
Abstract
As human-dominated land-use increases globally, urbanization is a prominent cause of a decline in natural areas, including lentic ecosystems (i.e. lakes, ponds and wetlands). Lentic ecosystems in urban green spaces, such as parks, golf courses or forest preserves, could provide important habitat for organisms within the urban matrix. However, there is a lack of data comparing lentic habitats across these land-use types. We quantified the area of golf courses and forest preserve habitats in the Chicago Metropolitan Region. We measured pesticide concentrations, water chemistry, physicochemical parameters and algal composition in lentic habitats of golf courses and forest preserves in the spring and summer of 2017. Permanent ponds, whether located within a golf course or forest preserve, had similar water quality, chemistry and chlorophyll a and phycocyanin, but differed from ephemeral ponds in forest preserves. Pesticide concentrations within golf course ponds were found in 11 of 25 courses, but only three pesticides (out of 10 measured) were detected. These data suggest golf courses can provide lentic habitats of similar quality to other adjacent land-use types. Additional study is needed to document their role in providing habitat for aquatic biota and sustaining ecosystem services in urban landscapes.
Keywords:
Introduction
Growing human populations have increased the area of urban regions worldwide, and urban expansion is likely to continue (Angel et al. Citation2011). One consequence of urbanization is the disappearance and fragmentation of lentic habitats (McCauley et al. Citation2013; Xu et al. Citation2018) that are critical for many terrestrial and aquatic species. Increasing urbanization and agricultural land use lowers wetlands’ abundance, quality and increases isolation (Gibbs Citation2000; Noble and Hassall Citation2015). The reduced abundance and increased isolation of wetlands result in the inability of many wetland animals to migrate and sustain metapopulations (Semlitsch and Bodie Citation1998; Gibbs Citation2000). However, golf courses could potentially provide habitat and prompt the conservation of such metapopulations in urban landscapes. Understanding the role of lentic systems in providing habitat, sustaining biodiversity and conducting ecosystem services within urban habitats is critical to conserve regional ecosystem functioning.
Wetlands that persist in urban green spaces, such as parks, cemeteries and golf courses, may be crucial to the conservation of lentic environments as they provide important habitats where many aquatic species are threatened by increasing human-driven change (White and Main Citation2005; Yasuda and Koike Citation2006; Hodgkison et al. Citation2007; Morrison et al. Citation2016). Golf courses in the United States comprise over 908,321 hectares of the landscape (Lyman et al. Citation2007). Lentic systems (i.e. standing water bodies) account for an estimated 7% of total acreage on golf courses (over 65,000 hectares in the United States) (Throssell et al. Citation2009). Golf courses can help maintain and enhance local biodiversity, particularly when the adjacent landscape is dominated by anthropogenic land-use (e.g. urbanization and agriculture) (Colding and Folke Citation2009; Petrosillo et al., Citation2019). However, the role of golf courses in the conservation of aquatic ecosystems is rarely addressed.
The landscape on golf courses is composed primarily of turfgrass ecosystems that require management practices potentially impacting habitat quality in ponds. For example, pesticides and nutrients applied to turfgrass can affect water quality, aesthetics and native beneficial organisms via runoff into surface waters (Yang et al. Citation2013; Petrosillo et al. Citation2019). The effect of pesticides, for example, is a function of their mode of action, loading rates and half-life (Christians et al. Citation2017), and pesticides can have nontarget effects on biofilms, invertebrates and amphibians in lentic ecosystems (e.g. growth and development) (Fairchild and Lowe Citation1984; Declerck et al. Citation2011; Griffis-Kyle Citation2007). In addition, turfgrass fertilization with nitrogen and phosphorus can exacerbate eutrophication in downstream aquatic ecosystems. Commitment to the sustainable management of non-turfgrass areas is valuable for maintaining native biodiversity (Tanner and Gange Citation2005; Semlitsch et al. Citation2007). Ecological benefits provided by golf courses are still understudied, and habitat quality and biodiversity of golf course ponds has not been measured or compared to more natural ecosystems. Therefore, understanding lentic ecosystems on golf courses, including the effects of turfgrass management, will provide insights into the capacity of golf course ponds to sustain biodiversity in urban areas.
In addition to golf courses, parks are key sites of conserved land in urbanized environments, and often include lentic habitats that serve as refugia for aquatic taxa and support ecosystem services. The Chicago Metropolitan Region (CMR) is the third largest metropolitan area in the United States (U.S. Census Bureau Citation2019). Governments of the five counties in the region manage thousands of hectares across nearly 200 forest preserves for recreation, storm-water management and wildlife and plant conservation. This green infrastructure network was designed to provide recreational opportunities, trail connections and biological conservation (Ruliffson et al. Citation2002). These protected, managed and restored landscapes are also a primary source of habitat for mammals (Magle et al. Citation2016; Gallo et al. Citation2017; Gallo et al. Citation2018), birds (Brawn and Stotz Citation2001), herpetofauna (Cassel et al. Citation2019) and plants (Brandt et al. Citation2017; Gharehaghaji et al. Citation2019), among other organisms. Variation across the landscapes within forest preserves leads to a variety of lentic habitats, from permanent water bodies to ephemeral, seasonal ponds and wetlands, which were created as a function of the geology across the landscape and management of storm-water runoff (Hackett Citation1966). Forest Preserves harbor more biodiversity compared to golf courses, city parks or cemeteries (Gallo et al. Citation2017). However, the role of forest preserves and golf courses in supporting aquatic taxa in ponds is not well known. Thus, comparing ponds across the types of conserved open space (i.e. golf courses and forest preserves) will allow for important inferences regarding aquatic ecosystem structure and function by land-use type.
