ABSTRACT
A new species of bothriodontine anthracothere, Arretotherium meridionale, is described from the early Miocene (Arikareean North America Land Mammal Age) Las Cascadas fossil assemblage in Panama, Central America. Fossils of A. meridionale are the first record of an anthracothere from the New World Tropics. Among anthracotheres, A. meridionale is most similar to A. acridens from the middle Arikareean from Texas in having a relatively deep and robust jaw, high and sharp cusps on the lower molars, short c–p1 diastema, and absence of a mesiolingual metacristid. A. meridionale differs from other species in being generally larger, two lower incisors (rather than three), prehypocristid never reaching the postprotocristid, more apical junction between postprotocristid and postmetacristid, mesiolingual entocristid transversely notching preentocrisitid, and transverse valley tapered lingually by prehypocristid. Although cladistic analysis of 28 anthracotheriids coded for 51 characters supports a relationship between A. meridionale and A. acridens, some presumably convergent dental characteristics are also similar to certain Oligocene-Miocene Eurasian bothriodontines. The presence of Arretotherium in the Las Cascadas Formation in Panama, and absence in the later Centenario Fauna, shows that primitive bothriodontines entered into Central America by the early Miocene before disappearing from the New World during the late early–middle Miocene.
SUPPLEMENTAL DATA—Supplemental materials are available for this article for free at www.tandfonline.com/UJVP
INTRODUCTION
Anthracotheriid artiodactlys likely split from helohyid artiodactyls (Coombs and Coombs, Citation1977; Ducrocq et al., Citation1997) in Eurasia (Suteethorn et al., Citation1988; Lihoreau and Ducrocq, Citation2007) before representatives of this family dispersed into North America in the Eocene (Douglass, Citation1901; Troxell, Citation1921; Scott, Citation1940; Macdonald, Citation1956; Macdonald and Shultz, Citation1956). Their North American fossil record is restricted to the late middle Eocene to middle Miocene of the Great Plains, with a few Oligocene and Miocene records from further south in Texas (Kron and Manning, Citation1998; Albright, Citation1999; Tedford et al., Citation2004).
North American anthracotheres are classified in two subfamilies (Scott, Citation1940; Kron and Manning, Citation1998): Anthracotheriinae Leidy, Citation1869, represented by the genus Heptacodon Marsh, Citation1894a, from the Whitneyan of North America, and Bothriodontinae Scott, Citation1940, which includes Aepinacodon Troxell, Citation1921, from the late Eocene of South Dakota and Nebraska (middle to late Chadronian), Arretotherium Douglass, Citation1901 (), ranging from the late Oligocene–early Miocene of North America (early Arikareean–early Hemingfordian), Kukusepasutanka Macdonald, Citation1956, from the late Arikareean deposits of Montana (Macdonald, Citation1956), and Bothriodon Aymard, Citation1846, from the Brule Formation in South Dakota (Orellan to early Arikareean) and from the early Chadronian of Saskatchewan (Kron and Manning, Citation1998). Bothriodontines, characterized by the presence of more selenodont molars, a relatively shorter rostrum, upper molars with four or five cusps, and the upper molar mesostyle being variably invaded by the transverse valley (Scott, Citation1940; Macdonald, Citation1956; Kron and Manning, Citation1998; Lihoreau and Ducrocq, Citation2007), have been recognized as possibly sharing a relationship with Hippopotamidae (Simpson, Citation1945; Boisserie and Lihoreau, Citation2006; Boisserie et al., Citation2010, Citation2011). They dispersed widely during the early Neogene, reaching all continents except Australia by the end of the Neogene. Although North American bothriodontines were probably derived from a primitive Eurasian form, such as Elomeryx or Bothriodon (Scott, Citation1940; Macdonald, Citation1956; Macdonald and Shultz, Citation1956; Lihoreau and Ducrocq, Citation2007), distinguishing between Old and New World bothriodontines has been one of the primary difficulties in clarifying the relationships between American forms and their Eurasian counterparts (Kron and Manning, Citation1998; Lihoreau and Ducrocq, Citation2007).
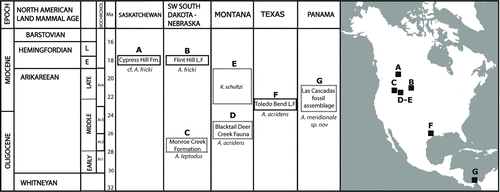
FIGURE 1 Location and biochronology of early Miocene bothriodotines discussed in this study. A, cf. Arretotherium fricki from the early Hemingfordian Cypress Hill Formation in Saskatchewan (Storer and Bryant, Citation1993); B, A. fricki Macdonald and Shultz, Citation1956, from the early Hemingfordian Upper Marsland Formation in Nebraska and also present in the Hemingfordian Flint Hill L. F. from South Dakota (Macdonald and Shultz, Citation1956; Macdonald and Martin, Citation1987); C, A. leptodus (= Ancodon leptodus) Matthew, Citation1909, from the early Arikareean Lower Rosebud beds, South Dakota (Scott, Citation1940; Macdonald, Citation1956; Macdonald, Citation1963; Tedford et al., Citation2004); D, A. acridens Douglass, Citation1901, from the early Arikarrean Blacktail Deer Creek Fauna., Montana (Hibbard and Keenmon, Citation1950); E, Kukusepasutanka Macdonald, Citation1956, from the late Arikareean deposits of Montana (Macdonald, Citation1956); F, Arretotherium acridens Douglass, Citation1901, from the early Miocene Toledo Bend L. F., Texas (Albright, Citation1999); G, A. meridionale, sp. nov., from the Las Cascadas fossil assemblage, Panama. Chronostratigraphy and biochronology modified from Albright et al. (Citation2008). Abbreviations: Ar, Arikareean Faunal Zone; E, early; L, late; L. F., Local Fauna.
The early Miocene bothriodontines Arretotherium and Kukusepasutanka are the last-known North American anthracotheres (). Although they are only known from a few specimens, these taxa are similar in lacking a paraconule on the upper molars; however, Kukusepasutanka is larger, has a more invaded mesostyle, and has more strongly developed stylar cusps (Macdonald, Citation1956). The only known species, K. shultzi, was recovered from early Miocene sediments in the Drummond Granite County in Montana. K. shultzi is characterized by an elongated and massive tubular snout and the presence of well-developed anterior diastema (Macdonald, Citation1956; Kron and Manning, Citation1998). Although no lower dentition of K. shultzi has been recovered, making its relationship with Arretotherium unclear, it is similar to Heptacodon in its development of stylar cusps on the upper molars. In contrast to K. schultzi, Arretotherium has a relatively short snout and a more robust mandible (Macdonald, Citation1956). Distinguishing characteristics originally noted by Douglass (Citation1901) for Arretotherium and further discussed by Macdonald (Citation1956) include (1) four-cusped upper molars; (2) teeth with sharp cusps or crescents; (3) small first upper premolar; (4) upper premolars increasing rapidly in size posteriorly, composed of a simple cone and a cingulum-like cusp; (5) upper molars high and square with heavy cinguli; (6) upper canine laterally compressed; and (7) upper premolars reduced, with protocone not connected labially to cingulum.
Despite the fact that only a few fossils have been referred to Arretotherium, there are three described species: (1) A. acridens Douglass, Citation1901 (type species), from the early Arikareean Blacktail Deer Creek Fauna in Montana (Douglass, Citation1901; Hibbard and Keenmon, Citation1950), thought to be equivalent to the Harrison Formation in Nebraska (Tedford et al., Citation1987; Tedford et al., Citation2004), and the middle to late Arikareean Toledo Bend Local Fauna (L. F.) in Texas (Albright, Citation1998, Citation1999); (2) A. leptodus (= Ancodon leptodus) Matthew, Citation1909, from the early Arikareean Lower Rosebud beds that overly the White River Group in South Dakota and is presumably the oldest record of this genus (Scott, Citation1940; Macdonald, Citation1956; Tedford et al., Citation2004); and (3) A. fricki Macdonald and Shultz, Citation1956, described from cranial material from the early Hemingfordian Runningwater Formation in Nebraska, and is also present in the Hemingfordian Flint Hill L. F. from South Dakota (Macdonald and Martin, Citation1987) and the early Hemingfordian Cypress Hill Formation in Saskatchewan (Storer and Bryant, Citation1993). A. acridens and A. leptodus from the early to middle Arikareean are characterized by a heavier, more robust skull and a deeper mandible than that of A. fricki. In contrast, although the Hemingfordian A. fricki might be derived from A. acridens (Macdonald and Martin, Citation1987), it is characterized by a more slender and gracile cranium (Kron and Manning,
To date, there is no agreement about the relationships between the North American early Miocene forms, which has caused several taxonomic problems. For example, there is an overlap between the defining molar characteristics of A. acridens and A. leptodus. Macdonald (Citation1956) discussed in detail this matter and suggested that A. leptodus could represent a nomen vanum because of the poorly preserved holotype. However, Macdonald later abandoned his argument, ratifying the validity of this taxonomic entity, but transferring this taxon to the genus Arretotherium (A. leptodus Macdonald, Citation1963) based solely on its stratigraphic position and the probable age of the fossil assemblage (Hibbard and Keenmon, Citation1950; Macdonald, Citation1963). Disregarding the poorly understood relationship between A. leptodus and A. acridens, A. fricki seems to represent the last appearance of Anthracotheriidae () in the fossil record of North America (Macdonald and Martin, Citation1987). Although it is beyond the scope of the present study, a more detailed analysis of the American Anthracotheriidae is clearly needed to further clarify the present taxonomic uncertainties associated with this family.
