ABSTRACT
Morphological studies of fossil and extant shark teeth have typically been qualitative in nature, with resulting taxonomic problems due to the complicated forms of heterodonty exhibited by many sharks. This is apparent in the designation of fossil species assigned to Isurus (Lamniformes), where the status of the putative Neogene fossil species Isurus xiphodon and I. hastalis is solely based upon supposed differences in tooth morphology. Here we apply the geometric morphometric techniques of Procrustes superimposition, resampling-based Goodall's F-test, and canonical variates analysis to extant Isurus teeth, with the goal of assessing whether these quantitative analytical techniques provide a more objective basis for taxonomic decisions than do traditional qualitative morphological descriptions. These quantitative techniques are then applied to I. hastalis and I. xiphodon to examine whether I. xiphodon should be considered a junior synonym of I. hastalis or a separate species. Results show that geometric morphometric techniques can differentiate between the two extant species as well as the two extinct species, supporting I. xiphodon as a valid taxon. We suggest that this type of analysis is applicable for examining tooth-based species delimitations of sharks with both extant and extinct members, and has the potential to be applied to other fossil shark species as well.
INTRODUCTION
Fossil chondrichthyan species are typically identified on the basis of qualitative descriptions of tooth morphology. The reliance on teeth results from the fact that teeth are often the only parts of the skeleton preserved in the fossil record (CitationCappetta, 1987). However, such studies have inherent problems. Many extant elasmobranchs exhibit various and sometimes complicated forms of heterodonty, including monognathic, dignathic, sexual, and ontogenetic heterodonty (CitationPeyer, 1968; CitationCompagno, 1988; CitationGottfried and Francis, 1996; CitationKajiura and Tricas, 1996). Variations in tooth morphology may also be geographic (CitationLucifora et al., 2003) or pathologic (CitationBecker et al., 2000). The presence of heterodonty or other variations in tooth morphology may not be immediately apparent in fossil taxa, as most shark teeth are not found as associated sets (CitationShimada, 2005).
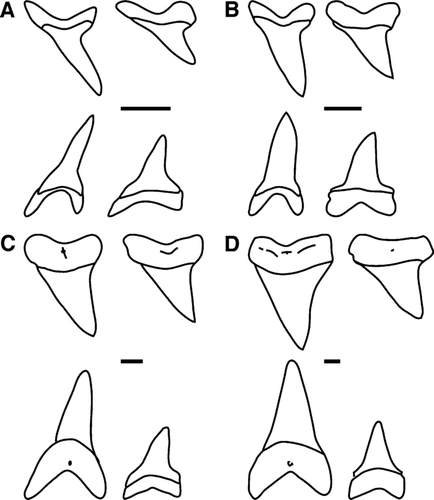
The problem of heterodonty in tooth morphology has cast doubt on qualitative studies of isolated fossil shark teeth (CitationCompagno, 1988; CitationApplegate and Espinosa-Arrubarrena, 1996; CitationHubbell, 1996; CitationGottfried and Fordyce, 2001; CitationPurdy et al., 2001; CitationShimada, 2005). This is apparent in the designation of fossil species assigned to Isurus (Lamniformes). There are two extant species of mako sharks, the shortfin mako Isurus oxyrinchus and the longfin mako I. paucus. Both are fast swimming pelagic predators. The species display sexual dimorphism, with females generally larger than males (CitationPratt and Casey, 1983; CitationCompagno, 1988). In addition to the relative lengths of their pectoral fins, tooth morphology has been used as a diagnostic feature to distinguish the species. Both species have unserrated, unicuspid teeth, but the teeth of I. oxyrinchus have relatively narrow cusps with reversed and labially recurved tips, whereas the teeth of I. paucus have broader cusps and are not recurved (CitationCompagno, 2001) (). Both extant species exhibit monognathic and dignathic heterodonty. Teeth can be divided into anterior, intermediate, and lateral sets on both the upper and lower jaws (CitationShimada, 2002). The status of the putative Neogene fossil species I. xiphodon and I. hastalis is solely based upon supposed differences in tooth morphology. Agassiz (1843) originally established these species based on the shape of the lingual crown foot. Eventually, the two species were synonymized, along with several of Agassiz's other species, into I. hastalis, based partially on a lack of a type specimen for I. xiphodon (CitationLawley, 1878; CitationLeriche, 1926). CitationPurdy et al. (2001) reestablished I. xiphodon as a separate species qualitatively on the basis of the broadness of the tooth and root lobe and mesial cutting edge shape (). Despite this recent change, some still consider I. xiphodon to be a nomen dubium (CitationWard and Bonavia, 2001).
FIGURE 1 Representative anterior and lateral teeth of Isurus species included in this study. For each figure portion, teeth in the top row are from the upper jaw and the bottom row from the lower jaw. Scale bar = 1.0 cm. A, I. oxyrinchus, labial view (Florida Museum of Natural History 030102013.28); B, I. paucus, labial view (GHC LONG1786); C, Isurus hastalis, lingual view (USNM 453155, 474480, 474986, 474988); D, I. xiphodon, lingual view (USNM 421913, 278765, 421916, 482217).