The goals of this study were to (1) quantify the area of golf course habitats in the CMR, and (2) compare water chemistry, physicochemical parameters, pesticide concentrations and algal composition in lentic habitats of golf courses and forest preserves. Due to the high percentage of urbanized land-use across the region, we hypothesized that water quality and chemistry of lentic systems within golf courses would be similar to permanent lentic systems within the forest preserve permanent ponds, but different than ephemeral lentic systems in forest preserves. Results from this study will provide insight as to the relative abundance of the golf course ponds, and whether lentic systems within golf courses in urban areas can help support broader objectives for the conservation of aquatic organisms and ecosystem services.
Materials and methods
Landscape analysis
A GIS database was developed for all golf courses and forest preserves across a five-county area in the CMR (McHenry, Lake, DuPage, Winnebago and Cook Counties, Illinois, USA). We used the USGS-GNIS, Esri world imagery map, and referred to aerial photographs obtained from Google Maps (2018) and forest preserve records in each county to quantify the area of each golf course and forest preserve habitat by minimum convex polygons in ArcMap version 10.4.1 (Redlands, CA, USA).
Site description
We quantified water quality, chemistry and algal concentrations across 25 permanent lentic ecosystems on golf courses that contained fish (herein golf course ponds) and compared those to the same parameters in 30 ponds located within adjacent forest preserves: 15 permanent ponds with fish (herein forest preserve ponds) and 15 fishless ephemeral wetlands (herein ephemeral ponds). All ponds chosen were <12,000 m2 in surface area. We further classified sites for sampling purposes: small sites; <5000 m2 and large sites; >5000 m2 surface area. All ponds were sampled once within each of three time periods in 2017: (1) early spring (31 March–15 April), (2) early summer (12–25 June) and (3) late summer (7–21 August). Precipitation data were gathered from weather stations nearest sampling locations and were analyzed by summing total precipitation for the 3 d prior to sampling (Appendix A; Young et al. Citation2018). Sampling was not conducted in ephemeral pond sites for the second and third sampling events if ponds were dry.
Water quality, primary producers, nutrients and pesticides
We collected several measurements of water chemistry on each visit using handheld devices. We used a YSI multiprobe (Model 556; YSI Inc. [Citationdate unknown]) to quantify dissolved oxygen (DO; mg/L), salinity (Sal; ppt), specific conductance (µS/cm), oxidation–reduction potential (ORP; mV), pH and temperature (°C) at three sites per pond on each sampling visit. We also measured chlorophyll a (µg/L) and phycocyanin (blue/green algae; µg/L) within the littoral zone using two FluoroSense Handheld Fluorometers (Turner Designs Co. [Citationdate unknown], linear ranges: 0−199 µg/L). Three (small sites) or five (large sites) chlorophyll a and phycocyanin readings were taken around the perimeter of each pond during each sampling period.
We collected water from each site for later measurement of nutrients and common chemical pollutants from the littoral zone (N = 3 at small sites, N = 5 at large sites). For nutrient analysis, we used 60 mL syringes to collect water samples from shoreline habitats and filtered water using handheld filter holders (MF-Millipore 0.45 µm MCE Membrane filters). Water samples were stored on ice in 20 mL scintillation vials and frozen within 10 h of collection until further analysis. In the lab, dissolved inorganic nutrients were measured using an Autoanalyzer III (Seal, Analytical, Inc. Mequon, WI), with the phenol hypochlorite method for ammonium (NH4+; µg N/L; Solorzano Citation1969), the sulfanilamide method for nitrate and nitrite (NO3− + NO2−; µg N/L; herein NOx−; APHA Citation1998), and the antimonyl tartrate method for soluble reactive phosphorus (µg P/L; Murphy and Riley Citation1962). Finally, we collected water from each of the 25 golf course ponds and seven accessible course inflows (e.g. courses with accessible wells or lotic systems filling course ponds) during the April and August sampling periods to measure 10 common pesticides. Water samples were taken by mixing approximately 10 L of water from the littoral zone and filtering 1 L of water through a 0.45 µm filter. Samples were stored on ice and immediately shipped to the University of Georgia: Agricultural and Environmental Services Laboratories for analysis within 48 h of collection. We examined concentrations of 10 analytes (minimum detection limit [ppb]): azoxystrobin (0.50), bifenthrin (0.20), α-chlordane (0.20), γ-chlordane (0.20), chlorpyrifos (0.20), cypermethrin (1.00), oxadiazon (0.50), cis-permethrin (1.30), trans-permethrin (1.30) and fenvalerate (0.30)
Statistical analysis
We used one-way analysis of variance (ANOVA) followed by Tukey Multiple Comparison tests to compare water quality, water chemistry and algal concentrations between golf course ponds, forest preserve ponds and ephemeral ponds within each sampling period. Statistical analyses were conducted using Statistica version 13.0 (TIBCO Software, Inc. Citation2017).
