
ABSTRACT
Stegosaurs are a major clade of ornithischian dinosaurs, yet because of their fragmentary fossil record, their interrelationships and early evolution are poorly understood. Here, we describe a new stegosaur, Bashanosaurus primitivus, gen. et sp. nov., and some other indeterminate stegosaur materials. We provide new U-Pb detrital zircon ages for horizons bounding the holotypic quarry in the Lower Member of the Shaximiao Formation, Yunyang, Chongqing Municipality, China, which indicate a Middle Jurassic (Bajocian) age. Bashanosaurus represents the earliest record of stegosaurs in Asia and one of the earliest records of this clade in the world. The dorsal vertebrae, scapula, coracoid, femur, and plates of Bashanosaurus primitivus possess several unique characters among Stegosauria, including the elevation of the parapophyses of dorsal vertebrae on stalks at the base of the transverse processes, a flared distal end of the scapula, and a small acromial process. Among armored dinosaurs (thyreophorans), these features are reminiscent of the basally branching taxon Scelidosaurus, indicating that Bashanosaurus possesses a unique mosaic of plesiomorphic thyreophoran and derived stegosaur features. Phylogenetic analysis shows that Bashanosaurus primitivus is the earliest-diverging stegosaur, along with Chungkingosaurus, from the Upper Member of the Shaximiao Formation, consistent with the early age of the taxon.
INTRODUCTION
The stegosaurs are characterized by the possession of two parasagittal rows of hypertrophied dermal armor plates and/or spines extending from the neck to the end of the tail and range from Middle Jurassic to Early Cretaceous in age (Galton and Upchurch, Citation2004). Stegosaur fossils have been found on all continents except for Antarctica and Australia. The first stegosaurs are known from the Middle Jurassic of Europe, China, Argentina, and Morocco: Loricatosaurus priscus is from the Middle Callovian Lower Oxford Clay Formation, Cambridgeshire, U.K. (Maidment et al., Citation2008); Huayangosaurus taibaii is from the Bathonian–Callovian Lower Member of the Shaximiao Formation, Sichuan Province, China (Zhou, Citation1984); Isaberrysaura is known from the Bajocian part of the Los Molles Formation of Argentina (Salgado et al., Citation2017), and Adratiklit boulahfa is from the Bathonian El Mers II Formation of Morocco (Maidment et al., Citation2020). Stegosaurian diversity appears to have decreased after the Jurassic, and they seem to have gone extinct by the middle Cretaceous (Maidment et al., Citation2020). Fourteen valid genera are currently recognized (Maidment et al., Citation2020). Fragmentary fossil material has hindered attempts to explore interrelationships between members of Stegosauria, and the taxonomy of various species and genera remains in flux (Carpenter and Galton, Citation2001; Galton and Upchurch, Citation2004; Maidment et al., Citation2008; Carpenter, Citation2010; Cobos et al., Citation2010; Galton and Carpenter, Citation2016).
In 2016, we discovered a new dinosaur quarry in the Middle Jurassic Lower Member of the Shaximiao Formation in Laojun Village, Pu’an Township, Yunyang County, Chongqing Municipality, China (Li et al., Citation2019; Dai et al., Citation2020; Tan et al., Citation2020). The Shaximiao Formation is divided into Lower Member and Upper Members, and the age of the Upper Member is generally considered to be Late Jurassic (Peng et al., Citation2005). However, the age of the Shaximiao Formation was regarded as Middle Jurassic by Wang et al. (Citation2010) and Li et al. (Citation2010). A number of radiometric ages have been reported for the Shaximiao Formation, including 167 Ma or 172 Ma (ESR: Electron Spin Resonance) (Zong and Shi, Citation1997), 167–172 Ma or 178–169 Ma (ESR) (Gou et al., Citation2000), 163 ± 3 Ma (Li et al., Citation2010), 160 ± 4 Ma (Luo et al., Citation2014), 158 ± 7 Ma (Qian et al., Citation2016), and 159 ± 2 Ma (Wang et al., Citation2018). Geotectonically, this location is situated in the southwest of the Yangzi Block where the red beds of the Shaximiao Formation are well exposed, and outcrops are generally continuous (Gu and Liu, Citation1997). The Lower Member of the Shaximiao Formation consists of purplish red muddy siltstone and silty mudstone interbedded with siltstone and mudstone (). The grayish-yellow massive fine-grained lithic feldspathic sandstone and purplish red silty mudstone form multiple rhythmic layers of uneven thickness, and the siltstones contain calcareous nodules. This development of ‘redbeds’ and calcareous nodules indicates a semi-arid, terrestrial paleoenvironment (Zhang et al., Citation2020).
FIGURE 1. Maps showing the location where the Bashanosaurus primitivus was recovered. A, sketch map of China showing the location of Chongqing Municipality. B, sketch map of Yunyang County. C, sketch map of the fossil locality. D, stratigraphic chart with Lower Member of the Shaximiao Formation in Yunyang County, Chongqing Municipality, China.
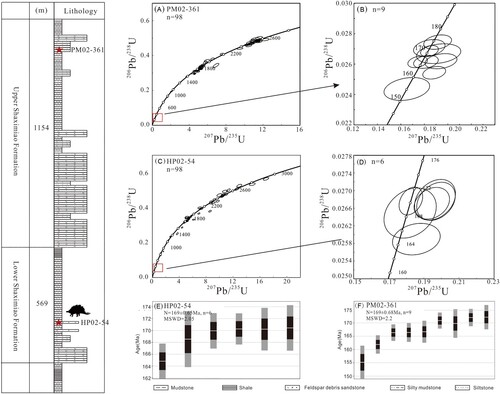
FIGURE 2. Lithological section of the Yunyang dinosaur locality in the Lower Member of the Shaximiao Formation in this study (left). A–D: concordia diagrams of detrital zircon U-Pb ages for the samples analyzed in this study; E–F: histogram of zircons for the samples analyzed in this study.
The new quarry was excavated systematically to form a wall of dinosaur fossils 150 m long and 8 m high. Further material was also discovered in an exploratory trench of the same stratigraphic unit about 900 m away from the original excavation.
Herein we describe the newly discovered stegosaur materials and examine their phylogenetic position in light of their early stratigraphic age. Our results shed new light on the early evolution and diversification of stegosaurs, as well as contributing to debates around the age of the Shaximiao Formation and its various faunas.
Institutional Abbreviations—CLGPR, Chongqing Laboratory of Geoheritage Protection and Research, Chongqing Municipality, China; NHMUK, Natural History Museum, London, U.K.
Anatomical Abbreviations—ACPL, anterior centroparapophyseal lamina; PPDL, paradiapophyseal lamina; IPZS, intraprezygapophyseal shelf; PODL, postzygodiapophyseal lamina. (Wilson, Citation1999).
METHODS
Geochronologic Methods
We collected two sandstones from the Shaximiao Formation for detrital U-Pb geochronology analyses. Sample HP02-54 comes from a gray-green tuffaceous sandstone from the Lower Member of the Shaximiao Formation, immediately beneath the bonebed. Sample PM02-361 comes from a gray-green sandstone from the Upper Member of the Shaximiao Formation, immediately above the bone bed ().
Zircon separation and cathodoluminescence (CL) image collection were conducted at Beijing Zhongke Kuangyan Test Technology Co., Ltd. The samples were crushed and sieved using standard methods, and were then washed, dried and processed with a magnetic separator. Euhedral grains were hand-picked under a binocular microscope, mounted in epoxy resin, and eventually polished. A Tescan MIRA 3 field emission scanning electron microscope (SEM) was used to collect detailed CL images of individual zircon grains.
Zircon U-Pb isotope data were conducted using an Agilent 7700e ICP-MS instrument coupled to a GeolasPro laser-ablation system at Wuhan Sample Solution Analytical Technology Co., Ltd, using methods described in Liu et al. (Citation2008), Hu et al. (Citation2015), and Zong et al. (Citation2017). Raw data are presented in Tables S1 and S2.
