ABSTRACT
The fossil record of dyrosaurid crocodyliforms spans the Late Cretaceous to Middle Eocene of Africa, Asia, Europe, North America, and South America. Prior to this study, specimens from South America have been limited to a few fossils with only two taxa diagnosed. We describe a nearly complete skull and unassociated mandible of a new dyrosaurid, Cerrejonisuchus improcerus gen. et sp. nov., from the Paleocene Cerrejón Formation of northeastern Colombia. The skull of C. improcerus has relatively elongate supratemporal fenestrae and well-developed occipital tuberosities, both diagnostic characteristics of Dyrosauridae. The rostrum of adult C. improcerus comprises 54–59% of the length of the skull, making it the shortest snout of any known dyrosaurid. A cladistic analysis using 82 cranial and mandibular characters for all species of Dyrosauridae known from crania yielded two most-parsimonious cladograms with C. improcerus as the sister taxon to a clade including Arambourgisuchus, Dyrosaurus, Hyposaurus, Congosaurus, Rhabdognathus, Atlantosuchus, and Guarinisuchus. Only Chenanisuchus, Sokotosuchus, and Phosphatosaurus, all known only from Africa, are more primitive within Dyrosauridae. Chenanisuchus from the Paleocene of Morocco, the only other known short-snouted dyrosaurid, is not closely related to C. improcerus and a short-snouted condition appears to have evolved independently at least twice within Dyrosauridae. Our analysis supports an African origin of Dyrosauridae with dispersals to the New World by the Late Cretaceous or earliest Paleocene. The presence of C. improcerus, together with undescribed taxa from the Cerrejón Formation, suggests a radiation of dyrosaurid crocodyliforms, possibly following the K-P boundary, in tropical South America.
INTRODUCTION
Dyrosauridae is an extinct family of mesoeucrocodylians typically found in transitional marine sediments from the Late Cretaceous through Late Eocene (CitationBrochu et al., 2002). The family was named by Giuseppe De Stefano in 1903 for the type genus Dyrosaurus, named by A. Pomel for the locality of the holotype, Djebel Dyr, near Tébessa, Algeria (CitationPomel, 1894). Dyrosaurid crocodyliforms are well known from the Late Cretaceous to Eocene of northern Africa and southwestern Asia (e.g., CitationSwinton, 1930, Citation1950; CitationArambourg, 1952; CitationHalstead, 1973, Citation1975; CitationHalstead and Middleton, 1976; CitationBuffetaut 1976a, Citation1976b, Citation1977, Citation1978, Citation1979, Citation1980; CitationStorrs 1986; CitationBuffetaut et al., 1990; CitationLangston, 1995) as well as the Late Cretaceous to Paleocene of eastern North America (e.g., CitationTroxell, 1925; CitationParris, 1986; CitationDenton et al., 1997), and possibly the Late Cretaceous of Europe (CitationBuffetaut and Lauverjat, 1978). However, the fossil record of dyrosaurids in South America is more limited.
Fossils of dyrosaurid crocodyliforms have previously been recovered from only four localities in South America. Hyposaurus derbianus is known from somewhat fragmentary fossils from the Maria Farinha Formation in Pernambuco, Brazil (CitationCope, 1885, Citation1886). The Maria Farinha Formation was originally thought to be Late Cretaceous (CitationCope, 1885, Citation1886; CitationBuffetaut, 1976a, Citation1980), but has been more recently placed in the Paleocene (CitationAlbertâo et al., 1993). CitationArgollo et al. (1987) described a Late Cretaceous or Paleocene crocodyliform assemblage from Huarachani in Bolivia that included two teeth, two osteoderms, and a vertebra attributed to Dyrosauridae gen. et sp. indeterminate. CitationMarshall and de Muizon (1988) produced a faunal list from Tiupampa, Bolivia, that included a crocodyliform referred to the dyrosaurid Sokotosuchus aff. ianwilsoni and thought to be Late Cretaceous in age. Fossils from this site were later referred to Dyrosauridae gen. et sp. indet. and considered Paleocene in age (CitationBuffetaut, 1991). A mandibular symphysis of Sulcusuchus erraini from Patagonia was attributed to Dyrosauridae (CitationGasparini and Spalletti, 1990; CitationGasparini, 1996), but was later referred to Plesiosauroidea (CitationGasparini and de la Fuente, 2000) outside of Crocodylomorpha (CitationGasparini, 2007). Additional dyrosaurid specimens from the Paleocene part of the Maria Farinha Formation have been found from the same unit that H. derbianus was discovered, including some isolated teeth and vertebrae (CitationCarvalho and Azevedo, 1997; CitationGallo et al., 2001). CitationBarbosa et al. (2008) also recently described a new genus and species, Guarinisuchus munizi, based on a complete skull and partial lower jaw from the Maria Farinha Formation. Most of these sites are south of the ancient tropical zone (CitationScotese, 2001; ). Thus very little is known of the diversity of Dyrosauridae within South America, and little is known of crocodyliforms as a whole for the ancient neotropics prior to the Eocene.
FIGURE 1 World map of locations of known crocodyliform fossil material during the Paleocene. Circles represent locations ascertained as Paleocene in age, squares represent locations that have been contested as possibly Late Cretaceous in age, and the star represents the new locality in northeastern Colombia. Map of the Paleocene by Scotese, 2001.
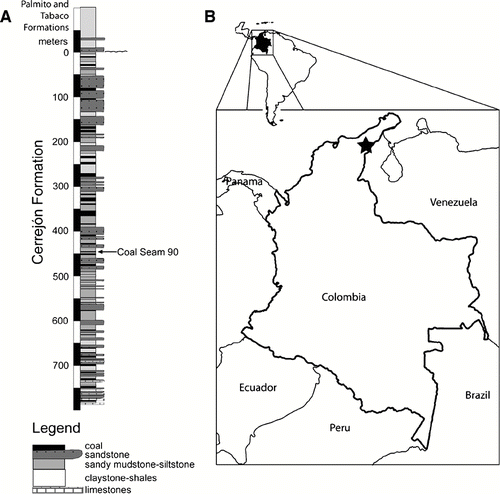
Here we describe dyrosaurid fossils from the Cerrejón coal mine in northeastern Colombia (CitationBloch et al., 2005; CitationHastings and Bloch, 2007, Citation2008). The fossil-bearing locality outcrops in the La Puente Pit and is situated in the underclay of Coal Seam 90 in the Cerrejón Formation. The age of the Cerrejón Formation has been estimated to be middle–late Paleocene based on carbon isotopes, pollen, spores, and dinoflagellate cysts (; Jaramillo et al., 2007). The surrounding matrix is comprised of bituminous coal and gray, fine-grained clay. The paleoenvironment of the section from which the fossils were recovered is transitional, with likely brackish water in a riverine-to-lagoonal setting. The environmental conditions implied by the geology of the site, as well as associated faunas, including large freshwater turtles, a giant boid snake, and dipnoan and elopomorph fishes (CitationBloch et al., 2008; CitationHead et al., 2009), are consistent with a transitional marine-freshwater environment. The environment of the dyrosaurids in Colombia is likely more inland than the estuarine environment of the dyrosaurid locality within the Umm Himar Formation of Saudi Arabia (CitationLangston, 1995), but nearer to the coast than the fluvial setting of the northern Sudan dyrosaurid locality (CitationBuffetaut et al., 1990).
FIGURE 2 Location and stratigraphic column from which the new fossil material was discovered. A, stratigraphic column including the layer that yielded the fossils herein described, marked by an arrow. B, map of Colombia, star marks location of the field site from which crocodyliform fossils were recovered. Stratigraphic column modified from Jaramillo et al., 2007.
We describe a new dyrosaurid based on three skulls, a lower jaw, and associated postcrania. Another 10 individual crocodyliforms, represented by at least somewhat complete cranial or mandibular material of at least two additional taxa of very different morphotypes, have also been recovered from the Cerrejón Formation (e.g., CitationHastings and Bloch, 2008) and will be described elsewhere.
Institutional Abbreviations—AMNH, American Museum of Natural History; IGM, Museo Geológico, at the Instituto Nacional de Investigaciones en Geociencias, Minería y Quimica, Bogotá, Colombia; UF, Florida Museum of Natural History, University of Florida; YPM, Yale Peabody Museum.
Terminology and Anatomical Abbreviations—Teeth and alveoli will be referred to by numbers, with 1 being the most anterior. Premaxillary teeth and alveoli will have the initial ‘pm,’ maxillary teeth and alveoli will use ‘m,’ and dentary teeth and alveoli will use ‘d.’ For example, the first tooth or alveolus of the premaxilla will be referred to as ‘pm1’ and the second tooth or alveolus of the maxilla will be referred to as ‘m2.’ This nomenclature is modified from CitationSereno et al. (2001) and de Lapparent de Broin (2002).
SYSTEMATIC PALEONTOLOGY
CROCODYLOMORPHA CitationWalker, 1970
CROCODYLIFORMES CitationHay, 1930
MESOEUCROCODYLIA CitationWhetstone and Whybrow, 1983
DYROSAURIDAE Citationde Stefano, 1903
CERREJONISUCHUS, gen. nov.
Etymology—Named for the Cerrejón Formation from which the fossils were recovered within the Cerrejón coal mine on the Guajira Peninsula of northeastern Colombia and -suchus, Greek for crocodile.
Type and Only Known Species—Cerrejonisuchus improcerus
Range—Middle–late Paleocene, Colombia
Diagnosis—As for type and only known species.
CERREJONISUCHUS IMPROCERUS, sp. nov.
Etymology—improcerus, Latin for diminutive, an allusion to not only its relatively short snout, but also its relatively small body size.
Holotype—UF/IGM 29, a nearly complete skull including the entire snout, 11 teeth, a complete dorsal skull table (postorbital, squamosal, parietal, and frontal) and a partial occipital region including exoccipitals, basioccipital, and partial basisphenoid.
Type Locality and Horizon—All known specimens are from the Cerrejón Formation, underclay of Coal Seam 90 at the La Puente Pit within the Cerrejón Coal Mine in Northeastern Colombia. Middle to Late Paleocene in age. Latitude 11.08.45.50 N, longitude 72.32.55.56 W.
Referred Specimens—UF/IGM 30, lower jaw including dentaries and splenials and a total of 11 partial teeth. Latitude 11.08.38.93 N, longitude 72.33.18.54 W. Additional unprepared fossils include: UF/IGM 31, a nearly complete skull with at least four teeth and associated postcranials including a humerus, ulna, left femur, fibula, tibia, left and right pubi, 17 vertebrae, one rib, and eight osteoderms; UF/IGM 32, a complete snout and partial orbital region.
Diagnosis—Shorter snout, approximately 54–59% of the dorsal skull length, than that of all other known dyrosaurids. Also differs from all other dyrosaurids in having approximatly 11 teeth in each maxillary, 8 of which are anterior to the orbits. Further differs from all known dyrosaurids except Chenanisuchus in having a wide interfenestral bar that is square-shaped in cross-section. Further differs from all other dyrosaurids except Phosphatosaurus (and possibly Arambourgisuchus) in having a reduced fourth premaxillary tooth. Further differs from Phosphatosaurus and Sokotosuchus in lacking a ‘festooned’ lateral margin of the snout in dorsal view. Further differs from Hyposaurus, Rhabdognathus, Atlantosuchus, and Guarinisuchus in having a mediolaterally straight posterodorsal margin of the parietal. Further differs from Chenanisuchus and Sokotosuchus in having well-developed occipital tuberosities. Further differs from Chenanisuchus in having ornamentation continuous across dorsal and lateral surfaces with no interruption across sutures and orbits medially and dorsally placed, most closely approximating the orbit position of Dyrosaurus. Further differs from Hyposaurus rogersii in having teeth with straight, rather than twisted anterior carinae.
Preservation
Although the type specimen (UF/IGM 29) is slightly dorsoventrally compressed, it is a nearly complete skull, preserving most cranial elements from the terminus of the premaxillae to the occipital condyle, with many features preserved in three dimensions (Figs. and ). The posterior portion of the palatines, both pterygoids, both ectopterygoids, the internal choanae, prootic bones, and quadratojugals are missing in UF/IGM 29 (). The lateral margin of the right quadrate is missing, as is the posterior portion of the jugal.
FIGURE 3 Skull of Cerrejinosuchus improcerus, UF/IGM 29, from the Cerrejón coal mine of northeastern Colombia, middle–late Paleocene, in dorsal view. A, photograph; B, sketch. Abbreviations: bo, basioccipital; en, external nares; eo, exoccipital; f, frontal; ifb, interfenestral bar; j, jugal; l, lacrimal; lsp, laterosphenoid; m, maxilla; n, nasal; or, orbit; ot, occipital tuberosity; p, parietal; pm3, third premaxillary tooth; pm, premaxilla; po, postorbital; prf, prefrontal; q, quadrate; sq, squamosal; stf, supratemporal fenestra; vt, ventral tubercle. Dotted line represents a suture which was not clear. Scale bar equals 10 cm.
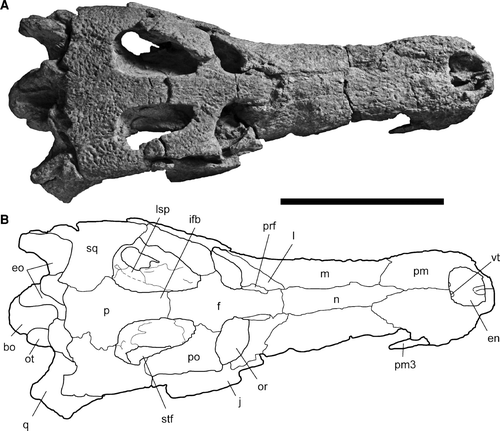
FIGURE 4 Skull of Cerrejinosuchus improcerus, UF/IGM 29, from the Cerrejón coal mine of northeastern Colombia, middle–late Paleocene. A, UF/IGM 29 in lateral view; B, UF/IGM 29 in occipital view; C, UF/IGM 29 sketch of occipital view. Abbreviations: bo, basioccipital; car, carotid foramen; eo, exoccipital; fm, foramen magnum; fv, foramen vagi; oc, occipital condyle; ot, occipital tuberosity; p, parietal; prp, paroccipital process; ptf, posttemporal fenestra; q, quadrate; so, supraocciptal; sq, squamosal. Both scale bars equal 10 cm.