Herein, we describe a new American bothridontine, Arretotherium meridionale, sp. nov., from the upper part of the volcanoclastic early Miocene Las Cascadas Formation from the Panama Canal area (Gaillard Cut) in Central America (). The new fossils represent the southernmost occurrence of Bothriodontinae, suggesting that it reached tropical habitats before its extinction in North America during the middle Miocene (Kron and Manning, Citation1998; Lihoreau and Ducrocq, Citation2007).
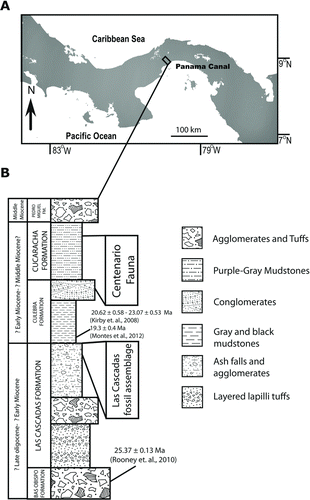
FIGURE 2 Location and stratigraphic position of fossil bothriodontines from the Gaillard Cut, Panama Canal area. A, map of North and Central America showing the location of the Panama Canal area and the Gaillard Cut. B, stratigraphic position of the Las Cascadas fossil assemblage and the Centenario Fauna. Modified from Kirby et al. (Citation2008), MacFadden et al. (Citation2010), and Montes et al. (Citation2012).
Abbreviations
The biochronology follows late Oligocene–
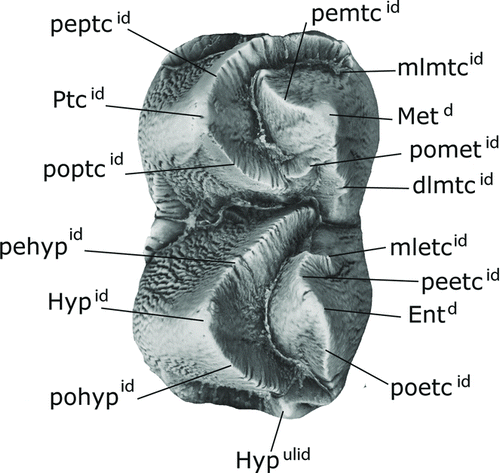
early Miocene biozonation developed in the Great Plains (Tedford et al., Citation1987, 2004; MacFadden and Hunt, Citation1998) and the subsequent recalibration proposed by Albright et al. (Citation2008) for the Arikareean North America Land Mammal Age (NALMA). For the most part, dental nomenclature () follows Lihoreau and Ducrocq (Citation2007). We also use the descriptive system of crests and crescents defined by Loomis (Citation1925) for buno-selenodont dentitions in artiodactyls. Additionally, we follow the tooth orientation terminology proposed by Smith and Dodson (Citation2003) for the incisors.
FIGURE 3 Diagram showing most of the dental cusp nomenclature applied to lower permanent dentitions of anthracotheres in this study. Abbreviations: dlmtcid , distolingal metacristid; Entd , entoconid; Hypd , hypoconid; Hypulid , hypoconulid; mletcid , mesiolingual entoconid; mlmtcid , mesiolingual metacristid; Metd , metaconid; poetcid , postentoconulid; pohypid , posthypoconid; pometid , postmetacristid; poptcid , postprotocristid; peetcid , preentocristid; pehypid , prehypocristid; pemtcid , premetacristid; peptcid , preprotocristid; Ptcd , protoconid. Modified from Lihoreau and Ducrocq (Citation2007).
Institutional Abbreviations
AMNH, American Museum of Natural History, New York, U.S.A; CM, Carnegie Museum, Pittsburgh, Pennsylvania, U.S.A.; F:AM, Frick: American Mammals collection at the AMNH; LSUMG-V, Louisiana State University Museum of Geoscience; Baton Rouge, Louisiana, U.S.A; SDSM, South Dakota School of Mines, Rapid City, South Dakota, U.S.A; UCMP, University of California Museum of Paleontology, Berkeley, California, U.S.A; UF, Vertebrate Paleontology Collection, Florida Museum of Natural History (FLMNH), University of Florida, Gainesville, Florida, U.S.A; UNSM, University of Nebraska State Museum, Lincoln, U.S.A.
AGE AND GEOLOGIC SETTING
The early Miocene Las Cascadas Formation represents the oldest continental sequence cropping out along the Gaillard Cut, Panama Canal area (, B). The basal part of the Las Cascadas Formation unconformably overlies the welded and massive marine to shallow marine volcanic agglomerates of the middle to late Oligocene Bas Obispo Formation (Woodring and Thompson, Citation1949; Woodring, Citation1957, Citation1982; Kirby et al., Citation2008; Montes et al., Citation2012). Although the thickness of the Las Cascadas Formation is still unknown, it is composed of a lower member with massive agglomerates and an upper member that includes paleosols, ash falls, lenticular bodies of conglomeratic sandstones, and purple to green mudstones interbedded with coarser-grained volcanic agglomerates. Fossil-bearing lithologies are restricted to local fluvial sequences in the uppermost part of the Las Cascadas Formation. The fossils described here were found in the northern part of the Gaillard Cut and were associated with other ungulates, including protoceratids, equids, peccaries, and camelids (Rincon et al., Citation2010, 2012). Although the better-preserved fossils are restricted to the fine-grained ash falls, additional fossils were also recovered from an overlying conglomeratic welded tuff.
Although the shallow marine Culebra Formation unconformably overlies the upper part of Las Cascadas Formation in the northern part of the Gaillard Cut (Montes et al., Citation2012), intense faulting and deformation obscures this contact in the Las Cascadas fossil locality (Lirio Norte). Fossil vertebrates from the Gaillard Cut (Whitmore and Stewart, Citation1965; Woodring, Citation1957, Citation1982; MacFadden and Higgins, Citation2004; MacFadden, Citation2006, Citation2009; MacFadden et al., Citation2010; Uhen et al., Citation2010) have been formally grouped into the Centenario Fauna (MacFadden et al., Citation2010). This fauna extends from the uppermost part of the Culebra Formation and includes the transitional to continental Cucaracha Formation (), representing about 120 m of fossil-bearing sediments that are widely distributed along the Gaillard Cut (MacFadden et al., Citation2010). The Centenario Fauna includes mammalian taxa known from the Arikareean to the Barstovian of North America, but does not obviously fit within the biochronological context of a specific Miocene NALMA (MacFadden and Higgins, Citation2004; MacFadden, Citation2006, Citation2009; MacFadden et al., Citation2010). Attempts to independently determine the age of the Centenario Fauna using geochronological analyses are currently in progress.
Despite the absence of a reliable isotopic date, the age of the upper part of the Las Cascadas Formation can be tentatively constrained through its stratigraphic relationships with other formations from the Gaillard Cut (). Consequently, the upper Las Cascadas fossiliferous sequence represents an interval older than the lower part of the Culebra Formation, for which several dates have been published, including 20.62 ± 0.58, 23.07 ± 0.53 (Kirby et al., Citation2008), and 19.3 ± 0.4 (Montes et al., Citation2012) Ma. The lower boundary of the Las Cascadas Formation can also be constrained using an Ar40/Ar39 age of 25.37 ± 0.13 Ma derived from the andesitic water-saturated arc lavas of the underlying Bas Obispo Formation (Rooney et al., Citation2010). Therefore, the duration of the Las Cascadas Formation may span the late Oligocene to early Miocene (<25 to ∼19 Ma), likely representing the middle to late Arikareean NALMA sensu MacFadden and Hunt (Citation1998), Tedford et al. (Citation2004), and Albright et al. (Citation2008). Based on the stratigraphic position and available biochronology, fossil vertebrates from the Las Cascadas fossil assemblage likely represent the late Arikareean and represent the oldest mammalian fauna so far found in the Gaillard Cut, indicating the arrival of northern populations into a recently emerged continental area (Rincon et al., Citation2012).
SYSTEMATIC PALEONTOLOGY
Class MAMMALIA Linnaeus, Citation1758
Order ARTIODACTYLA Owen, Citation1848
Family ANTHRACOTHERIIDAE Leidy, Citation1869
Subfamily BOTHRIODONTINAE Scott, Citation1940
ARRETOTHERIUM Douglass, Citation1901
Type Species
Arretotherium acridens (Douglass, Citation1901)
Holotype
CM 704. Right and left partial maxillae, with RI1–P2, M1–M3, and isolated LP3–P4 and LM1–M2.
Generic Diagnosis
Short rostrum without diastema, upper cheek teeth as in Elomeryx except molars have lost protoconule, cusps of upper and lower cheek teeth extremely high and pointed (Macdonald, Citation1956).