Phylogenetic Methods
To assess the phylogenetic position of Bashanosaurus primitivus, the taxon was added to the character-taxon matrix of Maidment et al. (Citation2020). Settings from the original analysis were retained: Pisanosaurus was set as the outgroup, Ankylosauria was represented by three taxa, Gastonia, Euoplocephalus, and Sauropelta, and all continuous characters (1–24) and characters 105 and 106 were ordered; all others were unordered. The matrix was analysed via parsimony in TNT v. 1.5 (Goloboff et al., Citation2008) using a New Technology search with the ratchet, sectorial, drift and tree fusing algorithms and 10 random addition sequences. Branch support was calculated using Bremer support and GC frequencies (1000 replicates, traditional search). In order to explore the phylogenetic placement of Bashanosaurus more thoroughly, the taxon was constrained to fall in several specified positions (see Results) using the ‘force’ command in TNT. The matrix and character list can be found in the Supplemental Data.
SYSTEMATIC PALEONTOLOGY
DINOSAURIA Owen, Citation1842
ORNITHISCHIA Seeley, Citation1887
STEGOSAURIA Marsh, Citation1877
BASHANOSAURUS PRIMITIVUS, gen. et sp. nov.
Etymology—The generic name “Bashan” refers to the ancient name of Chongqing, and the specific name “primitivus” indicates that this newly named species represents the earliest-diverging stegosaur.
Diagnosis—Bashanosaurus primitivus differs from all other stegosaurs by possession of the following autapomorphies: (1) the parapophyses of dorsal vertebrae are elevated on stalks at the base of the transverse process and are clearly visible in anterior and posterior views; (2) the scapula blade is slender and flares distally; (3) the acromial process of the scapula is small, triangular and poorly developed; (4) the 4th trochanter of the femur is positioned below the middle of the shaft; (5) the bases of plates are strongly convex and transversely thickened, and are separated from the anterior/posterior margins of the plates by a distinct groove.
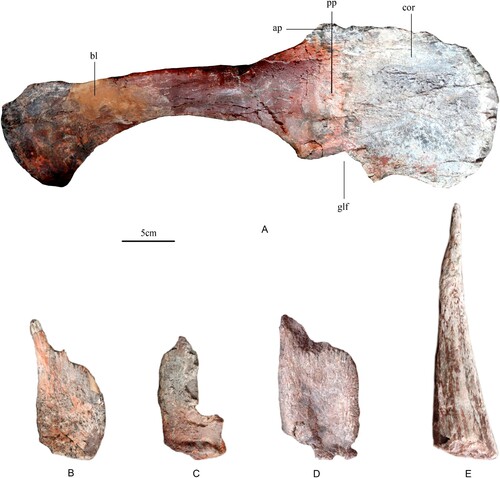
Holotype—A partial individual that is CLGPR V00006-1 is preserved on the west side of the wall of dinosaur fossils (), and comprises one dorsal vertebra, two caudal vertebrae, right scapula, right coracoid, left femur, left tibia, left fibula, one metatarsal, three pieces of dermal armor (1 plate and 2 spines) and some ribs.
FIGURE 3. A, photograph (top) and interpretative sketch (bottom) of Bashanosaurus primitivus (CLGPR V00006-1) on the west side of the Dinosaur Fossil Wall. B, photograph (top) and interpretative sketch (bottom) of Bashanosaurus primitivus (CLGPR V00006-2) in the middle of the Dinosaur Fossil Wall. Abbreviations: cav, caudal vertebra; co, coracoid; dv, dorsal vertebra; f, femur; fl, fibula; mt, metatarsal; pl, plate; r, rib; sc, scapula; sp, spine; t, tibia.

Referred Specimens—A partial individual that is CLGPR V00006-2 is preserved in the middle of the wall of dinosaur fossils (), and comprises five dorsal vertebrae, right tibia, right fibula, one piece of dermal plate, and some ribs.
CLGPR V00006-3 is a dorsal vertebra from the east side of the wall of dinosaur fossils ().
FIGURE 4. A–F, dorsal vertebrae referred to Bashanosaurus primitivus (CLGPR V00006-3) from the Middle Jurassic of China. A, anterior; B, left lateral; C, dorsal; D, posterior; E, right lateral; F, ventral; G, posterior views of dorsal vertebrae referred to Bashanosaurus primitivus (CLGPR V00006-1) from the Middle Jurassic of China. H, posterior view of dorsal vertebrae referred to Bashanosaurus primitivus (CLGPR V00006-2) from the Middle Jurassic of China. I, anterior view of dorsal vertebrae referred to Bashanosaurus primitivus (CLGPR V00006-2) from the Middle Jurassic of China. J–O, anterior caudal vertebrae referred to Bashanosaurus primitivus (CLGPR V00006-1) from the Middle Jurassic of China. J, anterior; K, left lateral; L, dorsal; M, posterior; N, right lateral; O, ventral. P–R, middle caudal vertebrae referred to Bashanosaurus primitivus (CLGPR V00006-1) from the Middle Jurassic of China. P, right lateral; Q, posterior; R, ventral. Abbreviations: acpl, anterior centroparapophyseal lamina; dia, diapophysis; ipzs, intraprezygapophyseal shelf; nc, neural canal; ns, neural spine; para, parapophysis; podl, postzygodiapophyseal lamina; pozyg, postzygapophysis; ppdl, paradiapophyseal lamina; przyg, prezygapophysis.

CLGPR V00006-1, V00006-2, and V00006-3 are all from the wall of dinosaur fossils. CLGPR V00006-2 is about 50 m away from CLGPR V00006-1. CLGPR V00006-3 is about 120 m away from CLGPR V00006-1 and about 70 m away from CLGPR V00006-2.
The size, morphological features of the bones and the preservation styles show that CLGPR V00006-1, CLGPR V00006-2, and CLGPR V00006-3 belong to three separate stegosaurian individuals.
Locality and Horizon—Laojun Village (108°55′25.86′′E, 30°50′45.44′′N), Pu’an Township, Yunyang County, Chongqing Municipality, People’s Republic of China; Lower Member of the Shaximiao Formation, Middle Jurassic (Bajocian).
DESCRIPTION
Dorsal Vertebrae
Seven dorsal vertebrae are preserved in the type and referred specimens. One middle dorsal vertebra is well-preserved in the referred specimen CLGPR V00006-3, with only the top of the neural spine missing (). The centrum is almost amphiplatyan, in contrast to the condition in Chungkingosaurus, in which the anterior articular facet is strongly convex (Maidment et al., Citation2018). The length, width, and height of the centrum are similar (Tables S3 and S4), as in the dorsal centra of other stegosaurs such as Stegosaurus (Maidment et al., Citation2015), Loricatosaurus (Galton, Citation1985), Kentrosaurus (Hennig, Citation1915), and Huayangosaurus (Zhou, Citation1984), but in contrast to Dacentrurus, in which the dorsal centra are wider than long (Galton Citation1990).
