
Abstract
Poyang Lake, the largest freshwater lake in China, has experienced a large decrease in wetland area due to long-term overutilization and especially after the hydrological drought events from 2003. This outdoor study was conducted to elucidate the impacts of drought and high concentrations of N on Vallisneria spinulosa. Compared with the 60 cm water depth, the 30 cm water depth caused no difference except lower leaf weight and greater winter bud number per plant growing in the sediment from Poyang Lake. After 400 mg/L NH4+-N was added to the sediment, the leaf weight, root weight, root length, winter bud number, ramet number and stolon weight of V. spinulosa declined greatly. The high NH4+-N content in sediment increased the influence of low water depth on V. spinulosa expressed as the relatively light fresh weight per winter bud of 0.11 g. The results indicated that both a low water depth in spring and high N concentration impacted V. spinulosa and the high concentration of NH4+-N limited the adaptation of V. spinulosa to the decline in water depths. Thus, besides water-level management, nutrient control is also required to protect and restore aquatic macrophytes in Poyang Lake.
Introduction
Poyang Lake, the largest freshwater lake in China, is located on the south bank of the Yangtze River, in northern Jiangxi Province, China. Poyang Lake receives inflow from five main tributaries, including the Ganjiang River, Fuhe River, Xinjiang River, Raohe River and Xiushui River. It regulates and stores water by diverting some of this incoming water into the Yangtze River through an estuary. The lake, its catchment and the Yangtze River form a natural lake-catchment-river system. The complex system comprises important ecological functions, such as serving as an important international wetland that is listed in the Ramsar Convention on Wetlands in 1992 (Zhao et al. Citation2007). Large seasonal variations occur in both lake storage and lake level. During the flood season (April to September), the area of Lake Poyang expands to more than 3000 km2 and the average water depth reaches 8.4 m (Zhao and Liu Citation2017). The lake area shrinks to less than 1000 km2 during the dry season (October to March). The Lake undergoes seasonal water-level fluctuation of 11 m with a change of water level from 9 to 20 m (Zhao and Liu Citation2017).
Due to environmental changes, Lake Poyang has experienced frequent hydrological drought events in past decades, resulting in a sharp decrease in the biodiversity in Poyang Lake Wetland. Compared with the 1980s, Lake Poyang has experienced a large decrease in wetland area due to long-term overutilization, especially after the hydrological drought events from 2003 (Zhang et al. Citation2017). In the spring and summer, aquatic plants move towards the lake center in the Poyang Lake Wetland with the decreasing water level, resulting in a decline of their growing space. The aquatic plants with good tolerance for low water levels and contaminants expanded rapidly, while other species such as Potamogeton malaianus and V. spinulosa declined because of their sensitivity to such environment changes (Hu et al. Citation2010, Citation2015). According to a field survey, the aquatic higher plant community declined to from 8 in 1984 to 3–4 in 2013, indicating that a significant degradation of the lake ecosystem has occurred (Hu et al. Citation2015). In the 1980s, the most common submerged plants were Potamogeton malaianus, Vallisneria spiralis and Hydrilla verticillata in Lake Poyang. Since then these plant species decreased constantly and eventually disappeared (Hu et al. Citation2015).
In the past 30 years, especially after 2004, the water quality of Lake Poyang has been significantly degraded because of the annual increases in pollution loads from industrial, agricultural and urban sources (Gao et al. Citation2014). Eutrophication originating from excessive nutrient loading is a serious environmental problem which leads to the degradation of lake ecosystems. Sediment is considered to be a major nutrient source for rooted submerged macrophytes, including V. spinulosa (Ye et al. Citation2009). Many researchers have confirmed that sediment had a profound effect on both the growth and reproduction of the plants, as well as on their population establishment (Low et al. Citation2001). Nitrogen is an essential nutrient for primary production in aquatic ecosystems, and is primarily released from sediment as NH4+-N. V. spinulosa, a previously dominant submerged macrophyte in Lake Poyang, has been widely distributed in China. It has strong adaptability and can grow in a wide range of sediment conditions, which is a useful trait where environmental changes and human activities have degraded sediments. In short, aquatic macrophytes have declined substantially since 2003 mainly owing to the hydrological drought in Poyang Lake. At the same time, economic development had created excessive nutrient loading in Poyang Lake, resulting in eutrophication, a serious environmental problem. It is assumed that the excessive nutrients in sediment and drought will have significant joint influences on V. spinulosa. Therefore, the major objective of this study was to investigate the response of V. spinulosa in morphology and winter buds to nutrient concentration and water level ().
