ABSTRACT
Three clutches and eight additional eggs are described from the Upper Cretaceous Chichengshan Formation in the Lishui Basin of Zhejiang Province, China. The three clutches contain 6–18 partial or complete spherical eggs, 14–18 cm in diameter. The eggshell thickness and reticulate pore system are identical to Dictyoolithus hongpoensis CitationZhao, 1994. However, the eggshell lacks superimposed shell units previously reported in this oospecies. The difference in microstructure most likely results from the reliance on thin section analysis in the original study; furthermore, superimposed shell units are inadequately documented in all Dictyoolithus oospecies due to the absence of scanning electron microscopy (SEM) or low (×45) magnification of SEM imaging. Cladistic analysis and the presence of the following characters support a theropod affinity of Dictyoolithus hongpoensis: relatively narrow spacing of nucleation sites, mammillary cones with blocky calcite cleavage, prismatic columns, and parallel laminae within the shell units. We reevaluate previous assignment of Dictyoolithus eggs to the Dinosauroid-spherulitic Basic Type and advocate discontinuation of Basic Type and Morphotype in fossil egg classification.
INTRODUCTION
The Upper Cretaceous formations of the Tiantai Group exposed within in the Lishui Basin of Zhejiang Province yield a variety of dinosaur fossils, including ankylosaur, sauropod, and theropod osteological remains (CitationLü et al., 2007). However, dinosaur eggs are previously unreported from this region. In December of 2004, more than 100 fossil eggs were discovered during construction of Riyuetonghui Park, in the suburbs of Lishui City (). These fossil eggs recovered from the Upper Cretaceous Chichengshan Formation represent a monospecific assemblage, referable to the oofamily Dictyoolithidae.
Known exclusively from China, eggs of the Dictyoolithidae are remarkable because of their unusual reticulate pore system and extremely high porosity. CitationZhao (1994) described Dictyoolithus eggshell as comprised of three to five superimposed shell units and, on the basis of this interpretation, hypothesized an evolutionary trend in the calcification of dinosaur eggs. Previous descriptions of Dictyoolithus eggs are very brief and rely on radial thin sections of eggshell, examined by transmitted light microscopy (CitationZhao, 1994; CitationWang et al., 2006). Only one study (CitationLiu and Zhao, 2004) includes scanning electron microscope (SEM) images, but the low magnification (×45) prohibits identification of the superimposed shell units that purportedly comprise the eggshell.
Here, we report the first Dictyoolithus egg clutches from the Lishui Basin, near Lishui City, Zhejiang Province, in southeastern China. We describe new eggshell characters previously unreported in the oofamily Dictyoolithidae and review previous interpretations of superimposed shell units in Dictyoolithus hongpoensis and other Dictyoolithus oospecies. The phylogenetic affinities of D. hongpoensis were examined by conducting an analysis of a selection of Mesozoic and modern egg types.
Institutional Abbreviations—FPDM, Fukui Prefectural Dinosaur Museum, Japan; LSM, Lishui Museum, Lishui City, Zhejiang Province, China; ZMNH, Zhejiang Museum of Natural History, Hangzhou, China.
MATERIALS AND METHODS
Preparation of three egg clusters (LSM F008a, LSM F008b, ZMNH M8708-3) included removal of sediment from the eggs with small hand tools and stabilizing loose eggshell fragments with glue. Eggshells were removed from the three clusters (), five nearly complete eggs (ZMNH M8708-1, ZMNH M8707-1, ZMNH M8707-2, ZMNH M8733, ZMNH M8734), and an incomplete egg (FPDM 7744) (). The eggshell fragments were prepared as standard petrographic thin sections (30 μm thick) and studied by a Nikon Eclipse polarized light microscope and Olympus BX61 epifluorescent/light microscope. Additional eggshells were coated with gold (10 nm), mounted on aluminum stubs, and imaged at 15 kV under a J.R. Lee Instrument Personal SEM and JEOL 6100 SEM with Backscattered Electron Imaging. Structural attributes (shell thickness, shell unit width) were measured with Scion image analysis software. Clutches LSM F008a and LSM F008b are catalogued at the Lishui Museum, Lishui City, and ZMNH M8708-3 is catalogued at the Museum of Natural History, Hangzhou; both facilities are in Zhejiang Province, China. Eggshell thin sections are catalogued at the Museum of Natural History, Hangzhou, China, and the Fukui Prefectural Dinosaur Museum, Japan.
FIGURE 1 Location of Lishui City in Zhejiang Provice, southeastern China.
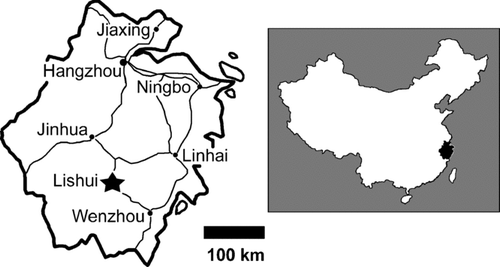
FIGURE 2 Three Dictyoolithus hongpoensis clutches and a partial egg from the Lishui Basin, Zhejiang Province, China. A, LSM F008b, a clutch containing at least 12 eggs; B, ZMNH M8708-3, a clutch containing 6 or more eggs; C, LSM F008a, a clutch of at least 18 eggs; D, FPDM 7744, a partial egg, black arrow points to thin shell on the egg surface, white arrow indicates unweathered eggshell contained within the matrix that fills the egg; scales bars equal 10 cm.