Both the anterior and posterior articular surfaces of the centrum are sub-circular in outline. There is no distinct keel on the ventral surface of the centrum. In lateral view, the surface of the centrum is smooth and gently longitudinally concave. The lateral surfaces of the centra lack deep central fossae, as seen on the dorsal vertebrae of Gigantspinosaurus (Maidment et al., Citation2008). The lateral surface of the neural arch is gently convex; the area immediately below this convex surface is concave and merges with the lateral surface of the centrum. The neural arch pedicle is not greatly expanded dorsally, similar to early-diverging stegosaurs such as Huayangosaurus (Maidment et al., Citation2006) and Gigantspinosaurus (Maidment et al., Citation2008), unlike in most stegosaurs such as Stegosaurus (Maidment et al., Citation2015), Kentrosaurus (Hennig, Citation1915), and Adratiklit (Maidment et al., Citation2020). The neurocentral suture is not visible. The height of the neural canal is about 22 mm, and it has a round outline. The parapophysis is situated at the base of the diapophysis, and it is elevated on a short stalk so that it is clearly visible in anterior and posterior view. The elevation of the parapophysis on a short stalk is greater than that seen in other stegosaurs such as Huayangosaurus (Zhou, Citation1984; Maidment et al., Citation2006); Adratiklit (Maidment et al., Citation2020), Gigantspinosaurus (Maidment et al., Citation2008), Kentrosaurus (Hennig, Citation1915), and Stegosaurus (Maidment et al., Citation2015), but is similar to that of the basally branching thyreophoran Scelidosaurus (NHMUK PV R1111). The parapophysis is separated from the base of the prezygapophysis by a distinct anterior centroparapophyseal lamina (ACPL). Its dorsal corner is linked to the tip of the diapophysis by a faint but distinct paradiapophyseal lamina (PPDL). The parapophysis has an oval outline with its long axis trending dorsolaterally, and it is gently concave. In anterior view, the prezygapophyses diverge from each other at an angle of ∼70° from vertical. The articular surfaces of prezygapophyses are sub-square in outline and flat. In dorsal view, the middle and posterior area of prezygapophyses has an intraprezygapophyseal shelf (IPZS) that is continuous with the base of neural spine, similar to the condition in Stegosaurus (Maidment et al., Citation2015), Kentrosaurus (Hennig, Citation1915), and Huayangosaurus (Zhou, Citation1984).
In anterior view, the diapophyses extend dorsolaterally, forming an angle of ∼45° to vertical. In lateral view, the diapophyses have a sub-oval cross-section, and their posterior margins are linked to the postzygapophyses by a postzygodiapophyseal lamina (PODL). In posterior view, the postzygapophyses are separated by a deep midline groove. This contrasts with the condition in Stegosaurus (Maidment et al., Citation2015), where a midline groove is entirely absent, and in other stegosaurs such as Kentrosaurus (Hennig Citation1915), Loricatosaurus (Galton, Citation1985), Hesperosaurus (Maidment et al., Citation2018), Adratiklit (Maidment et al., Citation2020), and Huayangosaurus (Zhou, Citation1984), where the postzygapophyses are separated by a much shorter midline groove. It is, however, similar to the condition in the basally branching thyreophoran Scelidosaurus (NHMUK PV R1111). A ridge extends from the base of the postzygapophyses down the midline of the neural arch to the dorsal margin of the neural canal. The articular facets of the postzygapophyses are sub-square in outline and flat. The postzygapophyses overhang the posterior margin of the centrum, as in Stegosaurus (Maidment et al., Citation2015), but different to Huayangosaurus (Zhou, Citation1984) and Kentrosaurus (Hennig, Citation1915), in which the postzygapophyses extend posteriorly to about the same level as the posterior centrum surface. The top of the neural spine is missing. In anterior view, the neural spine is a transversely compressed plate. The anterior margin of the spine is narrower than the posterior margin. The spine projects dorsally and slightly posteriorly.
One almost complete dorsal vertebra is preserved in situ in CLGPR V00006-1 (). Five dorsal vertebrae are preserved in situ in CLGPR V00006-2, but only the anterior or posterior surface can be seen (). Two of these are almost complete (); in the other three, only the centrum can be seen. They are all very similar to the dorsal vertebrae of CLGPR V00006-3.
Caudal Vertebrae
CLGPR V00006-1 preserves two caudal vertebrae. One is an almost complete anterior caudal vertebra (). The centrum is about 35 mm long, 56 mm wide, and 40 mm high. The anterior and posterior articular surfaces of the centrum are slightly concave. The anterior articular surface is slightly heart-shaped, and the posterior articular surface is sub-oval in outline. The ventral surface of the centrum is smooth aside from the distinct chevron facets. The left transverse process is better preserved than the right. The transverse processes are positioned on the anterolateral part of the centrum and extend laterally and slightly dorsally, as in Kentrosaurus (Hennig, Citation1915) and Dacentrurus (Galton, Citation1990), but different from Huayangosaurus (Zhou, Citation1984). The transverse processes are long, triangular in dorsal view and the base of the left transverse process is 17 mm anteroposteriorly. A dorsal process on the transverse process is absent, as in Huayangosaurus (Zhou, Citation1984), but in contrast to Stegosaurus, Kentrosaurus, and Dacentrurus armatus (Galton, Citation1991). In lateral view, the neural arch surface is flat. The outline of the neural canal is round in anterior view. The prezygapophyses clearly overhang the anterior margin of the centrum, different from Huayangosaurus, where the prezygapophyses overhang the anterior margin of the centrum only slightly (Zhou, Citation1984). The articular surfaces of prezygapophyses are flat and extend anterodorsally, differing from Kentrosaurus (Hennig, Citation1915), in which the prezygapophyses extend anteriorly. The postzygapophyses do not extend posteriorly over the posterior margin of the centrum, and their articular facets are flat. The top of the neural spine is missing; the preserved part of the neural spine is 54 mm long, which greater than the height of the centrum, as in other stegosaurs, for example: Kentrosaurus (Hennig, Citation1915), Stegosaurus (Maidment et al., Citation2015), and Dacentrurus armatus (Galton, Citation1990). In anterior view, the neural spine is a high and transversely compressed plate. Although the top of the spine is missing, the length anteroposteriorly is equal from base to top, different from Huayangosaurus, where the spine contracts from base to top (Zhou, Citation1984). The spine projects dorsally and posteriorly and is not bifurcated, as in Huayangosaurus (Zhou, Citation1984), Kentrosaurus (Hennig, Citation1915), and Hesperosaurus (Galton, Citation1990), but in contrast to Stegosaurus (Maidment et al., Citation2015).
A middle caudal vertebra is also preserved (), but only right lateral, posterior, and ventral surfaces can be seen. The centrum is longer anteroposteriorly than it is wide transversely and is as tall dorsoventrally as it is wide transversely. The posterior articular surface is nearly square in outline and concave. Ventrally, there is no keel, but distinct facets for chevrons are present. On the lateral surface, there is no evidence for transverse processes, suggesting these were not present posterior to the mid-caudal region. This is similar to Stegosaurus (Maidment et al., Citation2015), but different from Kentrosaurus (Hennig, Citation1915) and Huayangosaurus (Zhou, Citation1984), in which transverse processes are present on the distal half of the tail. The neural canal is teardrop-shaped. The prezygapophyses extend anterodorsally and do not overhang the anterior articular facet. The postzygapophyses and neural spine are missing.
Scapula
The right scapula is almost completely preserved, but a small portion of the distal end of the blade is missing (). The scapula and coracoid are fused. The proximal plate of the scapula is gently concave and short anteroposteriorly, forming the coracoid articulation anteriorly, the glenoid ventrally and the acromial process dorsally. The acromial process is present as a small, triangular process arising gradually from the scapular blade. In most stegosaurs, the acromial process is a rectangular process that projects dorsally to form the proximal plate of the scapula (Kentrosaurus, Hennig, Citation1915; Stegosaurus, Maidment et al., Citation2015; Hesperosaurus, Maidment et al., Citation2018; Miragaia, Mateus et al., Citation2009), but in Huayangosaurus it is less well-developed and forms a triangular process (Zhou, Citation1984; Maidment et al., Citation2006). The acromial process of Bashanosaurus primitivus is even less well-developed than that of Huayangosaurus, similar to that of the basally branching thyreophoran Scelidosaurus (Maidment et al., Citation2006). The scapular blade is slender and expands dorsoventrally at its posterior end (the ratio of the maximum height of blade to the minimum height of blade is about 3.4; Table S5), in contrast to the scapulae of other stegosaurs, in which the blade is relatively deeper dorsoventrally and does not expand distally (e.g., Kentrosaurus, Hennig, Citation1915; Stegosaurus, Maidment et al., Citation2015; Hesperosaurus, Maidment et al., Citation2018; Miragaia, Mateus et al., Citation2009; Huayangosaurus, Zhou, Citation1984; Maidment et al., Citation2006).