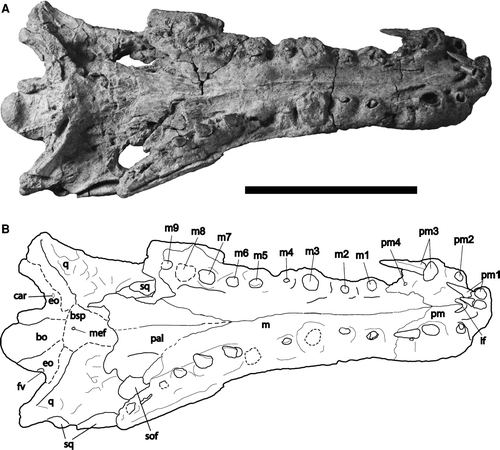
FIGURE 5 Skull of Cerrejonisuchus improcerus, UF/IGM 29, from the Cerrejón coal mine of northeastern Colombia, middle–late Paleocene, in ventral view. A, photograph; B, sketch. Abbreviations: bo, basioccipital; bsp, basisphenoid; car, carotid foramen; eo, exoccipital; fv, foramen vagi; if, incisive foramen; j, jugal; m, maxilla; m1–9, first through ninth maxillary alveoli/teeth; mef, medial eustachian foramen; pal, palatine; pm, premaxillary; pm1–4, first through fourth premaxillary alveloi/teeth; q, quadrate; sof, suborbital fenestra. Dotted lines represent features/sutures that were not clear. Scale bar equals 10 cm.
Description
General—Unless otherwise noted, all descriptions and measurements are taken from the holotype (UF/IGM 29; Figs. –). The snout is relatively narrow and comprises about 54–59% of the dorsal skull length, measured from the tip of the snout to the posterior margin of the parietal (). The lateral margins of the snout remain parallel in dorsal view from the external nares to anterior to the orbits (). Ornamentation consists of shallow pits consistent on the dorsal surfaces () as well as the preserved lateral surfaces of the specimen (), and is consistent across cranial bone sutures.
TABLE 1 Snout proportions of all members of Dyrosauridae with material complete enough for skull length estimation. Notably absent is Rhabdognathus aslerensis, which lacks most of the front of the snout needed to estimate dorsal skull length as well as preorbital skull length. Congosaurus bequaerti is only known from the anterior portion of the skull (CitationJouve and Schwarz, 2004); however, dorsal skull length and body length were estimated by CitationJouve et al. (2008). , dorsal skull length; , preorbital skull length; , ratio of preorbital skull length to dorsal skull length; , estimated total body length using method by CitationSereno et al. (2001). Citations are marked by numbered superscripts: 1Jouve et al., 2005a; 2an estimation from CitationJouve et al., 2008; 3an estimation from figure 2 in Barbosa et al., 2008; 4Jouve et al., 2005b; 5Jouve et al., 2006; 6 CitationJouve et al., 2008.
Cranial Openings—The external nares are mediolaterally oval, oriented dorsally, and surrounded exclusively by the premaxillae (). The external nares are located extremely anteriorly, with only a thin wall (6.57 mm) of the premaxillae around the anterior edge. These conditions are consistent across all specimens and are not likely the result of deformation. The anterior lateral margins are angled toward the midline of the snout. The lateral margins of the external nares are approximately level with the posterior and anterior margins, and do not slope ventrally or dorsally (Figs. , ). Two low ventral ridges extend posteriorly from the anterior margin of the external nares, paralleling what would have been the midline of the snout (). At the posterior end of the external nares are two small tubercles on the ventral surface of the external nares projecting from the posterior wall (). These tubercles are also present, but partly eroded, in UF/IGM 31 and 32 (Figs. and ), and therefore the feature seems representative of the species.
FIGURE 6 Referred skull of Cerrejonisuchus improcerus, UF/IGM 31, from the Cerrejón coal mine of northeastern Colombia, middle–late Paleocene, in dorsal view. A, UF/IGM 31 photograph; B, UF/IGM 31 sketch. Abbreviations: bo, basioccipital; en, external nares; eo, exoccipital; f, frontal; ifb, interfenestral bar; itf, infratemporal foramen; j, jugal; l, lacrimal; m, maxilla; n, nasal; or, orbit; ot, occipital tuberosity; p, parietal; pm3, third premaxillary tooth; pm, premaxilla; po, postorbital; prf, prefrontal; q, quadrate; qj, quadratojugal; sq, squamosal; stf, supratemporal fenestra; vert, vertebra vt, ventral tubercle. Dotted lines represent sutures that were not clear. Specimen has a partial thoracic vertebra fused to the ventral side of the skull as a result of deformation, visible also in dorsal view. Scale bar equals 10 cm.
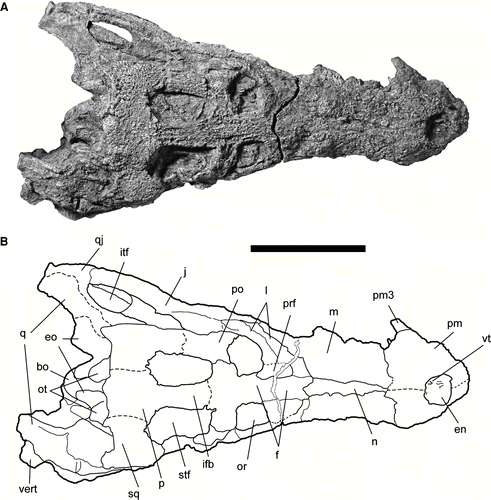
FIGURE 7 Referred snout of Cerrejonisuchus improcerus, UF/IGM 32, from the Cerrejón coal mine of northeastern Colombia, middle–late Paleocene. A, UF/IGM 32 in dorsal view; B, UF/IGM 32 in ventral view. Scale bar equals 5 cm.

Due to anteroventral flattening, the incisive foramen has been displaced laterally from the midline of the external nares (). The incisive foramen is entirely surrounded by the premaxillae. The incisive foramen is visible in dorsal view through the external nares as a small pit between the low anterior ridges (). In ventral view, the foramen has a triangular anteriorly directed process placed medially over the foramen, between the pm1 teeth (). The entire foramen is level with the pm2 alveoli.
The orbits of UF/IGM 29 are oval in outline and oriented anterodorsally, near the midline (Figs. , ). The prefrontals participate in the anteromedial corners of the orbits. The lateral margins of the orbits are comprised of the lacrimal, with the jugal only participating in the posterolateral corners. The medial margins of the orbits are entirely comprised of the frontal bone. The posterior margins of the orbits are slightly more than half comprised of the postorbitals that have prominent anterolateral processes. Less than half of the remaining posterior orbital margins are formed by the frontal (). The separation between the orbits by the frontal is approximately one-third the width of an orbit.
Flattening has distorted much of the infratemporal fenestrae of the holotype (). However, UF/IGM 31 has preserved the left infratemporal fenestra (). This fenestra is elongate, anteroposteriorly longer than it is dorsoventrally wide. The postorbitals constitute the anterodorsal portions of the fenestrae. Along the ventral margins of the infratemporal fenestrae the jugals rise dorsally at a point nearly even with the postorbital-squamosal joint and approximately halfway along the margin (Figs. , ). The posterodorsal corner in UF/IGM 31 is bound by the quadratojugal ().
The supratemporal fenestrae are 51.8% longer than they are wide and narrow anteriorly to 63.7% of the width across the posterior fenestrae (). The anterior and posterior margins of the supratemporal fenestrae are rounded, making the fenestrae longitudinally ovular with a straight medial margin, and lateral margins that taper anteriorly toward the midline. The fenestrae are bound anteromedially by the frontal. The postorbitals constitute the lateral halves of the anterior margins as well as the anterior halves of the lateral margins. The parietal forms approximately the medial three-quarters of the posterior margins of the supratemporal fenestrae. The posterior halves of the lateral margins, as well as the remaining one-quarter of the posterior margins, of the fenestrae are bound by the squamosal. The posterior walls of the supratemporal fenestrae are weakly sloped anteroventrally from the skull roof and are largely visible in dorsal view (). This feature may have been exaggerated slightly from dorsoventral flattening, but a significant amount of exposure is evident even in the less flattened specimens. The interfenestral bar is very wide, 24.60 mm at its widest and 16.71 mm at its narrowest. The cross-section of the interfenestral bar is rectangular in shape, and not T-shaped.
The temporal canals have largely been lost. The right parietal-squamosal suture is incomplete and slightly separated with a subtle widening of a crack (0.40 mm) at roughly the expected level of the temporal canal. This depression likely represents the cavity that was the temporal canal and is dorsolaterally bound by the squamosal and medioventrally bound by the parietal.
The posttemporal fenestrae are located dorsal to the occipital tuberosities and are oriented mediolaterally, and parallel to the skull roof (). The posttemporal fenestrae are typically dorsoventrally short among dyrosaurids (CitationJouve, 2005), but due to taphonomic dorsoventral distortion, this characteristic has been further exaggerated. The fenestrae are bound dorsolaterally by the squamosals and dorsomedially by the parietal. The ventral surface is formed by the occipital tuberosites of the exoccipitals. The supraoccipital is not complete, but evidently had little to no participation to the posttemporal fenestrae.
Much of the palatines have been crushed and/or lost, and both ectopterygoids are absent, thus little can be discerned of the suborbital fenestrae (). A small portion of the medial and anterolateral margins of the right suborbital fenestrae remain. The preservation of the right suborbital fenestra includes the anterolateral margin formed by the maxilla, and ceases anteriorly at the anteromedial corner, comprising about two-thirds of the anterior margin of the fenestra. This margin is round and smooth, with a slight protrusion of the palatine into the fenestra in the middle of the medial margin, near the m9 alveolus (). The fenestra reaches anteriorly to the level of the m8 alveolus. In addition, one-third of the anteromedial margin of the left suborbital fenestra is also present, where the palatine has been flattened so that it is flush dorsally against the braincase. Posteriorly the preservation of the left suborbital fenestra ceases with the posteromost preservation of the left maxilla, which is at the level of the m11 alveolus. The suborbital fenestrae are narrow and slender in shape and anteriorly acute. The suborbital fenestrae are very posteriorly placed, and their anterior margins do not reach the level of the orbit.
The foramen magnum has been crushed dorsoventrally, only leaving a shallow depression (). The foramen is bound laterally and dorsally by the exocciptals, comprising three-quarters of the margin. The ventral margin is formed by the basioccipital.
Premaxillae—The premaxillae are well preserved in UF/IGM 29. The anterior end of the snout has been dorsoventrally compressed with slight warping toward the left side (Figs. , ). This has resulted in five premaxillary teeth displaced from, but still adjacent to, their respective alveoli (). The premaxillae do not expand laterally with respect to the maxillae. The premaxilla-maxilla suture is level with the pm3 alveoli on the dorsal and ventral surfaces (Figs. , ). The posterodorsal process extends posteriorly to the interalveolar space between the m1 and m2, and penetrates between the maxillae and nasal (). Four alveoli are present on each premaxilla. The left pm1 tooth is notably shorter in length from the right (8.44 mm vs. 15.43 mm), likely due to different stages in replacement. The teeth of pm1 are placed very close together (2.11 mm apart). A deep gap is present between the pm1 and pm2 alveoli, large enough for lower dentition to occlude. These deep notches are visible on the dorsal surface as slight indentations along the anteromost margin of the snout (). The pm3 alveoli are notably raised from the adjacent alveoli and much more robust (). Measurements of alveolar size and spacing are provided in . The premaxillae have shallow pitted ornamentation, with anterior ornamentation directed toward the midline.
Maxillae—The maxillae of UF/IGM 29 comprise most of the snout. The left maxilla has 11 preserved alveoli, the right maxilla has 9 variably preserved alveoli (). The terminal end of the left maxilla represents the end of this bone posteriorly where it would have fused with the ectopterygoid that was not preserved. An indentation is present just posterior to the premaxilla-maxilla suture along the lateral snout margin, visible in both dorsal and ventral views, particularly on the right side (Figs. , ). The maxillae have smooth, even margins that gradually expand posterolaterally (). The maxillae are dorsally separated by the nasal (). The two maxillae contact each other posteriorly until the level of the interalveolar space between m4 and m5 where the maxillae are medially separated by the palatines and extend posterolaterally. Posteriorly, the maxillae extend to the level of the posterior margins of the supratemporal fenestrae, with the posteromost alveolus being just anterior to this level. In addition, the maxillae contribute posteriorly to the anteroventral portion of the lower temporal bar (). The alveoloar walls project ventrally, forming raised walls above the interalveolar space. A deep sulcus is located just anterior to the m1 alveolus (). Of the preserved dentition, the m3 alveoli are by far the largest. Interalveolar spaces are fairly consistent, and typically less than one-third the length of the adjacent alveoli. Deep sulci are present between the m4 and m5 alveoli, deeper than those between the pm4 and m1. Smaller, yet still deep, occlusal pits are found between the m3 and m4 alveoli as well as between the m5 and m6 alveoli. The maxillary alveoli are mostly circular in outline, but especially posteriorly they have a slight tendency toward an anteroposterior ovate shape. See for alveolar size and spacing. A small ridge is visible in ventral view along the suture between the left and right maxillae (). The maxillae are dorsally and laterally ornamented (Figs. , ).