Included Species
Arretotherium acridens Douglass, Citation1901, from the early Arikareean Blacktail Deer Creek from Montana (Douglass, Citation1901; Hibbard and Keenmon, Citation1950; Tedford et al., Citation2004) and the middle to late Arikareean Toledo Bend L. F. in Texas (Albright, Citation1999); A. leptodus (= Ancodon leptodus) Matthew, Citation1909, from the early Arikareean Lower Rosebud beds in South Dakota; A. fricki Macdonald and Shultz, Citation1956, from the early Hemingfordian Upper ‘Marsland Formation’ in Nebraska, the Hemingfordian Flint Hill L. F. from South Dakota (Macdonald and Shultz, Citation1956; Macdonald and Martin, Citation1987), and the early Hemingfordian Cypress Hill Formation in Saskatchewan (Storer and Bryant, Citation1993); A. meridionale, sp. nov., from the Las Cascadas fossil assemblage in Panama.
Remarks
Although only a single partial upper molar has been referred to Arretotherium meridionale, sp. nov. (see below), it lacks the distinctive strongly developed stylar cusps and deeply invaded mesostyle considered diagnostic for the similarly sized monospecific genus Kukusepasutanka (Macdonald, Citation1956:fig. 16). In all other comparable features, A. meridionale, sp. nov., is most similar to species classified in Arretotherium (see also Douglass, Citation1901; Scott, Citation1940; Macdonald, Citation1956; Macdonald and Shultz, Citation1956; Kron and Manning, Citation1998; Albright, Citation1999; Lihoreau and Ducrocq, Citation2007). Although description of this new taxon could result in an expanded concept of Arretotherium, we are reluctant to emend a generic diagnosis that is based solely on the morphology of the upper dentition (Macdonald, Citation1956) without more complete fossils of A. meridionale, sp. nov.
ARRETOTHERIUM MERIDIONALE, sp. nov.
(–; )
Holotype
UF 244187, juvenile left dentary with associated deciduous canine, partially erupted c1, p1, dp2–dp4, m1–m2, and associated right i1–i2 and mandibular symphysis.
Locality and Horizon
Lirio Norte (site key YPA-024 in University of Florida Vertebrate Paleontology Collection), Panama Canal area, Panama, Central America (). Fossils were collected in the upper part of the Las Cascadas Formation (), likely equivalent to the late Arikareean NALMA (Rincon et al., 2012).
Etymology
From the Latin ‘meridionale’ (southern). Corresponding to the southernmost record of Anthracotheriidae in the New World.
Referred Material
The following were found in association with the holotype: UF 245609, right upper incisor; UF 244295, left upper C1 (broken); UF 244181, right astragalus; and UF 244290, distal right metacarpal. Additionally, the following specimens were found in association in a different horizon of the same locality: UF 244174, partial left M3; UF 246821, partial skull fragment; and UF 244177, right distal femur. Although it is possible that associated specimens are from the same individuals (minimum number of individuals = 2), the lack of any sort of articulation makes this ambiguous and they are thus cataloged separately.
Diagnosis
Differs from all other species of Arretotherium in having overall larger size, two lower incisors, lower molars with prehypocristid never reaching postprotocristid, more apical junction between postprotocristid and postmetacristid, mesiolingual entocristid transversely notching preentocrisitid, and transverse valley lingually tapered by prehypocristid. Further differs from A. acridens in having incisors with stronger and more cylindrical roots and an upper canine with a serrated anterior edge pinched into a carina. Further differs from A. fricki in having a deeper and more robust jaw, and more sloping mandibular symphysis. Further differs from A. leptodus in having a more laterally compressed upper canine.
Description
Mandible
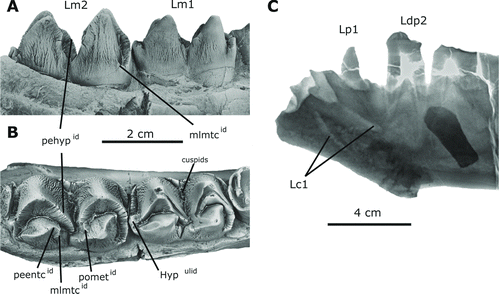
A single juvenile partial dentary is referred to Arretotherium meridionale, sp. nov., from the Las Cascadas Formation fossil assemblage (UF 244187, holotype). The mandible is deep, with short diastemata in front of, and behind, the p1 (–C). The anterior part is partially broken but exhibits two preserved incisor alveoli. The jaw is relatively narrow and deep, with no evidence of constriction at the level of the canine. The mandible posterior to the m2 is not preserved in this specimen. The labial surface of the horizontal ramus is uniformly convex along the jaw, whereas the lingual segment is convex below the molars and concave anterior to the dp3 (). An attenuated transverse constriction is present between the p1 and the dp2 and coincides with the position of the anterior mental foramen (). A second smaller posterior foramen is located just below the anterior root of the dp3. The anterolingual surface of the mandible is characterized by a relatively straight and oblique symphysis that ends ventrally with a distinct genial spine that extends posteriorly from the posteroventral end of the symphysis (, B). Both the ventral and dorsal borders of the symphysis are subparallel and slightly convex dorsally. The symphysis has a broader anterior segment that ends in a flattened ventral surface just below the genial spine. The symphysis is broad, long, and V-shaped in cross-section, with no evidence of fusion. A short diastema is present between the single-rooted p1 and the dp2. A relatively well-developed vascular groove is partially preserved at the posterior ventral end of the ramus, corresponding to a change in curvature of the ventral mandibular notch (, C).
FIGURE 4 Lower dentition of Arretotherium meridionale, sp. nov. A, UF 244187, juvenile left dentary with left dc1, partially erupted left canine, p1, dp2–dp4, m1–m2, and associated right i1–i2 and mandibular symphysis, occlusal view; B, lingual view; C, labial view; D, associated right i1, lingual view; E, labial view; F, associated right i2, lingual view; G, labial view; H, detailed view of the anterior part of UF 244187 showing the erupting Lc1. Abbreviations: pehypid , prehypocristid; postlgF, posterolingual furrow; antlgF, anterolingual furrow.
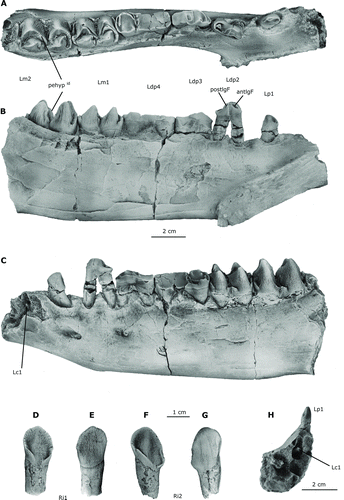
FIGURE 5 Detailed view of the lower permanent dentition of Arretotherium meridionale, sp. nov. A, UF 244187, left m1–m2, lingual view; B, occlusal view; C, X-ray detailed image of the anterior part of UF 244187 showing the erupting permanent Lc1. Abbreviations: pehypid , prehypocristid; mlmtcid , mesiolingual metacristid; peetcid , preentocristid; pometid , postmetacristid; hypulid , hypoconulid.
Lower Deciduous Dentition
The preserved deciduous dentition includes the left dc1 and dp2–dp4 (). The crown of the dc1 is very worn. It is elongated and posteriorly recurved. The posterior part of crown has an elongated constriction that is evident even in intense wear stages. The dp2 is double-rooted (–C). The crown of dp2 consists of an anterior crest, a large primary cusp, and a relatively elongated posterior crest with a small but distinct cingular segment enclosing a small, conical cuspule. These structures form the talonid of the tooth and the cingulid extends from the posterior end of the principal cusp along the labial and the lingual segment. Two small furrows are present lingual to the principal cusp; the anterior is deeper and elongated, whereas the posterior is shorter (). Despite intense wear, an additional anterior cusp is evident, occupying the position of the paraconulid, and an anterior crest connects this cusp with the primary cusp (paraconid). The paraconid is more separated from the posterior crest, resulting in a trenchant overall shape with a higher paraconid. The crown of dp3 is longer and larger than that of dp2 but preserves a similar trenchant morphology. The anterior crest is well developed and proportionally longer than that of the dp2. A lingually opened invagination of the enamel is present along the anterolingual segment of the paraconid (). A deep enamel valley separates the principal cusp from the posterior crescent, which is completely worn and exposes the posterior root below the cervix (enamel-dentine junction). The anterior crest is high, and, like that of dp2, has an anterolingual ridge located in the position of the paraconulid. The crown of dp4 is molariform and trilobed. The anterior lobe is similar in size to the intermediate lobe and both are slightly narrower than the posterior lobe (). The tooth is worn almost to the base of the crown, especially its anterior-most lobe. The anterior-posterior length of the dp4 exceeds that of m1 or m2. Even though intensive wear is present, a fully selenodont posterior lobe is perceptible. A distinctive cingular segment forms the posterolingual part of the dp4. The metaconid is separated from the entoconid by a lingual notch that is visible even in advanced wear stages. A strong, shelf-like cingulid is evident anterior to the hypoconid in the labial side (, C). The presence of a metastylid could not be evaluated due to strong wear on the lingual segment of the dp4.