Geometric morphometrics is a technique that allows for quantification of shape and visualization of complex morphology via the designation of landmarks. Taking the coordinates of these anatomical landmarks allows shapes to be objectively compared through various statistical analyses. These techniques has been applied to a variety of studies incorporating morphology (CitationRohlf et al., 1996; CitationZelditch et al., 2003; CitationBogdanowicz et al., 2005; CitationOettle et al., 2005; CitationPretorius, 2005; CitationCandioti, 2006), including studies of elasmobranch dentitions. Discriminant function analysis and quadratic discriminant analysis have been applied to linear measurements between landmarks to characterize recent Carcharhinus teeth (CitationNaylor and Marcus, 1994). Linear distances between landmarks have also been used to elucidate evolutionary relationships between fossil and extant species of Hexanchus (CitationAdnet, 2006a). Geometric morphometric techniques have been applied to modern and fossil Rhizoprionodon teeth and were able to distinguish between Rhizoprionodon and closely related genera, but could not distinguish between species of Rhizoprionodon (CitationGarry, 2004). Most recently, geometric morphometrics was used to explore the evolutionary origins of modern white sharks Carcharodon carcharias (CitationNyberg et al., 2006).
Here we apply the geometric morphometric techniques of Procrustes superimposition, resampling-based Goodall's F-test, and canonical variates analysis to extant Isurus teeth, with the goal of assessing whether these quantitative analytical techniques provide a more objective basis for taxonomic decisions than do traditional qualitative morphological descriptions. These techniques are then applied to two fossil species of Isurus, I. hastalis and I. xiphodon, to quantitatively examine whether I. xiphodon should be considered a junior synonym of I. hastalis or a valid separate species.
MATERIALS AND METHODS
Analysis of Extant Species
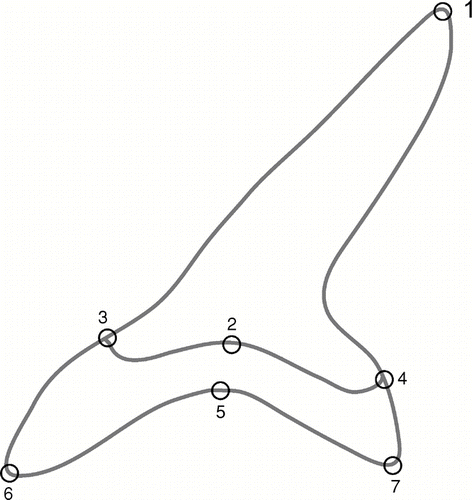
Tooth sets and jaws of adult I. oxyrinchus (N = 10) and I. paucus (N = 11) were obtained from the following collections: National Museum of Natural History (NMNH), Washington, D.C.; Field Museum of Natural History (FMNH), Chicago, Illinois; Kwa Zulu–Natal Sharks Board (KZN), Umhlanga, South Africa; and the collection of Dr. Gordon Hubbell, Gainsville, Florida (GHC) (). Specimens were a mix of tooth sets and dried articulated jaws. Each jaw or tooth set and each of the first seven teeth posterior to the symphysis within each jaw were photographed individually using an Olympus C3000 digital camera. Teeth were photographed such that the camera was orthogonal to the portion of the cusp adjacent to the tooth base. Images of teeth from the upper left and lower right jaws were reflected in about a vertical axis using Adobe Photoshop so that all teeth curved to the viewer's right for ease of comparison. Teeth were then digitized using TPSDig (computer program produced by F. J. Rohlf, which is freely available at http://life.bio.sunysb.edu/morph/). Seven landmarks were chosen from the labial side of each tooth to reflect the shape of the crown and the entire tooth (). The labial side was chosen because the majority of the extant specimens used in this study were from articulated tooth sets within jaws that do not expose the lingual side of teeth well. Broken teeth were not digitized. The landmark configurations were then superimposed using generalized least squares Procrustes superimposition (CitationRohlf and Slice, 1990). This procedure minimizes the variance between specimens through rotation, translation, and resizing (CitationZelditch et al., 2004). Procrustes superimposition was performed using CoordGen6 (part of Integrated Morphometrics Program [IMP] produced by H. D. Sheets; components of IMP are freely available at http://www2.canisius.edu/~sheets/morphsoft.html).
TABLE 1 Specimens used in morphometric analysis. : , Calvert Marine Museum; , Field Museum of Natural History; , Private collection of Gordon Hubbell; , Kwa-Zulu Natal Sharks Board; , United States National Museum of Natural History; , information not available; , female; , male.
FIGURE 2 Location of landmarks for geometric morphometric analysis. 1, cusp apex; 2, center of the inner edge of the root; 3, 4, junction of cusp edge and root; 5, maximum curvature of root center; 6, 7 apex of root lobe.
To determine if teeth of I. oxyrinchus and I. paucus could be differentiated from each other using geometric morphometric techniques, we used two approaches. The first step was to perform a resampling-based Goodall's F-test using TwoGroup6 (part of IMP) to determine if teeth from the two species are statistically different (400 bootstraps, α-level of P = 0.05). Following this, a canonical variates analysis (CVA) using CVAGen6 (part of IMP) was used to visualize which aspects of tooth morphology are the best discriminators between the two species. CVA creates a new coordinate system based on linear combinations of variables. Inter-group variation is used to scale the axes, making this suitable for visualizing differences among group means (CitationZelditch et al., 2004). Because CVA is based on eigenvalue decomposition, only one axis is possible when two groups are present.
Two sets of CVAs were performed using species as the grouping variable in each. The first was a single CVA disregarding tooth position. This grouping represents an unsorted group of Isurus teeth. The second set consisted of two separate CVAs: one for anterior teeth (tooth positions 1 and 2) and another for lateral teeth (tooth positions 4 through 7), taking advantage of the known pattern of notable monognathic heterodonty in these extant sharks (CitationShimada, 2002). The intermediate tooth position (position 3) was disregarded as it is a transitional morphology between the anterior and lateral regions of the jaw (CitationShimada, 2002). This a priori separation allows identification of morphological differences specific to the anterior and lateral teeth. To determine how well those aspects of morphology discriminate between the two groups, we used the reassignment test function in CVAGen6. It should be noted that this is not a formal statistical test, but rather is used as a general metric of the ability of identified morphological changes to differentiate between groups. This test computes the mean for each group, then compares the Mahalanobis distance between each individual and each group mean. Individuals are reassigned to the group with the closest mean (CitationZelditch et al., 2004). If two congeneric species that have very similar dental morphology can be differentiated using the above techniques, this method can then be used to differentiate fossil teeth of the same genus that are similar in morphology.