Figure 1. The leaf weight (mean ± SE) of V. spinulosa growing on different sediments at two water depths. Different small letters above columns indicate significant differences between treatments.
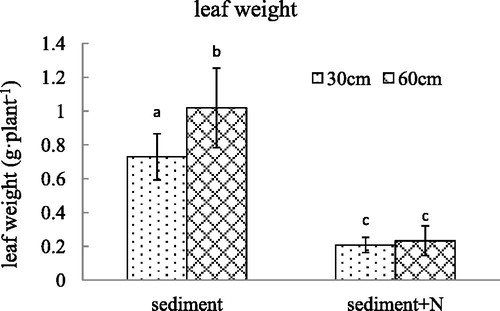
Figure 2. The root weight (mean ± SE) of V. spinulosa growing on different sediments at two water depths. Different small letters above columns indicate significant differences between treatments.
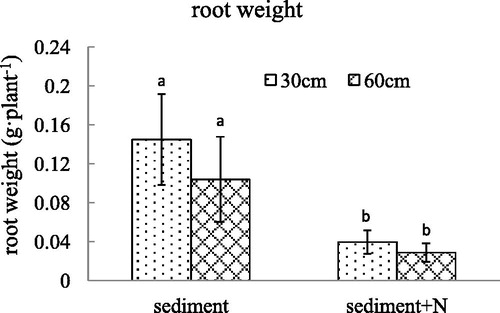
Figure 3. The root length (mean ± SE) of V. spinulosa growing on different sediments at two water depths. Different small letters above columns indicate significant differences between treatments.
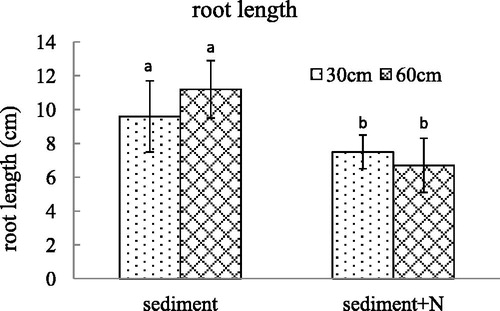
Figure 4. The ramet number (mean ± SE) of V. spinulosa growing on different sediments at two water depths. Different small letters above columns indicate significant differences between treatments.
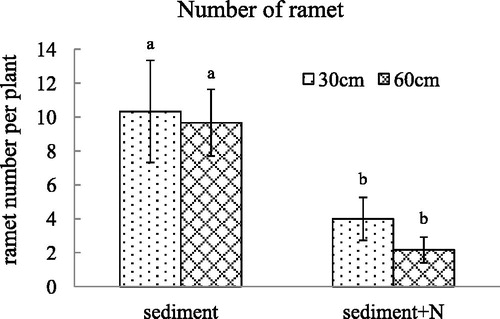
Figure 5. The stolon weight (mean ± SE) of V. spinulosa growing on different sediments at two water depths. Different small letters above columns indicate significant differences between treatments.