Lower Permanent Dentition
The permanent dentition includes Ri1–2, Lp1, and Lm1–m2 (UF 244187). Additionally, two incisor alveoli are preserved in the anterior part of the left dentary. Both incisors have long, deep cylindrical roots and procumbent broad and acutely pointed crowns (–G). The second incisor is considerably larger than the first (), and both are characterized by having a spatulate appearance. The basal part of the crown of i2 is extremely elongated, suggesting that it was located more distally than the smaller i1 (, F). The lingual surface of each incisor is concave, the labial surface is convex, and both surfaces are heavily crenulated (, G). There is a distinctive cingular segment along the lingual surface at the enamel-dentine junction, which is more developed on i2 than on i1 (, F).
TABLE 1 Summary of dental measurements (in mm) of A. meridionale, sp. nov., from the Las Cascadas Formation.
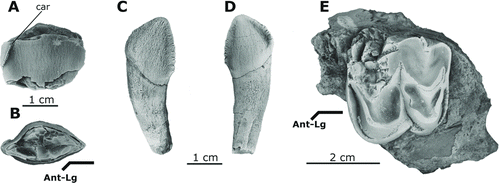
FIGURE 6 Upper dentition of Arretotherium meridionale, sp. nov. A, UF 244295, left upper canine, lingual view; B, occlusal view; C, UF 245609, right upper incisor (Ix), labial view; D, lingual view; E, UF 244174, partial left M3, occlusal view.
A partially erupted canine is replacing an extremely worn deciduous canine in the holotype (). Although c1 is only partially erupted, an X-ray image () shows it to be large and robust, which is characteristic of male anthracotheres. The crown of p1 is small and single-rooted and is isolated by short anterior and posterior diastemata. The p1 is oval, recurved, and has a well-defined main cusp that projects posteriorly. Two separate ridges extend from the crown apex. The anterior ridge is lingually inflected and extends lingually from the apex toward the cervix forming a small furrow (–C). In contrast, the posterior ridge extends uniformly along the posterior edge of the crown until it reaches the cervix. This ridge defines two small posterior concavities located along the lingual and labial sides of the tooth that extend upwards from the cervix (, B).
The lower molars of A. meridionale are bunoselenodont, with four somewhat sectorial, well-defined, high, sharply pointed, and strongly crenulated cusps. The two external cusps are crescentic and the two internal ones relatively conical (, C and 5A, B). The apex of each transverse pair of cusps is widely separated, similar to those of the longitudinal pairs. The lingual and labial cusps have almost the same transverse width in occlusal view (). The anterior and posterior lingual basal segments of each molar have strongly crenulated anterolabial cingulids that are more developed on the labial side, but incomplete lingually (). The lower molars increase in size posteriorly, with m1 smaller than m2 (). The lower molars have a metaconid that is high and conical and is surrounded by a labial anterior continuous crest (preprotocristid) that reaches the lingual margin of the tooth (, C). A premetacristid extends from the summit of the metaconid and reaches the preprotocristid at its midpoint, whereas a small postmetacristid extends to the postprotocristid, blocking the anterior longitudinal valley (). The mesiolingual metacristid is very reduced and restricted to the basal part of the crest, making the anterior portion of the metaconid conical in shape (). The protoconid is selenodont with two cristids. The preprotocristid reaches the lingual part of the lower molar crowns, whereas the postprotocristid reaches the postmetacristid, forming a distinctive shallow longitudinal valley apically isolated from the main transversely oriented valley. The labial opening of the transverse valley is blocked by a well-defined cingular segment formed by the junction of small cuspids issuing from the hypoconid and protoconid. Small cuspids are partially blocking the lingual part of transverse valley
().
The lower molars of A. meridionale are composed of a well-defined selenodont hypoconid located posterior to the transverse valley, a relatively elongated entoconid, and a relatively high and conical hypoconulid located distally between the postentocristid and the posthypocristid. Two main cristids are attached to the entoconid. The preentocristid is continuous and well defined close to the summit of the entoconid, but diverges into two anterior cristids, the mesiolingual entocristid and the preentocristid, at the same level of the junction between the postprotocristid and the postmetacristid in the anterior part of the molar. This divergence is responsible for a distinctive notch anterior to the entoconid that is clearly visible in the lingual view (, B). The hypoconid is selenodont. The prehypocristid runs anteriorly but it does not reach the postprotocristid, partially closing the transverse valley at its base. The postentocristid stretches posteriorly and does not reach the posthypocristid, leaving the posterior longitudinal valley opened. Both cristids are separated by a conical and small hypoconulid. A small posthypocristulid runs labially from the hypoconulid, redirecting the posterior longitudinal valley labially ().
Upper Dentition
A partial C1 (UF 244295) and a right upper incisor (UF 245609) were found in association with the dentary (UF 244187) and might be the same individual (–D). The C1 is robust and transversely compressed into the shape of a flattened cone. Despite lacking the apical part of the tooth, the lingual side is relatively flat, whereas the labial side is more convex. The posterior part of C1 is more constricted (), with an edge that is pinched lingually into a carina () that leads to the concave anterolingual side of the tooth. The lingual side anterior to the carina is convex (). The posterior basal part of the carina has small cuspules, giving it a serrated appearance (). The right upper incisor (UF 245609) is caniniform and has strongly crenulated enamel (, D). It has a distinctive serrated pattern along the mesial and distal edges. The crown and the deep cylindrical roots are mesially recurved. There is no noticeable lingual cingulum on the tooth ().
An isolated LM3 (UF 244174) has a square-shaped crown in occlusal view () and is posteriorly transversely reduced. The crown is heavily worn and its anterior crest is badly broken. The protocone and metaconule are crescentic and almost completely worn. A strong cingulum surrounds the protocone, whereas a weaker but strongly crenulated posterolingual cingulum encircles the posterior part of the metaconule. The lingual opening of the transverse valley is characterized by a stronger cingulum, resulting from the connection of the anterior and posterior cingula ().
Discussion and Comparisons
Although there are some dissimilarities, the well-preserved lower molars of A. meridionale clearly resemble those of A. acridens (LSUMG V-2270) from the Toledo Bend L. F., suggesting a close relationship with this late middle Arikareean form. They share several characteristics, such as a relatively deep and strong jaw morphology, four high and sharp cusps in the lower molars, the absence of a mesiolingual metacristid, the presence of a relatively short c–p1 diastema, the presence of a small hypoconulid on m1 and m2, and the presence of spurs blocking the labial segment of the transverse valley. The general morphology of the cingula on the M3 resembles the robust morphology present in the holotype of A. acridens from Montana (Douglass, Citation1901:pl. IX), whereas this cingulum is more reduced in A. acridens from the Toledo Bend L. F. (LSUMG V-2269; Albright, Citation1999:52). The overall shape of the partial upper canine of A. meridionale (, B) is also similar to that of the holotype of A. acridens (Douglass, Citation1901:270) and A. acridens (LSUMG V-2351) from the Toledo Bend L. F., only differing in the presence of a lingually inflected carina and the absence of convexities on the labial surface of the canine. The upper incisor (, D) closely resembles incisors referred to Elomeryx crispus crispus (Hellmund, Citation1991:pl. 2) in having a serrated distal ridge and generally mesially recurved shape.
The partially preserved dentition of A. meridionale has a number of diagnostic bothriondontine characters, including (1) a transversely compressed C1 with a posterior serrated margin; (2) a shelf-like lingual cingula on upper molars; (3) distinctly crescentic cusps on upper and lower molars (Scott, Citation1940; Macdonald, Citation1956; Macdonald and Shultz, Citation1956; Macdonald, Citation1963); and (4) reduced hypoconulids on m1–m2 (Kron and Manning, Citation1998). Additionally, it shares characteristic features with Arretotherium that include (1) lower cheek teeth with high and sharp cusps; (2) robust jaw; (3) reduced anterior diastemata; (4) selenodont cristids associated with the protoconid; (5) absence of a premetacristid; (6) a relatively deep transverse valley partially blocked by spurs from the hypoconid and protoconid; (7) anterior and posterior cingulids restricted to the labial part of the metaconid and entoconid; (8) lingual outlet of the transverse valley tapered by the junction of cingulids issued to the metaconid and entoconid; (9) single-rooted p1; (10) overall morphology of the mandibular symphysis; and (11) strongly crenulated enamel in the permanent dentition. Even though there is overlap in the dimensions of the molars of A. acridens and A. leptodus (Douglass, Citation1901; Macdonald, Citation1963; Albright, Citation1999), there are enough differences between these two forms and the molar dimensions of A. meridionale
() to warrant a specific differentiation.
TABLE 2 Summary table of the anterior–posterior length (APL) and maximum transverse width (MTW) of m1 and M3 for relevant specimens of Arretotherium.
The general morphology of the lower deciduous dentition of A. meridionale does not differ substantially from the juvenile dentition referred to from Elomeryx armatus Marsh, Citation1894b (SDSM 4084; Macdonald, Citation1956). However, compared with E. armatus (SDSM 4084), A. meridionale has a distinctly more reduced p1 and more selenodont lower molars, with no mesiolingual metacristid on the m1 (Macdonald, Citation1956:626).