GEOLOGY
Zhejiang Province in southeastern China () contains approximately 40 small sedimentary basins that range in size from 100 to 3000 km2 (CitationWang et al., 2000). The Cretaceous Lishui Basin contains, in ascending sequence, the Tangshang, Liangtoutang, and Chichengshan formations of the Tiantai Group. Rocks of the Tangshang and Liangtoutang formations are radiometrically dated at 109.2 Ma and 105.9–103.2 Ma, respectively, placing them within the Early Cretaceous (Albian) period (CitationWang et al., 2000). The fossil eggs occurred in the Chichengshan Formation, which CitationWang et al. (2000) considered as Late Cretaceous in age; however, Wang and his colleagues did not report stage or radiometric dates from the formation. The fossil eggs occurred within a single stratum comprised of purple to red-gray mudflow conglomerates that intercalate with argillaceous siltstone. In addition to this conglomerate and siltstone facies, which represent deposition within an alluvial system (CitationWang et al., 2000), the Chichengshan Formation contains two grayish-white, massive rhyolitic tuffs.
SYSTEMATIC PALEONTOLOGY
Oofamily DICTYOOLITHIDAE CitationZhao, 1994
Type Oogenus—Dictyoolithus CitationZhao, 1994.
Included Oogenus—Stromatoolithus CitationZhao, Ye, Li, Zhao, and Yan, 1991.
Diagnosis—Radial view of eggshell with reticulate organization framed by irregular basic structural units.
Distribution—Stromatoolithus from Maastrichtian Pingling Formation, Guangdong Province, China; for Dictyoolithus oogenera see and references therein.
TABLE 1 Dictyoolithus oospecies from China.
Oogenus DICTYOOLITHUS CitationZhao, 1994, sensu CitationLiu and Zhao, 2004
Type Oospecies—Dictyoolithus hongpoensis CitationZhao, 1994.
Included Oospecies—Four oospecies: Dictyoolithus hongpoensis CitationZhao, 1994; D. neixiangensis CitationZhao, 1994; D. jiangi CitationLiu and Zhao, 2004; and D. gongzhulingensis CitationWang et al., 2006.
Revised Diagnosis—Modified from CitationLiu and Zhao (2004). Eggs, oval or ovate in shape; basic structural units with parallel laminae; single or multiple superimposed shell units, low nodes on shell surface.
Distribution—See and references therein.
Oospecies DICTYOOLITHUS HONGPOENSIS CitationZhao, 1994:190, fig. 12.4 ()
Syntype—Two broken eggs, field number 79001.
Type Locality and Horizon—Hongpo, Xixia County, Henan Province, Lower Cretaceous Sigou Formation; however, CitationCheng et al. (1995) reported the formation as Upper Cretaceous. Liu and CitationZhao (1994) included the city of Chishuigou in Neixiang County, Henan Province in the type locality but provided no additional information.
Referred Material—Clutches LSM F008a, LSM F008b, and ZMNH M8708-3, from Lishui Basin, Zhejiang Province, China; partial eggs FPDM 7744, ZMNH M8735, ZMNH M8738, ZMNH M8734, ZMNH M8708-1, ZMNH M8708-2, ZMNH M8707-1, ZMNH M8707-2; eggshell thin sections, FPDM.V.7744-1, FPDM.V.7744-2, FPDM.V.7744-3, FPDM.V.7744-4.
Range—See .
Revised Diagnosis—Modified from CitationZhao (1994). Although no unambiguous autapomorphies are recognized, the oospecies can be differentiated from Dictyoolithus neixiangensis, D. gongzhulingensis, and D. jiangi by the following unique combination of features: clutches contain a single layer of up to 18 oval or ovate eggs, egg size from 12 to 16 cm in diameter; eggshell with closely spaced nucleation sites; shell thickness from 2.5 to 2.8 mm.
DESCRIPTION
Three egg clusters and eight isolated eggs were recovered from the Upper Cretaceous Chichengshan Formation in the suburbs of Lishui City in Zhejiang Province, China (). All clutches occurred within the same stratum. The outcrop was leveled during construction, thus the site provides limited stratigraphic and taphonomic information, and the original dimensions, distribution, and number of eggs in each cluster remains uncertain. Two clusters, LSM F008b and ZMNH M8708-3, contain at least 12 and 6 eggs, respectively (). The largest cluster, LSM F008a, consists of at least 18 partial and complete eggs (). In each cluster, the eggs comprise a single layer within in a red, matrix-supported pebble conglomerate. None of the eggs are superimposed. Some eggs touch one another, but most are separated by a few to several centimeters. Somewhat distorted by lithologic compression, the eggs are only partly exposed within the matrix, and their maximum diameters vary from 10.6 cm in a poorly exposed specimen, to 16 cm in a well-exposed egg. Eight relatively complete eggs from the same locality are free of matrix and likely provide an accurate size estimate; these range from 14.3 cm × 13.7 cm to 15.8 cm × 14.0 cm.
The eggshell samples vary in thickness in accordance with the degree of weathering. One eggshell preserved within the sediment that fills FPDM 7744 shows little evidence of abrasion or calcite dissolution and measures 2.8 mm; we consider this as the original shell thickness (). Substantially thinner shell (0.66–1.8 mm) covers the outer surface of the same egg and results from recent weathering.