Nasal—The nasals are fused in UF/IGM 29 () and UF/IGM 31 (), but are clearly unfused in UF/IGM 32 (). As UF/IGM 29 and 31 are similarly sized, and UF/IGM 32 is slightly smaller, this character may be related to ontogeny. The single nasal of UF/IGM 29 gradually widens from its anterior point to 12.99 mm at its posteromost contact with the premaxilla. The nasal contacts the premaxillae dorsally at the level of the pm3 alveolus. Anteriorly, the nasal comes to a point near the narial opening, with only a small space (20.46 mm) along which the premaxillae fuse together before the posterior margin of the external nares, such that the nasal does not participate in the posterior margin of the external nares (). The width is relatively constant; from this point of contact with the premaxilla (13.03 mm wide) until the first point of contact with the lacrimal (15.70 mm wide), a 17% increase in width. The area where the nasal joins the frontal, lacrimal, and prefrontal is distorted, with the posterior end of the nasal overlapping the anterior end of the frontal bone. The posterior end of the nasal terminates at the level of the m5 alveoli. Due to lack of preservation, whether or not the nasal bifurcates into two posterior processes cannot be discerned in UF/IGM 29. The posterior nasals are better preserved in UF/IGM 31 and 32 and do not bifurcate (). The nasal does not contact the orbit in either UF/IGM 29 or UF/IGM 32. The ornamentation of the nasal is relatively shallow and uniform.
Prefrontals—The size disparity between the prefrontals of UF/IGM 29 and 31 is 47% (∼16.7 mm anteroposterior length in UF/IGM 29 and 34.8 mm in UF/IGM 31) despite only a 17.8% difference in skull length (see ). The prefrontals of UF/IGM 32 are incomplete, but appear more similar to the large condition seen in UF/IGM 31. In all specimens, both prefrontals have a straight contact with the frontal, parallel to the midline.
Lacrimals—The left lacrimal of UF/IGM 29 is more complete than the right and bears a very prominent long and narrow ridge ascending from its contact with the left jugal. A portion of this long, narrow ridge is present on the right side, but neither its contact with the jugal, prefrontal, or nasal is preserved. The prominent lateral ridge composing the lateral margin of the orbit widens posterolaterally. The lacrimal constitutes most of the lateral margin of the orbit, with the jugal comprising only the posteromost portion. The lacrimal expands anteriorly and curves medially toward the midline, constituting much of the semicircular anterior margin of the orbit. The lacrimal extends anteriorly to the level of the m4 alveolus. The left lacrimal's length of contact with the prefrontal, 14.65 mm, is greater than that with the nasal, 10.72 mm. The ornamentation of the lacrimal is limited to the lateral surface.
Frontal—The single cruciform frontal of UF/IGM 29 is nearly complete, except where the frontal-nasal suture was not preserved (), and whether or not the frontal penetrates the nasal is unclear. The frontal reaches farther than the prefrontals anteriorly (Figs. , ). The frontal participates in the anterior 28% of the thick interfenestral bar. In cross-section, the anterior projection of the frontal is roughly triangular, with the point directed ventrally, and is roughly as wide as the interfenestral bar. The frontal contributes to the margins of both orbits and both supratemporal fenestrae. The bone comprises the entire medial margin of the orbit as well as the medial half of the posterior margin (). The suture with the postorbital is at the midline of the orbit. The participation of the frontal to the posteromedial corner of the orbit is a smooth, semicircular curve. The parietal-frontal contact occurs along the dorsal surface of the interfenestral bar and extends anterolaterally and ventrally through the supratemporal fenestra. The contact with the laterosphenoid is located anterior to the dorsal frontal-parietal suture. The anteromedial margin of the supratemporal fenestra formed by the frontal is slightly overhung. However, the interfenestral bar is barely overhung. On the ventral surface of the anterior portion of the frontal is a shallow midline groove, visible near the midline, indicating the pathway for the olfactory tract (CitationBrochu et al., 2002). The ornamentation of the frontal is shallowly pitted, present at sutures, and oriented anteroposteriorly (). Two faint shallow grooves can be discerned extending just anterior to the supratemporal fenestral margin in an anteromedial direction, ceasing anteriorly about level with the posteromedial corners of the orbits.
Parietal—The interfenestral bar is slightly crushed at the contact between the frontal and parietal, with the posterior margin of the frontal being uplifted onto the anterior margin of the parietal (). The parietal portion of the interfenestral bar is equally as thick as that portion of the frontal, and comprises 72% of the length of the interfenestral bar. The interfenestral bar is roughly square in cross-section, with slight overhang on the dorsal surface. The thickness of the interfenestral bar is 16.71 mm at the closest point to the frontal that gradually thickens to 24.60 mm at its base at the posterior margin of the supratemporal fenestrae. The anteroventral portion of the parietal within the right supratemporal fenestra has been pushed up relative to the rest of the parietal, overlaying its otherwise smoothly dipping surface and obscuring its suture with the laterosphenoid (). The parietal overhangs anterodorsally onto the posterior margin of the supratemporal fenestra. The overhang originates at the parietal-squamosal suture and deepens toward the midline, where it merges with the very slight overhang of the interfenestral bar. This is less pronounced than the overhang in the anteromedial corners of the supratemporal fenestrae by the frontal bone. The parietal-quadrate suture is parallel with the skull roof, and is not visible in dorsal view. The parietal-squamosal suture extends laterally from the point of the posterior margin of the supratemporal fenestra, and passes through the location of the presumed temporal canal, slightly lateral to the mid-width of the supratemproal fenestra, and continues to extend ventrolaterally. The right parietal-squamosal suture is relatively straight, without a zigzag pattern, but the left suture is too distorted to be certain. The posterior portion of the parietal comprising the skull roof contacts the left and right squamosals and forms the posterior margin of the supratemporal fenestrae is broad, thick, and flat anteroposteriorly and transversally straight (). The parietal is dorsoventrally thin, slopes gently posteroventrally, and is slightly exposed in occipital view (). A small process projects ventrally, visible in posterior view, which has a raised midline and depression on either side such that it forms a thin ridge projecting ventrally from the skull roof (). The parietal-supraoccipital suture is crescentic, and not ‘W-shaped.’ The parietal is ornamented evenly across its dorsal surface, including across the suture with the frontal on the interfenestral bar ().
Postorbitals—The right postorbital has better preservation than the left and is essentially complete (). The postorbital constitutes the anterolateral margin of the supratemporal fenestra, and the posterior margin of the orbit (). The right postorbital has a strong anterolateral ventrally directed process that contacts the jugal and marks the posterolateral corner of the orbit. The postorbital-frontal contact occurs roughly at the mid-width of the anterior dorsal margin of the supratemporal fenestra. The posterior extensions of the postorbitals extend to about one-third the length of the supratemporal fenestrae from the posterior edge (). As the infratemporal fenestrae are incomplete, the amount of participation of the postorbitals can only be estimated, but is likely less than half of the dorsal margin (Figs. , ), and whether or not it contacts the quadratojugal is uncertain. The postorbital bars have been crushed beneath the frontal and postorbitals, and little can be discerned of their thickness or relative participations of the postorbital and jugal. The postorbitals have similar ornamentation to the rest of the dorsal surface of the skull and ornamentation is continuous across sutures ().
Squamosals—The squamosal-parietal contact occurs at the mid-width of the posterior margin of the supratemporal fenestra (). The squamosal-postorbital suture occurs roughly in the middle of the supraoccipital fenestrae. The squamosal narrows slightly anteriorly toward the suture with the postorbital, and is thick posteriorly, along the posttemporal bar (). The squamosal contacts the postorbital ventrally, participating in the posterodorsal margin of the infratemporal fenestra (). The lateral margin of the squamosal bears a faint groove that does not flare anteriorly (). The squamosal terminates posterodorsally in a short posteriorly directed process that is roughly level with the posterior edge of the occipital tuberosity and the base of the occipital condyle (). The squamosal does not extend ventrally in occipital view, and its visibility in this view is limited to the dorsal-most portion (). Ornamentation is pronounced near the parietal, but otherwise consistent across the dorsal surface of the squamosal ().
Jugals—The anterolateral portion of the jugal of UF/IGM 29 is all that remains of the bone on the left side, from its contact with the lacrimal to the anterior three-quarters of its participation in the infratemporal fenestrae (). Less is preserved of the right jugal, only from the presumed contact with the lacrimal to nearly the anterior half of the infratemporal fenestra (). The jugal of UF/IGM 29 only participates minimally in the posterolateral corner of the orbit. The point where the maxillary-jugal and maxillary-lacrimal sutures meet is at the posterolateral corner of the orbit (). The anteromost extent of the jugal is roughly level with the posterior one-third of the orbit. The lacrimal-jugal contact is slanted anteroventrally-posterodorsally. There is no evidence of foraminae along the jugals, thus it seems unlikely that UF/IGM 29 had a well-developed anterior jugal siphonial foramen (as discussed for Rhabdognathus in Brochu et al., 2002).
Supraoccipital—The supraoccipital is recessed between the occipital tuberosities and only slightly contributes to the medial portion of the tuberosities (). The supraoccipital is not V-shaped, but instead more crescentic, bowing inward toward the braincase. The supraoccipital is dorsally bordered by the parietal and medioventrally by the exoccipital. The exoccipital separates the supraoccipital from the foramen magnum. The supraoccipital does not contribute to the posttemporal fenestrae or contact the squamosals.
Exoccipitals—The exoccipitals are mostly preserved, yet compressed dorsoventrally (). The bones are present along both sides of the occipital condyle, but displaced on the right side, and too distorted to make accurate measurements. Exoccipital participation to the width of the occipital condyle may be estimated from the better-preserved left side to be about 36.8% dorsally and 26.3% ventrally. The exoccipital participates to the dorsal three-quarters of the foramen magnum. Dorsally, the exoccipital comprises the majority of the occipital tuberosity, which is well developed, rounded, and directed posteriorly (, ). The exoccipital extends laterally to constitute the ventral portion of the paraoccipital process. Much of the basioccipital tubera have been crushed and lost, and participation of the exoccipitals cannot be estimated (). The left foramen vagi (X–XI), that houses the vagus (X) and accessory nerves (XI) as well as the jugular vein (CitationBrochu et al., 2002) is preserved, as well as the right carotid foramen (). The foramen vagi is located along the lateral margin of the base of the ocipital condyle and is directed ventroposteriorly. The exoccipital forms the entire ventral surface of the posttemporal fenestra.
Basioccipital—The basioccipital forms the bulk of the occipital condyle and extends ventrally, forming the basioccipital tubera, which have mostly been lost in UF/IGM 29 (Figs. , ). The basioccipital tubera are oriented posterolaterally relative to the midline, forming a V-shape in ventral view (). The tubera are of indeterminate shape in occipital view (). There is a relatively wide, flat rugosity extending along the anteroventral midline of the basioccipital that is more ventral than the lateral edges. The ventral surface of the occipital condyle lacks any distinct grooves paralleling this rugosity (Figs. , ). Due to poor preservation in this region, it is difficult to determine how much the basioccipital extends ventrally from the occipital condyle. The area between the occipital condyle and the basioccipital tubera is arched dorsally, creating a smooth, concave outline in lateral view.
Quadrates—The quadrates have been crushed dorsoventrally such that they are flat and parallel to the skull roof. The right quadrate is very well preserved, reaching as far anteriorly as the level of the postorbital-squamosal contact, and lies immediately ventral to the squamosal throughout its length (). The quadrates are posteroventrally oriented and the cranioquadrate canals are not preserved (Figs. , ). The condylar portion of the left quadrate is lacking the lateral portion, leaving only the medial-most one-third. The quadrate condyle is better preserved in UF/IGM 31, which shows a partly eroded participation of the quadratojugal to the condyle's articulation with the lower jaw (). The quadrates lack a crest at mid-width, which does not seem to be an artifact of preservation ().
Quadratojugals—The left quadratojugal is preserved in UF/IGM 31 only (), and comprises the posterodorsal margin of the infratemporal fenestrae. The quadratojugal-quadrate suture is somewhat obscured, but includes the quadratojugal in the articulation with the lower jaw.
Palatines—The palatines of UF/IGM 29 have been flattened against the skull, and most of the posterior portions are missing, but clearly expand posterolaterally. The anterior portions form a sharp V between the maxillaries that reach anteriorly to the interalveolar space between the m6 and m7 alveoli (). The palatine forms the medial margin of the suborbital fenestra as well as its anteromedial corner.
Laterosphenoids—Little identifiable material remains of the laterosphenoids. A small portion of them is visible in dorsal view in the supratemporal fenestrae ventral and lateral to the parietal and frontal (). Much of the laterosphenoids in ventral view are obscured by the palatines crushed upon them (). A small anterior portion is exposed, in which the sutures of the parietal and frontal are parallel with the skull roof. The laterosphenoid does not contact the squamosal.
Basisphenoid—The ventral portion of the basisphenoid that includes the basisphenoid rostrum is missing (). The posterior branch of the medial eustachian foramen is visible in ventral view roughly 2 mm anterior to the ventral-most point of the basisphenoid that surrounds it entirely. The medial eustachian foramen has a diameter of 1.41 mm.
