
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.Abstract
As a consequence of the climate change, localized torrential rainfall has increased in intensity and frequency in Northeast Asia. This work aims at studying the effect of rainfall intensity on benthic macroinvertebrate communities in a mid-sized mountain stream in Korea, where the summer monsoon flood is prevalent. Sampling was conducted along reaches in Gapyeong stream (42 km in length), central Korean Peninsula, from a period of July 2008–October 2009. The sampling collection was carried out within 48 h after every major rain occasion and rainfall intensity was quantified by using the total rainfall amount for three days. Both water velocity and Froude number were found to increase with respect to the increase of rainfall at all study sites. As a consequence, the habitat environment and benthic macroinvertebrate communities were apparently affected by the rainfall intensity along the stream reaches. The species richness and abundance of benthic macroinvertebrates were significantly decreased at a rainfall over 200 mm, while much less decreased at a rainfall below 100 mm. Furthermore, the richness and abundance of species generally decreased with an increase of rainfall and their decrease was more evident at the downstream site, as compared to the upstream and midstream one.
1. Introduction
Global warming is the most likely cause of climate changes that give rise to a number of local, regional and global effects (Hardy Citation2003). Over the last few decades, studies for the relationship between the climate changes and variation of seasonal precipitation in East Asian monsoon regions have been conducted (Lee et al. Citation2004; Yang Citation2006; Kim et al. Citation2014). The monsoon rainy season starts in early June, late June and early July in central China and southern Japan, in Korea and in northern China, respectively. In these regions, over 50% of the annual precipitation is concentrated in the summer monsoon season, in which the degree of rainfall concentration is the most severe (ca. 63%) in the Korean Peninsula (Lee et al. Citation2004; Ding and Johnny Citation2005; Jung et al. Citation2017). Recently, the summer rainfall in these regions has conformed to a different pattern from the past; the rainfall frequency and intensity are increasing (Ping et al. Citation2006; Lee and Bae Citation2011). These changes in the rainfall pattern deteriorate the structure and function of aquatic ecosystems in streams, rivers, lakes, and wetlands (Meyer et al. Citation1999; Poff et al. Citation2002). Rainfall events in the past were typical natural disturbances to which organisms became well-adapted in life history and drift behavior, and responded quickly (Ward Citation1992). However, catastrophic rainfall greatly magnifies the disturbances in streams, leading to fundamental alterations in benthic macroinvertebrate communities (Merigoux and Doledec Citation2004; Snyder and Johnson Citation2006; Bae and Park Citation2009; Ilg et al. Citation2009; Lee and Bae Citation2011).
In lotic ecosystems, aquatic organisms are disturbed by various natural disturbances including flood, typhoon and drought that may subsequently affect the coexistence, distribution and abundance of benthic macroinvertebrates (Ward Citation1992; Thomaz et al. Citation2007). Thus, the effects of precipitation and hydrologic disturbances on benthic macroinvertebrates have been considered as major indicators for the environmental assessment of aquatic ecosystems in Europe and the Americas (Matthaei et al. Citation2000; Poff et al. Citation2002; Merigoux and Doledec Citation2004; Snyder and Johnson Citation2006). However, in the monsoon regions in East Asia, only a few studies have been reported regarding the natural disturbances by rainfall on benthic macroinvertebrate communities (Kwak et al. Citation2004; Bae and Park Citation2009; Lee and Bae Citation2011).
In this study, we investigated the effects of rainfall frequency and intensity on the community structure of benthic macroinvertebrates in a typical mid-sized mountain stream in Korea during two monsoon seasons.
2. Methods
2.1. Study sites and sampling
Gapyeong stream, located in a preserved area in the central Korean Peninsula, is a branch of the Bukhan River (North Han River) with the length of about 42 km (). The stream order is 1–5 using a 1:25,000 scale map. From the upstream to the downstream, the stream becomes wider, the slope becomes slower and the substrate particle size becomes smaller, representing a typical mid-sized mountain stream in this region.
Figure 1. Location of study sites in the Gapyeong stream, South Korea.