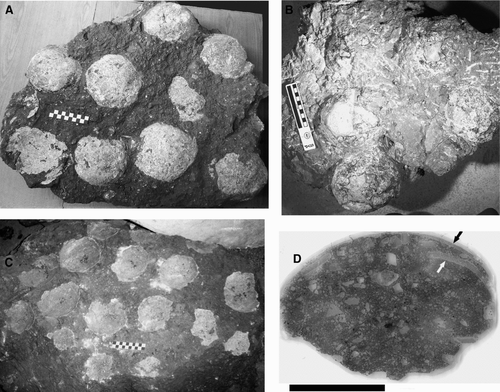
Thin sections generally reveal the complex structure of the eggshell more clearly than SEM (). Calcite nuclei at the inner surface of the eggshells are poorly preserved, but where present these nuclei are about 0.10 mm in diameter. The thin bifurcating shell units that comprise the eggshell exhibit no evidence of radiating calcite spherulites (). The eggshell displays abundant irregular pores, and the shell units occasionally exhibit faint parallel laminations and a columnar extinction pattern under polarized light microscopy (). Epifluorescent imaging resulted in no additional information beyond that provided by thin-section analysis; however, SEM reveals mammillae with blocky calcite structure and prismatic columns that exhibit horizontal laminations (tabular structure) (). Although not definitive, a structural transition may occur in the lower one-fifth of the eggshell in well-preserved specimens (). Qualitative assessment suggests that the pore diameters are greater in the inner portion of the eggshell, and pore area/shell unit area ratio decreases in an outward direction from the nucleation sites (). The shell unit widths vary from about 0.06 to 0.08 mm. Although the egg surface appears smooth, well-preserved eggshells display the rounded, narrow apical parts of the prismatic columns that are absent in more weathered samples ().
FIGURE 3 Dictyoolithus hongpoensis from the Lishui Basin, China. A, Radial thin section of eggshell (FPDM.V. 7744-3) in plain polarized light, showing shell units and reticulate pore system. Outer shell surface is at the top of the image, and the calcite-filled pores are lighter in color than the eggshell. A crack extends laterally through the upper portion of the specimen; scale bar equals 1 mm; B, same as A, under crossed polars; note columnar extinction pattern and possible structural transition in the lower one fifth of the eggshell; C, SEM image of M8735, triangles indicate width of mammillae, rectangular box shows location of prismatic column in E, scale bar equals 1 mm; D, mammillary cone shown in C; arrow indicates preserved calcite nuclei, scale bar equals 100 μm; E, close up of prismatic column with parallel laminae shown in D, scale bar equals 100 μm; F, apical portion of a shell unit in M8735 separated by pores, viewed under light microscopy; arrows indicate margin of one of two adjacent shell units; scale bar equals 100 μm.
Remarks
The highly porous nature of the eggshell contributes to lateral crack propagation during lithostatic compression (). When the shell breaks along these cracks, abrasion and calcite dissolution may further reduce the shell thickness. The greater pore diameter, possibly enhanced by dissolution that characterizes the inner portion of the eggshell, often causes these pores to appear more conspicuous in thin sections. These diagenetic changes may so alter individual specimens as to lead to misinterpretation of these ‘thin’ specimens as a different oospecies.
DISCUSSION
Comparisons
We interpret the three egg clusters as in situ but incomplete egg clutches. The 18 eggs in LSM F008a exceed the six and 12 eggs that comprise LSM F008b and ZMNH M8708–3, respectively. Although the eggs in LSM F008a occur at slightly different levels in the matrix, they are not superimposed, thus suggesting that they represent a single clutch, rather than eggs combined from different horizons. Eggshells removed from six eggs reveal reticulate pore system, surface ornamentation that consists of low nodes, and similar shell thickness as Dictyoolithus CitationZhao, 1994, one of two oogenus within the oofamily Dictyoolithidae CitationZhao, 1994. Comparison to the second oogenus, Stromatoolithus (CitationZhao et al., 1991), is difficult because of the very brief description of the specimens; however, Zhao and his colleagues report the surface ornamentation as “wormlike” ridges, suggesting sagenotuberculate morphology that differs from that of Dictyoolithus (CitationCarpenter, 1999).
The oogenus Dictyoolithus contains four oospecies known exclusively from China. Isolated specimens of Dictyoolithus hongpoensis and D. neixiangensis are reported from the Sigou and Gaogou formations, respectively, in the Xixia Basin of Henan Province (CitationZhao, 1994). CitationZhao (1994) considered both formations as Lower Cretaceous; in contrast, CitationCheng et al. (1995) reported these formations as Upper Cretaceous. Dictyoolithus jiangi and D. gongzhulingensis occurred in the Upper Cretaceous Jiangjunding Formation of Shandong Province (CitationLiu and Zhao, 2004) and the Lower Cretaceous Quantou Formation of Jilin Province (CitationWang et al., 2006), respectively.
The Lishui Dictyoolithus eggs are larger than D. gongzhulingensis (CitationWang et al., 2006) and Dictyoolithus neixiangensis (CitationZhao, 1994), but overlap with the upper size range of D. jiangi (CitationLiu and Zhao, 2004). The thickness of the Lishui eggshell exceeds that of all three of these oospecies (). Compared to the Lishui eggs, D. neixiangensis and D. gongzhulingensis exhibit greater pore diameter throughout the eggshell, widely spaced nuclei, and a continuous calcite ‘band’ or interlocking shell units at the outer eggshell surface (compare to CitationZhao, 1994:fig.12.5; CitationWang et al., 2006:fig. 4C, D). Light microscopy also suggests that shell units in D. gongzhulingensis consist of radiating spherulites, which are not present in the Lishui eggs. The Lishui fossil eggs closely resemble Dictyoolithus hongpoensis in eggshell thickness, narrow shell units, and reticulate pore system (CitationZhao, 1994). However, some microstructural characteristics differ from previous descriptions of eggs assigned to the Dictyoolithidae; therefore, we discuss the structure of dictyoolithid eggs and their classification in egg parataxonomy in more detail below.