Results
Quantification of golf course and forest preserve area
We identified 340 golf courses and 356 state or county forest preserves in the five county CMR. In total, golf courses represented 21,185 ha (mean size of courses = 72 ha, SD = 52 ha) and forest preserves represented 84,923 ha (mean size of forest preserves = 239 ha, SD = 512 ha). The five-county area of the CMR (Cook, Lake, DuPage, McHenry and Kane counties) represents a total of 1,130,787 ha, which equates to golf courses and forest preserves representing 1.9% and 7.5% of total land area, respectively.
Water quality
Measurements of water quality showed a distinction between ephemeral ponds relative to the golf courses and forest preserve ponds, although patterns varied by date and metric (Appendix B; ). Specific conductance and pH were significantly higher in golf course ponds and forest preserve ponds compared to ephemeral ponds during every month sampled (Specific conductance: April p = 0.025, June p = 0.026, August p = 0.047; pH: June p ≤ 0.001, August p ≤ 0.001) (; ). For example, pH and specific conductance in June were 21% and 47% lower, respectively, in ephemeral ponds compared to golf course ponds. Salinity was significantly lower in ephemeral ponds compared to golf course ponds for each sampled month (April p = 0.043, June p = 0.030, August p = 0.020), however, forest preserve ponds were not significantly different between golf course and ephemeral ponds (; ). Mean DO was significantly higher in golf course and forest preserve ponds than ephemeral ponds in June and August (June p = 0.001, August p ≤ 0.001) (; ), but not in April. Specifically, DO in June was 3.2 and 2.8 times higher in the golf courses and forest preserve ponds, respectively, compared to ephemeral ponds. Temperature in June was significantly higher in golf course ponds compared to forest preserve and ephemeral ponds (p = 0.001) (; ). There were no significant differences in temperature across pond types in April or August. There were no significant differences in ORP values across pond types; however, ORP values in all pond types were highest in August.
Table 1. One-way ANOVA results examining differences in water quality, algae and nutrient measures across golf course, forest preserve permanent and ephemeral lentic ecosystems in June, July and August 2017.
Chlorophyll a and phycocyanin
Relative concentrations of chlorophyll a and phycocyanin were high across all sampling dates. On average, all ponds in this study were determined to be eutrophic across all months (7–40 µg/L; Forsberg and So Citation1980). Hypereutrophic values (≥40 µg/L) were more frequent in golf course ponds compared to forest preserve and ephemeral ponds (; ). Among pond types, chlorophyll a concentration was significantly higher in ephemeral ponds compared to golf course and forest preserve ponds in August (p = 0.049) (; ), where chlorophyll a concentrations were 84% and 96% higher in ephemeral ponds than in golf course and forest preserve ponds, respectively.
Water chemistry and pesticides
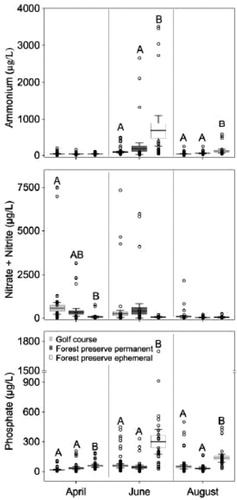
In general, water chemistry values were similar in forest preserve and golf course ponds but both were significantly different from ephemeral ponds for most parameters (, ). Ephemeral ponds had significantly higher ammonium concentrations compared to forest preserve and golf course ponds in June and August (June p ≤ 0.001, August p ≤ 0.001) (, ). Specifically, June mean ammonium values were 3.7 (forest preserve ponds) and 7.3 (golf course ponds) times lower than in ephemeral ponds. There was no significant difference in ammonium levels between pond types in April. In contrast, NOx− concentrations were significantly lower in ephemeral ponds relative to golf course ponds in April (p = 0.046) (, ), whereas in June and August no significant differences were found across the pond types. Forest preserve and golf course ponds were 4.3 and 8.1 times higher, respectively, in April than in ephemeral ponds. Phosphate concentrations were significantly lower in the forest preserve and golf course ponds compared to ephemeral ponds across all months sampled (April p ≤ 0.001, June p ≤ 0.001; August p ≤ 0.001) (, ). The greatest difference between pond types occurred in June. Phosphate in ephemeral ponds was 7 and 5.2 times higher compared to forest preserve and golf course ponds, respectively (, ).
Of the 10 pesticides we measured, only 3 were detected in August, and none were detected in April. Azoxystrobin, α-chlordane and oxadiazon were the only pesticides with concentrations above the detectable limit in any of the 25 golf courses in August (). Three of the seven-course inflows we examined had detectable pesticides that were likely contributing to the concentrations found in golf course ponds (). Ponds with α-chlordane or oxadiazon also had azoxystrobin. However, no ponds had both α-chlordane and oxadiazon. One of the pond inflows had both oxadiazon and azoxystrobin.
Table 2. Concentrations of pesticides (µg/L) within golf course ponds (25) and course inflows (7) in August 2017.
