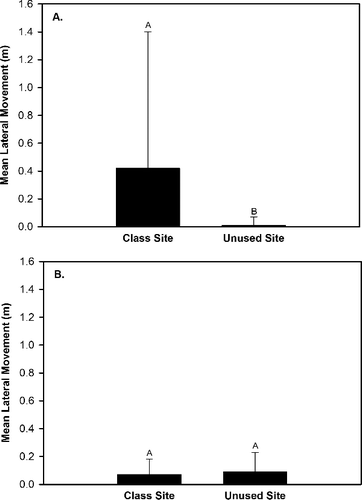
Our results indicated that four substrate movement variables (total movement, upstream movement, lateral movement, vertical movement) exhibited either a significant (P < 0.05) effect of site, an interaction effect of site × rock size class, an interaction effect of site × hydrological regime, and/or an interaction effect of site × rock size class × hydrologic regime (). Upstream movement was greater in the class site that served as the location for OE stream classes than the unused site that was not subjected to OE stream classes (). The generalized linear model analyses indicated that downstream movement, total movement, and vertical movement exhibited a significant interaction effect of site × rock size class (), but the Tukey Test did not identify differences in these three response variables between sites for each of the three rock size classes. Total movement and vertical movement also exhibited a significant interaction effect of site × hydrologic regime (). Total movement, vertical movement, and lateral movement were greater at the class site subjected to OE stream classes than the unused site during base flow conditions and no differences in total movement, vertical movement, and lateral movement occurred between sites during spate conditions (). The generalized linear model analyses indicated that lateral movement exhibited a significant interaction effect of site x rock size class × hydrologic regime (), but the Tukey Test did not identify significant differences (P > 0.05) in lateral movement trends among sites for each rock size class between the two hydrologic regimes. The generalized linear model analyses also indicated that upstream rock movement was significantly influenced (P < 0.05) by rock size class (), but a Tukey Test did not identify any differences in upstream movement among rock size classes.
Table 2. P-values from a three factor (site, rock size class, and hydrologic regime) generalized linear model analysis of rock movement response variables in Alum Creek, OH, from 19 April 2014 to 31 May 2014. Bolded P-values are those that had a significant effect (P < 0.05) on rock movement within the generalized linear model analysis. Abbreviations are: Size – rock size class and HR – hydrologic regime.
Figure 1. Mean upstream rock movement between the class site subjected to outdoor education stream classes and the unused site within Alum Creek, OH, from 19 April 2014 to 31 May 2014.
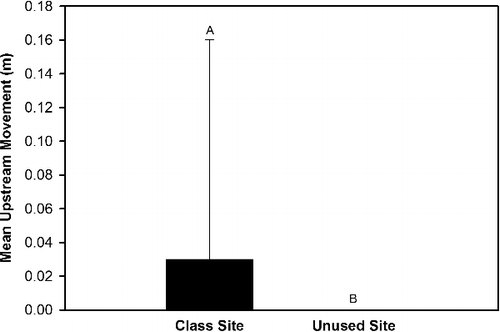
Figure 2. Differences in mean total rock movement between the class site subjected to outdoor education stream classes and an unused site during baseflow conditions (A) and spate conditions (B) within Alum Creek, OH, from 19 April 2014 to 31 May 2014.
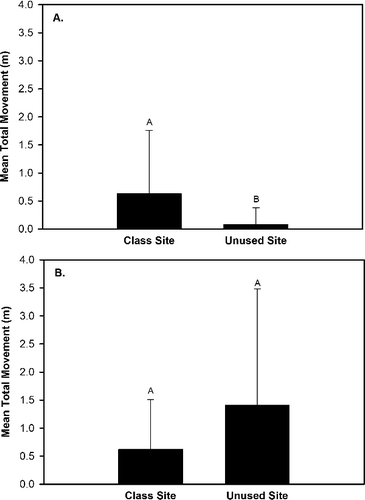
Figure 3. Differences in mean vertical rock movement (i.e. rock flipping) between the class site subjected to outdoor education classes and the unused site during baseflow conditions (A) and spate conditions (B) within Alum Creek, OH, from 19 April 2014 to 31 May 2014.