Analysis of I. hastalis and I. xiphodon
To determine if the teeth of I. hastalis and I. xiphodon differ in morphology indicating the possibility of different species, we applied the same analyses used for the extant species. Pictures of I. hastalis and I. xiphodon from CitationPurdy et al. (2001:figs. 27, 28a, b, f, 29), in addition to digital photographs of teeth from the NMNH and Calvert Marine Museum (CMM) collections were used (). In total, 54 I. xiphodon and 56 I. hastalis teeth were digitized from photographs of their lingual sides. Landmark data were subjected to Procrustes superimposition as described above. A resampling-based Goodall's F-test using TwoGroup6 determined whether or not teeth from the two species are significantly different (400 bootstraps, α-level of P = 0.05); CVA was then used to visualize how tooth morphology differs. One CVA was run on the pool of all teeth, and a second set of CVAs were run on separated lateral and anterior tooth sets. We separated these teeth by following the same heterodonty pattern shown in the extant species. Finally, to determine how well these aspects of morphology were able to distinguish between the two groups of teeth, the reassignment test function in CVAGen6 was employed.
RESULTS
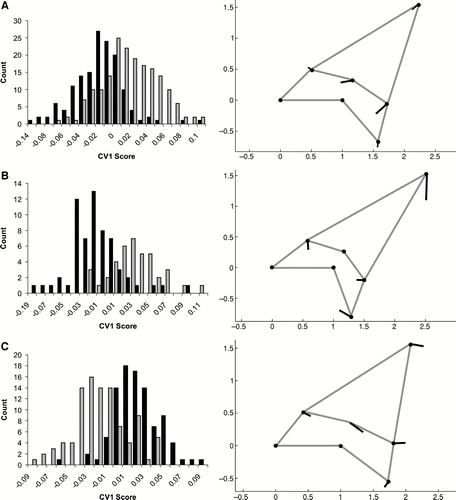
Despite considerable overlap of the canonical variates, the teeth from the two extant mako species, Isurus oxyrinchus and I. paucus, were significantly different (F = 9.29, P = 0.003) (). The differences represented by CV1 were slight. In general, teeth from I. oxyrinchus have more elongate, laterally angled cusps compared to those of I. paucus. When all teeth are considered together, the reassignment test assigned 72.9% of the teeth to the correct species. Performing separate CVAs on anterior and lateral teeth improved the assignment capability (). For the anterior teeth (positions 1 and 2), little separation between the two species occurs along CV1. Anterior teeth of I. oxyrinchus have narrower and slightly more laterally angled cusps compared to I. paucus. The lateral teeth (positions 4 through 7) are better at discriminating between species, with teeth of I. oxyrinchus having more laterally sheared bases. Correct species assignment occurred for 85.4% of the anterior teeth and 82.4% of the lateral teeth, indicating that a priori separation of anterior and lateral teeth provides better resolution of species differences.
FIGURE 3 Canonical variates analysis of Procrustes landmarks for I. oxyrinchus and I. paucus. Histograms on the left side of the figure are counts of CV1 scores. Black bars equal I. oxyrinchus; grey bars equal I. paucus. Vector plots on the right show the variation represented on CV1 as values become more positive. A, all tooth positions; B, anterior teeth only (positions 1 and 2); C, lateral teeth only (positions 4 through 7).
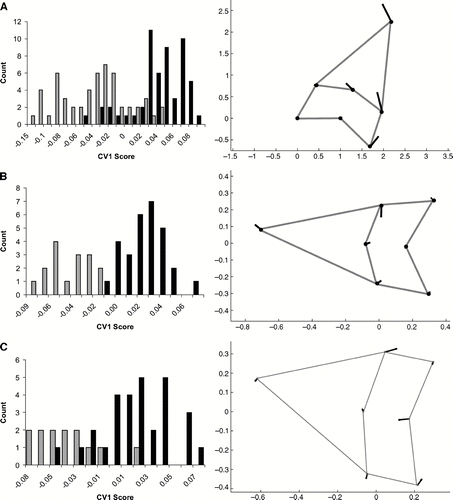
Teeth from Isurus hastalis and I. xiphodon were also found to be significantly different (F = 19.82, P = 0.003). The analysis of all fossil teeth showed more separation between the two species than the CVA of the extant species (). As shown by CV1, teeth from I. hastalis have more elongate, laterally angled cusps and a less curved and taller base than I. xiphodon. In this case, the reassignment test assigned 86.4% of the teeth to the corresponding species. Performing two separate CVAs improved assignment capabilities for anterior teeth, but not lateral teeth (). Anterior teeth of I. hastalis are slightly narrower than those of I. xiphodon. Lateral teeth of I. xiphodon have an apicobasally shortened base compared to I. hastalis. Whereas correct species assignments were made for 83.7% for the lateral teeth, 97.8% of the anterior teeth were assigned correctly.