Discussion
This study aimed to compare lentic systems between urban parks, i.e. forest preserves, and golf courses by examining a suite of limnological variables. Assessing golf course ponds is crucial, as golf courses comprise large areas of urban landscapes, and could be a valuable, but overlooked source for ecosystem services provided by natural ponds (Lyman et al. Citation2007). In the CMR, golf courses represent nearly 2% of land use in this highly urbanized region, thus providing important areas of greenspace. Our results suggest physicochemical characteristics, algae and water chemistry parameters were similar between permanent golf course and forest preserve ponds, although significantly different from ephemeral ponds in forest preserves. Overall, golf course ponds provide similar quality habitat for aquatic organisms and ecosystem processes compared to natural permanent ponds in urban landscapes.
Water quality
Differences in water quality between permanent ponds (i.e. in golf courses and forest preserves) and ephemeral ponds were driven by hydrology, seasonal drying and oxygen concentrations. Ephemeral ponds were in the process of drying up over the course of this study, unlike permanent ponds that had a continuous source of water. Specific conductance, DO, pH and salinity were significantly lower in ephemeral ponds compared to permanent ponds. Pond drying can elevate salinity, decrease DO and elevate ammonia and phosphate in the water column as they release from sediments (Baldwin et al. Citation2000; Kassila et al. Citation2001). Considering all our ponds were found within a matrix of urban to suburban land use, elevated salinity and specific conductance in permanent ponds could also be a result of continuous inputs of heavy metals, ions and nutrients in runoff from the surrounding landscape (Dow and Zampella Citation2000; Paul and Meyer Citation2001).
Our values for each water quality parameter were similar to reports from ponds within urban golf courses across the United States (Ryals et al. Citation1998; Davis and Lydy Citation2002; Lewitus et al. Citation2003; Devitt et al. Citation2005). This suggests golf course lentic habitats are providing similar habitat in urban areas that represent an important contribution to urban landscapes, and that golf course ecosystem management facilitates healthy lentic ecosystems. These results contradict some studies (Seminara et al. Citation2008), but are consistent with others (Fairchild et al. Citation2003), especially studies in the Midwestern United States (Lipsey Citation1980; Little and Church Citation2018). For example, values of pH, chlorophyll a, and DO are comparable to other studies (Lipsey Citation1980; Little and Church Citation2018). Although water quality measures across the CMR ponds differ considerably, these measures are consistently similar in golf course and forest preserve ponds.
Water chemistry and pesticides
Our results show a clear pattern of higher nutrient concentrations in ephemeral ponds compared to permanent ponds, whether located within a golf course or forest preserve. Few studies have compared water chemistry in temporary and permanent ponds; however, our results are consistent with published values. Drenner et al. (Citation2009) found no significant difference in phosphorus levels in temporary versus permanent ponds, however, total nitrogen concentrations were significantly lower in temporary ponds, likely as a result of redox. Higher concentrations of phosphorus and ammonium in our ephemeral ponds were likely influenced by oxygen levels. DO was significantly higher in permanent ponds (), likely as a function of increased water movement and continuous inflow. Oxygenated conditions sustain abiotic sorption of P with sediment particles (e.g. iron oxide), thereby decreasing phosphorus concentrations in the water column (Bertram Citation1993; Jiang et al. Citation2008). Similarly, ammonium concentrations are sustained under low oxygen conditions, which inhibit assimilatory and dissimilatory (i.e. nitrification) transformations (Howard‐Williams Citation1985). Land-use could also be a driving factor in nutrient concentrations. In the CMR, nitrogen and phosphorus concentrations in surface waters were associated with agricultural, barren land and developed space (Wilson and Weng Citation2010), and many ephemeral wetlands in our study, although forested, were surrounded by land-use that could facilitate nutrient runoff. These data suggest golf course and forest preserve ponds differ from ephemeral ponds with respect to water quality and chemistry parameters, but are similar to each other, which allows for quality habitat for aquatic biota in permanent ponds in both land-use types.
Pesticide use on golf courses is distinct from other types of urban land use and may represent a stressor on habitat quality in golf course ponds. Concentration and type of pesticides used vary across golf courses but over the past few decades pesticide accumulation from golf course sources has declined (Cohen et al. Citation1999; Baris et al. Citation2010Citation). In this study, only three of the 10 pesticides tested were found above the detectable limit from 25 golf courses in August and none were detected in April. Other studies examining pesticides on golf courses report inconsistent results. Miles et al. (Citation1992) examined surface water and groundwater associated with golf courses and detected 10 out of 17 pesticides analyzed (60%); whereas Ryals et al. (Citation1998) determined that the impact of golf course pesticide use was minimal to the golf course ponds as most values above the detectible limit were below the EPA’s health advisory level. In addition, studies have found concentrations of pesticides, when detected, are similar to those in our study (Ryals et al. Citation1998; Mackey et al. Citation2014). These results suggest, in this study system, commonly used pesticides were not detrimental to water quality, chlorophyll a and phycocyanin in golf course ponds; however, we did not measure biotic impacts in this study and note that indirect or sub-lethal impacts on biota could also occur.