The Structure of Dictyoolithid Eggs
In brief, preliminary description of the new oogenus and oospecies Dictyoolithus hongpoensis and D. neixiangensis, CitationZhao (1994) noted their unusual reticulate pore system and suggested that the eggshells consisted of superimposed shell units. Purportedly, Dictyoolithus hongpoensis included five or more, whereas D. neixiangensis exhibited two to three, shell units. CitationZhao (1994) compared the formation of Dictyoolithus eggshell to that of the extant tuatara (Sphenodon punctatus) in which the membrane and eggshell form simultaneously, rather than sequentially (as in modern archosaurs), within the oviduct. He postulated that simultaneous formation of membrane and shell in dictyoolithid eggs might have provided nucleation sites for the crystalline shell units during the initial stage of eggshell formation; subsequent shell units then formed over the first ones. Based on his interpretation of Dictyoolithus microstructure, CitationZhao (1994) proposed a trend in dinosaur eggshell evolution from Early to Late Cretaceous time.
The schematic diagram that illustrates this proposed trend in dinosaur egg calcification (CitationZhao, 1994:figs. 12.12, 12.13) shows widely spaced nuclei at the inner shell surface; in addition, calcite nuclei occur at the base of each interpreted superimposed shell unit in Dictyoolithus eggshell. These features, however, are not described in the paper that established D. hongpoensis and D. neixiangensis within the new oofamily Dictyoolithidae. The paper does not include SEM images of the eggshell, and the radial thin sections (viewed by light microscopy) do not show these nuclei or a structural change that allows differentiation of the purported superimposed shell units (CitationZhao, 1994:figs. 12.4, 12.5). Furthermore, CitationZhao (1994) did not include superimposed shell units in the diagnoses of the oofamily Dictyoolithidae or oogenus Dictyoolithus.
Subsequent papers also suggest that other Dictyoolithus oospecies consist of superimposed shell units, and this feature is used to distinguish Dictyoolithus ootaxa at the oospecies level (; CitationLiu and Zhao, 2004; CitationWang et al., 2006). In addition, CitationLiu and Zhao (2004) modified the oogenus diagnosis to include the interpreted superimposed eggshells. Although illustrated in a schematic diagram, CitationWang et al. (2006) did not describe or include photographs of the nuclei at the base of three purported superimposed shell units in D. gongzhulingensis; furthermore, the paper lacks SEM images, relying solely on thin section analysis.
CitationLiu and Zhao (2004) provide the only SEM image of Dictyoolithus eggs. However, the low magnification (i.e., ×45) prevents definitive identification of the purported superimposed shells in D. jiangi. in their paper shows a possible nucleation site at the left side of the eggshell; however, this feature may represent an imaging artifact. Other shell units in the same and presumably a different eggshell (CitationLiu and Zhao, 2004:fig. 2C, D) do not show nuclei or a structural change that would denote the two to three superimposed layers reported in the paper. Superimposed shell units, therefore, are inadequately documented in all Dictyoolithus oospecies. Based on our study of eggs from the Lishui Basin, we conclude that Dictyoolithus hongpoensis does not consist of superimposed layers; therefore, we modified the oogenus and oospecies diagnoses accordingly. The presence of superimposed shell units in other Dictyoolithus oospecies remains uncertain and requires further SEM study.
Classification of the Dictyoolithidae
CitationMikhailov et al. (1996) established uniform criteria for description and classification of fossil eggs. Dinosaur eggs were separated into three categories: Dinosauroid spherulitic, Dinosauroid prismatic, and Ornithoid. The Dictyoolithidae was assigned to the Dinosauroid-spherulitic Basic Type (see also CitationHirsch, 1996; CitationMikhailov, 1997; CitationCarpenter, 1999). However, other egg types referred to this category (e.g., Megaloolithidae, Spheroolithidae, Dendroolithidae) exhibit calcite spherulites that emanate from nucleation sites at the inner shell surface and radiate outward, forming the single structural layer of the eggshell. CitationZhao (1994), however, did not describe spherulitic microstructure in Dictyoolithus eggshell, nor is this structure apparent in thin sections of the type specimens D. hongpoenis or D. neixiangensis (CitationZhao, 1994:figs. 12.4,12.5). To the contrary, he states that the structure of Dictyoolithus eggshell differs from eggs of the Spheroolithidae (CitationZhao, 1994:194). Thin sections and SEM imaging of Dictyoolithus hongpoensis from Lishui (reported here) do not reveal shell units comprised of radiating spherulites; in contrast, the eggs exhibit a mixed suite of features that occur in theropod and non-theropod eggs.