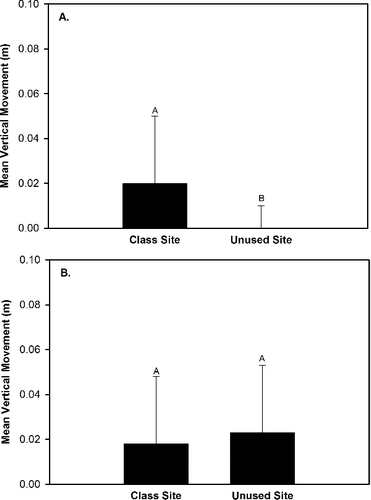
Figure 4. Differences in mean lateral rock movement between the class site subjected to outdoor education classes and the unused site during baseflow conditions (A) and spate conditions (B) within Alum Creek, OH, from 19 April 2014 to 31 May 2014.
Macroinvertebrate colonization
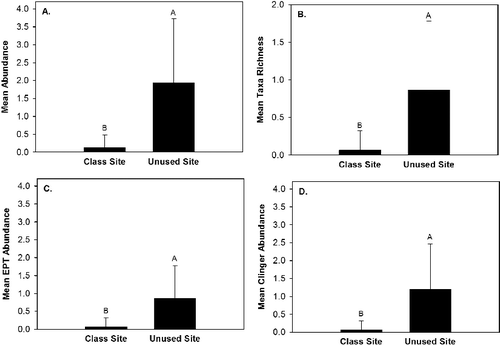
Only two aquatic macroinvertebrate taxa (Tricladida: Planariidae, Ephemeroptera: Heptageniidae) colonized rocks within the class site by the end of the six week study period (). In contrast, seven macroinvertebrate taxa colonized rocks within the unused site and the four most abundant colonizers were (1) Tricladida: Planariidae; (2) Trichoptera: Hydropsychidae; (3) Ephemeroptera: Baetidae; and (4) Coleoptera: Psephenidae (). Macroinvertebrate abundance, taxa richness, EPT abundance, and clinger abundance differed (P < 0.05) between sites (). Mean macroinvertebrate abundance, taxa richness, EPT abundance, and clinger abundance were greater in the unused site than the class site (). DVF clinger abundance did not differ (P > 0.05) between sites or among rock size classes (). The generalized linear model analysis indicated that macroinvertebrate abundance differed among rock size classes and that taxa richness exhibited a significant interaction effect of site and rock size class (), but the Tukey Test did not identify differences in means among the groups of interest.
Table 3. Abundance of aquatic macroinvertebrate families that colonized experimental rocks placed within a class site subjected to outdoor education stream classes and within the unused site within Alum Creek, OH, May 2014.
Table 4. P-values from a two factor (site and rock size class) generalized linear model analysis of macroinvertebrate colonization response variables in Alum Creek, OH, from 19 April 2014 to 31 May 2014. Bolded P-values are those that were documented to have significant effect (P < 0.05) on macroinvertebrate colonization within the generalized linear model analysis. Abbreviations are: EPT – Ephemeroptera, Plecoptera, Trichoptera and DVF – dorsoventrally flattened body form.
Figure 5. Mean macroinvertebrate abundance (A), taxa richness (B), Ephemeroptera, Plecoptera, Trichoptera (EPT) abundance (C), and clinger abundance (D) between the class site subjected to outdoor education stream classes and the unused site in Alum Creek, OH, from 19 April 2014 to 31 May 2014.
Discussion
Substrate movement
The results of our rock movement study confirm our hypothesis that rocks at the site impacted by student activity would move more than rocks at the unused site. The greater mean upstream rock movement, total rock movement, lateral movement, and vertical movement we observed at the class site than at the unused site during base flow conditions are attributable to student activity during OE stream classes. The 1418 participating students over the 71 individual stream classes conducted throughout the study caused weekly rock movement as students searched for aquatic macroinvertebrates during base flow weeks. Our results provide the first known documentation of the impacts of OE stream class on substrate movement.