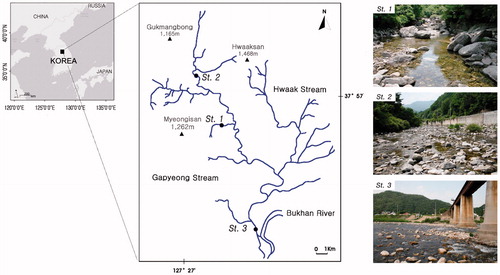
In this study, we investigated the stream width, depth, and substrate (subjectively estimated percentage cover of bedrock, bolder >256 mm, cobble 64–256 mm, pebble 16–64 mm, gravel 2–16mm, coarse sand 0.5–2 mm, fine sand and silt <0.5 mm and detritus) at three sites of the Gapyeong stream (upstream, midstream and downstream reaches). Site 1 (upstream site: 37˚ 58′ 36.4″ N and 127˚ 26′ 35.5″ E) was located on the upstream reach, a second-order branch stream of the Gapyeong stream, with the altitude of about 136 m, the stream width of 3.0 ± 2.8 m (mean ± SD), and the stream depth of 27.5 ± 11.3 cm at the sampling point. The substrate was dominantly occupied by bedrock and boulders, and the streamside was a forest. Site 2 (midstream site: 37˚ 56′ 10.9″ N and 127˚ 28′ 52.5″ E) was located in the midstream reach, a third-order stream of the Gapyeong stream, with the altitude of about 274 m, the stream width of 20.0 ± 14.1 m and the stream depth of 31.1 ± 10.8 cm at the sampling point. The substrate was dominantly occupied by boulders and cobbles, and the streamside was a small resort. Site 3 (downstream site: 37˚ 49′ 53.2″ N and 127˚ 31′ 4.92″ E) was located in the downstream reach near the stream mouth, a fifth-order stream of the Gapyeong stream, with the altitude of about 70 m, with the stream width of 175.0 ± 35.4 m and the stream depth of 35.2 ± 13.0 cm at the sampling point. The substrate was dominantly occupied by cobbles and pebbles, and the streamside was a town.
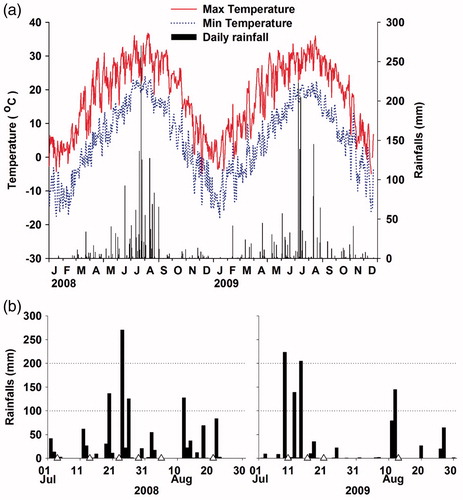
We conducted 13 surveys between July and October in 2008 and 2009 in order to investigate the characteristics of the benthic macroinvertebrate communities depending on the changing rainfall at each site. A contrast of rainfall pattern in terms of frequency and intensity was well represented during the years 2008–2009 in the area (). The survey was performed 10 times during July and August, which is the monsoon period and three times during the dry season. Quantitative sampling was conducted with a Surber sampler (50 × 50 cm, mesh size 0.25 mm, three replicates per site) at each site and collected approximately 20 cm in depth. The collected benthic macroinvertebrates were preserved in 80% ethyl alcohol.
Figure 2. Temperature and rainfall changes during the study period (a) and rainfall events during the rainy season in 2008 and 2009 (b) (Triangles show sampling dates on July 4, 14, 23 and 29 and August 5 and 21 in 2008; July 10, 16 and 21 and August 13 in 2009).
2.2. Monitoring of environmental factors
We used the daily rainfall amount data of automatic weather system (AWS) of the Korea Meteorological Administration (http://www.kma.go.kr), that is located the Site 2. To study the reaction of benthic macroinvertebrates according to varying rainfall amounts during the monsoon season; the sampling was conducted within 48 h (mostly two days after) after a rainfall. The rainfall amount used for the analysis was the total amount of rainfall which accumulated for three days prior to the stopping of the rain. In most rainfall events during study period, the rainfalls lasted less than three days except one case in 2008 (). The water level dropped relatively rapidly and the sampling was possible at that time even after a large amount of rainfall during the study period. The converted rainfall data were grouped in 100 mm increments. The Torricelli’s method was used to measure the flow rate at each survey site during the field sampling (Clanet Citation2000; (EquationEquation (1)(1)
(1) ). The Froude number (Fr) was acquired from the measured velocity (Merigoux and Doledec Citation2004; EquationEquation (2)
(2)
(2) ).