Eggshell attributes that support a theropod affinity for the Lishui eggs include closely spaced nuclei, mammillary cones with blocky calcite cleavage, and prismatic columns with parallel laminae (CitationMikhailov, 1997; CitationZelenitsky et al., 2002; CitationGrellet-Tinner and Chiappe, 2004; CitationVarricchio and Jackson, 2004). However, other features that are typical of most theropod eggs remain equivocal or absent in the Lishui eggs. For example, nearly all theropod eggs (but see CitationManning et al., 1997, Citation1999) exhibit squamatic texture and a gradual or abrupt transition from the mammillary to the overlying prismatic or cryptoprismatic layer (CitationMikhailov, 1997; CitationZelenitsky et al., 2002; CitationGrellet-Tinner and Chiappe, 2004; CitationVarricchio and Jackson, 2004). Squamatic texture is absent in Dictyoolithus eggs from the Lishui Basin. Change in extinction pattern suggests that a gradual transition may occur between the mammillary and prismatic layers in the lower one fifth of FPDM 7794 (). However, this feature may be difficult to identify in more altered eggshells because calcite dissolution typically enhances pore diameters at the inner eggshell surface, thereby limiting the opportunity to clearly view the basal portion of the eggshell.
Other characters that occur in the Lishui eggs are not characteristic of Late Cretaceous theropod eggs. These include ovate (rather than elongate) egg shape and massed eggs that comprise the clutch (rather than within-clutch egg pairing). However, ovate shape occurs in an egg containing a therizinosaurid embryo from the Cretaceous (Santonian-Campanian) Nanchao Formation, Henan Province, China (CitationManning et al., 1997, Citation1999). Dictyoolithus eggs are somewhat similar to Jurassic eggs from Portugal (CitationMateus et al., 1997); these allosaurid eggs are massed and do not display the elongate shape that typifies troodontid and oviraptorid eggs. These features may represent plesiomorphs in theropod eggs. To further assess the phylogenetic affinities of Dictyoolithus hongpoenis, we conducted an analysis of Mesozoic and modern egg types.
PHYLOGENETIC ANALYSIS
Methods and Characters
Phylogenetic analysis was conducted using PAUP 4.0 (CitationSwofford, 1998) on Dictyoolithus and 16 other taxa or ootaxa. Of the 17 taxa or ootaxa, seven represent extant reptiles or birds, seven are extinct dinosaurs with eggs identified on the basis of embryonic remains or closely associated adults, and three (including Dictyoolithus) are strictly Mesozoic ootaxa. With the exception of Deinonychus (cf. Elongatoolithus) (CitationGrellet-Tinner and Makovicky, 2006), Macroelongatoolithus (CitationJin et al., 2007), and Dictyoolithus (this paper), these taxa and ootaxa are taken from CitationVarricchio and Jackson (2004). The data matrix also includes 16 characters taken or modified from CitationVarricchio and Jackson (2004) and three new characters (Appendix 1, 2, characters 9, 10, and 11). Of these all but two are unordered (Appendix 1, characters 2, 18). Two characters (Appendix 1, characters 10 and 11) were uninformative but retained for future reference. Character and character-state phrasing follow the recommendations of CitationSereno (2007).
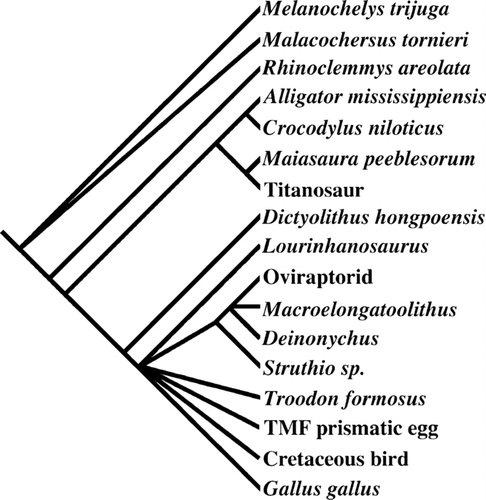
FIGURE 4 Majority-rule consensus tree for the shortest trees and all those up to three steps longer produced from the phylogenetic analysis of Dictyoolithus hongpoensis eggs. A strict consensus tree of the 39 shortest trees differs only in incorporating Dictyoolithus into the theropod polytomy. See Appendix 1 for oospecies names for eggs identified by embryonic elements or closely associated adult specimens.
It should be noted that CitationGrellet-Tinner and Makovicky (2006) were unable to replicate a previous cladistic analysis conducted by two of us (CitationVarricchio and Jackson, 2004) because the notation ‘ordered’ was inadvertently omitted from two character states. These characters are listed here as 2 and 18 (Appendix 1). Some characters in CitationVarricchio and Jackson (2004) also differ from CitationGrellet-Tinner and Makovicky (2006) because of incorrect coding in the latter or differences in interpretation. CitationZelenitsky and Therrien (2008) provide a detailed discussion of the incorrect coding and differences in character interpretation; therefore, we discuss only those characters not fully addressed in their paper.
CitationGrellet-Tinner and Makovicky (2006) reported a single structural layer in Alligator eggshell, whereas CitationFerguson (1985) reported multiple eggshell layers. Based on our SEM comparison of the calcite structure to the protein morphology of decalcified Alligator eggshell, we concur with CitationFerguson (1985). Previous descriptions report Troodon pores as straight (CitationHirsch and Quinn, 1990; CitationZelenitsky and Hills, 1996; CitationVarricchio et al., 2002), whereas CitationGrellet-Tinner and Makovicky (2006) score the pores as oblique. The putative pore in their paper, however, cuts across the prismatic column, rather than occurring parallel to the column, as typical of Troodon eggshell. We interpret this feature as a calcite-filled crack in the eggshell and score Troodon pores as straight in our analysis. In addition, a thin line occurs in the outer portion of Troodon eggshells, from different eggs and clutches recovered from the Two Medicine Formation of Montana. We interpret this feature as corresponding to the transition to the external layer (CitationVarricchio and Jackson, 2004). In contrast, CitationGrellet-Tinner and Makovicky (2006) consider this line in Troodon eggshell as diagenetic because it fluoresces under cathodoluminescence (CL). However, features that are biologic in origin and subsequently replace by magnesium also fluoresce under CL (CitationMarshall, 1988), as demonstrated by the mammillary layer of Lourinhanosaurus eggshell, also illustrated by CitationGrellet-Tinner and Makovicky (2006):fig. 4E, F). Finally, because coding of an external layer in Lourinhanosaurus (CitationVarricchio and Jackson, 2004) was not based on our personal examination, we coded this character as ‘unknown’ in the current analysis until further study.