FIGURE 4 Canonical variates analysis of Procrustes landmarks for I. hastalis and I. xiphodon. Histograms on the left side of the figure are counts of CV1 scores. Black bars equal I. hastalis; Grey bars equal I. xiphodon. Vector plots on the right show the variation represented on CV1 as values become more positive. A, all tooth positions; B, anterior teeth only (positions 1 and 2); C, lateral teeth only (positions 4 through 7).
DISCUSSION
Geometric morphometric techniques successfully distinguished between the teeth of the two extant mako species, Isurus oxyrinchus and I. paucus, despite the fact that the tooth morphologies are very similar. This suggests that this technique is applicable for analyzing teeth of two similar extinct putative species, I. hastalis and I. xiphodon. Differences in tooth morphology indicated by the canonical variates analysis for I. oxyrinchus and I. paucus were similar to those characters cited by CitationCompagno (2001): teeth of Isurus oxyrinchus have narrower cusps than I. paucus, though our two-dimensional analysis did not address recurving of the cusp apex. A difference in tooth base shape for the lateral teeth was also indicated by the CVA.
These techniques were also able to distinguish between the fossil mako species, Isurus hastalis and I. xiphodon. CitationPurdy et al. (2001) cite teeth of I. hastalis as having a convex mesial cutting edge and elongate root lobes, whereas teeth of I. xiphodon have a straight cutting edge, broader cusps, and shorter, more massive root lobes. The results of our CVA generally concur with this assessment of tooth morphology. The CVAs indicate that I. hastalis have narrow, laterally angled cusps and a less curved base, whereas I. xiphodon has broader, straighter cusps and a more curved base. Contrary to CitationPurdy et al. (2001), relative root lobe length was not an effective discriminator between the two species. When the results of the F-test and the CVA are taken together, our analysis supports I. hastalis and I. xiphodon as non-conspecific.
Despite this support, it is possible that other factors have contributed to the differences seen between I. hastalis and I. xiphodon. Statistical significance is not always equivalent to biological significance. As sample size increases, the ability of a statistical test to find small differences also increases. The F-tests ask whether there is a detectable difference between the mean shapes of the two groups being tested. Whether these differences reflect two species or two morphotypes should be considered anytime this type of testing is used. Some populations of modern elasmobranchs exhibit marked variation in tooth morphology among individuals, such as Carcharodon carcharias and Carcharhinus plumbeus (CitationNaylor and Marcus, 1994; CitationHubbell, 1996). Sexual heterodonty, with males and females possessing different tooth morphologies, also occurs in some modern elasmobranchs (CitationPeyer, 1968; CitationCompagno, 1988; CitationGottfried and Francis, 1996; CitationKajiura and Tricas, 1996). Geography may also play a role, as differences in tooth shape have been found among geographically distinct populations of Carcharias taurus (CitationLucifora et al., 2003). The modern specimens used in this study were taken from several locations around the world, including locations around the United States and South Africa. On the other hand, the fossil specimens came from two localities, Maryland (United States) and Peru. In this study, the CV plots show more separation between the two fossil species than in the modern species ( and ), indicating that there may be more of a difference in tooth morphology than between the modern established species and therefore supporting the separation of I. hastalis and I. xiphodon. It is possible that including more locations or new finds in future studies may challenge the results of our analysis. Indeed, this can be said of any study that erects a new species or attempts to examine taxonomic relationships between established fossil species.
Separating Isurus teeth into anterior and lateral groups based on previous analyses of heterodonty (CitationShimada, 2002) allowed for better resolution of differences in tooth morphology between species. The approach outlined here may also prove appropriate for other clades of sharks with both extant and extinct members by using information about extant heterodonty to perform a priori separations of teeth into bins. The genus Carcharhinus has been problematic in terms of fossil tooth identification. Many modern species have similar dental morphology (CitationCompagno, 1984, Citation1988). The fossil record shows similar trends, leading to misassignment and confusion (CitationNaylor and Marcus, 1994; CitationPurdy et al., 2001). For example, many Neogene teeth have been assigned to C. egertoni; however, it is likely that many belong to other species (CitationAdnet et al., 2007). Many species of Carcharhinus exhibit dignathic heterodonty, often with broader serrated teeth in the upper jaw and thin cusped teeth in the lower jaw in the regions close to the jaw symphysis (CitationCompagno, 1988). This would be an easy a priori separation for some species. The hexanchids would also benefit from this type of analysis, as these sharks also show strong dignathic heterodonty. CitationAdnet (2006) cites the need for a wide study of all fossil hexanchids to further elucidate phylogenetic relationships. Determining species validity would benefit this type of study, as there is debate over not only validity of extant species based on tooth morphology, but of fossil species as well (CitationAdnet, 2006a). Lastly, Isurus would also benefit from a wider geometric morphometric study. CitationCappetta (1987) lists eight fossil species of Isurus, which does not include the two extant species or I. xiphodon. CitationPurdy et al. (2001) suggest that I. paucus may be a junior synonym to I. hastalis, and that I. retroflexus may be a nomen dubium. CitationNyberg et al. (2006) cite a growing consensus that I. escheri should be split into two distinct species. The techniques outlined in this study may elucidate these relationships.