Mandible—A fairly complete mandible, UF/IGM 30 (), was recovered approximately 740 m from the site where UF/IGM 29 was discovered, within the same stratigraphic level. Based on its size, proportions, and number of teeth corresponding to the upper dentition, it is referred to Cerrejonisuchus improcerus gen. et sp. nov. UF/IGM 30 likely represents a second individual due to the distance between fossil localities and that the mandible is slightly smaller than would be expected for UF/IGM 29. The preservation of the mandible is very similar to that of UF/IGM 29; it has been dorsoventrally flattened toward the left lateral direction. Due to dorsoventral flattening, the height and width of the symphysis and splenials is too compressed for meaningful description or comparison.
FIGURE 8 Referred mandible of Cerrejonisuchus improcerus, UF/IGM 30, from the Cerrejón coal mine of northeastern Colombia, middle–late Paleocene, in dorsal view. A, UF/IGM 30 photograph; B, UF/IGM 30 sketch. Abbreviations: d, dentary; d1–13, dentary alveoli; dat, disassociated teeth; sp, splenial. Dotted lines represent features/sutures which were not clear. Scale bar equals 10 cm.
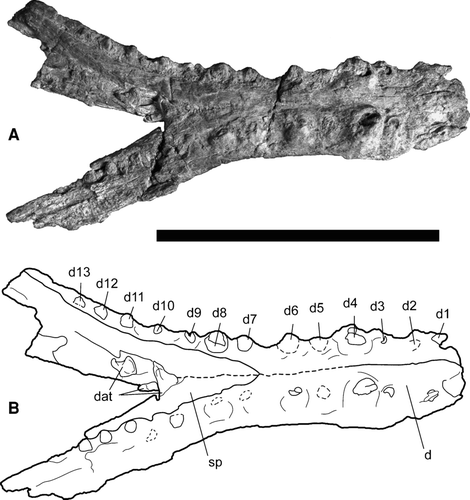
The specimen includes most of both dentaries and much of the splenials, but is missing the articular, angular, and surangular bones (). The d3–13 alveoli are present on the left dentary, and the d2–13 alveoli on the right dentary. The d1 alveoli on both dentaries are damaged; however, only a small part of the anteromost mandible is missing. The anteromost end of the right dentary suggests that the d1 alveolus for this tooth would have faced anterodorsally, not directly dorsal. The anterior portion of the mandible is not expanded, but rather continuous along most of the length of the symphysis (). The tooth of the d2 alveolus on the right dentary is flattened, but reduced relative to the space where the d1 alveolus would have existed. Measurements of the alveolar diameter and interalveolar spaces of UF/IGM 30 are compiled in . The d4 alveoli are the largest in the lower jaw and have dorsally higher alveolar walls. The dentition corresponds to the upper jaw of UF/IGM 29, such that the enlarged d4 teeth could occlude in the large interalveolar spaces between the p4 and m1 teeth. The large diastema between the d2 and d3 teeth would correspond well with the enlarged pm3 teeth. The m3 teeth would occlude well in the large interalveolar space between the d6 and d7 alveoli. The d7 alveoli are only slightly reduced relative to d8. The spreading angle of the dentaries measured at the labial midpoint of the left and right d13 alveoli to the anterior terminus of the splenials is approximately 40.7°. Ornamentation along the lateral and ventral surfaces is weak. The splenials fuse along the symphysis and taper anteriorly to a wedge between the two dentaries The splenials end anteriorly at the interalveolar spaces between d6 and d7 (). The splenials extend posteriorly beyond the last tooth at d13, as seen on the left dentary. The level at which the symphysis ends posteriorly is estimated to be level with the d9 teeth. The left d13 alveolus supports the final tooth in the lower dentition. The anterior alveoli are oriented dorsally and slightly anteriorly, grading to more dorsally directed alveoli and teeth toward the posterior end of the jaw.
TABLE 2 Alveolus dimensions and spacing for the upper dentition of the holotype, UF/IGM 29, and the lower dentition of the referred mandible, UF/IGM 30. Alveolus width measured transversally at anteroposterior midpoint. Alveolus length measured from anteromost to posteromost points along alveolar rim. Width between left and right alveoli measured from left medial anteroposterior midpoint to the right medial anteroposterior midpoint. Interalveolar length measured from posterior transverse midpoint to anterior transverse midpoint of adjacent alveolus. Dashes are used when preservation is not sufficient for measurement. All measurements in mm.
Dentition—Seven teeth were found in association with the maxilla, five of which were essentially preserved in place: the left and right m4, as well as the left m6–8 (). The premaxilla has preserved the right and left pm1 teeth, right and left pm3 teeth, and fragments of the right and left pm4. The posterior maxillary dentition is not preserved. UF/IGM 30 possesses an extremeley crushed and distorted right d2 tooth, very worn left d6–8 teeth, a misplaced tooth that may be referred to the right d9 alveolus, worn left d10–13, and worn right d11–12 (). The maxillary dentition of UF/IGM 29 is relatively homodont. The only significant deviations in size are the enlarged m3 alveoli and the reduced m4 alveoli (). Their general shape is conical and labiolingually compressed. The preserved teeth have a relatively rounded apex. The teeth are defined labially and lingually by strongly developed anterior and posterior carinae. Striae are not discernable on either surface of the teeth. Crenulations are present on at least the lingual side of the crown of the left m7 tooth, but preservation for the other teeth is too poor to assess this feature on the other maxillary teeth. The teeth do not increase or decrease in size posteriorly based on alveolar diameter, but without posterior teeth, it is difficult to be certain. The premaxillary tooth roots are more narrow and comparable to the apex width. In general the premaxillary teeth are thinner and longer than those of the maxillary. The maxillary tooth roots are wide relative to the crowns. The teeth do not curve distally. No preserved teeth possessed a ‘twisted’ carina.
Comparison
The elongate supratemporal fenestrae, posteriorly directed occipital tuberosities formed by the exoccipitals, strong participation of the exoccipitals to the occipital condyle, and the reduction of the seventh mandibular tooth and close placement to the eighth are major qualifying characters that unite Dyrosauridae (CitationJouve et al., 2006). The presence of all these characters in all preserved specimens indicates that Cerrejonisuchus improcerus belongs to the family Dyrosauridae.
Snout proportions vary among Dyrosauridae. UF/IGM 29 and 31 possess the shortest snout in proportion to the dorsal skull length of all known dyrosaurids (), at 59.30% and 54.78%, respectively. Chenanisuchus lateroculi (CitationJouve et al., 2005a) has the second shortest snout within Dyrosauridae, at 63.37%.
The incisive foramen is described for Dyrosaurus maghribensis as being heart-shaped and the posterior border as being comprised of the maxillae, as opposed to V-shaped and entirely surrounded by the premaxillae, as in C. improcerus (Figs. , ). The orbits of C. improcerus are placed much closer to the midline than those of Chenanisuchus lateroculi and the interorbital space is relatively small, more like the condition seen in D. phosphaticus. The orbits are more oval in shape than the circular shape of D. phosphaticus (CitationJouve, 2005) and Guarinisuchus munizi (CitationBarbosa et al., 2008). The supratemporal fenestrae of C. improcerus () are similar to all other known dyrosaurids in being much larger than the orbits (CitationBuffetaut, 1979) and longer than they are wide (CitationJouve et al., 2005b). The separation of the supratemporal fenestrae of C. improcerus is wide and thick, unlike the thin “sagittal crest” of Rhabdognathus aslerensis (CitationBrochu et al., 2002) and Guarinisuchus munizi (CitationBarbosa et al., 2008). The cross-section of the interfenestral bar is much more consistent with the square shape of Chenanisuchus lateroculi (CitationJouve et al., 2005a) than with the ‘T-shape’ of Dyrosaurus (CitationJouve, 2005; CitationJouve et al., 2006) and Atlantosuchus coupatezi (CitationJouve et al., 2008), or the smooth and narrow condition of Phosphatosaurus gavialoides (CitationBuffetaut, 1978; CitationMoody and Buffetaut, 1981), or the ‘V-shape’ of Rhabdognathus keiniensis (CitationJouve, 2007). The posterior walls of the supratemporal fenestrae are visible in dorsal view in C. improcerus as well as in Rhabdognathus (CitationJouve, 2007), but not in Dyrosaurus (CitationJouve, 2005). The lateral arches of the supratemporal fenestrae are robust, not thin as in Rhabdognathus (CitationJouve, 2007).
Cerrejonisuchus improcerus, like other dyrosaurids, possesses four premaxillary alveoli. The pm4 alveolus of UF/IGM 29 is greatly reduced, whereas in Dyrosaurus and C. bequaerti the pm1, pm2, and pm4 alveoli have similar diameters (CitationJouve and Schwarz, 2004; CitationJouve, 2005; CitationJouve et al., 2006). CitationJouve et al. (2005b) inferred that A. khouribgaensis also has a reduced pm4 tooth.
In dyrosaurids such as Phosphatosaurus gavialoides (CitationBuffetaut, 1978 and 1979) and Sokotosuchus ianwilsoni (CitationBuffetaut, 1979), the lateral margin of the snout is described as ‘festooned,’ referring to the undulations in the maxillae and premaxillae corresponding to alveoli and interalveolar spaces. This character is entirely absent in C. improcerus (); the margins are very smooth, as in Chenanisuchus lateroculi (CitationJouve et al., 2005a), Dyrosaurus phosphaticus (CitationJouve, 2005), Hyposaurus rogersii (CitationDenton et al., 1997), and Arambourgisuchus khouribgaensis (CitationJouve et al., 2005b). The total maxillary tooth count of C. improcerus is 11 teeth per maxilla. This number is more similar to estimates of 12 for Hyposaurus rogersii (CitationDenton et al., 1997), 13 for Chenanisuchus lateroculi (CitationJouve et al., 2005a), and 13–14 for Sokotosuchus ianwilsoni (CitationBuffetaut, 1979) and Guarinisuchus munizi (CitationBarbosa et al., 2008), than 15 for Phosphatosaurus gavialoides (CitationBuffetaut, 1978), 16 for Congosaurus bequaerti (CitationJouve and Schwarz, 2004), 17 for Arambourgisuchus khouribgaensis (CitationJouve et al., 2005b), 19–22 for Dyrosaurus (CitationJouve et al., 2006), or 23–24 of Rhabdognathus keiniensis (CitationJouve, 2007).
The nasals are entirely fused in UF/IGM 29 and 31, but unfused in the smaller UF/IGM 32 (). The two conditions for nasal fusing suggest the suture fuses with ontogeny in C. improcerus. The fused condition is consistent with other dyrosaurids such as Dyrosaurus phosphaticus (CitationJouve, 2005), Phosphatosaurus gavialoides (CitationBuffetaut, 1978, Citation1979), Hyposaurus rogersii (CitationDenton et al., 1997), and Congosaurus bequaerti (CitationJouve and Schwarz, 2004). Rhabdognathus aslerensis (CitationBrochu et al., 2002), Atlantosuchus coupatezi (CitationJouve et al., 2008), and Sokotosuchus ianwilsoni (CitationBuffetaut, 1979) have entirely unfused nasal bones, whereas Chenanisuchus lateroculi has an anteriorly fused nasal bone. The anterior portion of the preserved Rhabdognathus keiniensis is fused; however, the posterior portion is unpreserved (CitationJouve, 2007).
Posteromedially, the frontal participates in the thick interfenestral bar, contrasting the thin bar of A. khouribgaensis (CitationJouve et al., 2005b). The frontal participates 28.5% to the interfenestral bar in UF/IGM 29, as opposed to 20% in A. khouribgaensis (CitationJouve et al., 2005b) and Atlantosuchus coupatezi (CitationJouve et al., 2008), 25% of D. phosphaticus (CitationJouve, 2005) and Rhabdognathus keiniensis (CitationJouve, 2007), around 29% for Phosphatosaurus gavialoides (CitationBuffetaut, 1978), 33% of Chenanisuchus lateroculi (CitationJouve et al., 2005a) and Rhabdognathus aslerensis (CitationBrochu et al., 2002), and roughly 50% in Hyposaurus rogersii (CitationTroxell, 1925). UF/IGM 29 has a wide interfenestral bar, 16.71 mm at its minimum width, very similar to that of C. lateroculi with a value of 17 mm at its minimum width (CitationJouve et al., 2005a).
The frontoparietal suture of C. improcerus is much straighter () than the zigzag pattern described for D. phosphaticus (CitationJouve, 2005). C. improcerus lacks the V-shaped groove on the dorsal surface (see ) as in Rhabdognathus aslerensis (CitationBrochu et al., 2002; CitationJouve, 2007), and also lacks the deep sulcus and dorsal bulges at the posterior end when in occipital view (). C. improcerus also lacks the strong indentation of the posterior margin (see ) of the parietal as in Rhabdognathus (CitationJouve, 2007), Hyposaurus rogersii (CitationDenton et al., 1997), Atlantosuchus coupatezi (CitationJouve et al., 2008), and Guarinisuchus munizi (CitationBarbosa et al., 2008). C. improcerus does not have the W-shaped parietal-supraoccipital suture, seen in occipital view of D. phosphaticus (CitationJouve, 2005), making it more similar to the genus Rhabdognathus (CitationJouve, 2007), which also lacks this character.
The supraoccipital of C. improcerus does not display a V-shape () as in Dyrosaurus (CitationJouve, 2005; CitationJouve et al., 2006), Chenanisuchus lateroculi (CitationJouve et al., 2005a), and Arambourgisuchus khouribgaensis (CitationJouve et al., 2005b).
The occipital tuberosities are mostly comprised of the exoccipitals, but medially of the supraoccipital in C. improcerus (), and are posteriorly directed as in all dyrosaurids (CitationJouve et al., 2006). The occipital tuberosities of C. improcerus seem at least as well developed as the ‘moderately developed’ state of D. phosphaticus (CitationJouve, 2005); they are much more developed than in S. ianwilsoni and C. lateroculi (CitationJouve et al., 2005a), and most similar to the length and shape of R. keiniensis and R. aslerensis (CitationJouve, 2007).