FIGURE 5. A, lateral view of right scapula and coracoid referred to Bashanosaurus primitivus (CLGPR V00006-1) from the Middle Jurassic of China. B, C, E, dermal armor referred to Bashanosaurus primitivus (CLGPR V00006-1) from the Middle Jurassic of China. D, dermal armor referred to Bashanosaurus primitivus (CLGPR V00006-2) from the Middle Jurassic of China. B–D, lateral view of plates; E, lateral view of spine. Abbreviations: ap, acromial process; bl, scapula blade; cor, coracoid; glf, glenoid fossa; pp, scapula proximal plate.
Coracoid
The right coracoid is well preserved aside from part of its dorsal margin (). The coracoid is fused to the scapula. In lateral view, the coracoid is flat and has a sub-circular outline. There is no coracoid foramen. A coracoid foramen was not present in the scapulocoracoid of Huayangosaurus either (Maidment et al., Citation2006). There is a distinct corner on the anteroventral margin of the coracoid, which projects slightly ventral to the ventralmost extent of the glenoid fossa and forms a very small sternal process. A sternal process is not present on the coracoid of Stegosaurus (Maidment et al., Citation2015), but is present on the coracoid of Huayangosaurus (Zhou et al., Citation1984). The ratio of the proximal plate area to coracoid area is about 0.46, in contrast to the condition in most other stegosaurs in which the coracoid is smaller than the proximal plate (Galton and Upchurch, Citation2004).
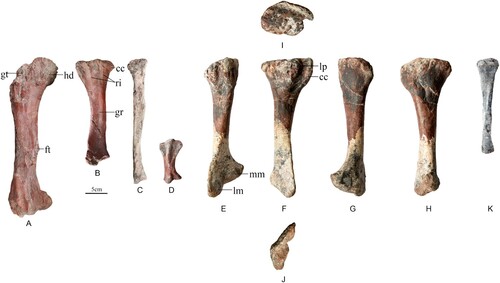
Femur
The left femur of CLGPR V00006-1 is crushed and only the posterior surface of the shaft can be seen (). The shaft of the femur is straight and about 370 mm long. The femoral head angles medially and slightly dorsally. The ventral margin of the femoral head is continuous with the medial margin of the shaft, describing a smooth curve. The greater trochanter lateral to the femoral head is slightly damaged. The anterior trochanter is fused to the greater trochanter, as in Huayangosaurus (Zhou, Citation1984), Kentrosaurus (Hennig, Citation1915), and Dacentrurus (Galton, Citation1990). The incomplete fourth trochanter is a rugose ridge on the posteromedial margin and forms a distinct process, different to Kentrosaurus (Hennig, Citation1915), Dacentrurus (Galton, Citation1990), and Stegosaurus (Maidment et al., Citation2015), in which the fourth trochanter is very poorly developed or absent. The fourth trochanter is positioned below the middle of the shaft (Tables S6 and S7), different to Huayangosaurus (Zhou, Citation1984), and Kentrosaurus (Hennig, Citation1915) where it is positioned at midshaft. The distal end of the femur bears a deep intercondylar groove that separates the two femoral epicondyles. The medial epicondyle has a bluntly rounded sub-elliptical outline with the long axis extending anteroposteriorly and it extends further posteriorly than the lateral epicondyle. The lateral epicondyle is slightly damaged.
FIGURE 6. A–D, hind limb referred to Bashanosaurus primitivus (CLGPR V00006-1) from the Middle Jurassic of China. A, left femur in posterior view; B, left tibia in medial view; C, left fibula in lateral view; D, left metatarsal III in lateral view. E–J, right tibia referred to Bashanosaurus primitivus (CLGPR V00006-2) from the Middle Jurassic of China. E, anterior; F, lateral; G, posterior; H, media; I, dorsal; J, ventral. K, lateral view of fibula referred to Bashanosaurus primitivus (CLGPR V00006-2) from the Middle Jurassic of China. Abbreviations: cc, cnemial crest; ft, fourth trochanter; gr, groove; gt, greater trochanter; hd, head; lm, lateral malleolus; lp, lateral process; mm, medial malleolus; ri, ridge.
Tibia
Two tibiae are preserved; our description focuses on the better-preserved right tibia of CLGPR V00006-2 (). In anterior view, the tibia comprises a straight shaft with a transversely compressed proximal end and a transversely expanded distal end. In dorsal view, the proximal surface is flat and is sub-oval in outline. The cnemial crest is a sharp ridge. The posterior surface of the proximal end forms an oval posterior process. The lateral (fibular) process is a large, round protuberance and situated in the middle of the proximal end, overhanging the lateral surface of the shaft. In lateral view the proximal end is expanded asymmetrically so that the cnemial crest extends further anteriorly than the posterior process does posteriorly. On the left tibia, the medial face of the proximal end bears two shallow, dorsoventrally oriented ridges that extend distally to bound a groove in the midshaft region. The shaft of bone is slender, twisted and has an oval-shaped cross-section. In medial view, the proximal of shaft is convex. The expansion degree of the distal end is the same as that of the proximal end. In ventral view, the medial malleoli and lateral malleoli are separated by a groove. The anterior surface of the lateral malleolus is flat for articulation with the fibula, is situated anteriorly to the medial malleolus and extends further ventrally than the medial malleolus. The medial surface of medial malleolus is convex. In posterior view, the two malleoli are flat and divided by a ridge. In ventral view, the distal end has a subtriangular outline. In morphology, the tibia is similar to that of other stegosaurs (Maidment et al., Citation2015).
Fibula
Two fibulae are preserved. The left fibula belongs to CLGPR V00006-1; it has suffered some crushing and is only exposed in lateral view (). The proximal and distal ends are slightly expanded anteroposteriorly. The proximal of the shaft is flat. The shaft is straight, slender, and slightly twisted along its length. The right fibula belongs to CLGPR V00006-2; it is similar to the left one of CLGPR V00006-1, and the distal end is missing ().
Metatarsal
Left metatarsal III of Bashanosaurus primitivus is well-preserved in CLGPR V00006-1 (). In lateral view, the proximal and distal ends are strongly expanded dorsoventrally. The ventral part of the proximal end is expanded more than the dorsal part. The middle surface of the proximal end has a well-developed ridge that extends beyond the dorsal surface onto the middle of the shaft. The distal end is expanded less than the proximal end. A deep groove is present at the distal end. The proximal and distal ends are joined by a waisted shaft.
Dermal Armor
Three plates and one spine were recovered. However, they are all incomplete. Two plates () and one spine () belong to CLGPR V00006-1. One plate () belongs to CLGPR V00006-2. The plates and spines are elongated dorsoventrally and compressed transversely. The plates have a rectangular outline, and the lateral surfaces of the plates are concave. Although the plates are all incomplete, the bases of plates are strongly convex and transversely thickened, and are separated from the anterior/posterior margins of the plates by a distinct groove in lateral view. This contrasts with the condition in Huayangosaurus (Zhou, Citation1984), in which the plates have an arc-shaped outline. The base of the spine is missing. In lateral view, it is smooth, tall, and slender. The dermal armor measurements can be found in Tables S8 and S9.
STEGOSAURIA gen. et sp. indet.
Specimen—A partial cranium, a dorsal vertebra, and a piece of dermal armor are preserved in CLGPR V00006-4. The specimen is from a ∼2 m2 exploratory trench that is about 900 m away from the Dinosaur Fossil Wall, and the fossil-bearing layer of these specimens is about 50 m higher stratigraphically than the Dinosaur Fossil Wall (i.e., the fossil-bearing layer of Bashanosaurus primitivus). Because Huayangosaurus is also known from the Lower Member of the Shaximiao Formation (Zhou, Citation1984), and CLGPR V00006-4 bears none of the autapomorphies of either Bashanosaurus primitivus or Huayangosaurus, we do not refer this specimen to either taxon.