The hydrologic forces within streams work primarily upon the substrate in a downstream direction and during spate conditions these forces are magnified by increased velocity and discharge. Spates are known to mobilize the streambed and move rocks of varying sizes downstream (Boulton et al. Citation1992; Cobb et al. Citation1992; Downes et al. Citation1998; Matthaei et al. Citation2000; Death Citation2002; Death Citation2003; Gjerlov et al. Citation2003). We also observed increased downstream movement of all rock sizes during spate flow conditions compared to baseflow conditions (Bossley Citation2016). However, upstream rock movement is not intuitive and we attribute it to student activity during the OE stream classes. Only one rock exhibited upstream movement in the absence of student activity. A single medium-sized rock along the right bank at the class site moved 5 cm upstream during Week 4 spate conditions when no stream classes were conducted. With the exception of that single incident, all other upstream rock movement occurred exclusively at the class site during base flow conditions when students were present.
The greater lateral movement of rocks at the class site may also be a result of student activity. Specifically, we believe the greater lateral movement within the class site may have been due to the way students entered the stream and how they proceeded to explore it during stream classes. The entry point for all stream classes within the class site is from a point bar along the left stream bank. During an observational study of stream classes at Heartland (Bossley Citation2016) we observed that some students worked methodically across the riffle from left bank to right bank after entering the stream. Rocks that were moved during that process were more likely to shift toward the right bank in the same direction as the students’ movement. During a return pass from right bank to left bank, students were more likely to move rocks toward the left bank. In comparison, natural base flow at the unused site does not appear to have caused lateral movement of the rocks.
During spate conditions the lack of difference (P > 0.05) in mean vertical movement between the class and unused sites suggests that spates were the primary mechanism for vertical movement in both sites. Conversely, during base flow conditions the increased mean vertical movement within the class site indicates that the students were the primary mechanism for vertical movement within the class site. Additionally, the similarity in mean vertical movement within the class site between spate and baseflow conditions suggests that students have a similar impact as spates on vertical rock movement (). Rock-flipping by students appears to be a function of student stream exploration methods and may have been influenced by whether the Heartland staff member teaching the stream class explained or demonstrated the purpose and benefit of overturning rocks (i.e. the underside of rocks is where many macroinvertebrates reside). Heartland staff members advocated students working together in groups of two or three while using dip nets when class sizes were exceptionally large (i.e. > 20 students per class) (Jon P. Bossley personal observation) due to the limited availability of equipment. During an observational study of Heartland's stream classes (Bossley Citation2016), it was frequently observed that one student would hold a net on the stream bottom while one or two others would move the rocks located directly upstream from the net. We observed that the large rocks exhibited greater percent frequency of rock movement during baseflow conditions than medium and small rocks in the class site, but not within the unused site (). Additionally, all large rocks within the class site were moved during all weeks of baseflow conditions (). These observations indicate that the students exhibited a preference for large rocks. Student preference for large rocks may be due to the greater visibility of the rocks in the stream, the perceived physical challenge they pose to students (i.e. can we flip this big rock?), or the perception that larger rocks must host more macroinvertebrates than smaller rocks.
Table 5. Mean percent frequency (minimum–maximum) in movement of rocks within three size classes between the class site subjected to outdoor education classes and the unused site during baseflow conditions within Alum Creek, OH, from 19 April 2014 to 31 May 2014.
The lack of difference (P > 0.05) in rock movement among rock size classes for all categories of movement was surprising. Other studies have shown differences in rock movement by size class in response to spates (Downes et al. Citation1998), but we observed no significant difference in movement by rock size class even during spate weeks. Rock placement along the transect in each site in our study was determined randomly, but all rocks in the study were equally loose and unembedded as compared to naturally placed rocks, which may have been partially embedded. Thus unrestrained, the human-placed rocks in all size classes in our study may have been equally prone to movement by spates at both sites. Also, since large rocks were moved more frequently () by students, large rocks at the class site experienced regular movement while the large rocks at the unused site remained stable. These counteracting movement tendencies resulted in a wide span in movement for large rocks, and may have prevented us from detecting an effect of rock size.