Chlorophyll a and phycocyanin
Differences in land-use types associated with lentic systems can impact phytoplankton (chlorophyll a and phycocyanin) biomass, which typically show that rural or forested systems have lower concentrations compared to agriculture or urban areas (Jones and Knowlton Citation1993; Knoll et al. Citation2003; Katsiapi et al. Citation2012). In our study, there was little difference in chlorophyll among pond types, and the range of concentrations was similar to those reported from northern Illinois (Lipsey Citation1980). Other studies have found higher chlorophyll a concentrations in urban ponds when compared to forested ponds (Kuczyńska‐Kippen and Joniak Citation2010; Marion et al. Citation2017). However, all of our study sites lie within urbanized watersheds, even when immediately surrounded by a forest preserve or golf course. Thus, we did not find differences in chlorophyll a and phycocyanin among our sites because they are all within a matrix of heavily developed landscape. The lack of significant differences across habitat types suggests golf course ponds provide viable habitat for phytoplankton, and possibly other biota, and are of similar concentration to those of ponds in forest preserves. Previous studies have reported significant relationships between chlorophyll a concentrations and phosphorus (Cunha Pereira et al. Citation2010), and nitrogen inputs can increase chlorophyll a concentrations when phosphorus is high (McCauley et al. Citation1989). Similar patterns between elevated nitrogen and phosphorus levels and chlorophyll a may explain the lack of differences in algal concentration between our study pond types ( and ).
Figure 1. Boxplots representing specific conductance (µS/cm), dissolved oxygen (DO; mg/L), temperature (°C), water pH, oxidation–reduction potential (ORP) and salinity (Sal; ppt), within golf course (n = 25 forest preserve permanent (n = 15), and forest preserve ephemeral ponds (n = 15) in April, June, and August 2017. Different upper-case letters indicate statistical significance (p ≤ 0.05) within months using Tukey HSD test. Line represents mean, boxes represent standard error, and whiskers represent 95% confidence intervals. Open circles are data values.
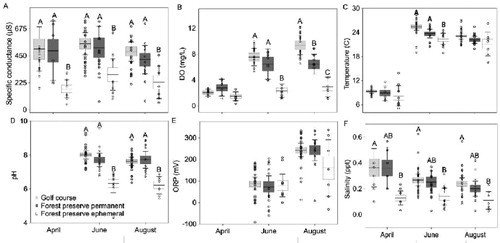
Figure 2. Boxplots representing chlorophyll a (µg/L) and phycocyanin (µg/L) concentrations within golf course (25), forest preserve permanent (15), and forest preserve ephemeral (15) ponds in April, June, and August 2017. Different upper-case letters indicate statistical significance (p ≤ 0.05) within months using Tukey HSD test. Line represents mean, boxes represent standard error, and whiskers represent 95% confidence intervals. Open circles are data values.
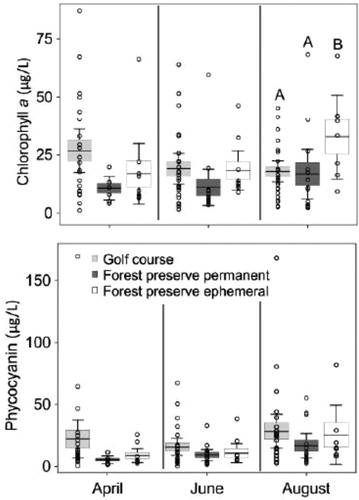
Figure 3. Boxplots representing ammonium (µg/L), nitrate and nitrite (µg/L), and phosphate (µg/L) concentrations within golf course, forest preserve permanent and forest preserve ephemeral ponds in April, June and August 2017. Different upper-case letters indicate statistical significance (p ≤ 0.05) within months using Tukey HSD test. Line represents mean, boxes represent standard error and whiskers represent 95% confidence intervals. Open circles are data values.
Golf courses showed similar habitat quality to permanent ponds located in urban parks in our study as well as other lentic habitats in the Midwest (Lipsey Citation1980; Little and Church Citation2018). However, habitat quality parameters of permanent ponds in our study, whether golf course or forest preserve, were different from ephemeral ponds, despite the hypereutrophic conditions in both ephemeral ponds and golf course ponds. Differences in abiotic factors, such as water quality, nutrient concentrations and chlorophyll a, between ephemeral ponds and permanent forest preserve/golf course ponds were likely driven by hydrology and oxygen dynamics, with important implications for biological communities and ecosystem processes. Whilst golf course ponds showed similar habitat quality to permanent ponds in forest preserves, golf course management can still improve the habitat quality of their lentic systems. Potential ways to improve habitat, minimize water quality issues and reduce pesticide input into these systems include adding native and unmaintained vegetative riparian buffers, reduce fertilizer application rate and volume, and make structural improvements of onsite drainage and wetland filtration systems (Davis and Lydy Citation2002; Winter et al. Citation2002; Winter and Dillon Citation2005). Future research on biotic factors across pond types is necessary to assess whether aquatic communities in golf course ponds are similar to those in natural permanent ponds and will provide insight as to whether golf courses can be considered a source for biodiversity of aquatic species in urban areas. We suggest future studies will benefit from including permanent and ephemeral ponds across a wider gradient of land use types, including golf courses in more rural areas adjacent to natural landscapes. Additional research that spans regions and landscape types will be valuable to evaluate the role of golf courses habitats for aquatic biota.
Acknowledgments
We thank S. Crites and J. Mattes for field assistance. The work was conducted under Illinois State permit NH17.6101, Lake County Forest Preserves Permit no. 2017-73 and Forest Preserve District of DuPage County Permit no. 17-06.