Locality and Horizon—Laojun Village (108°55′25.86′′E, 30°50′45.44′′N), Pu’an Township, Yunyang County, Chongqing Municipality, People’s Republic of China; Lower Member of the Shaximiao Formation, Middle Jurassic.
DESCRIPTION
Posterior Cranium
The partial cranium consists of frontals, parietals, and a braincase, which are fused together (Table S10). In anterior view (), the ventral surface of frontals consists of the cavity for cranial nerve I and a concave area lying lateral to it. They are separated by a thin orbitosphenoid whose distal end is curved to form the opening for cranial nerve I. The orbitosphenoid forms the sidewall of the narrow part of the otic tract and the adjacent part of the cerebrum. The lower part is the braincase.
FIGURE 7. A–F, braincase referred to Stegosauria gen. et sp. indet. (CLGPRV0005-4) from the Middle Jurassic of China. A, anterior; B, left lateral; C, right lateral; D, posterior; E, dorsal; F, ventral. G–L, dorsal vertebrae referred to Stegosauria gen. et sp. indet. (CLGPRV0005-4) from the Middle Jurassic of China. G, anterior; H, left lateral; I, dorsal; J, posterior; K, right lateral; L, ventral. M–R, dermal armor referred to Stegosauria gen. et sp. indet. (CLGPRV0005-4) from the Middle Jurassic of China. M, anterior; N, posterior; O, left lateral; P, right lateral; Q, dorsal; R, ventral. Abbreviations: boc, basioccipital; bpt, basipterygoid process; bsp, basiphenoid; cc, carotid canal; c.n.I, III, V, VII, XII exit for cranial nerves; dia, diapophysis; f, frontal; fime, fissura metotica; fm, foramen magnum; fo, foramen ovale; ipzs, intraprezygapophyseal shelf; ist, integumentary structure; nc, neural canal; ns, neural spine; oc, occipital condyle; os, orbitosphenoid; p, parietal; para, parapophysis; poc, paroccipital process; pozyg, postzygapophysis; przyg, prezygapophysis; ps, parasphenoid; soc, supraoccipital; sf, supratemporal fossae; sq, squamosal.
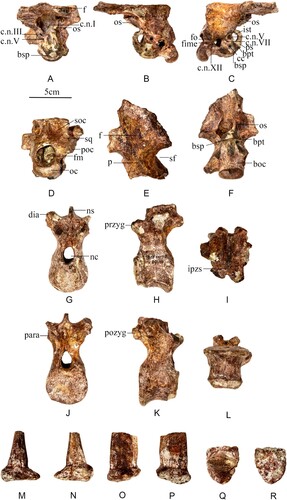
In lateral view (), the braincase is ovoid in shape. The basisphenoid extends less far ventrally than the occipital condyle, differing from the condition in Stegosaurus and Hesperosaurus (Carpenter et al., Citation2001). There is a carotid canal between the basisphenoid and the basipterygoid processes. The parasphenoid lies anterior to the basipterygoid processes, forming a circular fenestra, and it is fused to the ventral surface of the orbitosphenoid. There are three small foramina that represent the exits of the hypoglossal nerve (XII), as in the basally branching ornithischian Lesothosaurus (Sereno, Citation1991). Anterior to cranial nerve XII, there is a large, heart-shaped foramen, which is the combined fissura metotica and fenestra ovalis. The basisphenoid extends anteroventrally and lies ventral to this foramen. Anterior to the fenestra ovalis, a small foramen at the same level as the fenestra ovalis is for the facial nerve (VII). Anterior to VII, there are two foramina, presumably for cranial nerve III anteriorly and cranial nerve V posteriorly. The foramen for III is separated from that for V by a dorsoventrally extending ridge.
In posterior view (), the occipital condyle projects posteriorly, as in other stegosaurs (Hesperosaurus, Carpenter et al., Citation2001; Huayangosaurus, Zhou, Citation1984; Kentrosaurus, Hennig, Citation1925). The occipital condyle is ovoid (laterally wider than dorsoventrally tall) as in Huayangosaurus (Zhou, Citation1984) and Kentrosaurus (Hennig, Citation1925). The foramen magnum is circular, and is approximately similar in transverse width as the occipital condyle, but is much higher than the occipital condyle dorsoventrally, different to Hesperosaurus (Carpenter et al., Citation2001), but similar to Huayangosaurus (Zhou, Citation1984), Kentrosaurus (Hennig, Citation1925) and Stegosaurus (Carpenter et al., Citation2001). The incomplete squamosal extends laterally. The supraoccipital is concave and bears a transversely thick ridge on the midline that extends dorsoventrally. The incomplete squamosal lies lateral to the supraoccipital.
In dorsal view (), the frontal is flat, and the traces of the suture with the parietal are invisible, different to Tuojiangosaurus, and Kentrosaurus, in which the suture is visible (Maidment et al., Citation2018). The parietal is flat and expands anteroposteriorly, as in Stegosaurus, Huayangosaurus, and Hesperosaurus (Maidment et al., Citation2018). It narrows in its middle part and the margins are concave, forming the medial margins of the supratemporal fossae.
In ventral view (), the basioccipital forms the posterior floor of the braincase from the posterior part of the basisphenoid tubera posteriorly and it contributes to the spherical occipital condyle. Anterior to the occipital condyle, the basioccipital narrows transversely into a short neck, which is anteroposteriorly longer than the occipital condyle, differing from the condition in Huayangosaurus, Tuojiangosaurus, and Kentrosaurus (Maidment et al., Citation2018). The suture between the basisphenoid and basioccipital cannot be observed, as in other stegosaurs (Maidment et al., Citation2018). The basisphenoid forms the floor of the braincase from the rugose basisphenoid tubera to the basipterygoid processes. The basisphenoid is triangular with the apex pointing anteriorly.
Dorsal Vertebrae
One dorsal vertebra is preserved (). The centrum is 41 mm long and 46 mm wide. The diapophyses and neural spine are missing. It is very similar to the dorsal vertebrae of CLGPR V00006-3, differing from them only in the following respects. In posterior view, the postzygapophyses are angled posterolaterally at ∼45° from horizontal. Although the parapophysis is partially preserved, it is not elevated on a distinct stalk, which is an autapomorphy of Bashanosaurus primitivus. The parapophyses of posteriordorsal vertebrae project more obviously than the parapophyses of anterior dorsals in Huayangosaurus (Zhou, Citation1984), but there is little difference in the degree of projection along the column in Stegosaurus (Maidment et al., Citation2015). The lack of elevation of the parapophysis in CLGPR V00006-3 could therefore be related to location along the dorsal vertebral column, or a taxonomic difference. Only with more complete material can these options be assessed.
Dermal Armor
One spine was recovered (). The spine is elongated dorsoventrally and compressed transversely. The ventral surface of the base of the spine is flat, and the spine has gently convex edges laterally and sharp edges anteriorly and posteriorly. The edges are much more well-developed than these of Huayangosaurus (Zhou, Citation1984). It is much smaller than the spines associated with Bashanosaurus primitivus, although only few are known. The dermal armor measurements can be found in Tables S8 and S9.