(1)
(1)
(2)
(2)
In these equations, V (cm s−1) denotes the velocity, g the acceleration of gravity which is 980 cm s−2, h (cm) the difference of water height that was calculated using the difference in height between the parallel flow and horizontal direction of the water flow using the 1 m steel ruler and D (cm) the water depth.
2.3. Data analysis
The benthic macroinvertebrates were identified to the species level using available references (Yoon Citation1995; Kawai and Tanida Citation2005; Merritt et al. Citation2008), except for some oligochaete and Diptera groups and counted all of individuals at each sample. The mean species richness (species number per sample) and abundance (individual number per sample) were used for the data analysis. The benthic macroinvertebrate communities were divided into three habitat orientation groups (burrowers, clingers-sprawlers and swimmers) by the classification of Merritt et al. (Citation2008) for the analysis. The PC-ORD software was employed for the ordination analysis (McCune and Grace Citation2002). Nonmetric multidimensional scaling (NMS) analysis was used to determine the number of factors that determined the complexity of the benthic macroinvertebrate community. Rare species <5% of the community were excluded from the analysis (Clarke Citation1993). Multi-response permutation procedures (MRPP) were conducted with the software PC-ORD, a nonparametric protocol for testing the null hypothesis of no differences among groups in the data matrix. Two-way analysis of variance (ANOVA) was conducted to compare the mean species richness and abundance of the benthic macroinvertebrates by the Tukey-Kramer method. p values of less than 0.05 were considered statistically significant for all analyses (SAS Institute Citation2001).
3. Results
3.1. Environmental factors
The annual mean air temperature during the survey period (1 January 2008–31 December 2009) was about 10.6°C (Maximum: 36.8°C, Minimum: −17.5°C) and the mean annual precipitation was about 1700 mm of which 65% was focused on summer, especially the monsoon season from July to August. In 2008, the summer rainfalls (July 1 to August 31) was 1260mm and a relatively small amount of rainfalls continued for two or more days with irregular intervals. In 2009, the summer rainfalls (July 1 to August 31) was 1000.5 mm and relatively large rainfalls often continued for short periods of time (). The survey period was divided into four groups according to the three-day cumulative rainfalls. The first group included almost dry periods, not in the summer monsoon period: 17 October 2008, 30 May, and 12 October 2009. The second group of samples occurred during periods with moderate rainfalls during the summer monsoon period including five sampling days with a mean rainfall of 64.2 ± 19.5 mm (mean ± SD). The third group of samples occurred during periods with a normal rainfall which include July 23 and 29 in 2008 and the mean rainfall was 137.3 ± 14.5 mm. The fourth group of samples during periods with significant rainfall including July 10 and 16 and August 13 in 2009, with a mean rainfall of 220.8 ± 13.0 mm (F3,9=119.691; p > 0.05).
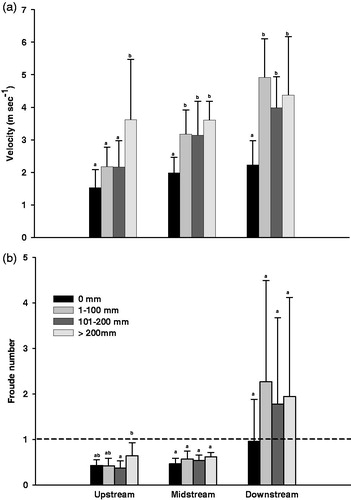
The stream velocity significantly increased as the rainfall increased at all study sites (F3,35=4.390 – 9.055; p < 0.05), but the Fr number increased at only upstream site (F3,35=3.562; p < 0.05) and other sites (F3,35=0.871–1.805; p > 0.05) was not significantly different (). The critical flow (Fr =1) was not exceeded at the upstream and midstream sites despite the high rainfall, but it was always exceeded at the downstream site even with limited rainfall.