Results of Phylogenetic Analysis
A heuristic search produced 39 equally parsimonious trees, each with a length of 41 steps, a C.I. of 0.76, and an R.I. of 0.85. The majority-rule consensus tree differs from the strict consensus tree in creating only one additional node: the placement of Dictyoolithus as the sister taxon to all other theropods. This topology was also maintained in majority-rule consensus trees generated from trees one to three steps longer (up to 44 steps) than the most-parsimonious ones. The consensus trees place the three turtles as outgroups to the clade of extant and extinct archosaurs ().
The archosaurs are united by the calcite composition of the shell. Within the archosaurs, the hadrosaur Maiasaura and the Argentine titanosaur fall as sister taxa to the two crocodilians. This may reflect the limited number of informative characters and low diversity among taxa and ootaxa available for the analysis. CitationGrellet-Tinner and Makovicky (2006) report similar placement in their analysis of eggs and reproductive attributes. The single remaining clade of nonavian and avian theropods are united by the following unambiguous synapomorphies: relatively narrow spacing of nucleation centers, presence of a strictly crystalline mammillary with blocky radiating crystals, and a second prismatic structural layer. Within the theropods, Dictyoolithus falls as the sister taxon to a clade of all theropods plus the ootaxa Macroelongatoolithus and the Two Medicine prismatic theropod egg. This clade exists as a large polytomy with only one resolved branch consisting of Struthio as the outgroup to a polytomy of Deinonychus, oviraptorid, and the oogenus Macroelongatoolithus. These taxa share a mammillary layer with truncated radiating crystals, an abrupt transition to the second structural layer, and squamatic structure that masks the prismatic nature of the second structural layer. Ornamentation and absence of a third layer separate the three Cretaceous forms from Struthio. An Adams consensus tree places both the allosauroid Lourinhanosaurus and Dictyoolithus as sister taxa to a clade of more derived theropods. These more-derived theropods unambiguously share squamatic structure in the second layer. Additional ambiguous features that may unite them include a third structural layer, a cuticle, asymmetric egg shape, and large eggs relative to adult size, and egg pairing within the clutch.
Bootstrap analysis of the data set using 1000 replicates and maximum trees saved value of 90,000 trees found the clade of all theropods plus the ootaxa Macroelongatoolithus, Dictyoolithus, and the Two Medicine prismatic egg to be represented in 67% and the archosaur clade in 53% of the replicates. The clade of Deinonychus, Macroelongatoolithus, and oviraptor was recovered in 74% and this group with Struthio in 53%. Cladistic analysis demonstrates that the theropod nature of Dictyoolithus is supported by the presence of the following characters: relatively narrow spacing of nucleation centers, blocky radiating crystals in the strictly crystalline mammillary layer, and a second prismatic structural layer (). However, the small taxonomic sample available for non-theropod dinosaurs perhaps mitigates the strength of this conclusion.
Reticulate Pores in Other Theropod Eggs
Manning et al. (Citation1997, Citation1999) reported an egg with a reticulate pore system containing a therizinosaurid embryo from the Cretaceous Nanchao Formation, Henan Province, China. However, Manning and his colleagues did not provide photographs or describe the eggshell structure, but merely assigned the egg to the Dendroolithidae. Insufficient documentation, therefore, prevents accurate comparison of the therizinosaurid egg to Dictyoolithus hongpoensis at this time. Nevertheless, the inclusion of Dictyoolithus and the therizinosaurid egg in Dinosauoid-spherulitic Basic Type (CitationHirsch, 1996; CitationMikhailov, 1997; CitationCarpenter, 1999) further demonstrates the paraphyly of some parataxonomic categories. CitationZelenitsky et al. (2002) suggested discontinuation of ‘Basic Types’ and ‘Morphotypes’ in fossil egg classification because these categories provide little additional information beyond the oofamily and oogenus diagnoses. We concur with this recommendation and therefore consider it unnecessary to reassign Dictyoolithidae to a Basic Type or Morphotype within egg parataxonomy. CitationCarpenter (1999) considered the Dictyoolithidae easily confused with Dendroolithidae. In our opinion, the absence of spherulitic structure and presence of closely space nuclei and prismatic columns with horizontal laminations clearly distinguish D. hongpoensis from eggs of the Dendroolithidae. Detailed descriptions and future studies using SEM, however, may clarify the microstructural characteristics of other Dictyoolithus oospecies.
CONCLUSIONS
Fossil eggs recovered from the Upper Cretaceous Chichengshan Formation in the Lishui Basin of Zhejiang Province, China, are referable Dictyoolithus hongpoensis CitationZhao, 1994. However, nucleation sites that originate at the inner surface of superimposed shell units (as interpreted by CitationZhao, 1994) are not present in the D. hongpoensis from the Lishui Basin. Furthermore, this feature is not conclusively demonstrated in radial thin sections or SEM photographs of any previously published Dictyoolithus oospecies. Therefore, the presence of this structure in dictyoolithid eggs remains questionable. The theropod nature of Dictyoolithus is supported by phylogenetic analysis and the following characters: relatively narrow spacing of nucleation centers, radiating crystals in the strictly crystalline, blocky mammillary layer, and the prismatic structure of the eggshell.