PHYLOGENETIC RESULTS
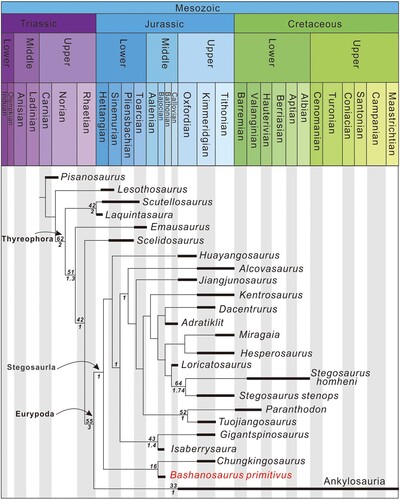
A single most parsimonious tree was recovered (); a second round of TBR branch swapping on trees held in RAM resulted in no further MPTs. The MPT was 269.61 steps in length, had a consistency index of 0.606 and a retention index of 0.664. Bashanosaurus primitivus is recovered in a clade with Chungkingosaurus, a taxon from the Upper Member of the Shaximiao Formation, and together these taxa are the earliest-diverging stegosaurs. To further explore the phylogenetic position of Bashanosaurus primitivus, four topological constraints were enforced in a series of subsequent analyses that otherwise used the same settings as the original analysis. First, the monophyly of all stegosaurs excluding Bashanosaurus was enforced. In this analysis, Bashanosaurus is found to lie basal to all other stegosaurs in a single MPT of length 269.65 steps. This is only slightly longer than the MPT of the original analysis and enforces the result that Bashanosaurus is a very early diverging stegosaur. The second constrained analysis enforced Bashanosaurus to fall within Ankylosauria and recovered a single MPT of 271.73 steps. The third analysis constrained Bashanosaurus to fall outside of Eurypoda as a basally branching thyreophoran and recovered a single MPT of 271.42 steps. Finally, because a sister-taxon relationship between Huayangosaurus and Chungkingosaurus has been recovered in previous analyses of this matrix (Maidment et al., Citation2020), and the fact that Bashanosaurus and Huayangosaurus are found in the same formation, Huayangosaurus, Chungkingosaurus, and Bashanosaurus were constrained to form a clade. This analysis returned a single MPT of length 270.8 steps.
FIGURE 8. A single most parsimonious tree recovered by the phylogenetic analysis. The numbers below the line are Bremer supports, the numbers above the line are GC frequencies. Ankylosauria was represented by three taxa, Gastonia, Euoplocephalus, and Sauropelta in the analysis, but these are shown here as a single terminal taxon for clarity.
GEOCHRONOLOGICAL RESULTS
The zircons from sample HP02-54 yielded Th/U ratios between 0.06–1.79 (only one grain is less than 0.1), which indicates that they are compatible with a magmatic origin. 107 grains of this sample gave 98 concordant grain ages. The age groups include ∼160–180 Ma, 240–260 Ma, 420–450 Ma, 1730–1900 Ma, and 2400–2600 Ma. The youngest zircons were used to calculate weighted average of 169 ± 0.65 Ma (n = 6, MSWD = 2.05). The zircons from sample PM02-361 yielded Th/U ratios between 0.1–1.98 that are compatible with a magmatic origin. 102 grains of this sample gave 98 concordant ages. The age groups include ∼160–180 Ma, 240–260 Ma, 420–440 Ma, 1730–1900 Ma, and 2400–2600 Ma. The youngest zircons were used to calculate a weighted average of 169 ± 0.68 Ma (n = 9, MSWD = 2.2). Since the zircon grains from samples HP02-54 and PM02-361 are detrital, they represent the maximum depositional age of the horizons just below and above the quarry, respectively, and indicate that the quarry was deposited no earlier than the Bajocian stage of the Middle Jurassic.
DISCUSSION
The geochronological results we report herein find the age of the bone-bearing layers of the Lower Member of the Shaximiao Formation to be Middle Jurassic at Pu’an township. In contrast, a recent study has suggested that the Lower Member of the Shaximiao Formation at Zigong is Upper Jurassic (Wang et al., Citation2018). Provenance analysis has shown that the depocenter of the Sichuan Basin was mainly in northeast Sichuan during the Middle Jurassic period (Shao et al., Citation2016); this indicates that the time limit of deposition in Yunyang may be earlier than that in Zigong. The intracontinental subduction of the Qinling orogenic belt and Daba Mountains (Dong et al., Citation2006), the heterogeneous distribution of bedrock thermochronology (Enkelmann et al., Citation2007), and the underlying conglomerate of the Qianfoya Formation (Luo et al., Citation2014) indicate that there may have been a large amount of denudation in the Jurassic period, which resulted in rapid filling of the basin. This evidence, combined with zircon dating results from the Shaximiao Formation in different locations (see Introduction and reported herein), indicates that the Shaximiao Formation may be laterally diachronous. As a result of material filling, the depocenter gradually migrated to the direction of the paleo-uplift region in central Sichuan. Therefore, it is possible that the beds in which Huayangosaurus were found are younger than those bearing Bashanosaurus. However, further research on the age of the Lower Member of the Shaximiao Formation at different localities across the Sichuan Basin is required to determine if this is the case.
Bashanosaurus primitivus differs from the coeval Huayangosaurus (Zhou, Citation1984) in several anatomical characters. In Bashanosaurus primitivus, the parapophyses of the dorsal vertebrae are elevated on a short stalk and the elevation is greater than that seen in Huayangosaurus (Zhou, Citation1984). In the anterior caudal vertebrae of Bashanosaurus primitivus, the transverse processes extend laterally and slightly dorsally in anterior view, in contrast to Huayangosaurus (Zhou, Citation1984), in which the transverse processes extend laterally and slightly ventrally. The prezygapophyses overhang the anterior margin of the centrum to a greater degree than that seen in Huayangosaurus (Zhou, Citation1984). In the pectoral girdle, the scapular blade flares distally and is more slender than Huayangosaurus (Zhou, Citation1984). The acromial process of the scapula is also smaller and more poorly developed than that of Huayangosaurus (Zhou, Citation1984). The area of the proximal plate-to-coracoid ratio is much larger in Bashanosaurus than in Huayangosaurus (Zhou Citation1984). On the femur, the 4th trochanter is positioned below the middle of the shaft in Bashanosaurus, in contrast to Huayangosaurus (Zhou, Citation1984) in which the 4th trochanter is positioned at midshaft. The bases of the plates of Bashanosaurus primitivus are strongly convex and transversely thickened, and are separated from the anterior/posterior margins of the plates by a distinct groove, different from the condition in Huayangosaurus (Zhou, Citation1984) in which the plates have an arc-shaped outline.
Loricatosaurus has an autapomorphy that is absent in Bashanosaurus primitivus: the anterior caudal vertebrae possess chevron facets that converge ventrally so that the vertebral centra are ‘v’ shaped in lateral view. In addition, the fourth trochanter of femur is poorly developed in Loricatosaurus and the dorsal plates are generally transversely thin, except at the base. Only a few bones are known from Adratiklit (Maidment et al., Citation2020), but its dorsal vertebrae have a small, triangular, rugose protuberance situated dorsally on the prezygapophysis posterior to the articular facet, and anterior centroparapophyseal laminae drawn into anteriorly-projecting rugosities either side of the neural canal. These autapomorphies of the dorsal vertebrae are absent in Bashanosaurus primitivus. The dorsal neural arch pedicles are also dorsally elongate in both Adratiklit and Loricatosaurus, contrasting with the condition in Bashanosaurus primitivus. The postcranial anatomy of Isaberrysaura has yet to be described. Bashanosaurus primitivus can therefore be distinguished from other Middle Jurassic stegosaurs, and clearly represents a new taxon based on autapomorphies of the dorsal vertebrae, scapulocoracoid, femur, and dermal armor.