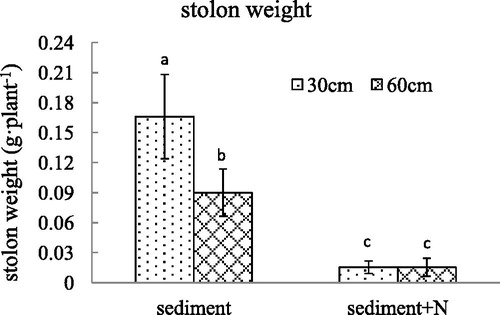
Figure 6. The winter bud number (mean ± SE) per V. spinulosa growing on different sediments at two water depths. Different small letters above columns indicate significant differences between treatments.
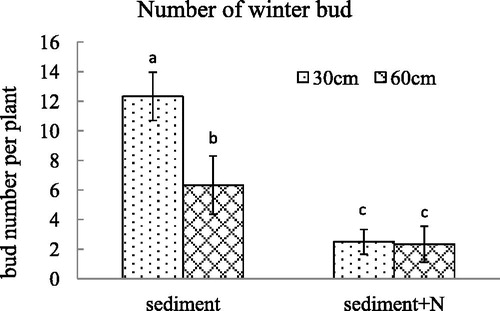
Figure 7. The fresh weight per winter bud (mean ± SE) of V. spinulosa growing on different sediments at two water depths. Different small letters above columns indicate significant differences between treatments.
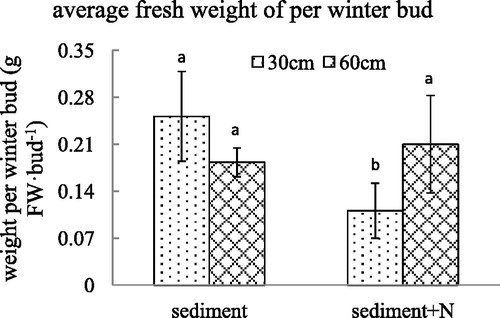
Figure 8. The total weight of winter buds (mean ± SE) per V. spinulosa growing on different sediments at two water depths. Different small letters above columns indicate significant differences between treatments.
Materials and methods
Materials
The sediment was collected from the shore of Lake Poyang in Xingzi City, China. Prior to the experiment, the sediment was dried, sieved (mesh size of 5.0 mm), and mixed to ensure homogeneity.
The seed of Vallisneria spinulosa found in Lake Poyang were planted in the rectangular container with the sediment obtained from the shore of Lake Poyang in spring. Two months later, the seedlings of similar size were selected for the experiment.
Experimental design
A randomized block 2 × 2 factorial experiment (water depths, sediment nutrient) was designed at Laboratory of Poyang Lake and Wetland Ecosystem Research Chinese Ecosystem Research Network located at the shoreline of Lake Poyang from June 15 to October 24, 2017. All treatments were run in triplicate. In this experiment, two substrates were used to grow V. spinulosa. Six sediment cores in Poyang Lake were collected and the total N (TN) content varied from 701.7 to 1294 mg/kg (Yang et al. Citation2015). Nitrogen is an essential nutrient for primary production in aquatic ecosystems and is primarily released from sediment as NH4+-N. According to Wang et al. (Citation2009), the addition of 400 mg/kg NH4+-N to sediments has had adverse effects on another submerged macrophytes, Myriophyllum spicatum. We expected that 400 mg/kg NH4+-N in sediments would be negatively affect the growth of V. spinulosa.
Substrate 1 was the unamended sediment taken from the shore of Lake Poyang. Substrate 2 was the Lake Poyang sediment amended with ammonium-nitrogen (NH4Cl, 400 mg N/kg). Two-month-old seedlings (26.2 ± 2.0 cm) were transplanted into 12 outdoor polyethylene buckets (height 70 cm, top diameter 56 cm and bottom diameter 44 cm), with 2 plants in each bucket. Each bucket held 160 L of tap water and a 10 cm layer of sediment covered by 3 cm of sand in order to prevent or minimize the N from dissolving in the water. Each bucket contained approximately 14 kg of sediment. After 2 weeks of incubation, water levels were adjusted to two treatment levels. Before the change of water level, the plant length of V. spinulosa growing in the unamended sediment was 32.27 ± 3.3 cm, while plant length in the other treatment was 29.3 ± 2.7 cm. Half of the buckets were kept at 60 cm water level depth and the other half were kept at 30 cm depth. To decrease the effects of algae on V. spinulosa in all buckets, algae were removed using a net manually once a day during July and August. Tap water was added to buckets once a week to maintain a constant water-level despite evaporation.