PHYLOGENETIC ANALYSIS
To evaluate the phylogenetic relationships of Arretotherium meridionale, sp. nov., within a sample of Anthracotheriidae, we performed a cladistic analysis of 28 anthracotheriid taxa, with the most primitive anthracothere, Siamotherium krabiense Suteerthorn et al., 1988, as the outgroup (Lihoreau and Ducrocq, Citation2007). Fifty-one dental and mandibular characters (Appendix 1) were scored and used in the analysis using a revised version of a previously published matrix (Lihoreau and Ducrocq, Citation2007:table 7.1). In our analysis (Appendix 2), Bothriogenys fraasi Schmidt, Citation1913, Bothriodon velaunus von Meyer, Citation1832, Elomeryx crispus Gervais, Citation1849, and Elomeryx armatus Marsh, Citation1894b, are scored as 1 (instead of 0) for character 20 (number of cristules issued from the metaconule) and Libycosaurus petrocchii Bonarelli, Citation1947, is scored as 1 (instead of 2) for character 15, which only presents two states: three roots for P4 (0) and fused roots of P4 (1) (Lihoreau, pers. comm., August, 2011).
The ingroup () includes taxa classified in three anthracotheriidae subfamilies: (1) Anthracotheriinae Leidy, Citation1869, is represented by five species; (2) Microbunodontinae Lihoreau and Ducroqc, Citation2007, is represented by four species; and (3) Bothriodontinae is represented by 19 species. To assess the relationship between Arretotherium meridionale and A. acridens from the Toledo Bend L. F. (Albright, Citation1999), A. acridens from Texas was scored as a distinct taxon from A. acridens Douglass, Citation1901, reported by Lihoreau and Ducrocq (2007). We believe that the inclusion of this Gulf Coast species in our preliminary analysis not only provides a test for the position of the new species from Panama, but also offers an opportunity to evaluate its specific designation and its relationship to the A. acridens holotype from Montana.
FIGURE 7 Biogeographic distribution and hypothesized phylogenetic relationships (calculated in PAUP 4.0b10) of Anthracotheriidae including Arretotherium meridionale, sp. nov., and A. acridens from the Toledo Bend L. F. (Albright, Citation1999). Data matrix includes 51 characters and 28 ingroup taxa; Siamotherium krabinese as the outgroup (modified from Lihoreau and Ducrocq, Citation2007). Pictured here is the strict consensus tree of the six most parsimonious trees resulted from the equally weighted branch-and-bound search (tree length = 120; CI = 0.583, RI = 0.829, HI = 0.41). At each node (bold numbers), the supporting unambiguous synapomorphies are 1, Antracotheriidae (20[1]); 2, Anthracotheriinae (7[1], 19[1]); 3, (18[1], 25[1], 35[1]); 4, (36[1]); 5, (16[1], 34[1], 38[1]); 6, Microbunodontinae (5[1], 6[1], 33[1]); 7, (26[1], 27[1], 32[1], 39[1], 40[1]; 8, (20[0], 37[1]); 9, Bothriodontinae (6[2], 17[1], 23[1], 39[2]); 10, (26[1], 27[1]); 11, (1[2], 2[2], 3[1], 4[2], 14[1], 20[0], 28[1], 44[2]); 12, (17[2], 18[1], 29[1], 35[1]); 13, (4[1], 33[1]); 14, (6[0], 7[1], 8[3], 24[1], 38[2], 39[1]); 15, (12[1]); 16, (20[0], 45[1]); 17, (4[1], 18[0]); 18, (6[1], 24[0]); 19, (11[1], 14[1], 23[0], 35[0], 44[1]); 20, (13[1], 21[1]); 21, (41[1]); 22, (18[0], 28[1]); 23, (39[0]); 24, (17[3], 40[2]); 25, (1[1], 9[2], 10[1], 11[2], 30[1], 31[1], 42[1], 49[1].
![FIGURE 7 Biogeographic distribution and hypothesized phylogenetic relationships (calculated in PAUP 4.0b10) of Anthracotheriidae including Arretotherium meridionale, sp. nov., and A. acridens from the Toledo Bend L. F. (Albright, Citation1999). Data matrix includes 51 characters and 28 ingroup taxa; Siamotherium krabinese as the outgroup (modified from Lihoreau and Ducrocq, Citation2007). Pictured here is the strict consensus tree of the six most parsimonious trees resulted from the equally weighted branch-and-bound search (tree length = 120; CI = 0.583, RI = 0.829, HI = 0.41). At each node (bold numbers), the supporting unambiguous synapomorphies are 1, Antracotheriidae (20[1]); 2, Anthracotheriinae (7[1], 19[1]); 3, (18[1], 25[1], 35[1]); 4, (36[1]); 5, (16[1], 34[1], 38[1]); 6, Microbunodontinae (5[1], 6[1], 33[1]); 7, (26[1], 27[1], 32[1], 39[1], 40[1]; 8, (20[0], 37[1]); 9, Bothriodontinae (6[2], 17[1], 23[1], 39[2]); 10, (26[1], 27[1]); 11, (1[2], 2[2], 3[1], 4[2], 14[1], 20[0], 28[1], 44[2]); 12, (17[2], 18[1], 29[1], 35[1]); 13, (4[1], 33[1]); 14, (6[0], 7[1], 8[3], 24[1], 38[2], 39[1]); 15, (12[1]); 16, (20[0], 45[1]); 17, (4[1], 18[0]); 18, (6[1], 24[0]); 19, (11[1], 14[1], 23[0], 35[0], 44[1]); 20, (13[1], 21[1]); 21, (41[1]); 22, (18[0], 28[1]); 23, (39[0]); 24, (17[3], 40[2]); 25, (1[1], 9[2], 10[1], 11[2], 30[1], 31[1], 42[1], 49[1].](/cms/asset/c51d26fd-c682-4cc1-b604-a9001ae42a76/ujvp_a_722573_o_f0007g.gif)
Morphologic data were compiled from the study of specimens and a literature review. Despite the fact that A. meridionale is only known from a juvenile partial lower dentition, we retained the 51 characters suggested in Lihoreau and Ducrocq (Citation2007) for both taxa, A. acridens from the Toledo L. F. (Albright, Citation1999) and A. meridionale, sp. nov., from Panama (Appendix 2 and Supplementary Data). The data matrix includes cranial and dental characters that are unordered and weighted equally. Characters not known for a taxon were coded as missing. Data were compiled in Mesquite version 2.72 (Maddison and Maddison, Citation2009) and the data set was analyzed under the parsimony criterion using the ‘branch and bound’ algorithm of PAUP version 4.0b10 (Swofford, Citation2003) with zero-length branches not collapsed. The analysis resulted in six equally most parsimonious trees (MPTs) with tree lengths of 120 steps, a consistency index (CI) of 0.583, a retention index (RI) of 0.829, and a homoplasy index (HI) of 0.414 ().
Despite the large amount of missing data for the upper dentition of A. meridionale, our results support the allocation of A. meridionale, sp. nov., with Arretotherium (node 17) in the strict consensus tree (). This association is based on the dimensions of the lower incisors (4[1]), on the presence of only one postprotocrista (18[0]), and partially on the presence of a mesial cusp in the lower premolars in A. acridens from the Toledo Bend L. F. (9[0]). Following our resulting topology, A. acridens from the Great Plains, equivalent to A. acridens in Lihoreau and Ducrocq (Citation2007), is consistently the sister taxon of A. acridens from the Toledo Bend L. F. and A. meridionale. The Toledo Bend form of A. acridens falls out from the holotype of A. acridens (node 18) based on the relative dimension of the labial and lingual cusp on the lower molars (24[0]) and the presence of a strong laterally compressed upper canine (6[1]). Furthermore, A. meridionale differs from both forms of A. acridens with the presence of a postentocristid not reaching the posthypocristid and leaving the longitudinal valley open (23[0]) and the reduction of one incisor (1[2]).
Results from our analysis support A. meridionale as a bothriodontine attributable to the genus Arretotherium; Arretotherium acridens from the Toledo Bend L. F. and A. meridionale as sister taxa with a common ancestor shared with A. acridens from the Great Plains. Our results also suggest that Elomeryx Marsh, Citation1894b (probably E. armatus, the American species), is the sister taxon of Arretotherium, and therefore sister taxon of the other more derived American bothriodontines (Arretotherium and probably Kukusepasutanka). However, the resolution of our analysis was not clear enough to clarify the relationship between the more primitive American anthracotheres (Aepinacodon and Bothriodon).
DISCUSSION AND CONCLUSIONS
Arretotherium meridionale, sp. nov., represents the southernmost occurrence of Anthracotheriidae in the New World. The inferred early Miocene (late Arikareean) age for the Las Cascadas fossil assemblage (Kirby et al., Citation2008; Montes et al., Citation2012; Rincon et al., Citation2012) places A. meridionale as one of the last anthracotheres reported in North America and the only occurrence of the family in Central America (). Although its relationship with other American bothriodontines is still unresolved, results from our cladistic analysis suggest a relationship with A. acridens from The Toledo Bend L. F. These tropical to subtropical forms share several plesiomorphic characters with coeval anthracotheres from different continents, such as Merycopotamus Falconer and Cautley, Citation1847, from the middle to late Miocene of Asia (Douglass, Citation1901; Troxell, Citation1921; Scott, Citation1940), Brachyudus Deperet, Citation1895, from the early Miocene of Eurasia, and Libycosaurus from the late Miocene of Libya. In addition, some plesiomorphic characters in the lower dentition are also present in A. meridionale and Elomeryx. These characters include (1) caniniform upper incisors; (2) the presence of a partially reduced mesiolingual entocristid that diverges from the preentocristid; (3) a relatively enlarged prehypocristid that does not reach the lingual margin of the lower molars; (4) a shallow junction between the postmetacristid and the postprotocristid, and (5) similar transverse widths of the labial and lingual cusps. In fact, A. meridionale seems to be less derived than other early Miocene forms, which are characterized by more conical inner posterior cusps on the lower molars, as well as the disappearance of the mesiolingual entocristid by the middle to late Arikareean (e.g., A. acridens from the Toledo Bend L. F.).