The lateral exoccipital forms the ventral portion of the paraoccipital process of C. improcerus (Figs. , ), as in Dyrosaurus phosphaticus (CitationJouve, 2005) and Dyrosaurus maghribensis (CitationJouve et al., 2006) and possibly other dyrosaurids as well. The ventral exoccipital contributes one-third to the basioccipital tuber in D. phosphaticus (CitationJouve, 2005), and less than one-fourth in Chenanisuchus lateroculi (CitationJouve et al., 2005a). The participation to the basiocciptial tuber of the better-preserved left exoccipital of UF/IGM 29 is 26%, more consistent with the condition of C. lateroculi (CitationJouve et al., 2005a). Similar participation to the basioccipital tubera is also noted in Rhabdognathus aslerensis (CitationBrochu et al., 2002). Both C. improcerus and D. phosphaticus (CitationJouve, 2005) exhibit an arched interspace between the occipital tuberosity and the basioccipital tubera.
No crest is visible on the ventral face of the quadrate of C. improcerus, which does not seem to be an artifact of preservation (). The lack of crest is dissimilar to A. khouribgaensis(CitationJouve et al., 2005b), Rhabdognathus aslerensis (CitationBrochu et al., 2002), and Rhabdognathus keiniensis (CitationJouve, 2007), which all possess a crest on the ventral side of the quadrate.
Dyrosaurus phosphaticus, like C. improcerus (), has a concavity at the middle of the medial margin of the suborbital fenestra, as a result of a restriction along the mid-length of the palatine (CitationJouve, 2005).
The anterior dentition of the dentary of C. improcerus is similar to that of Arambourgisuchus khouribgaensis (CitationJouve et al., 2005b), Hyposarus rogersii (CitationTroxell, 1925), and Dyrosaurus (CitationJouve, 2005; CitationJouve et al., 2006) in that the d2 alveoli are reduced in size and the interalveolar spaces between the d2 and d3 are greater than between d1 and d2, as well as d3 being placed very close to an enlarged d4 (). Arambourgisuchus khouribgaensis also has the raised d4 alveolar wall found in C. improcerus (CitationJouve et al., 2005b). The anterior orientation of the d1 alveolus of C. improcerus is similar to that of Hyposaurus derbianus (CitationCope, 1886) and unlike that of Dyrosaurus phosphaticus, which has a more dorsal orientation. The anterior portion of the C. improcerus mandible is not expanded, and thus not spatulate as in some Hyposaurus (CitationJouve, 2007), but more similar to H. derbianus (CitationCope, 1886).
The mandibular symphysis ends posteriorly in UF/IGM 30 at approximately the level of the d9 alveolus (), which is dissimilar to the d12 of Hyposaurus rogersii reported by CitationJouve (2007). This condition of the mandibular symphysis contrasts more strongly with that of other dyrosaurids, which have longer mandibles, such as the d14 alveolus in Guarinisuchus munizi (CitationBarbosa et al., 2008), the d15 in Hyposaurus sp. (CitationJouve, 2007), and Hyposaurus derbianus (CitationCope, 1886), the d16 in Congosaurus bequaerti (CitationJouve and Schwarz, 2004) and Arambourgisuchus khouribgaensis (CitationJouve et al., 2005b), the d17 in Dyrosaurus (CitationJouve, 2005), and beyond the d19 alveolus in Rhabdognathus (CitationJouve, 2007).
The splenials end anteriorly at the interalveolar space between the d6 and d7 alveoli in C. improcerus (), similar to that of Hyposaurus rogersii for which they end at the d7 alveolus (CitationJouve, 2007). This splenial-alveolus relationship contrasts that of other dyrosaurids with more posterior positions such as the interalveolar space between the d9 and d10 alveoli in Congosaurus bequaerti (CitationJouve and Schwarz, 2004) and Phosphatosaurus gavialoides (CitationBuffetaut, 1978), the interalveolar space between d10 and d11 in Arambourgisuchus khouribgaensis (CitationJouve et al., 2005b) and Hyposaurus derbianus (CitationCope, 1886), and d12 in Dyrosaurus (CitationBuffetaut, 1978).
The overall dentary tooth estimation for H. rogersii is as high as 17–20 (CitationBuffetaut, 1980) but as low as 15 (CitationDenton et al., 1997), somewhat comparable to the 13 of C. improcerus. The size of the d7 alveolus is much more comparable to the d8, contrary to the very reduced d7 alveolus relative to the d8 character traditionally used as a synapomorphy of Dyrosauridae. C. improcerus lacks the ‘festooned’ appearance () of H. rogersii along the lateral margins of the dentaries. Due to the flattening of UF/IGM 30, the diagnostic characters of width as compared to height of the lower jaw cannot be discerned.
The apices of the crowns of the preserved teeth of UF/IGM 29 are more rounded than acute, as in C. bequaerti (CitationJouve and Schwarz, 2004). The teeth of C. improcerus possess well-developed anterior and posterior carinae as in C. bequaerti (CitationJouve and Schwarz, 2004), D. phosphaticus (CitationJouve, 2005), D. maghribensis (CitationJouve et al., 2006), and H. derbianus (CitationCope, 1886), defining the labial and lingual surfaces. Unlike Congosaurus bequaerti (CitationJouve and Schwarz, 2004), Dyrosaurus (CitationJouve, 2005; CitationJouve et al., 2006), and Atlantosuchus coupatezi (CitationJouve et al., 2008), C. improcerus lacks noticeable striations on either face of the teeth, but instead possesses apical lingual crenulations on the left m7 tooth. Chenanisuchus lateroculi was noted as having lacked striations on at least the labial surfaces (CitationJouve et al., 2005a), as in C. improcerus. Hyposaurus rogersii has striations on the lingual surface (CitationDenton et al., 1997) and Hyposaurus derbianus possesses longitudinal ridges on the lingual face and a smooth condition on the exterior of its mandibular teeth (CitationCope, 1886). Arambourgisuchus khouribgaensis (CitationJouve et al., 2005b) was described as having a somewhat intermediate form of variably present and either absent or weak superficial striae. The teeth of C. improcerus do not curve distally, as in Dyrosaurus maghribensis (CitationJouve et al., 2006), Atlantosuchus coupatezi (CitationJouve et al., 2008), Guarinisuchus munizi (CitationBarbosa et al., 2008), and Phosphatosaurus gavialoides (CitationBuffetaut, 1978), nor are they slightly recurved as in Sokotosuchus ianwilsoni (CitationBuffetaut, 1979) and H. derbianus (CitationCope, 1886). The ‘twisted’ carinae characteristic of H. rogersii (CitationDenton et al., 1997) was not noticed in any of the preserved teeth.
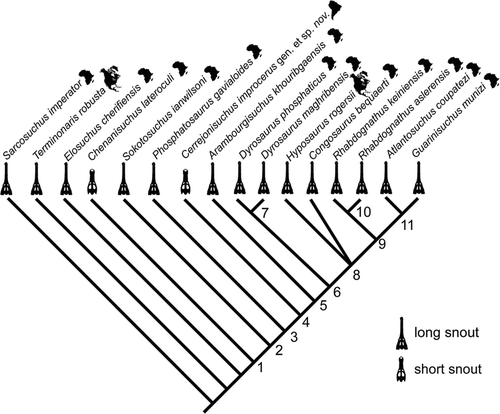
FIGURE 9 Results from phylogenetic analysis of 82 characters and 16 taxa (see Appendices 1 and 2). Single most-parsimonious cladogram of branch and bound search using both ordered and unordered characters. Tree lengths: 170 each; CI: 0.547; RI: 0.642; RC: 0.351; H.I.: 0.453. Unambiguous synapomorphies, with a superscript R indicating instances of reversal: Node 1, 1(1), 3(1), 6(1), 11(1), 15(1), 19(1), 27(1), 50(1), 52(1), 55(1), 60(1), 75(1); Node 2, 7(1), 8(1), 74(1); Node 3, 10(1), 32(2); Node 4, 31(1), 80(1); Node 5, 9(2), 22(1), 33(0)R; Node 7, 8(0)R, 9(1), 63(1); Node 8, 42(1), 50(2), 59(1), 81(1), 82(1); Node 9, 46(1), 48(1); Node 10, 39(1); Node 11, 5(0)R.
PHYLOGENETIC RELATIONSHIPS
Five phylogenetic analyses at the genus level of the family Dyrosauridae have been published previously (CitationBuffetaut, 1978; CitationJouve, 2005; CitationJouve et al., 2005b; CitationBarbosa et al., 2008; CitationJouve et al., 2008). CitationBuffetaut (1978) included Phosphatosaurus, Rhabdognathus, Dyrosaurus, and Hyposaurus in a manual cladogram. The most basal of which was Phosphatosaurus, with Hyposaurus and Dyrosaurus being more derived, and Rhabdognathus falling in between. CitationJouve (2005) conducted an analysis of 12 characters across five genera of Dyrosauridae. This analysis was expanded by CitationJouve et al. (2005b) to 30 characters and nine members of Dyrosauridae. CitationJouve et al. (2008) further expanded this analysis to 43 characters and 11 species of Dyrosauridae. These studies found Sokotosuchus and Phosphatosaurus to be basal genera of the family, supporting the subfamily Phosphatosaurinae, which at the time it was named included only these two genera (CitationBuffetaut, 1980). When Chenanisuchus was included, it was basal to all other members of Dyrosauridae (CitationJouve et al., 2005b; CitationJouve, 2008). CitationBarbosa et al. (2008) modified the 30 characters of CitationJouve et al. (2005b) and added their new taxon, Guarinisuchus munizi. This study also found a basal placement of Chenanisuchus, Sokotosuchus, and Phosphatosaurus, and placed Guarinisuchus as sister to a clade uniting Arambourgisuchus and Rhabdognathus.
New cranial specimens of Cerrejonisuchus improcerus allow for a reevaluation of the phylogenetic affinities of South American dyrosaurids. To help resolve the relationship of C. improcerus to other dyrosaurids, we performed a cladistic analysis of 82 cranial and mandibular characters (and one postcranial) for 13 dyrosaurid species and three outgroup taxa (Appendices 1 and 2). Early Cretaceous Sarcosuchus imperator (CitationSereno et al., 2001) and Elosuchus cherifiensis (de Lapparent de Broin, 2002) from Africa and Late Cretaceous Terminonaris robusta (CitationWu et al., 2001) from North America were used as outgroup taxa. The cladistic analysis was rooted with Sarcosuchus imperator because it is the geologically oldest mesoeucrocodylian thought to be closely related to dyrosaurids but clearly not in the ingroup (CitationSereno et al., 2001; CitationJouve et al., 2005b), and because its cranium has been thoroughly described (CitationSereno et al., 2001). We included only members of Dyrosauridae known from at least somewhat complete cranial material in this analysis. The known dyrosaurid species that were not included due to lack of cranial material are the Cretaceous- or Paleocene-aged Hyposaurus derbianus of Brazil (CitationCope, 1885, Citation1886), the Paleocene dyrosaurids Hyposaurus paucidens from Morocco (CitationArambourg, 1952; CitationJouve, 2007) and Congosaurus compressus from Mali (CitationJouve, 2007), and the Eocene dyrosaurid Rhabdognathus acutirostris from Tunisia (CitationBuffetaut, 1978). All characters except 26, 29, 30, 32, and 53 (continuous, multistate characters; e.g., location of the posteromost alveolus relative to the orbit/supratemporal fenestrae: character 30) were treated as unordered. Morphologic data were compiled from the literature (CitationTroxell, 1925; CitationBuffetaut, 1978, Citation1979; CitationBuffetaut and Wouters, 1979; CitationDenton et al., 1997; CitationSereno et al., 2001; CitationWu et al., 2001; CitationBrochu et al., 2002; de Lapparent de Broin, 2002; CitationJouve and Schwarz, 2004; CitationJouve, 2005, Citation2007; CitationJouve et al., 2005a, Citation2005b, Citation2006, Citation2008; CitationBarbosa et al., 2008) and study of specimens. Although postcranial data are known for Dyrosauridae (CitationJouve et al., 2006; CitationSchwarz et al., 2006), for the most part they have yet to be studied sufficiently for enough species to make them useful in this phylogenetic analysis. Three characters were variable within species (character 5, lateral margin of orbit; character 23, shape of maxillary margin; character 32, fusion of the nasals) and were coded as polymorphic. Characters not known for a taxon were coded as missing.
Data were compiled in Mesquite version 2.5 (CitationMaddison and Maddison, 2008) and the cladistic analysis was done in PAUP version 4.0b10 (CitationSwofford, 2003). A branch and bound search resulted in two most-parsimonious cladograms (strict consensus presented in ) with tree lengths of 170 steps, a consistency index of 0.547, a retention index of 0.640, and a homoplasy index of 0.453.
Our analysis corroborated the position of Chenanisuchus lateroculi at the base of Dyrosauridae, followed by Sokotosuchus ianwilsoni and Phosphatosaurus gavialoides. Cerrejonisuchus improcerus gen. et sp. nov., placed relatively basally between P. gavialoides and Arambourgisuchus khouribgaensis. C. lateroculi did not share a special relationship with the similarly short-snouted C. improcerus, as might have been expected based on snout proportions alone (Figs. , ). Both species of Dyrosaurus and both species of Rhabdognathus paired in separate monophyletic clades, and a polytomy occurred among Hyposaurus rogersii, Congosaurus bequaerti, and the clade uniting Rhabdognathus + Atlantosuchus coupatezi + Guarinisuchus munizi.