Figure 3. Velocity of the study area by Torricelli’s theorem (a) and the Froude number of the study area (b), according to rainfall ranges at the study sites and dotted horizontal line (b) the critical flow. Standard deviation bars with different letters are statistically different (p < 0.05); ab, no statistically significant difference between letters a and b (p > 0.05).
3.2. Benthic macroinvertebrate communities
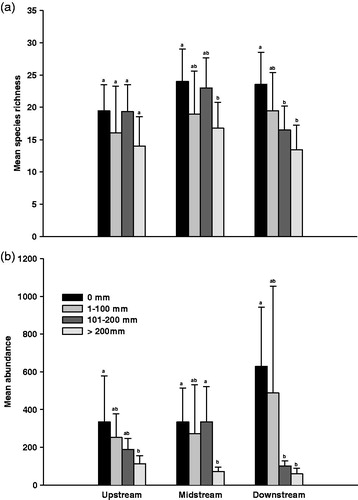
A total of 125 species of benthic macroinvertebrates were identified in 13 samplings. The identification was performed at the species level for all the taxa except for some Diptera. Ninety benthic macroinvertebrate species were analyzed, but rare species with the abundance of less than 5% total site samples were excluded. The mean species richness and abundance of the benthic macroinvertebrates decreased as the rainfall increased at all study sites (). In particular, the mean species richness and abundance at the downstream site decreased more drastically when compared with those of the upstream and midstream sites. In addition, the mean species richness and abundance at the downstream site in cases of rainfalls more than 100 mm of rainfall were less than those of the upstream and midstream sites.
Figure 4. Relationship between the range of rainfall and number of species (a) and individuals (b), according to rainfall ranges at the study sites. Standard deviation bars with different letters are statistically different (p < 0.05); ab, no statistically significant difference between letters a and b (p > 0.05).
The abundance of EPT-group (Ephemeroptera, Plecoptera and Trichoptera) and Diptera were accounted for more than 90% of the all taxa (). The mean abundance of EPT-groups and Diptera according to rainfall changes at each site apparently decreased at all the sites as the rainfall increased (). The mean abundance of Ephemeroptera at the upstream (F3,425=1.14; p > 0.05) and midstream (F3,425=2.23; p > 0.05) sites did not show a significant difference depending on the rainfalls, but it significantly decreased as the rainfall rose at the downstream (F3,425=4.51; p < 0.05) site (). In the case of Plecoptera, the abundance decrease according to the rainfall increase was not significant at the downstream (F3,126 = 0.57; p > 0.05) site, but the abundance decreased at the upstream (F3,126 = 2.16; p > 0.05) and midstream sites with increasing rainfall. In particular, the decrease of the mean abundance was significant at the midstream (F3,126 = 3.03; p < 0.05) site where the rainfall exceeded 200 mm (). The mean abundance of the Trichoptera (F3,360 =1.00–2.18; p > 0.05) and Diptera (F3,126 =0.60–1.80; p > 0.05) apparently decrease but not significant (). With respect to the habitat orientation groups (HOGs), clingers and sprawlers showed a higher mean abundance than the swimmers and burrowers at all study sites (). The mean abundance of each HOG apparently decreased with increasing rainfall at all sites; it decreased to a greater degree by the rainfalls <201 mm at the upstream and midstream sites as well as by the rainfall of 101–200 mm at the downstream site (). In particular, the decreases of the abundance of all HOGs at the downstream site were significant according to the amount of the rainfall (F3,152 –815=– 3.28–4.75; p < 0.05).
Figure 5. Mean abundance of taxa (a) and habitat orientation groups (HOGs) (b) at the study sites.