The recognition of prismatic columns and parallel laminae in the Lishui eggs resulted from the higher magnification possible with scanning electron microscopy. Therefore, we recommend the use of SEM in the study of fossil eggshell microstructure and detailed descriptions when describing fossil eggs. Finally, we concur with CitationZelenitsky et al. (2002) and advocate discontinuation of Basic Types and Morphotypes in fossil egg parataxonomy, on the basis of the paraphyletic nature of these categories.
APPENDIX 1
Egg and eggshell characters used in phylogenetic analysis. All characters are treated as unordered except numbers 2 and 18.
-
Eggshell, composition: aragonite (0); calcite (1).
-
Eggshell, nucleation centers, spacing relative to shell width: >0.40 (0); 0.33–0.40 (1); 0.24–0.33 (2); <0.24 (3) [ordered].
-
Eggshell, nucleation centers: at base of mammillary layer (0), within layer (1).
-
Eggshell, mammillary layer, composition: crystalline (0); organic and crystalline (1).
-
Eggshell, mammillary layer, radiating crystal form: truncated (0); irregular (1); blocky (2).
-
Eggshell, transition between first and second structural layers: abrupt (0); gradual (1).
-
Eggshell, second layer: absent (0); prismatic (1); wedge (2).
-
Eggshell, second layer, squamatic structure: absent (0); present (1).
-
Eggshell, second layer, squamatic structure type: regular, masks prisms (0); evenly distributed, visible prisms (1); irregular, prisms visible (2).
-
Eggshell, second layer, book-like horizontal laminations: absent (0); present (1).
-
Eggshell, transition between second and third structural layers: gradual (0); abrupt (1).
-
Eggshell, third layer: absent (0); present (1).
-
Eggshell, third layer, structure: horizontal crystals (0); vertical crystals (1); with porous and crystalline regions (2).
-
Eggshell, pore system: straight with consistent width (0); straight with enlarged base (1); straight to oblique (2); irregular and varying (3); branched (4); reticulate (5).
-
Eggshell, cuticle: absent (0); present (1).
-
Eggshell, ornamentation: absent (0); present (1).
-
Egg, shape: symmetrical (0); asymmetrical (1).
-
Egg, size, relative to adult body size: small, <0.10 Eb (0); medium, 0.10–0.30 Eb (1); large, >0.30 Eb (2) [ordered]. Eb is the mass of an egg predicted for a bird of similar adult size using an allometric equation based on modern data (CitationBlueweiss et al., 1978).
-
Clutch, arrangement: random/massed (0); eggs paired (1); open, one layer (2).
ACKNOWLEDGMENTS
We thank G. Wang and X. Zhou (Lishui Museum) for collecting the specimens, M. Ding, Y. Sheng, and H. Wu (Zhejiang Museum of Natural History) and Y. Noda (Fukui Prefectural Dinosaur Museum) for thin-section preparation. We thank R. Avci for use of the Imaging and Chemical Analysis Laboratory (ICAL) at Montana State University, Melody Bergeron for technical support, and J. Horner for use of the Gabriel Laboratory for Cellular and Molecular Paleontology in the Paleontology Department of the Museum of the Rockies. We thank J. Varricchio for translations and guiding service in China and B. Andres for logistical assistance. B. Jackson and J. Schmitt provided technical support and editorial comments. We thank two anonymous reviewers for their helpful suggestions that improved the manuscript. The College of Letters and Sciences and the Donald L. Smith Scholarship, Department of Earth Sciences, Montana State University, provided funding to FJ, and the Department of Culture and Land Resources, Zhejiang Province provided support to XJ for this project.
Related Research Data
LITERATURE CITED
- Blueweiss , L. , Fox , H. , Kudzma , V. , Nakashima , D. , Peters , R. and Sams , S. 1978 . Relationships between body size and some life history parameters . Oecologia , 37 : 257 – 272 .
- Carpenter , K. 1999 . Eggs, Nests, and Baby Dinosaurs: A Look at Dinosaur Reproduction , 336 Bloomington, , Indiana : Indiana University Press .
- Cheng , Z. , Wang , Y. and Li , G. 1995 . New advances in the study of dinosaur egg fossil strata in Xixia basin, Henan Province . Chinese Science Bulletin , 40 : 1487 – 1490 .
- Ferguson , M. 1985 . “ Reproductive biology and embryology of crocodilians; pp ” . In Biology of the Reptiles , Edited by: Billet , F. and Gans , C. Vol. 14 , 330 – 491 . New York : John Wiley & Sons .
- Grellet-Tinner , G. and Chiappe , L. M. 2004 . “ Dinosaur eggs and nesting: implications for understanding the origins of birds ” . In Feathered Dinosaurs , Edited by: Currie , P. J. , Koppelhus , E. B. , Shugar , M. A. and Wright , J. L. 184 – 214 . Bloomington, , Indiana : Indiana University Press .
- Grellet-Tinner , G. and Makovicky , P. 2006 . A possible egg of a dromaeosaur Deinonychus antirrhopus: phylogenetic and biologic implications . Canadian Journal of Earth Sciences , 43 : 705 – 719 .
- Hirsch , K. F. 1996 . Parataxonomic classification of fossil chelonian and gecko eggs . Journal of Vertebrate Paleontology , 16 : 752 – 763 .