Measurements
After four months of growth, V. spinulosa were harvested and washed under gently running water. The mother plant was picked out and measured to determine the leaf length and root length. The leaves, stolon and roots of V. spinulosa were separated to measure their respective weights. The winter buds were collected from sediment and washed as well. The winter buds were gently dried on tissue paper for determining their individual fresh weight. Dry weights of roots, leaves, stolon and total winter buds per plant were determined after drying to a constant weight for at least 48 h at 70 °C.
Two-way analysis of variance (ANOVA) was used to determine whether substrate and water depth had joint effects on growth and reproduction of V. spinulosa using SPSS software. Differences were considered significant at p < 0.05.
Results
Leaf and root growth
The leaf weight per plant declined significantly after the additional N was added in the sediment. For the sediment without additional N, the water depth of 60 cm is more beneficial to leaf growth of V. spinulosa expressed by leaf weight compared with 30 cm of water depth. The addition of N eliminated the influence of water depth on leaf weight of V. spinulosa (p = 0.015). Similar to the leaf weight, root weight decreased sharply when N was added in the sediment. The root length also declined substantially when N was added in the sediment. The decline in water depth had no effect on the length and weight of roots. There was no interaction between nitrogen content in sediment and water depth on the root length and root weight (p = 0.089, p = 0.276).
Ramet growth
Under the unamended sediment from Lake Poyang, the stolon weight increased after the water depth declined to 30 cm. After N was added to the sediment, there was no significant difference in stolon weight of V. spinulosa planted in the two kinds of sediments. There was significant interaction between high N content in sediments and water depth on stolon weight (p = 0.001). The ramet number of V. spinulosa planted in the sediment with high N content was less than that of V. spinulosa growing in the sediment without additional N. There is no interaction between nitrogen content in sediment and water depths on the ramet number of V. spinulosa (p = 0.471).
Winter bud
For the unamended sediment from Lake Poyang, the decline in water depth caused an increase in winter bud number per plant and total weight of winter buds per plant. However, after the N was added in the sediment, there was no difference in the number or total weight of winter buds per plant between the 60 cm and 30 cm water depth. For the unamended sediment, the decline in water depth did not caused a difference in the weight per winter bud. The winter bud weight per plant generated under a water depth of 30 cm was smaller than that under a water depth of 60 cm with additional N in sediments. The total weight of winter buds per V. spinulosa growing on sediment with high N content decreased dramatically and there was no difference between water depth of 60 and 30 cm. The high N content in sediment altered the influence of water depth on winter bud number, fresh weight per winter bud and total weight of winter buds per V. spinulosa (p < .001, p = .001, p < .001, respectively).