The delimiting of species can be difficult when dealing with extant organisms, species concept debates aside. In addition to molecular characters, suites of morphological characters are available from the entire body. Despite the abundance of data available for extant forms, species delimitations can still be cloudy if the characters used do not show much variation. This problem becomes more obvious when dealing with the fossil record. The vast majority of fossils supply limited sets of isolated morphological information that may or may not be useful for delimiting species. Often the designation of two very similar forms as one species or two is subjective, especially for fossil sharks, where the skeleton does not often fossilize. A survey of the literature from 2000 through 2007 to determine which criteria are most often used when designating or reevaluating fossil chondrichthyan species reveals that of the 45 papers published during this time period, 33 (73.3%) used only teeth (CitationBalbino and Cappetta, 2000; CitationPopov and Lapkin, 2000, Citation2001; CitationRees, 2000; CitationCappetta and Adnet, 2001; CitationSiverson and Cappetta, 2001; CitationCappetta and Stringer, 2002; CitationMurry and Kirby, 2002; CitationRees and Underwood, 2002; CitationSchwimmer et al., 2002; CitationUnderwood, 2002; CitationKriwet, 2003; CitationPopov, 2003; CitationIvanov and Rodina, 2004; CitationUnderwood and Ward, 2004; CitationCappetta and Nolf, 2005; CitationHoganson and Erickson, 2005; CitationMutter and Rieber, 2005; CitationRees, 2005; CitationRicher, 2005; CitationZiegler et al., 2005; CitationAdnet, 2006b; CitationAdnet et al., 2006, Citation2007; CitationConsoli, 2006; CitationDuffin and Ginter, 2006; CitationGarvey and Turner, 2006; CitationKriwet, 2006; CitationReinecke and Hoedemakers, 2006; CitationHampe and Ivanov, 2007; CitationMollen, 2007; CitationSiverson et al., 2007). Five (11.1%) used other characters in addition to teeth, such as fin spines and other parts of the skeleton (CitationFarres, 2002; CitationHovestadt and Hovestadt-Euler, 2002; CitationIvanov and Rodina, 2004; CitationThies, 2005; CitationMilner and Kirkland, 2006), and seven papers did not use teeth at all (15.6%) (CitationCoates and Sequeira, 2001; CitationHeidtke and Kraetschmer, 2001; CitationMarss and Gagnier, 2001; CitationStamberg, 2001; CitationMiller et al., 2003; CitationdeCarvalho et al., 2004; CitationMutter and Rieber, 2005). In 16 (42.1%) of the papers utilizing teeth, the tooth-related characters and comparisons described were solely qualitative. Twenty-two (57.9%) papers reported linear measurements such as height or width, but only four (10.5%) specifically used these measures to compare the new species to other taxa. No study employed rigorous statistical testing, though one paper plotted tooth length versus tooth width for the new and previously designated species (CitationReinecke and Hoedemakers, 2006). The use of geometric morphometrics may be useful in more objective species delimitation. In the papers cited above, many of the new species were described for genera without extant representatives. However, pools of teeth for the possible new species can be compared to similar contemporaneous species without specifically taking heterodontic patterns into account. In this study, F-tests and CVAs were both performed using only species as a grouping variable. This treats all teeth of one species, regardless of position within the jaw, as one group, allowing variation due to heterodonty to contribute to variance. Despite well-demonstrated dignathic and monognathic heterodonty, the F-test and CVA were able to separate known species and questionable fossil Isurus species when all teeth were grouped only by species.
CONCLUSION
Geometric morphometric techniques were able to successfully distinguish between teeth of the two extant species of Isurus, I. oxyrinchus and I. paucus, and between teeth of the two selected extinct species, I. hastalis and I. xiphodon. These results support I. xiphodon as a valid taxon. We propose that this type of quantitative analysis is applicable for examining species identification and delimitation among other lamnids and other clades of sharks with both extant and extinct members.
ACKNOWLEDGMENTS
We thank G. Cliff (Kwa Zulu–Natal Sharks Board), R. Purdy (NMNH), G. Hubbell (Gainesville, Florida), and M. Rogers (Field Museum of Natural History) for specimen loans. We also thank D. Sheets, M. Zelditch, P. Motta, G. Herbert, K. Shimada, J. Burris, D. Brandt, and R. Anstey for comments and help with this study. The manuscript was improved by comments from G. Naylor and an anonymous reviewer. The Michigan State University Graduate School and the University of South Florida student government supported this research.
Notes
†Current address: Department of Biology, Allegheny College, 520 North Main Street, Meadville, Pennsylvania, 16335, U.S.A.
Related Research Data
LITERATURE CITED
- Adnet , S. 2006a . Biometric analysis of the teeth of fossil and recent hexanchid sharks and its taxonomic implications . Acta Palaeontologica Polonica , 51 : 477 – 488 .
- Adnet , S. 2006b . Nouvelles faunes de selaciens (Elasmobranchii, Neoselachii) de l’Eocene moyen des Landes (sud-ouest, France); implication dans la connaissance des communautes de selaciens d’eaux profondes; in F. Pfeil (ed.), Palaeo-Ichthyologica, Volume 10 . Verlag Friedrich Pfeil, München , : 128
- Adnet , S. , Cappetta , H. and Reynders , J. 2006 . Nouveaux genres de Squaliformes (Chondrichthyes) du Paleogene des Landes (Sud-Ouest de la France) . Palaeontologische Zeitschrift , 80 : 60 – 67 .
- Adnet , S. , Antoine , P.-O. , Hassan Baqri , S. R. , Crochet , J.-Y. , Marivaux , L. , Welcomme , J.-L. and Metais , G. 2007 . New tropical carcharhinids (Chondrichthyes, Carcharhiniformes) from the Late Eocene–Early Oligocene of Balochistan, Pakistan: paleoenvironmental and paleogeographic implications . Journal of Asian Earth Sciences , 30 : 303 – 323 .
- Applegate , S. P. and Espinosa-Arrubarrena , L. 1996 . “ The fossil history of Carcharodon and its possible ancestor, Cretolamna: a study in tooth identification ” . In Great White Sharks: Biology of Carcharodon carcharias , Edited by: Klimley , A. P. and Ainley , D. G. 19 – 36 . San Diego : Academic Press .