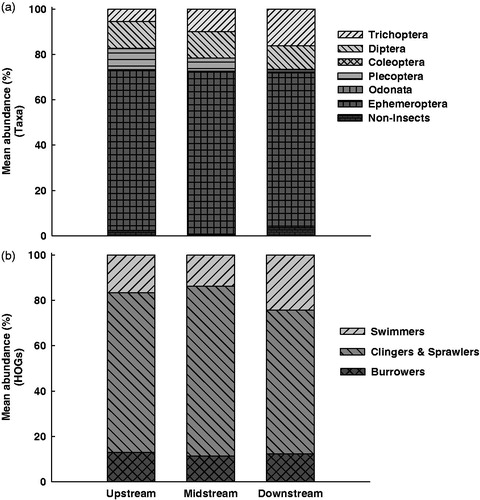
Figure 6. Mean abundance of major benthic macroinvertebrate taxa, Ephemeroptera (a), Plecoptera (b), Trichoptera (c) and Diptera (d), which comprise over 90% abundance of all taxa, according to rainfall ranges at the study sites. Standard deviation bars with different letters are statistically different (p < 0.05); ab, no statistically significant difference between letters a and b (p > 0.05).
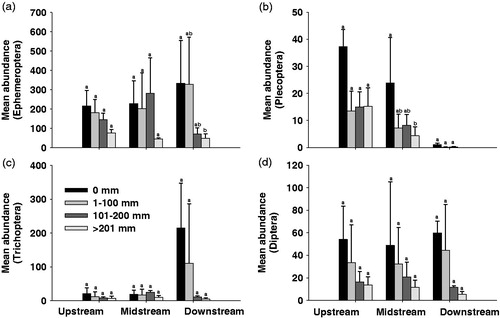
Figure 7. Mean abundance of habitat orientation groups, burrowers (a), clingers and sprawlers (b) and swimmers (c), according to rainfall ranges at the study sites. Standard deviation bars with different letters are statistically different (p < 0.05); ab, no statistically significant difference between letters a and b (p > 0.05).
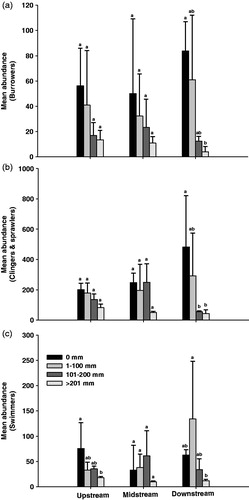
The NMS analysis () was conducted with two qualitative variables (site and year) and two quantitative variables (rainfall and velocity) on the basis of the benthic macroinvertebrate communities at each site. The analytical result showed that the variance of Axis 1 and 2 were 23.0 and 42.9%, respectively. The benthic macroinvertebrate communities were divided into groups of the year 2008 and 2009 by Axis 1. In particular, some parts of the group of the year 2009 were located in the group of the year 2008, which was found to be for the dry period in 2009. The communities were divided into two groups of upstream, midstream, and downstream by Axis 2. Thus, Axis 1 showed the distribution of benthic macroinvertebrate communities according to the rainfall, whereas Axis 2 showed the characteristics of the benthic macroinvertebrate communities at each study site. The MRPP analysis showed that the clusters at the study sites showed a significant difference among them (see the A statistics; ).
Figure 8. Nonmetric multidimensional scaling (NMS) of benthic macroinvertebrate communities (for taxa present in over 5% of samples) at the sites during the study period. Circles: upstream site in 2008 (closed) and 2009 (open); triangles: midstream site in 2008 (closed) and 2009 (open); squares: downstream site in 2008 (closed) and 2009 (open).
Table 1. Results of MRPP to detect differences in species composition between the study sites.
4. Discussion
In Asian monsoon regions, more than 50% of annual precipitation is concentrated in July and August of the monsoon season and there is an increasing trend during recent years in the rainfall frequency and intensity (Ping et al. Citation2006; Oh et al. Citation2009). Our results also showed that over 65% of the rainfall was concentrated on summer season and in particular, the total rainfall in the monsoon season in 2008 was 25% higher than that of 2009. However, the continuous rainfall days during the monsoon season in 2008 was 3.2 ± 2.3 days (mean ± SD) and in 2009 was 1.8 ± 0.9 days, indicating that the rainfall intensity was higher in 2009 than 2008 since rainfalls with more than 100 mm rainfall occurred in 1–2 days in 2009. Increased rainfall has a significant correlation with increased velocity and Fr and this caused the observed decrease in the species richness and abundance of the benthic macroinvertebrate communities.