The differences in morphology and geographic distribution during the late Arikareean early Hemingfordian () between A. meridionale and similarly aged A. fricki suggest that the two species occupied two separate ecosystems during the earliest Miocene. A. acridens fron the Toledo Bend L. F. and A. meridionale occupied a distinctive biogeographic area connecting the Gulf Coast with southern Central America (Albright, Citation1998, Citation1999; Kirby and MacFadden, Citation2005), whereas A. fricki was restricted to more temperate and open habitats in the northern and central Great Plains (Strömberg, Citation2002, Citation2006). We also note that there is a remarkable difference in the dimensions of the molars in the later forms of Arretotherium. The Hemingfordian A. fricki from the Great Plains is relatively smaller than A. acridens from Montana (Douglass, Citation1901) and A. acridens from the Toledo Bend L. F. (Albright, Citation1999), whereas A. meridionale seems to indicate an increase in overall body size (). This interpretation should be reevaluated in the light of more complete material to avoid possible bias due to sexually dimorphic morphologies. It is also possible that the morphological differences in A. meridionale could be explained by insular isolation (Eisenberg, Citation1981; Gould and MacFadden, Citation2004) in Panama due to a rapid emergence of new tropical volcanic terrains (Las Cascadas Formation) and rapid changes in the relative sea level during the late Oligocene–earliest Miocene (Zachos et al., Citation2001). It is our hope that ongoing excavation associated with the expansion of the Panama Canal and continued field activities in the Gaillard Cut will reveal new fossil material and add to our knowledge of A. meridionale, allowing us to better address some of these remaining questions.
Rincon et al Supplemental Data
Download Zip (3.9 KB)ACKNOWLEDGMENTS
We thank J. Bourque at FLMNH, who prepared the specimens in the laboratory; J. Schiebout and S. Ting at the Louisiana State University Department of Geology for access to relevant research collections; M. Warren at C.A. Pound Human Identification Laboratory (University of Florida) for access to the X-ray equipment; C. Montes, S. Suarez, M. Vallejo, and F. Moreno (STRI) who helped in the collection of the specimens; R. Hulbert Jr. for help with anatomical terminology and taxonomic nomenclature; F. Lihoreau and B. Shockey for help with the phylogenetic character matrix; and C. Manz, L. Oviedo, K. Cummings, E. Woodruff, F. Herrera, A. Hastings, and J. Pardo, who edited an earlier version of the manuscript and made helpful comments for its improvement. We thank the Panama Canal Authority (ACP) and the Ministerio de Comercio e Industria (MICI) for assess to relevant fossil sites. This research was supported by University of Florida Research Opportunity Grant; the U.S. National Science Foundation Partnerships in International Research and Education grant 0966884 (OISE, EAR, DRL), EAR 0824299, and EAR 0418042; STRI-Tupper Paleontological Fund; STRI-Panama Canal Authority Fund; and Ricardo Perez Toyota, Panama. This is a University of Florida Contribution to Paleobiology number 643.
Handling editor: Jessica Theodor
Related Research Data
LITERATURE CITED
- Albright , L. B. III. 1998 . “ The Arikareean Land Mammal Age in Texas and Florida: southern extension of the Great Plains and Gulf Coastal Plain endemism ” . In Depositional Environments, Lithostratigraphy, and Biostratigraphy of the White River and Arikaree Groups (Late Eocene to Early Miocene, North America) , Edited by: Terry , D. O. Jr. , LaGarry , H. E. and Hunt , R. M. Jr. 167 – 183 . Geological Society of America Special Paper 325 .
- Albright , L. B. III. 1999 . Ungulates of the Toledo Bend Local Fauna (late Arikareean, early Miocene), Texas Coastal Plain . Bulletin of the Florida Museum of Natural History , 42 : 1 – 80 .
- Albright , L. B. III , Woodburne , M. , Fremd , T. , Swisher , C. , MacFadden , B. J. and Scott , G. R. 2008 . Revised chronostratigraphy and biostratigraphy of the John Day Formation (Turtle Cove and Kimberly Members), Oregon, with implications for updated calibration of the Arikareean North American Land Mammal Age . Journal of Geology , 116 : 211 – 327 .
- Aymard , A. 1846 . Essai monographique sur un nouveau genre de mammifére fossile trouvé dans la Haute-Loire, et nommé Entelodon . Annales de la Sociéte d’Agriculture, Sciences, Arts et Commerce du Puy , 12 : 227 – 267 .
- Boisserie , J. R. and Lihoreau , F. 2006 . Emergence of Hippopotamidae: new scenarios . Comptes Rendus Palevol , 5 : 749 – 756 .
- Boisserie , J. R. , Fisher , R. E. , Lihoreau , F. and Weston , E. M. 2011 . Evolving between land and water: key questions on the emergence and history of the Hippopotamidae (Hippopotamoidea, Cetancodonta, Cetartiodactyla) . Biological Reviews , 86 : 601 – 625 . 10.1111/j.1469-185X.2010.00162.x
- Boisserie , J. R. , Lihoreau , F. , Orliac , M. , Fisher , R. E. , Weston , E. M. and Ducrocq , S. 2010 . Morphology and phylogenetic relationships of the earliest known hippopotamids (Cetartiodactyla, Hippopotamidae, Kenyapotaminae) . Zoological Journal of the Linnean Society , 158 : 325 – 366 .
- Bonarelli , G. 1947 . Dinosauro fossile del Sahahra Cirenaico . Rivista Biologica Colonial Roma , 8 : 23 – 33 .
- Coombs , M. C. and Coombs , W. P. 1977 . Dentition of Gobiohyus and a revaluation of the Helohyidae (Artiodactyla) . Journal of Mammalogy , 58 : 291 – 308 .
- Depéret , C. 1895 . Über die Fauna von miocänen Wirbelthieren aus der ersten Mediterranstufe von Eggenburg . Sitzungsberichten der Österreiche Akademie der Wisssenschaften , 4 : 395 – 416 .
- Douglass , E. 1901 . Fossil Mammalia of the White River Beds of Montana . Transactions of the American Philosophical Society, New Series , 20 : 237 – 279 .
- Ducrocq , S. , Chaimanee , Y. , Suteethorn , V. and Jaeger , J. J. 1997 . First discovery of Helohyidae (Artiodactyla, Mammalia) in the Late Eocene of Thailand: a possible transitional form for Anthracotheriidae . Comptes Rendus de l’Academie des Sciences, Paris (Sciences de la Terra) , 325 : 367 – 372 .
- Eisenberg , J. F. 1981 . The Mammalian Radiations. An Analysis of Trends in Evolution, Adaptation, and Behavior , 509 Chicago , Illinois : The University of Chicago Press .
- Falconer , H. and Cautley , P. T. 1847 . Fauna Antiqua Sivalensis, Being the Fossil Zoology of the Siwalik Hills, in the North India, Atlas , 25 – 80 . London : Smith, Lender and Co. . plates
- Gervais , P. 1849 . Recherches sur les mammifères fossils des genres Palaeotherium et Lophiodon et sur les autres animaux de la même classe que l’on a trouvés avec eux dans le midi de la France . Comptes Rendus de l’Academie des Sciences , 29 : 568 – 579 .
- Gould , G. C. and MacFadden , B. J. 2004 . Gigantism, dwarfism, and Cope's rule: nothing in evolution makes sense without a phylogeny . Bulletin of the American Museum of Natural History , 285 : 219 – 237 .
- Hellmund , M. 1991 . Revision der europäischen Species der gattung Elomeryx Marsh, 1894 (Anthracotheriidae, Artiodactyla, Mammalia)—odontologische Untersuchungen . Palaeontographica Abteilung A , 22 : 1 – 101 .
- Hibbard , C. W. and Keenmon , K. A . 1950 . New evidence of the Lower Miocene age of the Blacktail Deer Creek Formation in Montana . Contributions from the Museum of Paleontology University of Michigan , 8 : 193 – 204 .
- Kirby , M. X. and MacFadden , B. J. 2005 . Was southern Central America an archipelago or a peninsula in the middle Miocene? A test using land-mammal body size . Palaeogeography, Palaeoclimatology, Palaeoecology , 228 : 193 – 202 .
- Kirby , M. X. , Jones , D. S. and MacFadden , B. J. 2008 . Lower Miocene stratigraphy along the Panama Canal and its bearing on the Central American peninsula . PLoS ONE , 3 : e2791 doi: 10.1371/journal.pone.0002791.
- Kron , D. G. and Manning , E. 1998 . “ Anthracotheriidae; pp ” . In Evolution of Tertiary Mammals of North America. Volume 1: Terrestrial Carnivores, Ungulates, and Ungulatelike Mammals , Edited by: Janis , C. M. , Scott , K. M. and Jacobs , L. L. 381 – 388 . Cambridge , , U.K : Cambridge University Press .