Discussion
Responses to water depths and substrates
The results indicate that water depth and nutrient contents in sediments influenced not only the plant growth but also asexual reproduction of V. spinulosa. In this research, nutrient content in sediments exhibited more significant effects on V. spinulosa, compared to water depth. Before the change of water level, the plant length of V. spinulosa growing on the unamended sediment was 32.27 ± 3.3 cm, while plant length of the other V. spinulosa was 29.3 ± 2.7 cm. It is for this reason that the adjustment of water depth started after the seedlings were planted in the treated sediments for 2 weeks of incubation. The water depth and nutrient content in sediment had extremely significant joint influences (p < 0.05) on leaf weight, stolon weight, fresh weight of each winter bud, total weight of winter bud per plant and winter bud number of V. spinulosa. The high nutrient levels (400 mg/L NH4+-N) in amended sediments probably affected the growth and reproduction of the submerged macrophyte. In this investigation, root morphology changes mainly included the root dry weight and root length. Because N is an essential element for plant growth, moderate concentrations of N should promote its growth (John et al. Citation1999). Root morphological plasticity is considered an important adaptive mechanism for resource acquisition in fertile sediment because when nutrients are abundant in sediments, there is no need for plants to develop large root systems to absorb nutrients (Xu et al. Citation2016). However, in this research, the high N content in sediments also greatly influenced the leaf weight of V. spinulosa, which may be related to the reason that the plant growth was inhibited by adding 400 mg/L NH4+-N in sediments. Submerged macrophytes are highly sensitive to NH4+ toxicity at high N concentrations. Many researchers hold that high concentrations of NH4+ have a direct toxic effect on submerged macrophytes (Cao et al. Citation2004). Under NH4+ stress, most plants show slow growth rates and oxidative damage (Nimptsch and Pflugmacher Citation2007). In other research, it was found that a high concentration of NH4+ inhibited photosynthesis.
In this research, the water depth did not alter the influence of sediment on the growth of V. spinulosa roots. However, the leaf weight declined after the water level changed to 30 cm from 60 cm. Many researchers also found that the submerged macrophyte V. spinulosa allocated more biomass to the leaves in deep water to acquire more light, because the effect of water depth on the growth was an indirect effect of lack of light (Thouvenot et al. Citation2013). In this research, the biomass transfer from leaf to root occurred when the water depth was lowered to 30 cm. It is partly because V. spinulosa tried its best to acquire light to reach the optimum state and partly because V. spinulosa leaves were exposed to air, resulting in leaf damage.
The ammonia nitrogen in sediment not only influenced the leaf and root growth of V. spinulosa, but also affected its asexual reproduction. In the unamended sediment, the low water depth increased the number of winter buds per plant without significant difference in fresh weight of each winter bud. Similarly, the decrease of water depth caused an increment in stolon weight per plant with the same ramet number per plant. The results indicated that the decrease in water depth influenced asexual reproduction of V. spinulosa, which is an adaptation to the change in water depth. Yuan’s et al. (Citation2012) discovery that V. spinulosa produced fewer tubers at greater depth or shade unfavorable conditions are consistent with this result. After NH4+-N was added to the sediment, the ramet number, stolon weight and winter bud number declined greatly, and there was no significant difference between 30 and 60 cm water depth. A decline in water depth led to a decrease in winter bud number per plant growing in the unamended sediment, generating no difference in average fresh weight of each winter bud. The high content of NH4+-N enhanced the effect of water depth on fresh weight per winter bud of V. spinulosa. When V. spinulosa was growing on the sediment with high content of ammonia nitrogen, not only the winter bud number but also its fresh weight declined radically after the water depth was lowered to 30 cm, which means that the high N content inhibited the flexibility of V. spinulosa in the water depth decline. Winter buds which are smaller produce smaller plants the following spring. In an adverse environment, plants arising from large winter buds will have the best opportunity for survival (Korschgen et al. Citation1997), which indicates that the combination of high N content and low water depth had a strong impact on V. spinulosa reproduction.
Implication for macrophyte management
The morphological plasticity and asexual reproduction adaptation of V. spinulosa are important features which could explain its high tolerance to adverse environments. For the sediment from Poyang Lake, the low water depth caused a decline in leaf weight and little influence on the root growth of V. spinulosa. V. spinulosa adapted to the decline in water depth by increasing the stolon weight and winter bud number. As is well known, the drought period of Poyang Lake has been growing longer and starting earlier each year. Frequent occurrence of spring droughts for Poyang Lake resulted in the drought conditions at the early growth stage of submerged macrophytes (Gao et al. Citation2014). At the early growth stage, V. spinulosa can adapt to the low water depth of 30 cm by asexual reproduction. V. spinulosa is able to exist under drought conditions even in low water depths. With economic development, the content of N and phosphorus in Poyang Lake has increased and antibiotic contaminants are present in Poyang Lake (Ding et al. Citation2017). Ammonia nitrogen is the main form of nitrogen in sediment. The results of this research imply that high contents of ammonia nitrogen in sediment amplify the influence of water depth decline on V. spinulosa. However, many factors have not been taken into account in this study, such as the influence of water velocity or water-level fluctuation on the growth and asexual reproduction. All the other factors would likely aggravate the influence of decline in water depth on V. spinulosa.