Disclosure statement
No potential conflict of interest was by reported the author(s).
Additional information
Funding
Notes on contributors
Jennifer N. Piacente
Jennifer N. Piacente, M.S., is a graduate of Loyola University Chicago and studied invasive macrophytes and aquatic macroinvertebrates. She is currently employed by the Florida Department of Environmental Protection.
Joseph R. Milanovich
Dr. Joseph R. Milanovich is an Associate Professor of Biology at Loyola University Chicago. He studies the relationship between humans and wildlife conservation.
Martin B. Berg
Dr. Martin B. Berg is a Professor of Biology at Loyola University Chicago. He studies aquatic insects and their role in energy flow of lentic and lotic systems.
Timothy J. Hoellein
Dr. Timothy J. Hoellein is an Associate Professor of Biology at Loyola University Chicago. His research addresses the movement and biological transformations of pollutants in aquatic ecosystems.
Andrés G. Muñoz
Andrés G. Muñoz is a PhD Candidate at the University of Denver. He studies synthetic organic chemistry and develops photochemical methodology for application in medicinal chemistry.
Armand A. Cann
Armand A. Cann, M.S., is a Biology Lab Instructor at Loyola University Chicago. He studies the wildlife conservation.
Isabella S. Lentini
Isabella S. Lentini is a graduate of Loyola University Chicago with a B.S. in Ecology. She is currently pursuing a Masters of Nursing at DePaul University.
References
- Angel S, Parent J, Civco DL, Blei A, Potere D. 2011. The dimensions of global urban expansion: estimates and projections for all countries, 2000–2050. Prog Plann. 75(2):53–107.
- APHA. 1998. Standard methods for the examination of water and wastewater. 20th ed. Baltimore (MD): United Book Press Inc.
- Baldwin DS, Mitchell AM, Rees GN. 2000. The effects of in situ drying on sediment-phosphate interactions in sediments from an old wetland. Hydrobiologia. 431(1):3–12.
- Baris RD, Cohen SZ, Barnes NL, Lam J, Ma Q. 2010. Quantitative analysis of over 20 years of golf course monitoring studies. Environ Toxicol Chem. 29(6):1224–1236.
- Bertram PE. 1993. Total phosphorus and dissolved oxygen trends in the central basin of Lake Erie, 1970–1991. J Great Lakes Res. 19(2):224–236.
- Brandt LA, Lewis AD, Scott L, Darling L, Fahey RT, Iverson L, Nowak DJ, Bodine AR, Bell A, Still S. 2017. Chicago Wilderness region urban forest vulnerability assessment and synthesis: a report from the Urban Forestry Climate Change Response Framework Chicago Wilderness pilot project. Gen Tech Rep. 142:168.
- Brawn JD, Stotz DF. 2001. The importance of the Chicago region and the “Chicago Wilderness” initiative for avian conservation. Avian ecology and conservation in an urbanizing world. Berlin, Germany: Springer; p. 509–522.
- Cassel KW, Vanek JP, Glowacki GA, Preuss TS, Nielsen CK. 2019. Multiscale habitat factors influence the occupancy and turnover of the suburban herpetofauna of Chicago, Illinois, USA. Herpetol Conserv Biol. 14(2):438–454.
- Christians N, Patton AJ, Law QD. 2017. Fundamentals of turfgrass management. Hoboken (NJ): Wiley & Sons, Inc.
- Cohen S, Svrjcek A, Durborow T, Barnes NL. 1999. Water quality impacts by golf courses. J Environ Qual. 28(3):798–809.
- Colding J, Folke C. 2009. The role of golf courses in biodiversity conservation and ecosystem management. Ecosystems. 12(2):191–206.
- Cunha Pereira H, Allott N, Coxon C. 2010. Are seasonal lakes as productive as permanent lakes? A case study from Ireland. Can J Fish Aquat Sci. 67(8):1291–1302.
- Davis NM, Lydy MJ. 2002. Evaluating best management practices at an urban golf course. Environ Toxicol Chem. 21(5):1076–1084.
- Declerck SAJ, Bakker ES, van Lith B, Kersbergen A, van Donk E. 2011. Effects of nutrient additions and macrophyte composition on invertebrate community assembly and diversity in experimental ponds. Basic Appl Ecol. 12(5):466–475.
- Devitt DA, Morris RL, Baghzouz M, Lockett M, Fenstermaker LK. 2005. Water quality changes in golf course irrigation ponds transitioning to reuse water. Hort Sci. 40(7):2151–2156.
- Dow CL, Zampella RA. 2000. Specific conductance and pH as indicators of watershed disturbance in streams of the New Jersey Pinelands, USA. Environ Manage. 26(4):437–445.
- Drenner SM, Dodson SI, Drenner RW, Pinder JE. III 2009. Crustacean zooplankton community structure in temporary and permanent grassland ponds. Hydrobiologia. 632(1):225–233.
- Fairchild GW, Cruz J, Faulds AM, Short AEZ, Matta JF. 2003. Microhabitat and landscape influences on aquatic beetle assemblages in a cluster of temporary and permanent ponds. J North Am Benthol Soc. 22(2):224–240.
- Fairchild GW, Lowe RL. 1984. Artificial substrates which release nutrients: effects on periphyton and invertebrate succession. Hydrobiologia. 114(1):29–37.