- Leidy , J. 1869 . The extinct mammalian fauna of Dakota and Nebraska, including an account of some allied forms from other localities, together with a synopsis of the mammalian remains of North America . Proceedings of the Academy of Natural Sciences, Philadelphia , 7 : 1 – 472 .
- Lihoreau , F. and Ducrocq , S. 2007 . “ Family Anthracotheriidae ” . In The Evolution of Artiodactyls , Edited by: Prothero , D. R. and Foss , S. E. 89 – 105 . Baltimore : The Johns Hopkins University Press .
- Linnaeus , C. 1758 . Systema Naturae per Regna tria Naturae, secundum Classes, Ordines, Genera, Species, cum Characteribus, Differentis, Synonymis, Locis. , Tenth edition , 824 Stockholm : Laurentii, Slavi .
- Loomis , F. B. 1925 . Dentition of artiodactyls . Proceedings of the Paleontological Society , 36 : 583 – 604 .
- Macdonald , J. R. 1956 . The North American anthracotheres . Journal of Paleontology , 30 : 615 – 645 .
- Macdonald , J. R. and Shultz , C. B. 1956 . Arretotherium fricki, a new Hemingfordian anthracothere from Nebraska . Bulletin of the University of Nebraska State Museum , 4 : 53 – 58 .
- Macdonald , J. R. 1963 . The Miocene faunas from the Wounded Knee area of western South Dakota . Bulletin of the American Museum of Natural History , 125 : 139 – 238 .
- Macdonald , J. R. and Martin , J. E. 1987 . “ Arretotherium fricki (Artiodactyla, Anthracotheridae) from the Hemingfordian (Miocene) Flint Hill Local Fauna in South Dakota ” . In Papers in Vertebrate Paleontology in Honor of Morton Greene , Edited by: Martin , J. E. and Ostrander , G. E. 57 – 62 . Dakoterra 3 .
- MacFadden , B. J. 2006 . North American land mammals from Panama . Journal of Vertebrate Paleontology , 26 : 720 – 734 .
- MacFadden , B. J. 2009 . Three-toed browsing horse Anchitherium (Equidae) from the Miocene of Panama . Journal of Paleontology , 83 : 489 – 492 .
- MacFadden , B. J. and Higgins , P. 2004 . Ancient ecology of 15 million-year-old browsing mammals within C3 plant communities from Panama . Oecologia , 140 : 169 – 182 .
- MacFadden , B. J. and Hunt , R. M. Jr . 1998 . “ Magnetic polarity stratigraphy and correlation of the Arikaree Group, Arikareean (late Oligocene–early Miocene) of northwestern Nebraska ” . In Depositional Environments, Lithostratigraphy, and Biostratigraphy of the White River and Arikaree Groups (Late Eocene to Early Miocene, North America) , Edited by: Terry , D. O. , LaGarry , H. E. and Hunt , R. M. Jr. 143 – 165 . Geological Society of America Special Paper 325 .
- MacFadden , B. J. , Kirby , M. , Rincon , A. , Montes , C. , Moron , S. , Strong , N. and Jaramillo , C. 2010 . Extinct peccary “Cynorca” occidentale (Tayassuidae) from the Miocene of Panama and correlations to North America . Journal of Paleontology , 84 : 288 – 298 .
- Maddison , W. P. and Maddison , D. R. 2009 . Mesquite: A Modular System for Evolutionary Analysis . Version 2.72 , Available at http://mesquiteproject.org. Accessed December 2, 2010
- Marsh , O. C. 1894a . A new Miocene mammal . American Journal of Science (series 3) , 47 : 409
- Marsh , O. C. 1894b . Description of Tertiary artiodactyls . American Journal of Science (series 3) , 48 : 259 – 274 .
- Matthew , W. D. 1909 . Observations upon the genus Ancodon . Bulletin of the American Museum of Natural History , 26 : 1 – 7 .
- von Meyer , H. 1832 . Palaeologica, zur Geschichte der erde und ihrer Geschöpfe XI . Frankfurt am Main , : 560
- Montes , C. , Cardona , A. , McFadden , R. , Moron , S. E. , Silva , C. A. , Restrepo-Moreno , S. , Ramirez , D. A. , Hoyos , N. , Wilson , J. , Farris , D. , Bayona , G. A. , Jaramillo , C. A. , Valencia , V. , Bryan , J. and Flores , J. A. 2012 . Evidence for middle Eocene and younger emergence in Central Panama: implications for Isthmus closure . Geological Society of America Bulletin , 124 : 780 – 799 . 10.1130/B30528.1
- Owen , R. 1848 . Description of teeth and portions of two of two extinct anthracotheroid quadrapeds (Hyopotamus vectianus and H. bovinus) discovered by the Marchioness of Hastings in the Eocene deposits on the N. W. coast of the Isle of Wight, with an attempt to develop Cuvier's idea of the classification of pachyderms by the number of their toes . Quarterly Journal of the Geological Society of London , 4 : 104 – 141 .
- Rincon , A. F. , Bloch , J. I. , MacFadden , B. J. , Suarez , S. and Jaramillo , C. 2010 . New early Miocene camelids from the Las Cascadas Formation, Panama Canal, Central America. Presented at 70th Society of Vertebrate Paleontology Meeting, Pittsburgh, PA . Journal of Vertebrate Paleontology , 30 ( Program and Abstracts ) : 10–13 October 2010 Available at http://www.vertpaleo.org/meetings/documents/SVP10Abstracts_WEB.pdf. Accessed October 8, 2010, 151A
- Rincon , A. F. , Bloch , J. I. , MacFadden , B. J. , Suarez , C. and Jaramillo , C. A. 2012 . New floridatragulines (Mammalia, Camelidae) from the early Miocene Las Cascadas Formation, Panama . Journal of Vertebrate Paleontology , 32 : 456 – 475 .
- Rooney , T. , Franceschi , P. and Hall , C. 2010 . Water saturated magmas in the Panama Canal region: a precursor to Adakite-like magma generation . Contributions to Mineralogy and Petrology , 161 : 373 – 388 .
- Scott , W. B. 1940 . “ Artiodactyla ” . In The Mammalian Fauna of the White River Oligocene , Edited by: Scott , W. B. and Jepsen , G. L. 363 – 746 . Philadelphia : Transactions of the American Philosophical Society . 28, 4, 1–980
- Schmidt , M. 1913 . Uber Paarhufer der fluviomarinen Schichten des Fajum: odontographisches und osteologisches Material . Geologische und Palaeontologische Abhandlungen , 11 : 153 – 264 .
- Simpson , G. G. 1945 . The principles of classification and a classification of mammals . Bulletin of the American Museum of Natural History , 85 : 1 – 350 .
- Smith , J. B. and Dodson , P. 2003 . A proposal for a standard terminology of anatomical notation and orientation in fossil vertebrate dentitions . Journal of Vertebrate Paleontology , 23 : 1 – 2 .
- Storer , J. E. and Bryant , H. N. 1993 . Biostratigraphy of the Cypress Hills Formation (Eocene to Miocene), Saskatchewan: equid types (Mammalia: Perissodactyla) and associated faunal assemblages . Journal of Paleontology , 67 : 660 – 669 .
- Strömberg , C. A. E. 2002 . The origin and spread of grass-dominated ecosystems in the Late Tertiary of North America: preliminary results concerning the evolution of hypsodonty . Palaeogeography, Palaeoclimatology, Palaeoecology , 177 : 59 – 75 .
- Strömberg , C. A. E. 2006 . Evolution of hypsodonty in equids: testing a hypothesis of adaptation . Paleobiology , 32 : 236 – 258 .
- Suteethorn , V. , Buffetaut , E. , Helmcke-Ingavat , R. , Jaeger , J. J. and Jongkanjanasoontorn , Y. 1988 . Oldest known Tertiary mammals from South-East Asia: Middle Eocene primate and anthracotheres . Neues Jahrbuch fur Geologie und Palaontologie Monatshefte , 9 : 563 – 570 .
- Swofford , D. A. 2003 . PAUP* 4.0 , Sunderland , Massachusetts : Sinauer Associates .
- Tedford , R. H. , Skinner , M. F. , Fields , R. W. , Rensberger , J. M. , Whistler , D. P. , Galusha , T. , Taylor , B. E. , Macdonald , J. R. and Webb , S. D. 1987 . “ Faunal succession and biochronology of the Arikareean through Hemphillian interval (Late Oligocene through Earliest Pliocene Epochs) in North America ” . In Cenozoic Mammals of North America , Edited by: Woodburne , M. O. 153 – 210 . Berkeley, California : University of California Press .
- Tedford , R. H. , Albright , L. B. III , Barnosky , A. D. , Ferrusquia-Villafranca , I. , Hunt , R. M. Jr. , Storer , J. E. , Swisher , C. C. III , Voorhies , M. R. , Webb , S. D. and Whistler , D. P. 2004 . “ Mammalian biochronology of the Arikareean through Hemphillian interval (late Oligocene through early Pliocene epochs) ” . In Late Cretaceous and Cenozoic Mammals of North America: Biostratigraphy and Geochronology , Edited by: Woodburne , M. O. 169 – 231 . New York : Columbia University Press .
- Troxell , E. L. 1921 . The American bothriodonts . American Journal of Science , 3 : 325 – 339 .