The numerous shallow lakes in the middle and lower reaches of the Yangtze River in China are eutrophic where the submerged macrophytes disappeared (Wang et al. Citation2012). Therefore, the restoration of submerged macrophytes has been proposed as an important ecological measure for the rehabilitation of degraded lake ecosystem and improvement of water quality of shallow eutrophic lakes. Due to environmental changes, drought in Lake Poyang in winter became more frequent from 2003 to 2013 than from 1960 to 2003 (Zhang et al. Citation2017). The drought was considered as one of the most important factors leading to the degeneration of the Poyang Lake Wetland (Hu et al. Citation2010). Water-level management was proposed by many scholars to maintain suitable water levels in the spring and winter drought period (Ge et al. Citation2010). At the same time, nutrient content is one of the important factors that restricted the restoration of submerged macrophytes. Thus, In order to protect and restore aquatic macrophytes like V. spinulosa in Poyang Lake, not only water-level management, but also nutrient control is necessary. Other factors would intensify the influences of low water depths on aquatic macrophytes even if their own impacts were not as high as shown.
Conclusion
A low water depth in spring had a negative influence on leaf growth but little influence on root growth of V. spinulosa and greatly affected its asexual reproduction. The high content of nitrogen in sediment had serious impacts on the growth and asexual reproduction of V. spinulosa. The high concentration of NH4+-N increased the decline in average fresh weight per winter bud caused by a low water depth, and limited the adaptation of V. spinulosa to the decline in water depths. Thus, besides the water-level management, the nutrient control is also required to protect and restore aquatic macrophytes in Poyang Lake.
Acknowledgements
The study was processed in the Laboratory of Poyang Lake and Wetland Ecosystem Research Chinese Ecosystem Research Network research platform. The authors thank Yuwei Chen and Qi Zhang for providing experimental platform and technical assistance.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
Caixia Kang worked in East China Jiaotong University, China in 2015. She is working in the field of restoration technologies on eutrophic waters.
Xingzhao Dai is the professor of Jiangxi Academy of Agricultural Sciences, China. In his laboratory, researches mainly related to the planning and management of watershed, and GIS. He have published about 20 related papers and the relevant researches won the second class prize of Jiangxi province advanced science and technology.
Zhengong Tong is a professor of East China Jiaotong University, China. His current and previous interests are the purification techniques for surface water, the advanced treatment technologies for wastewater and their application, restoration of environmental ecology. Lots of relevant high-quality paper have been published and the technological invention won second award in Jiangxi Province.
Yunlu Shen is the graduate student in East China Jiaotong University, China.
References
- Cao T, Ni LY, Xie P. 2004. Acute biochemical responses of a submersed macrophyte, Potamogeton crispus L., to high ammonium in an aquarium experiment. J Freshwater Ecol. 19(2):279–284.
- Ding HJ, Wu YX, Zhang WH, Zhong JY, Lou Q, Yang P, Fang YY. 2017. Occurrence, distribution, and risk assessment of antibiotics in the surface water of Poyang Lake, the largest freshwater lake in China. Chemosphere. 184:137–147.
- John PML, Frank BJM, H. van der P W, Cornelis WPMB. 1999. Effects of sediment type and water level on biomass production of wetland plant species. Aquat Bot. 64:151–165.
- Gao JH, Jia JJ, Kettner AJ, Xing F, Wang YP, Xu XN, Yang Y, Zou XQ, Gao S, Qi SH, et al. 2014. Changes in water and sediment exchange between the Changjiang River and Poyang Lake under natural and anthropogenic conditions, China. Sci Total Environ. 481:542–553.