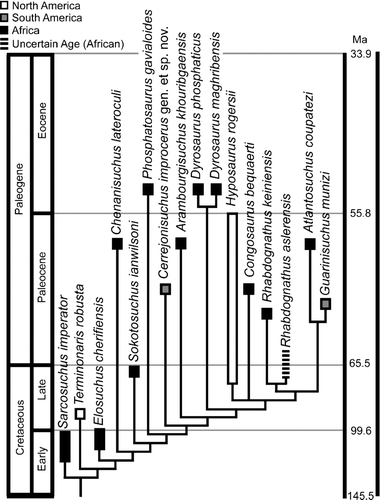
FIGURE 10 Phylogenetic relationship placed in stratigraphic and paleobiogeographic context. Dates from Gradstein et al., 2004.
DISCUSSION
Results from our study agree with previous placement of Chenanisuchus lateroculi, Sokotosuchus ianwilsoni, and Phosphatosaurus gavialoides as basal members of Dyrosauridae (CitationJouve, 2005; CitationJouve et al., 2005b, Citation2008; CitationBarbosa et al., 2008). The consensus cladogram presented here (Figs. , ) is similar to that of CitationBarbosa et al. (2008), only differing in a more basal position of Arambourgisuchus khouribgaensis relative to Hyposaurus rogersii and that instead corroborates the results of CitationJouve et al. (2008). The polytomy at node 8 may be due to the relatively little cranial material known for Congosaurus bequaerti (known from only a skull missing all bones posterior to the orbits; coding for this taxon limited to 41% of the characters in the matrix), this placement is tenuous and could change with discovery of more complete skulls. When C. bequaerti is removed, C. lateroculi still places as most basal, but the next node is a large polytomy containing S. ianwilsoni, P. gavialoides, Cerrejonisuchus improcerus, H. rogersii, and the clade uniting A. khouribgaensis, Dyrosaurus, Rhabdognathus, Atlantosuchus coupatezi, and Guarinisuchus munizi. When all characters were treated as unordered, the analysis resulted in four equally most-parsimonious cladograms reflecting four alternate relationships among H. rogersii, the two Rhabdognathus species, A. coupatezi, G. munizi, and C. bequaerti. However, the relationship of Cerrejonisuchus was not affected.
As for previous analyses, our results support the monophyly of Rhabdognathus (CitationJouve et al., 2005b; CitationBarbosa et al., 2008; CitationJouve et al., 2008) united by frontal participation to the interfenestral bar of equal to or greater than one-third (character 38). The Dyrosaurus clade is supported by three synapomorphies: a reversal to an ornamented interfenestral bar (character 8), T-shaped interfenestral bar (character 9), and a posteriorly descending parietal-laterosphenoid suture (character 63).
Despite the lack of sufficient cranial material, species not included in the phylogenetic analysis have biogeographic relevance to the current study (). If Hyposaurus derbianus is at least considered a valid member of the genus Hyposaurus, its presence in Brazil (CitationCope, 1886) implies a radiation of the genus Hyposaurus into the New World during the Late Cretaceous. Furthermore, the presence of Hyposaurus paucidens in Morocco, as well as other indeterminate Hyposaurus material (CitationJouve, 2007), implies a form of Hyposaurus remained in Africa into the Paleocene. Neither Congosaurus compressus nor Rhabdognathus acutirostris would imply drastically different paleobiogeography because both genera are already known from Africa (CitationJouve, 2007).
The monophlyly of Dyrosauridae is supported by 12 unambiguous synapomorphies in the current study. Presence of all basal dyrosaurid taxa, together with two of three outgroups, in Africa strongly supports previous paleobiogeographic hypotheses for an African origin of Dyrosauridae (CitationJouve et al., 2005b, Citation2008).
South American dyrosaurids are now known from the Paleocene of Bolivia, the Early Paleocene of Brazil, and the middle or late Paleocene of northeastern Colombia, as well as possibly from the Late Cretaceous in Bolivia and Brazil. The most complete material prior to this study is from the Pernambuco region of Brazil (CitationCope, 1885, Citation1886; CitationBarbosa et al., 2008), Hyposaurus derbianus and G. munizi. The genus Hyposaurus is best known from Late Cretaceous and Paleocene deposits of eastern North America, all referred to the species Hyposaurus rogersii (CitationDenton et al., 1997). Hyposaurus is also known from much more fragmentary remains from the Paleogene of Africa (CitationJouve, 2007).
Considering the temporal placement of the New World taxa (see ), we suggest three independent dispersals from Africa to the New World. Presence of C. improcerus together with a number of undescribed taxa from the Paleocene of northeastern Colombia suggests that there was a radiation of dyrosaurid crocodyliforms, possibly following the K-P boundary, in tropical South America. CitationBarbosa et al. (2008) propose a dispersal from Africa to Brazil with continued immigration into North America. Our finding supports this dispersal route and adds to it evidence of coastal dyrosaurids in northern South America in Colombia, transitional between the Brazilian occurrence and North American occurrences. Also, presuming these crocodilians dispersed along coasts, this would also give a transitional location for the occurrence of dyrosaurids in South America between the Brazilian specimens and the Bolivian specimens.
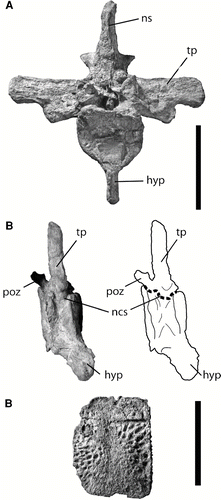
FIGURE 11 Associated postcrania of UF/IGM 31. A–B, anterior dorsal vertebra (likely the third or fourth position); A, in cranial view; B, in right oblique view of ventral aspect showing fusion of neurocentral suture; C, lateral osteoderm. Abbreviations: hyp, hypapophysis; ncs, neurocentral suture; ns, neural spine; poz, postzygapophysis; tp, transverse process. Scale bar equals 5 cm.
In addition, C. improcerus has the shortest snout known within Dyrosauridae (). The next shortest snout within the family belongs to C. lateroculi from the Paleocene of Morocco (CitationJouve et al., 2005a). Separation of C. improcerus and C. lateroculi within the phylogenetic analysis (Figs. , ) suggests that the short-snouted condition evolved at least twice within Dyrosauridae. The short-snouted condition most likely evolved to adapt to a more generalized diet, still including fish but invertebrates and small vertebrates as well, as opposed to the presumed strongly piscivorous diet of the typical long-snouted dyrosaurids (CitationBuffetaut, 1979; CitationDenton et al., 1997). The morphological similarities between the two short-snouted species are fairly limited beyond their snout proportions. The interfenestral bar for both species is relatively wide and square-shaped in cross-section. In addition, the posterior margin is relatively transversally straight in both short-snouted species. In long-snouted species, the margin can be either straight (e.g., Dyrosaurus) or indented (e.g., Rhabdognathus). The most noteworthy differences are their relative orbital placement and development of the occipital tuberosities.
Regarding ontogeny, all cranial sutures seem to be fully fused with the only possible exception being the nasal of UF/IGM 32. The partially fused nasal of UF/IGM 32 indicates this individual was slightly less mature than UF/IGM 29 or 31, particularly considering its smaller size (). Moreover, the associated postcrania of UF/IGM 31 further indicate morphologically mature status (). In crocodilians, the neurocentral sutures close from tail to neck during ontogeny (CitationBrochu, 1996), implying that the closed neurocentral sutures of the anterior dorsal vertebrae of C. improcerus, UF/IGM 31, indicate a morphologically mature individual (). Furthermore, no isolated centra were associated with this partial skeleton. Further evidence of maturity comes from the preserved osteoderms. Osteoderms in Alligator begin calcification around 1 year into their growth period as a small round nucleus, which then grows during life to articulate with other osteoderms and provide the dermal shield (CitationVickaryous and Hall, 2008). Assuming similar developmental mechanisms governed dyrosaurid osteoderm growth, these osteoderms likely represent an individual that has reached morphological maturity. Due to the similar size between the holotype (UF/IGM 29) and the referred UF/IGM 31 and fully fused cranial sutures, these both likely represent morphologically mature specimens, and the characters that distinguish C. improcerus from other dyrosaurid species likely reflect unique taxonomic features, as opposed to the features of younger stages in a known dyrosaurid species. Thus, several features, including snout proportion and supratemproal fenestrae shape and orientation, are considered to be morphologically distinct characters from other members of Dyrosauridae. Until further specimens are discovered, we set the estimated upper size limit of the species based on the largest specimen (2.22 m; UF/IGM 31; see ), which is similar to, but still smaller than, the lower size limit of 2.48 m for next smallest dyrosaurid, Hyposaurus rogersii.
Following CitationSereno et al. (2001), the body size of C. improcerus was approximated based on dorsal skull length of the morphologically mature specimens UF/IGM 29 and 31 (UF/IGM 32 is too incomplete for an accurate estimate, and likely morphologically immature). Using the known relationship between these measurements for Gavialis gangeticus and Crocodylus porosus, we estimate the total body length of morphologically mature C. improcerus to have been between 1.22 and 2.22 m (). As compared to other estimates of dyrosaurid length by CitationJouve et al. (2005a), C. improcerus has the shortest total body length in the family (). The longest dyrosaurid from this study was Phosphatosaurus gavialoides, at 7.22–8.05 m. Therefore the new body length range for mature dyrosaurids is between 1.22 and 8.05 m.
APPENDIX 1
Description of characters used in phylogenetic analyses. Outgroup consists of Sarcosuchus imperator, Terminonaris robusta, and Elosuchus cherifiensis. Ordered characters are labeled as such; all other characters are unordered. Except where otherwise stated, characters are original to this publication. Characters that have been modified from other publications are cited with the character number in bold after the citation.
General
-
Length of snout: greater than 68% (0); lesser than 68% (1) of total skull length, measured from tip of snout to posterior margin of parietal. Modified from Jouve, 2005 5.
Cranial Openings
-
Medial tubercles of external nares on the posterior margin: absent (0); dorsal (1); ventral (2).
-
Thickness of anterior margin of external nares: less than half anteroposterior length (0); greater than half anteroposterior length (1).
-
Orbits positioned: medially (0); laterally (1).
-
Lateral margin of the orbit comprised dominantly by: lacrimal (0); jugal (1).
-
Supratemporal fenestrae length and width: subequal or wider than long (0); longer than wide (1). Modified from: CitationClark, 1994 68; CitationOrtega et al., 2000 41; CitationWu et al., 2001 67; Sereno et al., 2003 4; CitationJouve et al., 2005b 15; CitationJouve et al., 2006 12; CitationJouve et al., 2008 15.
-
Interfenestral bar: wide (0); narrow (1). Modified from: CitationClark, 1994 33; CitationWu et al., 2001 33; CitationSereno et al., 2001 26; CitationJouve, 2005 10; CitationJouve et al., 2005b 18; CitationJouve et al., 2006 90; CitationJouve et al., 2008 18.
-
Interfenestral bar: ornamented (0); unornamented (1). Modified from: CitationJouve et al., 2005b 30; CitationJouve et al., 2008 30.
-
Cross section of interfenestral bar: square-shaped (0); shaped as an upright T (1); upside-down V-shaped, in the form of a sagittal crest (2). Following of Jouve et al., 2005a.
-
Posterior walls of supratemporal fenestrae: almost vertical and barely visible in dorsal view (0); dorsally inclined and largely visible in dorsal view (1). Modified from: CitationJouve et al., 2005b 10; CitationJouve, 2005 6; CitationJouve et al., 2008 10.
-
Lateral arches of supratemporal fenestrae: thick (0); thin (1). Modified from: CitationJouve et al., 2005b 16; CitationJouve et al., 2008 16.
-
Suborbital fenestrae anteriorly reach: posterior to the anterior 2/3 of the maxillary alveoli count (0); anterior/proximal to the anterior 2/3 (1).
Premaxillae
-
Posterodorsal processes of premaxillae reach: proximal or distal to the third maxillary alveolus (0); proximal or anterior to second alveolus (1).
-
Lateral expansion of the premaxillae anterior to premaxillary-maxillary suture: present (0); absent (1).
-
Premaxillae: possess 5 or more alveoli (0); possess 4 alveoli (1). Modified from: Gasparini et al., 1991 5; CitationOrtega et al., 2000 133; CitationJouve, 2005 12; CitationJouve et al., 2005b 26; CitationJouve et al., 2006 182; CitationJouve et al., 2008 26.
-
Third premaxillary alveoli: not enlarged relative to both second and fourth premaxillary alveoli (0); third alveoli enlarged relative to both adjacent alveoli (1).
-
Fourth premaxillary alveolus: enlarged or similar in size to first and second alveoli (0); reduced relative to all other premaxillary alveoli (1).
-
First notch for mandibular occlusion: similar in size to adjacent occlusal spacing (0); significantly larger than adjacent spacing (1).
-
Left and right first premaxillary alveoli: similar or wider interalveolar space than adjacent spacing (0); first premaxillary teeth closely spaced (1).
-
Last premaxillary alveoli position: anterolateral to first maxillary tooth (0); anterior to first maxillary tooth (1). Modified from: CitationSereno et al., 2001 70; CitationJouve et al., 2006 193.
-
Anteroventral expansion of premaxillae: present (0); absent (1).
-
Premaxillary length as compared to width: slightly longer than wide (0); nearly three times longer than wide or more than three times longer than wide (1). Modified from CitationJouve et al., 2008 41.
Maxillae
-
Margins of maxillae: not linear (0); linear for anterior 2/3 or less (1); linear for over the anterior 2/3 (2).
-
Lateral expansion of maxillae: absent, lateral margins linear (0); present, lateral margins ‘festooned’ (1). Modified from CitationJouve et al., 2008 40.