We showed that the EPT-groups and Diptera, the major benthic macroinvertebrate fauna of the stream, accounted for about 90% of the total community. Because of the characteristics of the stream, the clinger-sprawler group occupied more than 50% of the community. The compositions of benthic macroinvertebrate communities were different at the upstream, midstream and downstream sites, which corresponded with many other studies that the benthic macroinvertebrate communities were dependent on the physical properties of the stream (Wright et al. Citation1983; Ward Citation1992; Rabeni et al. Citation2002).
Our results showed that the species richness and abundance of the benthic macroinvertebrate communities apparently decrease with increasing rainfall. Particularly, the species richness and abundance of the benthic macroinvertebrate community at the downstream site decrease more drastically than those of the upstream and midstream sites even during periods of moderate rainfall. There are three possible explanations for our observations.
First, more branched streams are merged at the downstream site than those of the upstream and midstream sites, which makes the water quantity and the velocity increase more rapidly with increasing rainfall (Giller and Malmqvist Citation1998; Allan and Castillo Citation2007). The downstream site is at a fifth-order stream, so a great amount of water flows in from the branched upstream and midstream reaches during raining. Thus, the velocity and Fr increased with increasing amount of water. Fr is a good indicator of the hydraulic stress on benthic organisms and the value is generally 0.18 or lower in lentic waters and 0.41 or higher in lotic waters (Jowett Citation1993). The species richness and abundance of a benthic macroinvertebrate community are directly affected by the critical flow (Fr =1; Jowett Citation1993; Merigoux and Doledec Citation2004). At the downstream site, even a moderate rainfall results in a value over the critical threshold and we have shown that the species richness and abundance of the benthic macroinvertebrate community were decreases. Quinn and Hickey (Citation1994) conducted a study on the effect of hydraulic factors such as velocity and Fr on the changes in a benthic macroinvertebrate community. They reported that the increased hydraulic stress decreased the species richness and abundance. Suren and Jowett (Citation2006) also found significant positive relationships between stream macroinvertebrate density decrease and flood magnitude.
Second, the stream characteristics constitute one of the critical factors that determine the reaction of benthic macroinvertebrates to rainfalls. A rough substrate of heterogeneous proportions decreases the velocity by forming various turbulences and eddies, providing refuges for benthic macroinvertebrates (Way et al. Citation1995; Matthaei et al. Citation2000). Flooding disturbances to benthic macroinvertebrates are mainly the rolling of the substrate rather than the flow of water (Hart et al. Citation1996). Hence, a flood may move a substrate, but a benthic macroinvertebrate community can be well-adjusted to the general types of flood (Snyder and Johnson Citation2006). In this study, the upstream site dominantly occupied by bed rock and boulders of 256 mm or greater showed more diverse substrate structures than those of the downstream site, dominantly occupied pebbles (16–64 mm) and gravels (2–16 mm). Roughness of the substrate has been previously shown to be correlated with the velocity (Carling Citation1992). The roughness and variety of the substrate structure may have a positive effect on the reaction of the benthic macroinvertebrates to rainfall at the upstream site (Bae and Park Citation2017).
Third, a recovery from disturbances at upstream site, a well-conserved forest, was faster than the downstream site because the immigration of benthic macroinvertebrates from the adjacent branched streams was faster (Lee and Bae Citation2011). Lytle et al. (Citation2008) conducted a study on the relationship between benthic macroinvertebrate response rate (percentage of the population that exhibits flood escape) and catchment size. They reported that, in small catchment, floods tend to occur after longer rainfall events, that they are not a significant mortality threat by floods. In our study, the upstream site is at a second-order stream such as small catchment and the surrounding area is a forest which is better-conserved than the downstream site, with many branched streams nearby. The midstream and downstream sites show the characteristics of lowland streams such as large catchment, passing through the farming areas. In other words, the differences in species composition and abundance may be attributed to the ecological differences (Akankali et al. Citation2014).