Macroinvertebrate colonization
The results of our study confirm our hypothesis that the class riffle exhibiting greater substrate movement will exhibit reduced macroinvertebrate colonization compared to macroinvertebrate colonization at an unused riffle. Mean macroinvertebrate abundance, mean taxa richness, mean EPT abundance, and mean clinger abundance were greater at the unused site than the class site. Our study is the first to quantify that OE stream classes can negatively impact aquatic macroinvertebrates within streams.
During every week of the study, marked rocks at the class site were moved as a result of student activity during base flow conditions or increased water velocity during spates. All of the large rocks at the class site were moved every week of the study () making the large rock size class the least stable rock size class within the class site. In contrast, Downes et al. (Citation1998) observed a linear increase in stability with increasing rock size. Rock movement at the unused site was largely confined to the spate events during Weeks 2 and 4. Consequently, the marked rocks at the unused site experienced less frequent disturbance than marked rocks at the class site prior to the end of our study. In contrast, the marked rocks at the class site subjected to OE stream classes experienced more frequent substrate disturbance via student-activity and spates. Significant differences (P < 0.05) in mean macroinvertebrate abundance, mean taxa richness, mean EPT abundance, and mean clinger abundance between the class site subjected to OE stream classes and the unused site at the end of the study are consistent with findings in other studies of anthropogenic disturbance in which disturbance caused a decrease in macroinvertebrate colonization (Clifford Citation1982; Reice Citation1985; Robinson and Minshall Citation1986; Doeg et al. Citation1989; Death Citation1996; Matthaei et al. Citation1996; Wright and Li Citation1998; McCabe and Gotelli Citation2000; Laing Citation2008; Robinson et al. Citation2011). Thus, our results indicate that the greater frequency of student-induced substrate movement as a result of OE stream classes at the class site suppressed macroinvertebrate colonization of rocks.
Conclusions
Our study documents that student activity as part of OE stream classes in the upper Alum Creek led to increased substrate movement at the class site compared to an unused site. Specifically, our substrate movement results indicated that student movement of rocks occurred primarily in three directions – upstream, laterally toward the right or left banks, and vertically (i.e. rock flipping). The assertion by Erman and Erman (Citation1984) that changes in the substrate can lead directly to changes in aquatic macroinvertebrates community structure is corroborated by our findings related to aquatic macroinvertebrate colonization on the rocks. Macroinvertebrate abundance, taxa richness, EPT abundance, and clinger abundance were suppressed on rocks within the class site subjected to frequent movement by student activity as a result of OE stream classes. Our study is the first to quantify that OE stream classes can negatively impact stream substrates and aquatic macroinvertebrates within streams.
The negative impact of frequent OE stream classes on macroinvertebrate colonization has important implications for organizations that conduct OE stream programs. Repetitive use of the same stream site for OE stream classes can diminish biodiversity at the site, which creates a misleading educational experience. Students are taught to assess the overall health of the stream based on results obtained from a class site, but conditions at the class site may not be representative of the stream as a whole. Directors of OE programs and managers of OE centers should consider the value of maintaining multiple stream sites for OE classes and rotating use among them for OE stream classes to avoid overuse of a single site. Given the dearth of information on the environmental impact of OE programs, future research is needed to determine if this impact is influenced by the size and frequency of OE stream classes and whether the impacts differ among stream types.
Acknowledgments
We thank R. Stoakes, S. Fisher, R. Moore, K. Lekies, and D. Denlinger for helpful comments on an earlier draft of this manuscript. We also thank program managers J. Hughes and A. Buzbee of Heartland Outdoor School for their support of the project and permission to access sites on the Heartland Outdoor School property for sampling. We are grateful for H. Nguyen's guidance on statistical analyses. Finally, we thank the National Science Foundation for their support of the research conducted by Jon P. Bossley through a fellowship provided by the grant Linking Watershed Education with GK-12 Education in a Watershed Context.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Notes on contributors
Jon P. Bossley
Jon P. Bossley earned his PhD degree (2016) from the Ohio State University Environmental Science Graduate Program. His research interests include outdoor education and aquatic entomology. He is now an assistant professor of biology at Mount Vernon Nazarene University and a volunteer for the Triplehorn Insect Collection at the Ohio State University Museum of Biological Diversity.