-
Alveolar walls: raised relative to ventral surface of the maxillae in lateral view (0); walls level with this surface (1).
-
Each maxilla possesses: 18 or greater alveoli (0); 15–17 alveoli (1); 10–14 alveoli (2). Modified from: Gasparini et al., 1991 6; CitationOrtega et al., 2000 164; CitationSereno et al., 2001 51; CitationJouve et al., 2006 183; Ordered.
-
Maxillary alveoli posterior to anterior margin of orbits: absent (0); present (1).
-
Posteromost contact between maxillae and palatines on ventral surface relative to posteromost maxillary alveoli: posterior (0); anterior (1).
-
Maxillae extend posteriorly to: anterior or proximal to level of postorbital bars (0); even with the anteroposterior midlength of supratemporal fenestrae (1); proximal to the level of the posterior margins of the supratemporal fenestrae (2); Ordered.
-
Posteromost alveolus: proximal to level of anterior margin of orbit (0); posterior margin of orbit (1); proximal/posterior to anteroposterior midlength of supratemporal fenestrae (2); Ordered.
-
Maxillary tooth size: variable (0); homodont (1). Modified from: CitationJouve et al., 2005b 25; CitationJouve et al., 2008 25.
Nasals
-
Nasal in adult form: unfused (0); fused anteriorly (1); entirely fused (2); Ordered.
-
Anteromost extension of the nasal relative to level of first maxillary tooth: posterior (0); anterior (1). Modified from CitationJouve et al., 2008 42.
Lacrimal
-
Lacrimal extends anteriorly: only to posterior 1/3 maxillary alveoli (0); to anterior 2/3 alveoli (1).
-
Lacrimal-nasal contact relative to prefrontal-nasal contact in adult form: about equal (0); about twice as long (1). Modified from: CitationJouve et al., 2005b 12; CitationJouve et al., 2008 12.
-
Posteromost lateral contact of the maxillary and lacrimal bones as seen in dorsal view: proximal to posterolateral corner of orbits (0); proximal to anterolateral corner of orbits (1).
Frontal
-
Interorbital space, including prefrontals if applicable: narrow, less wide than minimal width of snout, at level of premaxillary-maxillary suture (0); broad, wider than minimal width of snout (1). Modified from: CitationClark, 1994 20; CitationWu et al., 2001 20; CitationJouve et al., 2005b 22; CitationJouve et al., 2006 83; CitationJouve et al., 2008 22.
-
Anterior frontal extension position: posteriorly located, level with or slightly anterior to the prefrontal (0); very anterior to the prefrontal (1). Modified from CitationJouve et al., 2008 31.
-
Frontal contribution to interfenestral bar: less than one-third (0); greater than or equal to one-third (1). Modified from: CitationClark, 1994 23; CitationSereno et al., 2001 20.
Parietal
-
Frontoparietal suture on the interfenestral bar: straight/smooth (0); zigzag (1).
-
Parietal-quadrate suture relative to the skull table: vertical (0); parallel (1).
-
Posterodorsal margin of parietal: transversely oriented (0); indented anteriorly (1). Modified from: CitationJouve, 2005 7; CitationJouve et al., 2005b 11; CitationJouve et al., 2008 11.
-
Medial parietal crest, visible in occipital view: present (0); absent (1).
-
Parietal in occipital view: ‘W-shaped’ (0); concave (1). Modified from CitationJouve et al., 2008 32.
-
Dorsal portion of parietal: strongly ornamented with deep pits (0); smooth (1); slightly ornamented (2). Modified from: CitationJouve et al., 2005b 27; CitationJouve et al., 2008 27.
-
Posterior lamina of posterior margin of skull roof above posttemporal fenestrae and the occipital tuberosities which overhangs, hiding part of the occipital tuberosity from dorsal view: absent (0); present (1). Modified from CitationJouve et al., 2008 34.
Postorbital
-
Antero-lateral process: absent/small (0); pronounced and reaches dorsal margin of jugal (1). Modified from: CitationClark, 1994 28; CitationOrtega et al., 2000 32; CitationWu et al., 2001 28; CitationSereno et al., 2001 24; CitationJouve, 2005 4; CitationJouve et al., 2005b 9; CitationJouve et al., 2006 58; CitationJouve et al., 2008 9.
Squamosal
-
Posterior margin of squamosal lateral to occipital tuberosity: straight (0); anteriorly concave (1). From: CitationJouve et al., 2005b 29; CitationJouve et al., 2008, 29.
Jugal
-
Ventral portion of postorbital bar: medially displaced with ridge separating postorbital bar from lateral surface of jugal (0); medially displaced from lateral surface of jugal with no ridge (1); nearly flush with lateral surface of jugal (2). Modified from: CitationOrtega et al., 2000 34; CitationJouve et al., 2006 72.
-
Postorbital bar: nearly vertical (0); slightly anterodorsally inclined (1); strongly anterodorsally inclined (2). From CitationJouve et al., 2008 35.
Supraoccipital
-
Supraoccipital participation to posttemporal fenestrae: present (0); absent (1).
Exoccipitals
-
Exoccipital participation to occipital condyle: slight (0); large (1). Modified from: Jouve, 2004 96; CitationJouve et al., 2005b 5; CitationJouve et al., 2006 104; CitationJouve et al., 2008 5.
-
Occipital tuberosities: absent (0); small and reduced (1); strongly developed (2); Ordered. Modified from: CitationJouve, 2005 1; CitationJouve et al., 2005b 3; CitationJouve et al., 2008 3.
Basisphenoid
-
Basisphenoid rostrum: short (0); extremely long anteriorly (1). From: CitationJouve, 2005 2; CitationJouve et al., 2005b 7; CitationJouve et al., 2008 7.
Basioccipital
-
Single wide ventral rugosity of basioccipital medial to basioccipital tubera: absent (0); present (1).
-
Basioccipital tubera in ventral view: oblong (0); V-shaped/tear-drop shaped (1).
-
Paired grooves along ventral surface, extending from base of occipital condyle to basioccipital tubera: absent (0); present (1).
-
Lateral eustachian foramen: level with medial eustachian foramen (0); dorsolateral to medial eustachian foramen, largely hidden from occipital view by the basioccipital (1). Modified from CitationJouve et al., 2006 145.
-
Ventral part of basioccipital: vertical, largely visible in occipital view (0); strongly inclined, weakly visible in occipital view (1). From: CitationJouve et al., 2005b 13; CitationJouve et al., 2008 13.
Quadratojugals
-
Extension of quadratojugal to cranial condyle for articulation with jaw: does not extend (0); extends largely (1). From: CitationJouve et al., 2005b 19; CitationJouve et al., 2008 19.
Palatines
-
Extension of the palatines to choanae: anterior margin or not (0); antermedial margin (1). From: CitationJouve et al., 2005b 4; CitationJouve et al., 2008 4.
Pterygoids
-
Anteromost point of posterior margin of pterygoidian wing: far anterior to medial eustachian foramen (0) about level with medial eustachian foramen (1). Modified from: CitationJouve, 2005 8; CitationJouve et al., 2005b 14; CitationJouve et al., 2008 14.
Laterosphenoids
-
Laterosphenoid sutures with parietal: parallel to skull table (0); descends posteriorly relative to the skull table (1).
Mandible
-
Size of second mandibular alveoli relative to first mandibular alveoli: similar in size (0); reduced in size relative to both adjacent alveoli (1).
-
Interalveolar space between second and third mandibular alveoli relative to that of first and second: roughly equal in proportion (0); greater (1).
-
Third mandibular alveoli placement: interalveolar space is roughly equal to that between second and third mandibular alveoli (0); close to mandibular fourth (1).
-
Fourth alveolus in proportion to other mandibular alveoli: enlarged (0); similar in size (1).
-
Fourth alveolar wall: level with adjacent alveoli (0); raised relative to adjacent alveoli (1).
-
First alveolus: directed entirely dorsal (0); slants anteriorly as well as dorsally (1).
-
Anterior portion of mandible: typically exhibits expansion (0); exhibits straight margins without significant lateral expansion (1). Modified from CitationSereno et al., 2001 72.
-
Point at which mandibular symphysis ends posteriorly: anterior to anterior 3/4 alveoli (0); posterior to anterior 3/4 alveoli (1).
-
Occlusal pits for maxillary occlusion: minimal (0); prolific (1).
-
Seventh alveolus: comparable in size to adjacent alveoli (0); reduced in size and close to eighth alveolus (1). Modified from: Jouve, 2004 153; CitationJouve, 2005 3; CitationJouve et al., 2005b 8; CitationJouve et al., 2006 164; CitationJouve et al., 2008 8.
-
Symphysis: wider than high (0); about as wide as high (1). From: CitationJouve et al., 2005b 17; CitationJouve et al., 2008 17.
-
Retroarticular process: short (0); extremely long and posterodorsally curved (1). From: CitationJouve et al., 2005b 1; CitationJouve et al., 2008 1.
-
Posteromedial wing of retroarticular process: dorsally situated or at mid-height (0); ventral on the retroarticular process (1). From: CitationJouve et al., 2005b 2; CitationJouve et al., 2008 2.
-
Coronoid: present (0); absent (1). From: CitationJouve et al., 2005b 6; CitationJouve et al., 2008 6.
-
External mandibular fenestra: absent or reduced to a thin slot (0); wide (1). Modified from: CitationJouve et al., 2005b 20; CitationJouve et al., 2008 20.
-
Posterior process of angular relative to posterior extremity of the retroarticular process: reaches process (0); remains far anterior (1). From: CitationJouve et al., 2008 39.
Dentition
-
Teeth: robust with very wide alveoli (0); thin and long teeth (1). From: CitationJouve, 2005 11; CitationJouve et al., 2005b 21; CitationJouve et al., 2008 21.
-
Distance between teeth from middle of maxilla or mandibular symphysis: nearly constant (0) much longer than in anterior portion of maxilla or mandibular symphysis (1). From CitationJouve et al., 2008 43.
Postcrania
-
Lateral margin of medial dorsal osteoderms: linear (0); serrated and curved, sutured with the lateral osteoderm (sends an acute process laterally) (1). From CitationJouve et al., 2008 37.
ACKNOWLEDGMENTS
Funding for field work was made possible by the National Science Foundation grants DEB-0733725 and DSGC 640179, the Smithsonian Tropical Research Institute Paleobiology Fund, the Geological Society of America Graduate Student Research Grant, the R. Jerry Britt Jr. Paleobiology Award (Florida Museum of Natural History), and the Cerrejón Coal mine. Many thanks to L. Teicher, F. Chavez, G. Hernandez, C. Montes, and the rest of the crew at the Cerrejón coal mine for permission to study the material, access to the site, and housing during field work. Thanks to A. Rincon, F. Herrera, and M. Carvalho for field collecting. Thanks to J. Nestler for her tireless efforts of research assistance, literature translation, and photograph production. Thanks also to the efforts of K. and N. Hastings for their assistance in literature translation. In addition, thanks go to J. Nestler and J. Bourque for their revision comments of the manuscript. Further thanks to C. Brochu for informative conversations. Thanks to P. Lanzarone who produced initial sketches of the specimens. Thanks also to the two reviewers of the manuscript for their thorough and helpful revisionary comments. Continued thanks to W. Joyce and S. Chester for access to and assistance with the collections at YPM and to C. Mehling and J. Conrad for access to and assistance with the collections at the AMNH.
Related Research Data
LITERATURE CITED
- Albertâo , G. , Koutshoukos , E. , Regali , M. , Attrep , M. and Martin , P. 1993 . The Cretaceous-Tertiary boundary (KTB) at Pernambuco/Paraíba Basin, Northeastern Brazil . Acta Geologica Leopoldensia , 16 : 59 – 71 .
- Arambourg , C. 1952 . Les vertébrés fossiles des gisements de phosphates (Maroc, Algérie, Tunisie) . Notes et Mémoires du Service géologique du Maroc , 92 : 1 – 372 .
- Argollo , J. , Buffetaut , E. , Cappetta , H. , Fornari , M. , Herail , G. , Laubacher , G. , Sigé , B. and Vizcarra , G. 1987 . Découverte de vertébrés aquatiques présumés Paléocènes dans les Andes Septentrionales de Bolivie (Rio Suches, Sinclinorium de Putina) . Geobios , 20 : 123 – 127 .
- Barbosa , J. A. , Kellner , A. W. A. and Viana , M. S. S. 2008 . New dyrosaurid crocodylomorph and evidences for faunal turnover at the K-P transition in Brazil . Proceedings of the Royal Society B , 275 : 1385 – 1391 .
- Bloch , J. , Cadena , E. , Herrera , F. , Wing , S. L. and Jaramillo , C. 2005 . Paleocene vertebrates from the Cerrejón Formation, Guajira Peninsula, northeastern Colombia . Journal of Vertebrate Paleontology , 25 : 37A – 38A .
- Bloch , J. , Cadena , E. , Hastings , A. , Rincon , A. and Jaramillo , C. 2008 . Vertebrate faunas from the Paleocene Bogotá Formation of Northern Colombia . Journal of Vertebrate Paleontology , 28 : 53A
- Brochu , C. A. 1996 . Closure of neurocentral sutures during crocodilian ontogeny: implications for maturity assessment in fossil archosaurs . Journal of Vertebrate Paleontology , 16 : 49 – 62 .
- Brochu , C. A. , Bouaré , M. L. , Sissoko , F. , Roberts , E. M. and O’Leary , M. A. 2002 . A dyrosaurid crocodyliform braincase from Mali . Journal of Paleontology , 76 : 1060 – 1071 .