- Hirsch , K. F. and Quinn , B. 1990 . Eggs and eggshell fragments from the Upper Cretaceous Two Medicine Formation of Montana . Journal of Vertebrate Paleontology , 10 : 491 – 511 .
- Jin , X. , Azuma , Y. , Jackson , F. D and Varricchio , D. J. 2007 . Giant dinosaur eggs from the Tiantai basin, Zhejiang Province, China . Canadian Journal of Earth Sciences , 44 : 81 – 88 .
- Liu , J. and Zhao , Z. 2004 . A new oospecies of the dinosaur eggs (Dictyoolithus) from Laiyang, Shandong, Province . Vertebrate PalAsiatica , 142 : 166 – 170 .
- Lü , J. , Jin , X. , Sheng , Y. , Li , Y. , Wang , G. and Azuma , Y. 2007 . New nodosaurid dinosaur from the Late Cretaceous of Lishui, Zhejiang Province, China . Acta Geologica Sinica , 81 : 344 – 350 .
- Manning , T. W. , Joysey , K. A. and Cruickshank , A. R. I. 1997 . “ Observations of microstructures within dinosaur eggs from Henan Province, Peoples’ Republic of China ” . In Dinofest International , Edited by: Wolberg , D. L. , Stump , E. and Rosenberg , G. D. 287 – 290 . Philadelphia, , Pennsylvania : Academy of Natural Sciences .
- Manning , T. W. , Joysey , K. A. and Cruickshank , A. R. I. 1999 . “ In ovo tooth replacement in a therizinosauroid dinosaur ” . In First International Symposium on Dinosaur Eggs and Babies, Extended Abstracts , Edited by: Bravo , A. M. and Reyes , T. 129 – 134 . Catalonia, , Spain : Isona .
- Marshall , D. 1988 . Cathodoluminescence of geological materials , 146 Boston, Massachusetts : Unwin Hyman .
- Mateus , I. , Mateus , H. , Antunes , M. T. , Mateus , O. , Tacquet , P. , Ribeiro , V. and Manuppella , G. 1997 . Couvee, oeufs et embryons d’un dinosaure theropode du Jurassique superieur de Lourinha. Les Comptes Rendus de l’Académie des Sciences , Vol. 325 , 71 – 89 . Paris. Sciences de la terre e des planètes .
- Mikhailov , K. E. 1997 . Fossil and recent eggshell in amniotic vertebrates: fine structure comparative morphology and classification . Special Papers in Palaeontology , 56 : 1 – 80 .
- Mikhailov , K. E. , Bray , E. S. and Hirsch , K. F. 1996 . Parataxonomy of fossil egg remains (Veterovata): principles and applications . Journal of Vertebrate Paleontology , 16 : 763 – 769 .
- Sereno , P. C. 2007 . Logical basis for morphological characters in phylogenetics . Cladistics , 23 : 1 – 23 .
- Swofford , D. L. 1998 . PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods). Version 4 , Sunderland, Massachusetts : Sinauer Associates .
- Varricchio , D. J. and Jackson , F. D. 2004 . A phylogenetic assessment of prismatic dinosaur eggs from the Cretaceous Two Medicine Formation of Montana . Journal of Vertebrate Paleontology , 24 : 931 – 937 .
- Varricchio , D. J. , Horner , J. R. and Jackson , F. D. 2002 . Embryos and eggs for the Cretaceous theropod dinosaur Troodon formosus . Journal of Vertebrate Paleontology , 22 : 564 – 576 .
- Wang , Y. , Jiang , Y. and Chen , K. 2000 . The study of dinosaur fossil and Late Cretaceous basin of Tiantai in Zhejiang. Unpublished file report of the Hydrology and Engineering Geological Brigade of Zhejiang Province and the Chinese Academy of Geological Sciences Available from the library of the Land and Resources Department of Zhejiang Province, China.
- Wang , Q. , Zan , S. , Jin , L. and Chen , J. 2006 . A new oospecies, Dictyoolithus gongzhulingensis, from the Early Cretaceous Quantou Formation in the Central Jilin Province . Journal of Jilin University (Earth Science Edition) , 36 : 153 – 157 .
- Zelenitsky , D. K. and Hills , L. V. 1996 . An egg clutch of Prismatoolithus levis oosp. nov. from the Oldman Formation (Upper Cretaceous), Devil's Coulee, southern Alberta . Canadian Journal of Earth Science , 33 : 1127 – 1131 .
- Zelenitsky , D. K. and Therrien , F. 2008 . Phylogenetic analysis of reproductive traits of maniraptoran theropods and its implications for egg parataxonomy . Palaeontology , 51 : 807 – 816 .
- Zelenitsky , D. K. , Modesto , S. P. and Currie , P. J. 2002 . Bird-like characteristics of troodontid theropod eggshell . Cretaceous Research , 23 : 297 – 305 .
- Zhao , Z. 1994 . “ Dinosaur eggs in China: on the structure and evolution of eggshells ” . In Dinosaur Eggs and Babies , Edited by: Carpenter , D. , Hirsch , K. F. and Horner , J. R. 184 – 203 . Cambridge : Cambridge University Press .
- Zhao , Z. K. , Ye , J. , Li , H. M. , Zhao , Z. H. and Yan , Z. 1991 . Extinction of the dinosaurs across the Cretaceous-Tertiary boundary in Nanxiong basin, Guangdong Province . Vertebrata PalAsiatica , 29 : 1 – 20 .