Peter C. Smiley
Peter C. Smiley Jr. is a research ecologist with the USDA Agricultural Research Service specializing in restoration ecology and headwater stream ecology. He received his BA degree in art and biology from Hiram College (1991), his MSc degree in biological sciences from the University of Mississippi (1993), and PhD degree in forest resources from Mississippi State University (2002).
References
- Bossley JP. 2016. Environmental impact from outdoor/environmental education programs: effects of frequent stream classes on aquatic macroinvertebrates [dissertation]. Columbus (OH): The Ohio State University.
- Boulton AJ, Peterson CG, Grimm NB, Fisher SG. 1992. Stability of an aquatic macroinvertebrate community in a multiyear hydrologic disturbance regime. Ecology. 73:2192–2207.
- Bretz F, Hothorn T, Westfall P. 2010. Multiple comparisons using R. Boca Raton (FL): Chapman & Hall/CRC Press.
- Caires AM, Vinson MR, Brasher AMD. 2010. Impacts of hikers on aquatic macroinvertebrates in the North Fork of the Virgin River, Utah. Southwest Nat. 55:551–557.
- Clifford HF. 1982. Effects of periodically disturbing a small area of substratum in a brown-water stream of Alberta, Canada. Freshw Invertebr Biol. 1:39–47.
- Cobb DG, Galloway TD, Flanagan JF. 1992. Effects of discharge and substrate stability on density and species composition of stream insects. Can J Fisheries Aquat Sci. 49:1788–1795.
- Death RG. 1996. The effect of patch disturbance on stream invertebrate community structure: the influence of disturbance history. Oecologia. 08:567–576.
- Death RG. 2002. Predicting invertebrate diversity from disturbance regimes in forest streams. Oikos. 97:18–30.
- Death RG. 2003. Spatial patterns in lotic invertebrate community composition: is substrate disturbance actually important? Can J Fisheries Aquat Sci. 60:603–611.
- Death RG, Winterbourn MJ. 1994. Environmental stability and community persistence: a multivariate perspective. J North American Benthol Soc. 13:125–139.
- Doeg TJ, Lake PS, Marchant R. 1989. Colonization of experimentally disturbed patches by stream macroinvertebrates in the Acheron River, Victoria. Aust J Ecol. 14:207–220.
- Downes BJ, Lake PS, Glaister A, Webb JA. 1998. Scales and frequencies of disturbances: rock size, bed packing and variation among upland streams. Freshwater Biol. 40:625–639.
- Elliott AG, Hubert WA, Anderson SH. 1997. Habitat associations and effects of urbanization on macroinvertebrates of a small, high-plains stream. J Freshw Ecol. 12:61–73.
- Erman DC, Erman NA. 1984. The response of stream macroinvertebrates to substrate size and heterogeneity. Hydrobiologia. 108:75–82.
- Falk JH. 1974. Estimating experimenter-induced bias in field studies: a cautionary tale. Oikos. 25:374–378.
- Firehock K, Marshall L, Pinto I. 1995. Hands on save our streams: the save our streams teacher's manual. Gaithersburg (MD): Izaak Walton League of America.
- Giglio VJ, Osmar JL, Schiavetti A. 2016. Recreational diver behavior and contacts with benthic organisms in the Abrolhos National Marine Park, Brazil. Environ Manag. 57:637–648.
- Gjerlov C, Hildrew AG, Jones JI. 2003. Mobility of stream invertebrates in relation to disturbance and refugia: a test of habitat templet theory. J North American Benthol Soc. 22:207–223.
- Gore JA, Milner AM. 1990. Island biogeographical theory: can it be used to predict lotic recovery rates? Environ Manag. 14:737–753.