- Buffetaut , E. 1976a . Sur la répartition géographique hors d’Afrique des Dyrosauridae, crocodiliens mésosuchiens du Crétacé terminal et du Paléogène . Comptes Rendu de l’Académie des Sciences, D , 283 : 486 – 490 .
- Buffetaut , E. 1976b . Une nouvelle définition de la famille des Dyrosauridae de Stefano, 1903 (Crocodylia, Mesosuchia) et ses conséquences: inclusion des genres Hyposaurus et Sokotosuchus dans les Dyrosauridae . Geobios , 9 : 333 – 336 .
- Buffetaut , E. 1977 . Donnees nouvelles sur les crocodiliens paléogènes du Pakistan et de Birmanie . Comptes Rendus de l’Academie des Sciences de Paris , 285 : 869 – 872 .
- Buffetaut , E. 1978 . Les Dyrosauridae (Crocodylia, Mesosuchia) des phosphates de l’Eocene inférieur de Tunisie: Dyrosaurus, Rhabdognathus, Phosphatosaurus . Géologie Mediterranéene , 5 : 237 – 256 .
- Buffetaut , E. 1979 . Sokotosuchus ianwilsoni and the evolution of the dyrosaurid crocodiles . Nigerian Field Monograph , 1 : 31 – 41 .
- Buffetaut , E. 1980 . “ Les crocodiliens paléogenes du Tilemsi (Mali), un aperçu systematique; Palaeovertebrata, Mémoire Jubilaire Rene Lavocat ” . 15 – 35 . Montpelier : Laboratoire de Paléontologie des Vertébrés de l’Ecole Practique des Hautes Études .
- Buffetaut , E. 1991 . Fossil crocodilians from Tiupampa, (Santa Lucia Formation, Early Paleocene) Bolivia: a preliminary report . Revista Técnica de YPFB, Fosiles y Facies da Bolivia, I, Vertebrados , 12 : 541 – 544 .
- Buffetaut , E. and Lauverjat , J. 1978 . Un crocodilian d’un type particulier dans le Cénomanien de Nazaré (Portugal) . Comptes Rendus sommaires des Séances de la Société Géologique de France , 20 : 78 – 82 .
- Buffetaut , E. and Wouters , G. 1979 . Atlantosuchus coupatezi, n. g., n. sp., un nouveau Dyrosauride (Crocodylia, Mesosuchia) des phosphates Montiens du Maroc . Bulletin Trimestriel de la Société Géologique de Normandie et des Amis du Muséum du Havre , 66 : 85 – 90 .
- Buffetaut , E. , Bussert , R. and Brinkmann , W. 1990 . A new nonmarine vertebrate fauna in the Upper Cretaceous of northern Sudan . Berliner Geowissenschaftlische Abhandlungen , 120 : 183 – 202 .
- Carvalho , L. B. de and de Azevedo , S. A. K. 1997 . Um crocodile marinho (Mesosuchia: Dyrosauridae) no paleoceno da Bacia Pernambuco-Paraiba, Brasil . Ameghiniana , 34 : 532
- Clark , J. M. 1994 . “ Patterns of evolution in Mesozoic Crocodyliformes. In the Shadow of the Dinosaurs ” . Edited by: Fraser , N. C. and Sues , H.-D. 84 – 97 . New York : Cambridge University Press .
- Cope , E. D. 1885 . A contribution to the vertebrate paleontology of Brazil . Paleontological Bulletin , 40 : 1 – 20 .
- Cope , E. D. 1886 . A contribution to the vertebrate paleontology of Brazil . Proceedings of the American Philosophical Society , 23 : 1 – 20 .
- de Lapparent de Broin , F. 2002 . Elosuchus, a new genus of crocodile from the Lower Cretaceous of the North of Africa . Comptes Rendu Palevol , 1 : 275 – 285 .
- De Stefano , G. 1903 . Nuovi rettili degli strati a fosfato della Tunisia . Bolletino della Societa Geologia Italiana , 22 : 51 – 80 .
- Denton , R. K. , Dobie , J. L. and Parris , D. C. 1997 . “ The marine crocodilian Hyposaurus in North America. Ancient Marine Reptiles ” . Edited by: Callaway , J. M. and Nicholls , E. L. 375 – 397 . New York : Academic Press .
- Gallo , V. , Figueiredo , F. J. D. , de Carvalho , L. B. and de Azevedo , S. A. K. 2001 . Vertebrate assemblage from the Maria Farinha Formation after the K-T boundary . Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen , 219 : 261 – 284 .
- Gasparini , Z. and Spalletti , L. A. 1990 . Un nuevo cocodrilo en los depósitos mareales Maastrichtianos de la Patagonia noroccidental . Ameghiniana , 27 : 141 – 150 .
- Gasparini , Z. , Chiappe , L. M. and Fernandez , M. 1991 . A new Senonian peirosaurid (Crocodylomorpha) from Argentina and a synopsis of the South American Cretaceous Crocodilians . Journal of Vertebrate Paleontology , 11 : 316 – 333 .
- Gasparini , Z. 1996 . Biogeographic evolution of the South American crocodilians. Münchner Geowissenschaftliche Abhandlungen . Reihe A, Geologie und Palaontologie , 30 : 159 – 184 .
- Gasparini , Z. and de la Fuente , M. 2000 . Tortugas y plesiosaurios de la Formación La Colonia (Cretácico Superior) de Patagonia, Argentina . Revista Española de Paleontología , 15 : 23 – 35 .
- Gasparini , Z. 2007 . “ Plesiosauria. Patagonian Mesozoic Reptiles ” . Edited by: Gasparini , Z. , Salgado , L. and Coria , R. 292 – 313 . Indianapolis, Indiana : Indiana University Press .
- Gradstein , F. , Ogg , J. and Smith , A. 2004 . “ A Geologic Time Scale 2004 ” . 589 Cambridge, , Uniked Kingdom : Cambridge University Press .
- Halstead , L. B. 1973 . Hunting pre-historic reptiles in Nigeria . The Nigerian Field , 38 : 1 – 14 .
- Halstead , L. B. 1975 . Sokotosuchus ianwilsoni n. g., n. sp., a new teleosaur crocodile from the Upper Cretaceous of Nigeria . Journal of Mining and Geology , 11 : 101 – 103 .
- Halstead , L. B. and Middleton , J. 1976 . Fossil vertebrates of Nigeria, Pt. III . The Nigerian Field , 41 : 166 – 174 .
- Hastings , A. and Bloch , J. 2007 . New short-snouted dyrosaurid (Crocodylomorpha) from the Paleocene of Northern Colombia . Journal of Vertebrate Paleontology , 27 : 87A – 88A .
- Hastings , A. and Bloch , J. 2008 . New long-snouted dyrosaurid (Crocodylomorpha, Mesoeucrocodylia) from the Paleocene of Northeastern Colombia . Journal of Vertebrate Paleontology , 28 : 89A
- Hay , O. P. 1930 . “ Second bibliography and catalogue of the fossil Vertebrata of North America. Vol. 2 ” . Vol. 2 , 1 – 1074 . Carnegie Institute of Washington Publication .
- Head , J. , Bloch , J. , Hastings , A. , Bourque , J. , Cadena , E. , Herrera , F. , Polly , P. and Jaramillo , C. 2009 . Giant boid snake from the Paleocene neotropics reveals hotter past equatorial temperatures . Nature , 457 : 715 – 718 .
- Jaramillo , C. , Pardo-Trujillo , A. , Rueda , M. , Harrington , G. , Bayona , G. , Torres , V. and Mora , G. 2007 . Palynology of the Upper Paleocene Cerrejón Formation, Northern Colombia . Palynology , 31 : 153 – 189 .
- Jouve , S. 2004 . Etude des Crocodyliformes fini Crétacé-Paléogène du Bassin des Oulad Abdoun (Maroc) et comparaison avec les faunes africaines contemporaines: systématique, phylogénie et paléobiogéographie . : 651 Unpublished Ph.D. dissertation, Muséum National d’Histoire Naturelle, Paris
- Jouve , S. 2005 . A new description of Dyrosaurus phosphaticus (Thomas, 1893) (Mesoeucrocodylia: Dyrosauridae) from the Lower Eocene of North Africa . Canadian Journal of Earth Sciences , 42 : 323 – 337 .
- Jouve , S. 2007 . Taxonomic Revision of the Dyrosaurid Assemblage (Crocodyliformes: Mesoeucrocodylia) from the Paleocene of the Iullemmeden Basin, West Africa . Journal of Paleontology , 81 : 163 – 175 .
- Jouve , S. and Schwarz , D. 2004 . Congosaurus bequaerti, a Paleocene dyrosaurid (Crocodyliformes; Mesoeucrocodylia) from Landana (Angola) . Bulletin de l’Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre , 74 : 129 – 146 .
- Jouve , S. , Bouya , B. and Amaghzaz , M. 2005a . A short-snouted dyrosaurid (Crocodyliformes, Mesoeucrocodylia) from the Palaeocene of Morocco . Palaeontology , 48 : 359 – 369 .
- Jouve , S. , Bouya , B. and Amaghzaz , M. 2008 . A long-snouted dyrosaurid (Crocodyliformes, Mesoeucrocodylia) from the Paleocene of Morocco: phylogenetic and palaeobiogeographic implications . Palaeontology , 51 : 281 – 294 .
- Jouve , S. , Iarochène , M. , Bouya , B. and Amaghzaz , M. 2005b . A new crocodyliform dyrosaurid from the Palaeocene of Morocco and a phylogenetic analysis of Dyrosauridae . Acta Palaeontologica Polonica , 50 : 581 – 594 .
- Jouve , S. , Iarochène , M. , Bouya , B. and Amaghzaz , M. 2006 . A new species of Dyrosaurus (Crocodylomorpha, Dyrosauridae) from the early Eocene of Morocco: phylogenetic implications . Zoological Journal of the Linnean Society , 148 : 603 – 656 .
- Langston , W. L. 1995 . Dyrosaurs (Crocodilia, Mesosuchia) from the Paleocene Umm Himar Formation, Kingdom of Saudi Arabia. U.S . Geological Survey Bulletin , 2093-F : F1 – F36 .
- Maddison , W. P. and Maddison , D. R. 2008 . Mesquite: a modular system for evolutionary analysis. Version 2.5 . Available at http://mesquiteproject.org. Accessed October 8, 2008
- Marshall , L. G. and de Muizon , C. 1988 . The dawn of the age of Mammals in South America . National Geographic Research , 4 : 23 – 55 .
- Moody , R. T. J. and Buffetaut , E. 1981 . Notes on the systematic and palaeoecology of the crocodiles and turtles of the Metlaoui Phosphates (Eocene) of Southern Tunisia . Tertiary Research , 3 : 125 – 140 .
- Ortega , F. , Gasparini , Z. B. , Buscalioni , A. D. and Calvo , J. O. 2000 . A new species of Araripesuchus (Crocodylomorpha, Mesoeucrocodylia) from the Lower Cretaceous of Patagonia (Argentina) . Journal of Vertebrate Paleontology , 20 : 57 – 76 .
- Parris , D. C. 1986 . Biostratigraphy of the fossil crocodile Hyposaurus Owen from New Jersey . New Jersey State Museum Investigation Number , 4 : 16
- Pomel , A. 1894 . Découverte de champsosauriens dans les gisements de phosphorite du suessonien de l’Algérie . Comptes Rendu de l’Académie des Sciences , 118 : 1309 – 1310 .
- Scotese , C. R. 2001 . “ Digital Paleogeographic Map Archive on CD-ROM ” . Arlington, Texas : PALEOMAP Project .
- Schwarz , D. , Frey , E. and Martin , T. 2006 . The postcranial skeleton of the Hyposaurinae (Dyrosauridae; Crocodyliformes) . Palaeontology , 49 : 695 – 718 .
- Sereno , P. C. , Larsson , H. C. E. , Sidor , C. A. and Gado , B. 2001 . The giant Crocodyliform Sarcosuchus from the Cretaceous of Africa . Science , 294 : 1516 – 1519 .
- Storrs , G. W. 1986 . A dyrosaurid crocodile (Crocodylia: Mesosuchia) from the Paleocene of Pakistan . Postilla Number , 197 : 1 – 16 .
- Swinton , W. E. 1930 . On the fossil Reptilia from Sokoto Province . Geological Survey of Nigeria , 13 : 1 – 62 .
- Swinton , W. E. 1950 . On Congosaurus bequaerti Dollo . Annales du Musée du Congo Belge , 13 : 9 – 56 .
- Swofford , D. 2003 . Massachusetts : Sunderland . PAUP* 4.0b10. Sinauer Associates
- Troxell , E. L. 1925 . Hyposaurus, a marine crocodilian . American Journal of Science , 8 : 489 – 514 .
- Vickaryous , M. K. and Hall , B. K. 2008 . Development of the dermal skeleton in Alligator mississippiensis (Archosauria, Crocodylia) with comments on the homology of Osteoderms . Journal of Morphology , 269 : 398 – 422 .
- Walker , A. D. 1970 . A revision of the Jurassic Reptile Hallopus victor (Marsh), with remarks on the classification of crocodiles . Philosophical Transactions of the Royal Society of London Series B, Biological Sciences , 257 : 323 – 372 .
- Whetstone , K. N. and Wybrow , P. J. 1983 . A ‘‘cursorial’’ crocodilian from the Triassic of Lesotho (Basutoland), southern Africa . Occasional Papers of the University of Kansas Museum of Natural History , 106 : 1 – 37 .
- Wu , X.-C. , Russell , A. P. and Cumbaa , S. L. 2001 . Terminonaris (Archosauria: Crocodyliformes): new material from Saskatchewan, Canada, and comments on its phylogenetic relationships . Journal of Vertebrate Paleontology , 21 : 492 – 514 .