In conclusion, increased rainfall frequency may lead to a long-term decrease of aquatic organisms (Ilg et al. Citation2009) but rainfall intensity has an even greater effect on the species richness and abundance of the benthic macroinvertebrate communities than that of rainfall frequency. With regards to the characteristics of the river basin, the benthic macroinvertebrate communities were more affected by the increased rainfall at the downstream reach than at the upstream and midstream reaches because the flow rate increase and disturbances were relatively higher. Generally, the catastrophic floods recovery time tends to be longer than after normal floods (Lee and Bae Citation2011; Gerisch et al. Citation2012). In our study we showed that the communities in the periods with the three-day cumulative rainfall as small as 100 mm or less and the dry periods were not significantly different, whereas the decrease of the species richness and abundance was relatively much more when the rainfall was 200 mm or more. This may also be related to the intermediate disturbance hypothesis that an intermediate disturbance may increase the diversity of an animal community (Townsend and Scarsbrook Citation1997) since the three-day cumulative rainfall of 100 mm or less may belong to a normal disturbance category.
Notes on contributors
Dong Gun Kim, PhD, is an assistant professor of Smith Liberal Arts College at Sahmyook University, Korea. He is broadly interested in benthic macroinvertebrate communities in freshwater ecosystems.
Tae Joong Yoon, PhD, is a research professor in Korean Entomological Institute at Korea University and has worked on the ecology focusing on endangered insect conservation.
Min Jeong Baek, PhD, a post-doctoral researcher of Environmental Safety Group in KIST-Europe, Germany.
Yeon Jae Bae, PhD, is a professor of Division of Environmental Science and Ecological Engineering at Korea University and Director of Korean Entomological Institute. He is broadly interested in systematics and ecology of aquatic insects.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Akankali JA, Abowei JFN, Charles E. 2014. Effects of flooding on Amassoma flood plain benthic macroinvertebrates Niger Delta, Nigeria. Ann Exp Bio. 2:45–52.
- Allan JD, Castillo MM. 2007. Stream ecology. Netherland: Springer.
- Bae MJ, Park YS. 2009. Change in benthic macroinvertebrate communities in response to natural disturbances in a stream. J Ecol Field Biol. 32:197–206.
- Bae MJ, Park YS. 2017. Diversity and distribution of endemic stream insects on a nationwide scale, South Korea: conservation perspectives. Water. 9, 833.
- Carling PA. 1992. The nature of the fluid boundary layer and the selection of parameters for benthic ecology. Freshwater Biol. 28: 273–284.
- Clanet C. 2000. Clepsydrae, from Galilei to Torricelli. Phys Fluids. 12:2743–2751.
- Clarke KR. 1993. Non-parametric multivariate analyses of changes in community structure. Aust J Ecol. 18:117–143.
- Ding Y, Johnny CLC. 2005. East Asian summer monsoon: an overview. Meteorol Atmos Phys. 89: 117–142.
- Gerisch M, Dziock F, Schanowski A, Ilg C, Henle K. 2012. Community resilience following extreme disturbances: the response of ground beetles to a severe summer flood in a Central European lowland stream. River Res Appl. 28:81–92.
- Giller PS, Malmqvist B. 1998. The biology of streams and rivers. Oxford and New York: Oxford University Press.
- Hardy JT. 2003. Climate change. England: John Wiley and Sons Ltd.
- Hart DD, Clark BD, Jasentuliyana A. 1996. Fine-scale field measurement of benthic flow environments inhabited by stream invertebrates. Limnol Oceanogr. 41:297–308.
- Ilg C, Foeckler F, Deichner O, Henle K. 2009. Extreme flood events favour floodplain mollusc diversity. Hydrobiologia. 62: 63–73.
- Jowett IG. 1993. A method for objectively identifying pool, run, and riffle habitats from physical measurements. New Zeal J Mar Fresh Res. 27: 241–248.
- Jung Y, Shin JY, Ahn H, Heo JH. 2017. The spatial and temporal structure of extreme rainfall trends in South Korea. Water. 9, 809.
- Kawai T, Tanida K. 2005. Aquatic insects of Japan: manual with keys and illustrations. Kanagawa, Japan: Tokai University Press.
- Kim DG, Cha YL, Choi LJ, Kang HJ, Baek MJ, Kim JG, Bae YJ. 2014. Drought effects on the colonization of benthic macroinvertebrate communities in the early successional phases in experimental mesocosm wetlands. J Freshwater Ecol. 29:507–524.