In our MPT, Bashanosaurus primitivus is recovered in a clade with Chungkingosaurus, and together they are the earliest-diverging stegosaurs. The constrained analysis in which Bashanosaurus was forced to lie outside of a clade of other stegosaurs was only very slightly less parsimonious, raising the possibility that the taxon may represent the earliest-diverging stegosaur, although further material of both Chungkingosaurus and Bashanosaurus primitivus is required to further test the exact phylogenetic relationships of these taxa. This accords with the stratigraphic position of Bashanosaurus primitivus as one of the earliest stegosaurs known worldwide. Bashanosaurus primitivus is from the Bajocian, making it the oldest stegosaur, along with Isaberrysaura, which has yet to be fully described. Bashanosaurus primitivus is united with Thyreophora by the presence of dermal armor. It is a euryopodan (Stegosauria + Ankylosauria) because it possesses fusion of the anterior and greater trochanter. It shares with other stegosaurs postzygapophyses that do not extend posteriorly over the posterior articular facet of caudal vertebrae. However, Bashanosaurus primitivus bears a number of plesiomorphic characteristics compared with other stegosaurs. A dorsal process on the transverse processes of anterior caudal vertebrae is absent in Bashanosaurus primitivus, along with early-branching thyreophorans and other early-diverging stegosaurs (Huayangosaurus and Gigantspinosaurus). Bashanosaurus primitivus also possesses elongate posterior caudal vertebrae, which it plesiomorphically shares with early thyreophorans and early-diverging stegosaurs (Huayangosaurus, Chungkingosaurus, and Gigantspinosaurus). Finally, it possesses an acromial process of the scapula that is convex upwards dorsally, a feature shared with basally branching thyreophorans and the early-diverging stegosaur Huayangosaurus. Furthermore, the elevation of the parapophysis of dorsal vertebrae on stalk-like processes is similar to the condition in the early-diverging thyreophoran Scelidosaurus (NHMUK PV R1111). Bashanosaurus primitivus and Chungkingosaurus are united in a clade that is supported by continuous data, primarily the ratio of the height of the centrum to the neural canal.
Bashanosaurus primitivus, Chungkingosaurus, and Huayangosaurus are the earliest-diverging stegosaurs and Bashanosaurus is also one of the oldest. They are all from the Middle to Late Jurassic Shaximiao Formation of China, a pattern that suggests that stegosaurs may have originated in Asia. However, the fossil record of early-diverging stegosaurs is sparse and likely to be biased by a lack of well-known Middle Jurassic terrestrial dinosaur-bearing units worldwide (SCRM pers. obs.).
The length of Huayangosaurus is about 3.6 m and the length of its femur is about 47 cm (Zhou, Citation1984). Based on the hindlimb size of Bashanosaurus primitivus, we estimate its total length is about 2.8 m. It is possible that this small size indicates that the holotype may be a subadult, an inference supported by a lack of fusion between the scapula and coracoid and the tibia and fibula; fusion between these elements is seen in adults of Stegosaurus but not in juveniles (Galton, Citation1982). However, the anterior and greater trochanters of the femur, and the scapula and coracoid, are fused in Bashanosaurus, features only observed in adult individuals of Stegosaurus (Galton, Citation1982). Unfortunately, due to a paucity of specimens representing multiple growth stages in stegosaurs other than Stegosaurus, the robusticity of skeletal fusion and size characteristics to assess ontogeny in stegosaurs is unknown, and thus we are not able to adequately assess the ontogenetic stage of Bashanosaurus at this time.
CONCLUSIONS
Bashanosaurus primitivus is the earliest record of Stegosauria in Asia and represents one of the earliest records of this clade from anywhere in the world. Geochronological data and analysis support a Middle Jurassic (Bajocian) age for the Shaximiao Formation in the vicinity of Pu’an Township, China. Phylogenetic analysis indicates that Bashanosaurus primitivus is the earliest-diverging stegosaur along with Chungkingosaurus supporting an Asian origin for Stegosauria based on currently available data.
Supplemental Material
Download Zip (86.5 KB)ACKNOWLEDGMENTS
We thank H. L. You (Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing) for helpful discussion and offering many valuable suggestions, Y. C. Lu for coordinating the fieldwork in Yunyang, X. S. Han for collecting the specimen, Y. Q. Zhang for preparing the specimen, Q. Li for helping with photography, Y. Ye, S. Jiang for providing information on Huayangosaurus taibaii of Zigong Dinosaur Museum. This work is supported by Natural Science Foundation of Chongqing, China (No. cstc2020jcyj-msxmX0082), Postdoctoral Project of Chongqing, China (No. Xm2017069) and Project of Dinosaur Fossil Protection and Research of Chongqing Planning and Natural Resources Bureau (No. kj-2018035). The corresponding authors contributed equally to this work.
Related Research Data
LITERATURE CITED
- Carpenter, K. 2010. Species concepts in North American stegosaurs. Swiss Journal of Geosciences 103:155–162.
- Carpenter, K., and P. M. Galton. 2001. Othniel Charles Marsh and the myth of the eight-spiked Stegosaurus; pp. 76–102 in K. Carpenter (eds.), The Armored Dinosaurs. Indiana University Press, Bloomington, Indiana.
- Carpenter, K., C. A. Miles, and K. Cloward. 2001. New Primitive Stegosaur from the Morrison Formation, Wyoming; pp. 55–75 in K. Carpenter (eds.), The Armored Dinosaurs. Indiana University Press, Bloomington, Indiana.
- Cobos, A., R. Royo-Torres, L. Luque, L Alcalá, and L. Mampel. 2010. An Iberian stegosaurs paradise: the Villar del Arzobispo Formation (Tithonian-Berriasian) in Teruel (Spain). Palaeogeography, Palaeoclimatology, Palaeoecology 293:223–236.
- Dai, H., R. Benson, X.-F. Hu, Q.-Y. Ma, Ch. Tan, N. Li, M. Xiao, H.-Q. Hu, Y.-X. Zhou, Zh.-Y. Wei, F. Zhang, Sh. Jiang, D.-L. Li, G.-ZH. Peng, Y.-L. Yu, and X. Xu. 2020. A new possible megalosauroid theropod from the Middle Jurassic Xintiangou Formation of Chongqing, People’s Republic of China and its implication for early tetanuran evolution. Scientific Reports 10:139.
- Dong, S.-W., J.-M. Hu, W. Sh, Zh.-Y. Zh, G. L. 2006. Jurassic superposed folding and Jurassic foreland in the Daba Mountain, Central China. Acta Geoscientia Sinica 27:403–410. [In Chinese with English abstract]
- Enkelmann, E. A. Weislogel, L. Ratschbacher, E. Eide, A. Renno, and J. Wooden. 2007. How was the Triassic Songpan-Ganzi basin filled? A provenance study. Tectonics 26:TC4007.
- Galton, P. M. 1982. Juveniles of the stegosaurian dinosaur Stegosaurus from the Upper Jurassic of North America. Journal of Vertebrate Paleontology 2:47–62.
- Galton, P. M. 1985. British plated dinosaurs (Ornithischia, Stegosauridae). Journal of Vertebrate Paleontology 5:211–254.
- Galton, P. M. 1990. A partial skeleton of the stegosaurian dinosaur Lexovisaurus from the uppermost Lower Callovian (Middle Jurassic) of Normandy, France. Geologica et Paleontologica 24:185–199.
- Galton P. M. 1991. Postcranial remains of the stegosaurian dinosaur Dacentrurus from the Upper Jurassic of France and Portugal. Geologica et Paleontologica 25:299–327.
- Galton, P. M., and K. Carpenter. 2016. The plated dinosaur Stegosaurus longispinus Gilmore, 1914 (Dinosauria: Ornithischia; Upper Jurassic, western USA), type species of Alcovasaurus n. gen. Neues Jahrbuchfür Geologie und Paläontologie-Abhandlungen 279:185–208.
- Galton, P. M., and P. Upchurch. 2004. Stegosauria; pp. 343–362 in P. M. Galton (eds.), The Dinosauria. University of California Press, Berkeley.
- Goloboff, P. A., J. S. Farris, and K. C. Nixon. 2008. TNT, a free program for phylogenetic analysis. Cladistics 24:774–786.
- Gou, Z. H., B. Zhao, and S. Wu. 2000. Jurassic system of Dayi, Chongzhou, Wenchuan and Dujiangyan, Sichuan. Journal of Chengdu University of Technology 27:31–39.
- Gu X. D., and X. H. Liu. 1997. Lithostratigraphy of Sichuan. China University of Geosciences Press, Hubei, 1-417pp. [In Chinese]
- Hennig, E. 1915. Kentrosaurus aethiopicus, der stegosauridae des Tendaguru. Sitzungs-berichte der Gesellschaft Naturforschender Freunde zu Berlin 219–247.