- Forsberg C, So R. 1980. Eutrophication parameters and trophic state indices in 30 Swedish waste-receiving lakes. Arch. Hydrobiol. 89(1–2):189–207.
- Gallo T, Fidino M, Lehrer EW, Magle SB. 2017. Mammal diversity and metacommunity dynamics in urban green spaces: implications for urban wildlife conservation. Ecol Appl. 27(8):2330–2341.
- Gallo T, Lehrer EW, Fidino M, Kilgour RJ, Wolff PJ, Magle SB. 2018. Need for multiscale planning for conservation of urban bats. Conserv Biol. 32(3):638–647.
- Gharehaghaji M, Kobal S, Reklau R, Minor ES. 2019. Management slows down invasion by non-native plants but does not prevent community change over 35 years in urban forests of the Midwestern USA. For Ecol Manage. 448:424–431.
- Gibbs JP. 2000. Wetland loss and biodiversity conservation. Conserv Biol. 14(1):314–317.
- Griffis-Kyle KL. 2007. Sublethal effects of nitrite on eastern tiger salamander (Ambystoma tigrinum tigrinum) and wood frog (Rana sylvatica) embryos and larvae: implications for field populations. Aquat Ecol. 41(1):119–127.
- Hackett JE. 1966. An application of geologic information to land use in the Chicago metropolitan region. Environ Geol. 8:1–22.
- Hodgkison S, Hero J-M, Warnken J. 2007. The efficacy of small-scale conservation efforts, as assessed on Australian golf courses. Biol Conserv. 135(4):576–586.
- Howard‐Williams C. 1985. Cycling and retention of nitrogen and phosphorus in wetlands: a theoretical and applied perspective. Freshw Biol. 15(4):391–431.
- Jiang X, Jin X, Yao Y, Li L, Wu F. 2008. Effects of biological activity, light, temperature and oxygen on phosphorus release processes at the sediment and water interface of Taihu Lake, China. Water Res. 42(8–9):2251–2259.
- Jones JR, Knowlton MF. 1993. Limnology of Missouri reservoirs: an analysis of regional patterns. Lake Reserv Manag. 8(1):17–30.
- Kassila J, Hasnaoui M, Droussi M, Loudiki M, Yahyaoui A. 2001. Relation between phosphate and organic matter in fish-pond sediments of the Deroua fish farm (Béni-Mellal, Morocco): implications for pond management. Hydrobiologia. 450(1–3):57–70.
- Katsiapi M, Mazaris AD, Charalampous E, Moustaka-Gouni M. 2012. Watershed land use types as drivers of freshwater phytoplankton structure. Phytoplankt responses to human impacts at different scales. Berlin, Germany: Springer; p. 121–131.
- Knoll LB, Vanni MJ, Renwick WH. 2003. Phytoplankton primary production and photosynthetic parameters in reservoirs along a gradient of watershed land use. Limnol Oceanogr. 48(2):608–617.
- Kuczyńska‐Kippen N, Joniak T. 2010. The impact of water chemistry on zooplankton occurrence in two types (field versus forest) of small water bodies. Int Rev Hydrobiol. 95(2):130–141.
- Lewitus AJ, Schmidt LB, Mason LJ, Kempton JW, Wilde SB, Wolny JL, Williams BJ, Hayes KC, Hymel SN, Keppler CJ. 2003. Harmful algal blooms in South Carolina residential and golf course ponds. Popul Environ. 24(5):387–413.
- Lipsey LL. 1980. Phytoplankton of selected borrow pit ponds in northern Illinois. Ohio Acad Sci. 5:108–113.
- Little AM, Church JO. 2018. Ephemeral pond vegetation within the glaciated Upper Midwest: a comparison with permanent wetlands. Freshw Sci. 37(1):31–41.
- Lyman GT, Throssell CS, Johnson ME, Stacey GA, Brown CD. 2007. Golf course profile describes turfgrass, landscape, and environmental stewardship features. Appl Turfgrass Sci. 4(1):0.
- Mackey MJ, Connette GM, Peterman WE, Semlitsch RD. 2014. Do golf courses reduce the ecological value of headwater streams for salamanders in the southern Appalachian Mountains? Landsc Urban Plan. 125:17–27.
- Magle SB, Lehrer EW, Fidino M. 2016. Urban mesopredator distribution: examining the relative effects of landscape and socioeconomic factors. Anim Conserv. 19(2):163–175.
- Marion JW, Zhang F, Cutting D, Lee J. 2017. Associations between county-level land cover classes and cyanobacteria blooms in the United States. Ecol Eng. 108:556–563.
- McCauley E, Downing JA, Watson S. 1989. Sigmoid relationships between nutrients and chlorophyll among lakes. Can J Fish Aquat Sci. 46(7):1171–1175.
- McCauley LA, Jenkins DG, Quintana-Ascencio PF. 2013. Isolated wetland loss and degradation over two decades in an increasing urbanized landscape. Wetlands. 33(1):117–127.
- Miles CJ, Leong G, Dollar S. 1992. Pesticides in marine sediments associated with golf course runoff. Bull Environ Contam Toxicol. 49(2):179–185.