- Kwak IS, Song MY, Chon TS. 2004. The effects of natural disturbances on benthic macro-invertebrate. Korean J Limnol. 37:87–95.
- Lee DR, Kim UT, Yoo C. 2004. Climate change impacts on meteorological drought and flood. J Korea Water Resour Assoc. 37:315–328.
- Lee HG, Bae YJ. 2011. Recovery of aquatic insect communities after a catastrophic flood in a Korean stream. Anim Cells Syst. 15:169–177.
- Lytle DA, Bogan MT, Finn DS. 2008. Evolution of aquatic insect behaviours across a gradient of disturbance predictability. Proc Biol Sci. 275:453–462.
- Matthaei CD, Arbuckle J, Townsend CR. 2000. Stable surface stones as refugia for invertebrates during disturbance in a New Zealand stream. J N Am Benthol Soc. 19:82–93.
- McCune B, Grace JB. 2002. Analysis of ecological communities. MjM Software Design: Gleneden Beach, OR.
- Merigoux S, Doledec S. 2004. Hydraulic requirements of stream communities: a case study on invertebrates. Freshwater Biol. 49:600–613.
- Merritt RW, Cummins KW, Berg MB. 2008. An introduction to the aquatic insects of North America. 4th ed. Dubuque, Iowa, USA: Kendall/Hunt.
- Meyer JL, Sale MJ, Mulholland PJ, Poff NL. 1999. Impacts of climate change on aquatic ecosystem functioning and health. J Am Water Resour Assoc. 35:1373–1386.
- Oh TS, Kim MS, Moon YI, Ahn JH. 2009. An analysis of the characteristics in design rainfall according to the data periods. J KOSHAM. 9:115–127.
- Ping F, Zhexian L, Jianhua J. 2006. Differences between dynamics factors for interannual and decadal variations of rainfall over the Yangtze River valley during flood seasons. Chin Sci Bull. 51: 994–999.
- Poff NL, Brinson MM, Day JW. 2002. Aquatic ecosystems and global climate change: potential impacts on inland freshwater and coastal wetland ecosystems in the United States. Arlington, VA: Pew Center on Global Climate Change.
- Quinn JM, Hickey CW. 1994. Hydraulic parameters and benthic invertebrate distributions in two gravel-bed New Zealand rivers. Freshwater Biol. 32: 489–500.
- Rabeni CF, Doisy KE, Galat DL. 2002. Testing the biological basis of a stream habitat classification using benthic invertebrates. Ecol Appl. 12:782–796.
- SAS Institute. 2001. PROC user’s manual. 6th ed. Cary, NC: SAS Institute.
- Snyder CD, Johnson ZB. 2006. Macroinvertebrate assemblage recovery following a catastrophic flood and debris flows in an Appalachian mountain stream. J N Am Benthol Soc. 25:825–840.
- Suren AM, Jowett ID. 2006. Effects of floods versus low flows on invertebrates in a New Zealand gravel-bed river. Freshwater Biol. 51: 2207–2227.
- Thomaz SM, Bini LM, Bozelli RL. 2007. Floods increase similarity among aquatic habitats in river-floodplain systems. Hydrobiologia. 579:1–13.
- Townsend C, Scarsbrook MR. 1997. The intermediate disturbance hypothesis, refugia, and biodiversity in streams. Limnol Oceanogr. 42: 938–949.
- Ward JV. 1992. Aquatic insect ecology. New York, NY: John Wiley.
- Way CM, Burky AJ, Bingham CR, Miller AC. 1995. Substrate roughness, velocity refuges, and macroinvertebrate abundance on artificial substrates in the lower Mississippi River. J N Am Benthol Soc. 14:510–518.
- Wright JF, Hiley PD, Cameron DC, Wigham ME, Berrie AD. 1983. A quantitative study of the macroinvertebrate fauna of five biotypes in the River Lambourn, Berkshire, England. Arch Hydrobiol. 96:271–292.
- Yang HK. 2006. Impact of climate change on hydrological disasters. J Korean Geomorophol Assoc. 13:45–54.
- Yoon IB. 1995. Aquatic insects of Korea. Seoul, Korea: Jeonghaengsa.