- Balbino , A. C. and Cappetta , H. 2000 . Paragaleus antunesi (Hemigaleidae, Carcharhiniformes) a new shark species from the latest Miocene of Portugal . Tertiary Research , 20 : 1 – 6 .
- Becker , M. A. , Chamberlain , J. A. and Stoffer , P. W. 2000 . Pathologic tooth deformities in modern and fossil chondrichthyans: a consequence of feeding-related injury . Lethaia , 33 : 103 – 118 .
- Bogdanowicz , W. , Juste , J. , Owen , R. D. and Sztencel , A. 2005 . Geometric morphometrics and cladistics: testing evolutionary relationships in mega- and microbats . Acta Chiropterologica , 7 : 39 – 49 .
- Candioti , M. F. V. 2006 . Ecomorphological guilds in anuran larvae: an application of geometric morphometric methods . Herpetological Journal , 16 : 149 – 162 .
- Cappetta , H. 1987 . Chondrichthyes II. Handbook of Paleoichthyology , 191 New York : Gustav Fischer Verlag .
- Cappetta , H. and Adnet , S. 2001 . Decouverte du genre actuel Trigonognathus (Squaliformes; Etmopteridae) dans le Lutetien des Landes (sud-ouest de la France); remarques sur la denture de l’espece actuelle Trigonognathus kabeyai . Palaeontologische Zeitschrift , 74 : 575 – 581 .
- Cappetta , H. and Stringer , G. 2002 . A new batoid genus (Neoselachii, Mylobatiformes) from the Yazoo Clay (late Eocene) of Louisiana, U.S.A . Tertiary Research , 21 : 51 – 56 .
- Cappetta , H. and Nolf , D. 2005 . Revision de quelques Odontaspididae (Neoselachii, Lamniformes) du Paleocene et de l’Eocene du Bassin de la mer du Nord. Bulletin de l’Institut Royal des Sciences Naturalles de Belgique . Sciences de la Terre , 75 : 237 – 266 .
- Coates , M. I. and Sequeira , S. E. K. 2001 . A new stethacanthid chondrichthyan from the lower Carboniferous of Bearsden, Scotland . Journal of Vertebrate Paleontology , 21 : 438 – 459 .
- Compagno , L. J. V. 1984 . “ Sharks of the World ” . In An Annotated and Illustrated Catalogue of Shark Species Known To Date, Part 2: Carcharhiniformes; FAO Species Catalogue , 249 Rome : Food and Agriculture Organization of the United Nations .
- Compagno , L. J. V. 1988 . Sharks of the Order Carcharhiniformes , 570 Princeton : Princeton University Press .
- Compagno , L. J. V. 2001 . Sharks of the World: An Annotated and Illustrated Catalogue of Shark Species Known To Date, Volume 2: Bullhead, Mackerel and Carpet Sharks (Heterodontiformes, Lamniformes and Orectolobiormes) . : 269 pp FAO Species Catalogue for Fishery Purposes. Food and Agriculture Organization of the United Nations, Rome
- Consoli , C. P. 2006 . Edaphodon kawai, sp. nov. (Chondrichthyes: Holocephali): a Late Cretaceous chimaeroid from the Chatham Islands, southwest Pacfic . Journal of Vertebrate Paleontology , 26 : 801 – 805 .
- deCarvalho , M. R. , Maisey , J. G. and Grande , L. 2004 . Freshwater stingrays from the Green River Formation of Wyoming (early Eocene), with the description of a new genus and species and an analysis of its phylogenetic relationships (Chondrichthyes, Myliobatiformes) . Bulletin of the American Museum of Natural History , 284 : 1 – 136 .
- Duffin , C. J. and Ginter , M. 2006 . Comments on the selachian genus Cladodus Agassiz, 1843 . Journal of Vertebrate Paleontology , 26 : 253 – 266 .
- Farres , F. 2002 . Mesopristis nov. gen. osonensis nov. sp., new genus and species of fish; Eocene of Vic, Catalonia, northeastern Spain . Batalleria (Barcelona) , 11 : 93 – 113 .
- Garry , A. 2004 . Geometric morphometric analysis of shark teeth of the genus Rhizoprionodon: modern tooth shape analysis and test of ancestral prediction methods by comparison of fossil shapes . Abstracts with Programs—Geological Society of America , 36 : 315
- Garvey , J. M. and Turner , S. 2006 . Vertebrate microremains from the presumed earliest Carboniferous of the Mansfield Basin, Victoria . Alcheringa , 30 : 43 – 62 .
- Gottfried , M. D. and Francis , M. P. 1996 . Developmental changes in white shark tooth morphology: implications for studies on fossil sharks . Journal of Vertebrate Paleontology , 17 : 50
- Gottfried , M. D. and Fordyce , R. E. 2001 . An associated specimen of Carcharodon angustidens (Chondricthyes, Lamnidae) from the Late Oligocene of New Zealand, with comments on Carcharodon interrelationships . Journal of Vertebrate Paleontology , 21 : 730 – 739 .
- Hampe , O. and Ivanov , A. 2007 . First xenacanthid shark from the Pennsylvanian (Moscovian) of the Northern Caucasus (Russia) . Mitteilungen aus dem Museum fuer Naturkunde in Berlin , 10 : 179 – 189 .
- Heidtke , U. H. J. and Kraetschmer , K. 2001 . Gladbachus adentatus nov. gen. et sp., ein primitiver Hai aus dem Oberen Givetium (Oberes Mitteldevon) der Bergisch Gladbach, Paffrath-Mulde (Rheinisches Schiefergebirge) . Mainzer Geowissenschaftliche Mitteilungen , 30 : 105 – 122 .