- Ge G, Ji WT, Liu CL, Xiong S, Wu ZQ. 2010. Hydraulic project and wetland ecological protection in Poyang lake. Resourc Environ Yangtze Basin. 19(6):606–613. in Chinese)
- Hu ZP, Ge G, Liu CL, Chen FS, Li S. 2010. Structure of Poyang lake wetland plants ecosystem and influence of lake water level for the structure. Resources and Environment in the Yangtze Basin. 19(6):597–605. in Chinese)
- Hu ZP, Ge G, Liu CL. 2015. Cause analysis and early warning for wetland vegetation degradation in Poyang Lake. Resourc Environ Yangtze Basin. 24(3):381–386. in Chinese)
- Korschgen CE, Green WL, Kenow KP. 1997. Effects of irradiance on growth and winter bud production by Vallisneria americana and consequences to its abundance and distribution. Aquat Bot. 58(1):1–9.
- Low EF, Battoe LE, Coveney MF, Schelske CL, Havens KE, Marzolf ER, Reddy KR. 2001. The restoration of Lake Apopka in relation to alternative stable states: an alternative view to that of Bachmann et al. (1999). Hydrobiologia. 448:11–18.
- Nimptsch J, Pflugmacher S. 2007. Ammonia triggers the promotion of oxidative stress in the aquatic macrophyte Myriophyllum mattogrossense. Chemosphere. 66(4):708–714.
- Thouvenot L, Haury J, Thiébaut G. 2013. Seasonal plasticity of Ludwigia grandiflora under light and water depth gradients: An outdoor mesocosm experiment. Flora. 208(7):430–437.
- Wang SR, Jin XC, Jiao LX, Wu FC. 2009. Response in root morphology and nutrient contents of Myriophyllum spicatum to sediment type. Ecol Eng. 35(8):1264–1270.
- Wang L, Dronova I, Gong P, Yang WB, Li YR, Liu Q. 2012. A new time series vegetation–water index of phenological–hydrological trait across species and functional types for Poyang Lake wetland ecosystem. Remote Sens Environ. 125:49–63.
- Xu WW, Hu WP, Deng JC, Zhu JG, Zhou NN, Liu X. 2016. Impacts of water depth and substrate type on Vallisneria natans at wave-exposed and sheltered sites in a eutrophic large lake. Ecol Eng. 97:344–354.
- Yang ZP, Wang LQ, Tao L, Huang MX. 2015. Nitrogen distribution and ammonia release from the overlying water and sediments of Poyang Lake, China. Environ Earth Sci. doi:10.1007/s12665-015-4081-8.
- Ye C, Yu HC, Kong HN, Song XF, Zou GY, Xu QJ, Liu J. 2009. Community collocation of four submerged macrophytes on two kinds of sediments in Lake Taihu. China Ecol Eng. 35:1656–1663.
- Yuan LY, Li W, Liu GHA, Deng G. 2012. Effects of different shaded conditions and water depths on the growth and reproductive strategy of Vallisneria spinulosa. Pak. J. Bot. 44(3):911–918.
- Zhang D, Chen P, Zhang Q, Li XH. 2017. Copula-based probability of concurrent hydrological drought in the Poyang lake-catchment-river system (China) from 1960 to 2013. J Hydrol. 553:773–784.
- Zhang XL, Zhang Q, Werner AD, Tan ZQ. 2017. Characteristics and causal factors of hysteresis in the hydrodynamics of a large floodplain system: Poyang Lake (China). J Hydrol. 553:574–583.
- Zhao XS, Liu YB. 2017. Phase transition of surface energy exchange in China’s largest freshwater lake. Agricultural and Forest Meteorol. 244–245:98–110.
- Zhao QG, Huang GQ, Qian HY. 2007. Ecological environment and sustainable development of Poyang Lake. Acta Pedolog Sin. 44:318–326. in Chinese)