- Hennig, E. 1925. Kentrurosaurus aethiopicus; die Stegosaurierfunde vom Tendaguru, Deutsch-Ostafrika. Palaeontographica-Supplementbände (Supplement 7):101–254.
- Hu, Z.-CH., W. Zhang, Y.-SH. Liu, SH. G, M. Li, K.-Q. Zong, H.-H. Ch, and Sh.-H. Hu. 2015."Wave"Signal-Smoothing and Mercury-Removing Device for Laser Ablation Quadrupole and Multiple Collector ICPMS Analysis: Application to Lead Isotope Analysis. Analytical Chemistry 87:1152–1157.
- Li, N., H. Dai, Ch. T, X.-F. Hu, Zh.-Y. Wei, Y. L, G.-B. Wei, D.-L. Li, L. M, B.-Q. H, H.-L. Y, and G.-Zh. Peng. 2019. A neornithischian dinosaur from the Middle Jurassic Xintiangou Formation of Yunyang, Chongqing, China: the earliest record in Asia. Historical Biology DOI:10.1080/08912963.2019.1679129.
- Li R. B., X.-Z. Pei, Z.-Q. Liu, Z.-C. Li and X.-L. Wang. 2010. Basin-Mountain Coupling Relationship of Foreland Basins between Dabashan and Northeastern Sichuan––The Evidence from LA-ICP-MS U-Pb Dating of the Detrital Zircons. Acta GeologicaSinica 84:1118–1134. [In Chinese with English abstract]
- Liu, Y.-Sh., Zh.-Ch. Hu, Sh. Gao, D. Günther, J. Xu, Ch.-G. Gao, and H.-H. Chen. 2008. In situ analysis of major and trace elements of anhydrous minerals by LA-ICP-MS without applying an internal standard. Chemical Geology 257:4–43.
- Luo, L., J.-F. Qi, M.-Zh. Zhang, K. Wang, and Y.-Zh. Han. 2014. Detrital zircon U–Pb ages of Late Triassic–Late Jurassic deposits in the western and northern Sichuan Basin margin: constraints on the foreland basin provenance and tectonic implications. International Journal of Earth Sciences 103:1553–1568.
- Maidment, S. C. R., C. Brassey, and P. M. Barrett. 2015. The postcranial skeleton of an exceptionally complete individual of the plated dinosaur Stegosaurus stenops (Dinosauria: Thyreophora) from the Upper Jurassic Morrison Formation of Wyoming, U.S.A. PLoS One 10:e0138352.
- Maidment, S. C. R., G. B. Wei, and D. B. Norman. 2006. Re-description of the postcranial skeleton of the Middle Jurassic stegosaur Huayangosaurus taibaii. Journal of Vertebrate Paleontology 26: 944–956.
- Maidment, S. C. R., D. C. Woodruff, and J. R. Horner. 2018. A new specimen of the ornithischian dinosaur Hesperosaurus mjosi from the Upper Jurassic Morrison Formation of Montana, USA, and implications for growth and size in Morrison stegosaurs. Journal of Vertebrate Paleontology 38:e1406366.
- Maidment, S. C. R., D. B. Norman, P. M. Barrett, and P. Upchurch. 2008. Systematics and phylogeny of Stegosauria (Dinosauria: Ornithischia). Journal of Systematic Palaeontology 6:364–407.
- Maidment, S. C. R., T. J. Raven, D. Ouarhache, and P. M. Barrett. 2020. North Africa's first stegosaur: Implications for Gondwanan thyreophoran dinosaur diversity. Gondwana Research 77:82–97.
- Marsh, O. C. 1877. A new order of extinct Reptilia (Stegosauria) from the Jurassic of the Rocky Mountains. American Journal of Science 14:34–35.
- Mateus, O., S. C. R. Maidment, and N. A. Christiansen. 2009. A new long-necked ‘sauropod-mimic’ stegosaur and the evolution of the plated dinosaurs. Proceedings of the Royal Society of London B: Biological Sciences 276:1815–1821.
- Owen, R. 1842. Report on British fossil reptiles. Reports of the British Association for the Advancement of Science 11:60–204.
- Peng G. Z., Y.-H. Gao and Ch.-K. Shu. 2005. Jurassic Dinosaur Faunas in Zigong. Chengdu. Sichuan People’s Publishing House 236.
- Qian T., Sh.-F. Liu, Z.-X. Wang, W.-P. Li, and X.-L. Chen. 2016. A detrital record of continent-continent collision in the Early-Middle Jurassic foreland sequence in the northern Yangtze foreland basin, South China. Journal of Asian Earth Sciences 131:123–137.
- Salgado, L., J. Canudo, A. C. Garrido, M. M. Azanza,L. C. A. Martínez, R. A. Coria, and J. M. Gasca. 2017. A new primitive neornithischian dinosaur from the Jurassic of Patagonia with gut contents. Scientific Reports 7:42778.
- Seeley, H. G. 1887. The classification of the Dinosauria. Reports of the British Association for the Advancement of Science 58:698–699.
- Sereno, P. C. 1991. Lesothosaurus, “fabrosaurids,” and the early evolution of Ornithischia. Journal of Vertebrate Paleontology 11:168–197.
- Shao T., N. Cheng, and M. Song. 2016. Provenance and tectonic-paleogeographic evolution: Constraints from detrital zircon U–Pb ages of Late Triassic-Early Jurassic deposits in the northern Sichuan basin, central China. Journal of Asian Earth Science 127:12–31.
- Tan C., M. X, H. D, X.-F. Hu, N. Li, Q.-Y. Ma, Zh.-Y. Wei, H.-D. Yu, C. X, G.-Zh. Peng, Sh. J, X.-X. Ren and H.-L. You. 2020. A new species of Omeisaurus (Dinosauria: Sauropoda) from the Middle Jurassic of Yunyang, Chongqing, China. Historical Biology. DOI:10.1080/08912963.2020.1743286
- Wang, J., Y. Ye, R. Pei, Y. Tian, C. Feng, D. Zheng, and S. Chang. 2018. Age of Jurassic basal sauropods in Sichuan, China: a reappraisal of basal sauropod evolution. Geological Society of America Bulletin 130:1493–1500.
- Wang, Y., B.-H. Fu, X.-P. Xie, Q.-S. Huang, and Z.-K. Jiang. 2010. The Terrestrial Triassic and Jurassic Systems in the Sichuan Basin, China. University of Science and Technology of China Press, Anhui, 1–214pp.
- Wilson, J. A. 1999. A nomenclature for vertebral laminae in sauropods and other saurischian dinosaurs. Journal of Vertebrate Paleontology 19(4):639–653.
- Zhang, H., W. Yang, Y. Zhou, D. Li, H. Dai, C. Tan and H. Yu. 2020. Geochemical characteristics and geological significance of the dinosaur fossils burial clasticrocks in the member 1 of Shaximiao Formation in the Yunyang area, Chongqing. Mineralogy and Petrology 40: 26–35. [In Chinese].
- Zhou, S. 1984. The Middle Jurassic Dinosaurian Fauna from Dashanpu, Zigong, Sichuan. Sichuan Scientific and Technological Publishing House, Sichuan52. [In Chinese].
- Zong, H., and H. Shi. 1997. The Jurassic System of Dujiangyan, Pengzhou and Shifang, Sichuan. Journal of Stratigraphy 21:192–202. [In Chinese with English abstract]
- Zong, K., R. Klemd, Y. Yuan, Z. He, J. Guo, X. Shi, Y. Liu, Z. Hu, and Z. Zhang. 2017. The assembly of Rodinia: The correlation of early Neoproterozoic (ca. 900 Ma) high-grade metamorphism and continental arc formation in the southern Beishan Orogen, southern Central Asian Orogenic Belt (CAOB). Precambrian Research 290:32–48.