- Hoganson , J. W. and Erickson , J. M. 2005 . A new species of Ischyodus (Chondrichthyes, Holocephali, Callorhynchidae) from upper Maastricthian marine fascies of the Fox Hills and Hell Creek formations, Williston Basin, North Dakota, USA . Palaeontology , 48 : 709 – 721 .
- Hovestadt , D. C. and Hovestadt-Euler , M. 2002 . The remains of a carchardinid shark with a new triakid species in its digestive track from the Oligocene of Germany . Tertiary Research , 21 : 171 – 182 .
- Hubbell , G. 1996 . “ Using tooth structure to determine evolutionary history of the white shark ” . In Great White Sharks: The Biology of Carcharodon carcharias , Edited by: Klimley , A. P. and Ainley , D. G. 9 – 18 . San Diego : Academic Press .
- Ivanov , A. and Rodina , O. 2004 . A new omalodontid-like shark from the Late Devonian (Famennian) of western Siberia, Russia . Fossils and Strata , 50 : 82 – 91 .
- Kajiura , S. M. and Tricas , T. C. 1996 . Seasonal dynamics of dental sexual dimorphism in the Atlantic Stingray, Dasyatis sabina . Journal of Experimental Biology , 199 : 2297 – 2306 .
- Kriwet , J. 2003 . Neoselachian remains (Chondricthyes, Elasmobranchii) from the Middle Jurassic of SW Germany and NW Poland . Acta Palaeontologica Polonica , 48 : 583 – 594 .
- Kriwet , J. 2006 . Biology and dental morphology of Priscusurus adruptodontus, gen. et sp. nov. (Chondrichthyes, Lamniformes) from the Albian (Early Cretaceous) of Peru . Journal of Vertebrate Paleontology , 26 : 538 – 543 .
- Lawley , R. 1878 . Confronto di denti fossili che si trovano nellecolline Toscane con la dentizione dell’ Oxyrhina spallanzanii Bonp. vivente in Mediterraneo , Vol. 3 , 343 – 349 . Memorie : Atti della Societa Toscana di Scienze Naturali Residente in Pisa .
- Leriche , M. 1926 . Les poissons Neogenes de la Belgique . Memoires du Musee Royal d’Histore Naturelle de Belgique , 32 : 367 – 472 .
- Lucifora , L. O. , Cione , A. L. , Menni , R. C. and Escalante , A. H. 2003 . Tooth row counts, vicariance, and the distribution of the sand tiger shark Carcharias taurus . Ecography , 26 : 567 – 572 .
- Marss , T. and Gagnier , P.-Y. 2001 . A new chondrichthyan from the Wenlock, lower Silurian, of Baillie-Hamilton Island, the Canadian Arctic . Journal of Vertebrate Paleontology , 21 : 693 – 702 .
- Miller , R. F. , Cloutier , R. and Turner , S. 2003 . The oldest articulated chondrichthyan from the Early Devonian period . Nature , 425 : 501 – 504 .
- Milner , A. R. C. and Kirkland , J. I. 2006 . Preliminary review of the Early Jurassic (Hettangian) freshwater Lake Dixie fish fauna in the Whitmore Point Member, Moenave Formation in southwest Utah . Bulletin—New Mexico Museum of Natural History and Science , 37 : 510 – 521 .
- Mollen , F. H. 2007 . A new species of Abdounia (Elasmobranchii, Carcharhinidae) from the base of the Boom Clay Formation (Oligocene) in northwest Belgium . Geologica Belgica , 10 : 69 – 77 .
- Murry , P. A. and Kirby , R. E. 2002 . A new hybodont shark from the Chinle and Bull Canyon formations, Arizona, Utah and New Mexico . Bulletin—New Mexico Museum of Natural History and Science , 21 : 87 – 106 .
- Mutter , R. J. and Rieber , H. 2005 . Pyknotylacanthus spathianus gen. et sp. nov., a new ctenacanthoid from the Early Triassic of Bear Lake (Idaho, USA) . Revista Brasileira de Paleontologia , 8 : 149 – 158 .
- Naylor , G. J. P. and Marcus , L. F. 1994 . Idenitfying isolated shark teeth of the genus Carcharhinus to species: relevance for tracking change through the fossil record . American Museum Novitates , 3109 : 1 – 53 .
- Nyberg , K. G. , Ciampaglio , C. N. and Wray , G. A. 2006 . Tracing the ancestry of the great white shark, Carcharodon carcharias, using morphometric analyses of fossil teeth . Journal of Vertebrate Paleontology , 26 : 806 – 814 .
- Oettle , A. C. , Pretorius , E. and Steyn , M. 2005 . Geometric morphometric analysis of mandibular ramus flexure . American Journal of Physical Anthropology , 128 : 623 – 629 .
- Peyer , B. 1968 . Comparative Odontology , 347 Chicago, Illinois : University of Chicago Press .
- Popov , Y. V. 2003 . A new genus of elephant fishes (Holocephali, Callorhinchidae) from the upper Callovian of the Volga region near Saratov, Russia . Palaeontological Journal , 37 : 507 – 513 .
- Popov , Y. V. and Lapkin , A. V. 2000 . A new shark species of the genus Galeorhinus (Chondrichthyes, Triakidae) from the Cenomanian of the lower Volga River basin . Palaeontological Journal , 34 : 435 – 438 .