- Morrison JL, Gottlieb IGW, Pias KE. 2016. Spatial distribution and the value of green spaces for urban red-tailed hawks. Urban Ecosyst. 19(3):1373–1388.
- Murphy J, Riley JP. 1962. A modified single solution method for the determination of phosphate in natural waters. Anal Chim Acta. 27:31–36.
- Noble A, Hassall C. 2015. Poor ecological quality of urban ponds in northern England: causes and consequences. Urban Ecosyst. 18(2):649–662.
- Paul MJ, Meyer JL. 2001. Streams in the urban landscape. Annu Rev Ecol Syst. 32(1):333–365.
- Petrosillo I, Valente D, Pasimeni MR, Aretano R, Semeraro T, Zurlini G. 2019. Can a golf course support biodiversity and ecosystem services? The landscape context matter. Landscape Ecol. 34(10):2213–2228.
- Ruliffson JA, Gobster PH, Haight RG, Homans FR. 2002. Niches in the urban forest: organizations and their role in acquiring metropolitan open space. J For. 100(6):16–24.
- Ryals SC, Genter MB, Leidy RB. 1998. Assessment of surface water quality on three eastern North Carolina golf courses. Environ Toxicol Chem. 17(10):1934–1942.
- Seminara M, Vagaggini D, Margaritora FG. 2008. Differential responses of zooplankton assemblages to environmental variation in temporary and permanent ponds. Aquat Ecol. 42(1):129–140.
- Semlitsch RD, Bodie JR. 1998. Are small, isolated wetlands expendable? Conserv Biol. 12(5):1129–1133.
- Semlitsch RD, Boone MD, Bodie JR. 2007. Using golf courses to bolster amphibian communities. USGA Green Sect Rec. 45:7–11.
- Solorzano L. 1969. Determination of ammonia in natural waters by the phenolhypochlorite method. Limnol Oceanogr. 14(5):799–801.
- Tanner RA, Gange AC. 2005. Effects of golf courses on local biodiversity. Landsc Urban Plan. 71(2–4):137–146.
- Throssell CS, Lyman GT, Johnson ME, Foundation G, Highway SUS. 2009. Golf course environmental profile measures water use, source, cost, quality, and management and conservation strategies applied turfgrass science applied turfgrass science. Plant Manag Netw. 6(1):0–0.
- TIBCO Software, Inc. 2017. Global leader in integration and analytics software. TIBCO Software Inc. www.tibco.com.
- Turner Designs Co. [date unknown]. Fluorometers for water quality. United States: Turner Designs. www.turnerdesigns.com.
- U.S. Census Bureau. 2019. U.S. Census Bureau QuickFacts: United States. Census Bureau QuickFacts. [accessed 2020 May 21]. https://www.census.gov/quickfacts.
- White CL, Main MB. 2005. Waterbird use of created wetlands in golf‐course landscapes. Wildl Soc Bull. 33(2):411–421.2.0.CO;2]
- Wilson C, Weng Q. 2010. Assessing surface water quality and its relation with urban land cover changes in the Lake Calumet Area, Greater Chicago. Environ Manage. 45(5):1096–1111.
- Winter JG, Somers KM, Dillon PJ, Paterson C, Reid RA. 2002. Impacts of golf courses on macroinvertebrate community structure in Precambrian Shield streams. J Environ Qual. 31(6):2015–2025.
- Winter JG, Dillon PJ. 2005. Effects of golf course construction and operation on water chemistry of headwater streams on the Precambrian Shield. Environ Pollut. 133(2):243–253.
- Xu X, Xie Y, Qi K, Luo Z, Wang X. 2018. Detecting the response of bird communities and biodiversity to habitat loss and fragmentation due to urbanization. Sci Total Environ. 624:1561–1576.
- Yang Y, He Z, Wang Y, Fan J, Liang Z, Stoffella PJ. 2013. Dissolved organic matter in relation to nutrients (N and P) and heavy metals in surface runoff water as affected by temporal variation and land uses–A case study from Indian River Area, south Florida, USA. Agric Water Manag. 118:38–49.
- Yasuda M, Koike F. 2006. Do golf courses provide a refuge for flora and fauna in Japanese urban landscapes? Landsc Urban Plan. 75(1–2):58–68.
- YSI Inc. [date unknown]. Water quality sampling and monitoring meters and instruments for dissolved oxygen, PH, turbidity. YSI. www.ysi.com/.
- Young AH, Knapp KR, Inamdar A, Hankins W, Rossow WB. 2018. The international satellite cloud climatology project H-Series climate data record product. Earth Syst Sci Data. 10(1):583–593.
Appendix A
Total precipitation (mm) data for golf course and forest preserve sites 3 d prior to the sampling event for (1) early spring (31 March–15 April), (2) early summer (12–25 June) and 3) late summer (7–21 August) sampling periods. Data were derived from five separate weather stations using the National Centers for Environmental Information (Young et al. Citation2018) located within 20 miles of each golf course, respectively.
Appendix B
Mean (SE) specific conductance (SC, µS), dissolved oxygen (DO; mg/L), temperature (T, °C), water pH, oxidation–reduction potential (ORP) and salinity (S; ppt), within golf course (GC, n = 25 forest preserve permanent (FP-P, n = 15), and forest preserve ephemeral ponds (FP-E, n = 15) in April, June and August 2017.