- Guo T, Smith JW, Leung YF, Seekamp E, Moore RL. 2015. Determinants of responsible hiking behavior: results from a stated choice experiment. Environ Manag. 56:765–776.
- Laing KD. 2008. Impacts of human physical disturbance on benthic communities in tributaries of the Niobrara national scenic river [thesis]. Lincoln (NE): University of Nebraska.
- Lee TH, Jan FH. 2015. The effects of recreation experience, environmental attitude, and biospheric value on the environmentally responsible behavior of nature-based tourists. Environ Manag. 56:193–208.
- Matthaei CD, Arbuckle CJ, Townsend CR. 2000. Stable surface stones as refugia for invertebrates during disturbance in a New Zealand stream. J North American Benthol Soc. 19:82–93.
- Matthaei CD, Uehlinger U, Meyer EI, Frutiger A. 1996. Recolonization by benthic invertebrates after experimental disturbance in a Swiss prealpine river. Freshw Biol. 35:233–248.
- Matthaei CD, Townsend CR. 2000. Long-term effects of local disturbance history on mobile stream invertebrates. Oecologia. 125:119–126.
- McCabe DJ, Gotelli NJ. 2000. Effects of disturbance frequency, intensity, and area on assemblages of stream macroinvertebrates. Oecologia. 124:270–279.
- Minshall GW. 1984. Aquatic insect-substratum relationships. In: Resh VH, Rosenberg DM, editors. The ecology of aquatic insects. New York (NY): Praeger Publishers; p. 358–400.
- Priskin J. 2003. Tourist perceptions of degradation caused by coastal nature-based recreation. Environ Manag. 32:189–204.
- R Core Team. 2015. R (version 3.2.3): a language and environment for statistical computing. Vienna (AT): R Foundation for Statistical Computing.
- Reice SR. 1985. Experimental disturbance and the maintenance of species diversity in a stream community. Oecologia. 67:90–97.
- Robinson CT, Minshall GW. 1986. Effects of disturbance frequency on stream benthic community structure in relation to canopy cover and season. J North American Benthol Soc. 5:237–248.
- Robinson CT, Blaser S, Jolidon C, Shama LNS. 2011. Scales of patchiness in the response of lotic macroinvertebrates to disturbance in a regulated river. J North American Benthol Soc. 30:374–385.
- Roche RC, Harvey CV, Harvey JJ, Kavanaugh AP, McDonald M, Stein-Rostaing VR, Turner JR. 2016. Recreational diving impacts on coral reefs and the adoption of environmentally responsible practices within the SCUBA diving industry. Environ Manag. 58:107–116.
- Rosgen D. 1996. Applied river morphology. Pagosa Springs (CO): Wildland Hydrology.
- Santos GS, Burgo DC, Lira SMA, Schwamborn R. 2015. The impact of trampling on reef macrobenthos in northeastern Brazil: how effective are current conservation strategies? Environ Manage. 56:847–858.
- Stanley EH, Powers SM, Lottig NR. 2010. The evolving legacy of disturbance in stream ecology: concepts, contributions, and coming challenges. J North American Benthol Soc. 29:67–83.
- Townsend CR, Scarsbrook MR, Doledec S. 1997. The intermediate disturbance hypothesis, refugia, and biodiversity in streams. Limnol Oceanogr. 42:938–949.
- Voshell JR Jr 2002. A guide to common freshwater invertebrates of North America. Blacksburg (VA): McDonald and Woodward.
- Wall G. 1997. Is ecotourism sustainable? Environ Manag. 21:483–491.
- Weston MA, Schlacher TA, Lynn D. 2014. Pro-environmental beach driving is uncommon and ineffective in reducing disturbance to beach-dwelling birds. Environ Manag. 53:999–1004.
- Wright KK, Li JL. 1998. Effects of recreational activities on the distribution of Dicosmoecus gilvipes in a mountain stream. J North American Benthol Soc. 17:535–543.
- Zuur AF, Ieno EN, Walker NJ, Saveliev AA, Dmith GM. 2009. Mixed effects models and extensions in ecology with R. New York (NY): Springer-Verlag.