- Popov , Y. V. and Yarkov , A. A. 2001 . A new giant species of Edaphodon (Holocephali; Edaphodontidae) from the Beryozovaya Beds (lower Paleocene) of the Volgograd Volga region . Palaeontological Journal , 35 : 183 – 187 .
- Pratt , H. L. and Casey , J. G. 1983 . Age and growth of the shortfin mako, Isurus oxyrinchus, using four methods . Canadian Journal of Fisheries and Aquatic Science , 40 : 1944 – 1957 .
- Pretorius , E. 2005 . Using geometric morphometrics to investigate wing dimorphism in males and females of Hymenoptera—a case study based on the genus Tachysphex Kohl (Hymenoptera: Sphecidae: Larrinae) . Australian Journal of Entomology , 44 : 113 – 121 .
- Purdy , R. , Schnieder , V. P. , Applegate , S. P. , McLellan , J. H. , Meyer , R. L. and Slaughter , B. H. 2001 . The Neogene sharks, rays, and bony fishes from Lee Creek Mine, Aurora, North Carolina. Geology and paleontology of the Lee Creek Mine, North Carolina, III . Smithsonian Contributions to Paleobiology , 90 : 71 – 202 .
- Rees , J. 2000 . A new Pliensbachian (Early Jurassic) neoselachian shark fauna from southern Sweden . Acta Palaeontologica Polonica , 45 : 407 – 424 .
- Rees , J. 2005 . Neoselachian shark and ray teeth from the Valanginian, Lower Cretaceous, of Wawal, central Poland . Palaeontology , 48 : 209 – 221 .
- Rees , J. and Underwood , C. J. 2002 . The status of the shark genus Lissodus Brough, 1935, and the position of nominal Lissodus species within the Hybodontoidea (Selachii) . Journal of Vertebrate Paleontology , 22 : 471 – 479 .
- Reinecke , T. and Hoedemakers , K. 2006 . Physogaleus hemmooriensis (Carcharhinidae, Elasmobranchii); a new shark species from the early to middle Miocene of the North Sea Basin . Palaeovertebrata , 34 : 1 – 25 .
- Richer , M. 2005 . A new xenacanthid shark (Chondrichthyes) from the Teresina Formation, Permian of the Paraná Basin, southern Brazil . Revista Brasileira de Paleontologia , 8 : 129 – 158 .
- Rohlf , F. J. and Slice , D. E. 1990 . Extensions of the Procrustes method for the optimal superimposition of landmarks . Systematic Biology , 39 : 40 – 59 .
- Rohlf , F. J. , Loy , A. and Corti , M. 1996 . Morphometric analysis of old world Talpidae (Mammalia, Insectivora) using partial-warp scores . Systematic Biology , 45 : 344 – 362 .
- Schwimmer , D. R. , Hooks , G. E. III and Johnson , B. 2002 . Revised taxonomy, age, and geographic range of the large lamniform shark Cretodus semiplicatus . Journal of Vertebrate Paleontology , 22 : 704 – 707 .
- Shimada , K. 2002 . Dental homologies in lamniform sharks (Chondrichthyes: Elasmobranch) . Journal of Morphology , 251 : 38 – 72 .
- Shimada , K. 2005 . Types of tooth sets in the fossil record of sharks, and comments on reconstructing dentitions of extinct sharks . Journal of Fossil Research , 38 : 141 – 145 .
- Siverson , M. and Cappetta , H. 2001 . A skate in the lowermost Maastrichthian of southern Sweden . Palaeontology , 44 : 431 – 445 .
- Siverson , M. , Lindgren , J. and Kelley , L. S. 2007 . Anacoracid sharks from the Albian (Lower Cretaceous) Pawpaw Shale of Texas . Palaeontology , 50 : 939 – 950 .
- Stamberg , S. 2001 . Fin spine of a ctenacanthoid shark (Elasmobranchii, Ctenacanthoidea) from the upper Stephanian of the Krkonose piedmont basin; Bohemia . Bulletin of Geosciences (Praha) , 76 : 141 – 148 .
- Thies , D. 2005 . Catshark (Neoselachii, Carcharhiniformes, Scyliorhinidae) from the Late Jurassica of Germany . Palaeontologische Zeitschrift , 79 : 339 – 348 .
- Underwood , C. J. 2002 . Sharks, rays, and a chimaeroid from the Kimmeridgian (Late Jurassic) of Ringestead, southern England . Palaeontology , 45 : 297 – 325 .
- Underwood , C. J. and Ward , D. J. 2004 . Neoselachian sharks and rays from the British Bathonian (Middle Jurassic) . Palaeontology , 47 : 447 – 501 .
- Ward , D. J. and Bonavia , C. G. 2001 . Additions to, and a review of, the Miocene shark and ray fauna of Malta . The Central Mediterranean Naturalist , 3 : 131 – 146 .
- Zelditch , M. L. , Lundrigan , B. L. , Sheets , H. D. and Garland , T. Jr . 2003 . Do precocial mammals develop at a faster rate? A comparison of rates of skull development in Sigmodon fulviventer and Mus musculus domesticus . Journal of Evolutionary Biology , 16 : 708 – 720 .
- Zelditch , M. L. , Swiderski , D. L. , Sheets , H. D. and Fink , W. L. 2004 . Geometric Morphometrics for Biologists: A Primer , 443 New York : Elsevier Academic Press .
- Ziegler , K. E. , Lucas , S. G. and Heckert , A. B. 2005 . A new genus of Rhombodontidae (Chondricthyes, Myliobatiformes) from the Eocene of Wyoming, U.S.A. Neues Jahrbuch fuer Geologie und Palaeontologie . Monatschefte , 2005 : 373 – 384 .