- Uhen , M. D. , Coates , A. G. , Jaramillo , C. A. , Montes , C. , Pimiento , C. , Rincon , A. , Strong , N. and Velez-Juarbe , J. 2010 . Marine mammals from the Miocene of Panama . Journal of South American Earth Sciences , 30 : 167 – 175 .
- Whitmore , F. C. and Stewart , R. H. 1965 . Miocene mammals and Central American seaways . Science , 148 : 180 – 185 .
- Woodring , W. P. 1957 . Geology and paleontology of Canal Zone and adjoining parts of Panama; description of Tertiary mollusks; gastropods; Trochidae to Turritellidae . US Geological Survey Professional Paper 306-A , : 1 – 239 .
- Woodring , W. P. 1982 . Geology and paleontology of Canal Zone and adjoining parts of Panama; description of Tertiary mollusks; pelecypods, Propeamussiidae to Cuspidariidae; additions to families covered in P306-E; additions to gastropods; cephalopods . US Geological Survey Professional Paper 306-7 , : 541 – 759 .
- Woodring , W. P. and Thompson , T. F. 1949 . Tertiary formations of Panama Canal Zone and adjoining parts of Panama . American Association of Petroleum Geologists Bulletin , 33 : 223 – 247 .
- Zachos , J. C. , Pagani , M. , Sloan , L. , Thomas , E. and Billups , K. 2001 . Trends, rhythms, and aberrations in global climate 65 Ma to present . Science , 292 : 686 – 693 .
APPENDIX 1. Description of dental characters used in the phylogenetic analysis. All characters are treated as unordered. Modified from Lihoreau and Ducrocq (Citation2007).
| 1. | Lower incisors: three (0); from two to three (1); two (2). | ||||
| 2. | Upper incisors: three of equal size (0); three with i3 reduced (60% or less) (1); two (2). | ||||
| 3. | Lower incisor morphology: not caniniform (0); at least one caniniform (1). | ||||
| 4. | Relative dimension of lower incisors: all equal size (0); i2 larger (1); i3 larger (2). | ||||
| 5. | Wear on lower canine: distal wear facet caused by the contact with upper C (0); mesial wear facet cause by contact with I3 (1). | ||||
| 6. | Upper canine morphology: strong with subcircular cross-section (0); strong and laterally compressed (1); premolariform (2). | ||||
| 7. | Lower canine in males: premolariform (0); large (1); ever-growing (2). | ||||
| 8. | Lower canine cross-section at cervix: subcircular (0); elliptical with rounded mesial margin and distal keel (1); elliptical with a mesial and distal crest (2); elliptical with a concave buccal margin and a distal keel (3). | ||||
| 9. | Accessory cusps on the mesial crest of lower premolars: none (0); only one (1); several (2). | ||||
| 10. | Presence of five upper premolars: no (0); yes (1). | ||||
| 11. | Distolabial crest on upper premolars: simple (0); with a maximum of two accessory cusps (1); with more than two (2). | ||||
| 12. | Accessory cusp on p4: no (0); yes (1). | ||||
| 13. | p1 roots: one (0); two (1). | ||||
| 14. | Mesial crests on P1–P3: one (0); two (1). | ||||
| 15. | Number of P4 roots: three (0); two (1); one (2). | ||||
| 16. | Accessory cusp on distolingual margin of P3: one (0); none (1). | ||||
| 17. | Upper molar mesostyle; simple (0); V-shaped and invaded by a transversal valley (1); loop-like (2); divided into two (3). | ||||
| 18. | Number postprotocristae: one (0), two (1). | ||||
| 19. | Accessory cusp on upper molar mesial cingulum: no (0); yes (1). | ||||
| 20. | Number of cristules issued from the metaconule: two (0); three (1). | ||||
| 21. | Preprotocristids and prehypocristids on lower molars: do not reach the lingual margin of the tooth (0); reach the lingual margin (1). | ||||
| 22. | Hypoconulid on m3: loop-like (0); single cusp (1). | ||||
| 23. | Postentocristid on lower molars: does not reach the posthypocristid and leaves the ling. valley open (0); reaches the posthypocristid and closes the long valley (1). | ||||
| 24. | Dimension of the lingual and labial cusps: equal (0); different (1). State (1): labial cusps twice as large at their basis as the lingual cusp. | ||||
| 25. | Entoconulid on m3: absent (0); present (1). | ||||
| 26. | Number of cristids issued to from the hypoconulid: three (0); two (1). | ||||
| 27. | Position of the preentocristid on lower molars: reaches the hypoconid summit (0); reached the prehypocristid (1). | ||||
| 28. | Premetacristid on lower molars: present (0); absent (1). | ||||
| 29. | Mesial part of looplike hypoconulid: open (0); pinched (1). | ||||
| 30. | Entoconid fold on lower molars: absent (0); present (1). | ||||
| 31. | Ventral vascular groove on mandible: slightly marked (0); absent (1); strongly marked (2). | ||||
| 32. | Morphology of mandibular symphysis cross-section: ‘U’-shaped (0); ‘V’-shaped (1). | ||||
| 33. | Transverse constriction of mandible at Cp1 diastema: no (0); yes (1). | ||||
| 34. | Cp1 diastema: absent (0); present (1). | ||||
| 35. | p1-p2 diastema: absent (0); present (1). | ||||
| 36. | Lateral mandibular tuberosity: absent (0); present (1). | ||||
| 37. | Dentary bone fussion at the symphysis: no (0); yes (1). | ||||
| 38. | Morphology of the symphysis is sagittal section: elliptic (0); dorsally concave (1); ventrally concave (2). | ||||
| 39. | Maximal thickness of the symphysis in sagittal section: in the middle part (0); in the anterior part (1); in the posterior part (2). | ||||
| 40. | Number and position of main external mandibular foramina: only one foramen, below the anterior part of the premolar row (0); two foramina, one below the anterior part of the premolar row, the other below its posterior part (1); one foramen, below the posterior part of the premolar row (2). | ||||
| 41. | Tuberosity on the dorsal border of the mandible at c–p1 diastema: no (0); yes (1). | ||||
| 42. | Palatine depression between the canines: no (0); yes (1). | ||||
| 43. | Canine fossa: short (0); long (1). | ||||
| 44. | Aperture of the main palatine foramen: between M3 and P3; between P2 and P1 (1); between P1 and C. | ||||
| 45. | Morphology of the frontonasal suture: V-shaped (0); rounded or straight (1). | ||||
| 46. | Lachrymal extension: separated from the nasal by the frontal (0); in contact with the nasal (1). | ||||
| 47. | Supraorbital foramina on the frontal: one (0); several (1). | ||||
| 48. | Facial crest: horizontal (0); oblique (1). | ||||
| 49. | Anterior border of premaxillary in lateral view: concave (0); convex (1). | ||||
| 50. | Postglenoid foramen position: posterior to the styloid process of the tympanic bulla (0); anterior to the styloid process of the tympanic bulla (1). | ||||
| 51. | Opening of internal choanes: at M3 (0); behind M3 (1). | ||||
APPENDIX 2. Character-taxon matrix used in the phylogenetic analyses of Anthracotheriidae. See Appendix 1 for character descriptions.
10 20 30 40 50
S. krabiense 000??00?00000000000000000000000000000000000?00?00?0
M. minimun 000011010000000100000000011000011100111100001000000
M. silistrensis 0?001?0100000?010000000001100001110011110??????????
A. magnum 00000010000000?00111000010000000001110010000????0?0
A. mosvialense 00000010000000?001110000100000000011100?0000???00?0
A. chaimanei 0?00?0?00000?0?001110000100000000?10?00?00000?00??0
A. pangan ???0?01?0?00000001110000100000???0?00???0??????????
A. thailandicus 0?001?01000000?10001000001100001110001110?000?0???0
A. ulnifer 000011010000000101010000000000001110010000000000000
H. occidentalis 0?0000140000000000110010000000?00000100?000000000?0
B. fraasi 00??0202000000?110010010000000000100012000?00?000?1
B. aequatorialis 22120202000001?110000010011100000100112000020000001
B. onoideus 22120202000001?110000010011100000100112000020??0001
A. americanus 0001020200000001210000100110100011101120000000000?0
B. velaunus 0001020200000001210100100110100011101120000?0000000
E. crispus 000000130001000121010011011010000110021000000000001
E. armatus 000000130000000121010011011010000110021000000000001
E. borbonicus 00000013100100012100001101101000011002100000??000??
A. acridens 00010013000000?120000011011010000110021000001?1?001
A. zelteni 10000013101101?121000001011010000100021100?11?1?0?1
S. palaeindicus 0?000?13101111012100100101101000010002111??????????
H. blandfordi ??0???132??11???2200100101101000010002101??????????
M. pusillus 000000131011110120001001011110000100020000110111011
M. dissimilis 000000231011110130001101011111200100020200111111011
M. medioximus 000000131011110130001001011110000100020200111111011
L. anisae 1?00001321211111200010010111111001000211011????11?1
L. petrocchii 110000132121112120001101011111100100021101121111111
A. acridens(TBend) 0?0101100?0100113000001011101?0?011?0200???????????
A. meridionale 2?0??11?????0?????0?0?00???0??0??11?0210???????????