
ABSTRACT
With advances in molecular phylogeny, the Ursidae affinity of Ailuropoda is no longer controversial. However, the early evolution of Ailuropoda and its close relatives (the tribe Ailuropodini) is still unclear. In this study, we describe a new fossil discovery from Bulgaria, which represents a new taxon of Ailuropodini, ?Agriarctos nikolovi. The materials of Ailurarctos are restudied and the evolution and dispersal of Ailuropodini are discussed. Early Ailuropodini split into two lineages, one in Europe as Agriarctos (three species, whose assignment to the same genus is not certain), and one in southeastern Asia as Ailurarctos and later Ailuropoda. Ailurarctos is a paraphyletic group, with both known species as successive direct ancestors to Ailuropoda. Subtribe Ailuropodina is proposed here to include Ailurarctos and Ailuropoda. Turolian European Agriarctos paralleled with Ailuropodina in many aspects, which reflects similar adaptation towards a specific herbivorous diet.
INTRODUCTION
Ailuropodini are a group of peculiar bears that evolved towards a herbivorous diet, and the terminal taxon Ailuropoda is a bamboo specialist, with bamboo accounting for more than 99% of food components in its diet (Zhou et al., Citation1997). The tribe was first proposed by Hendey (Citation1980a), including Ailuropoda and “Ursavus” depereti. Hunt (Citation2004) added Ailurarctos in Ailuropodini, but excluded “Ursavus” depereti. Abella et al. (Citation2012) included Ailuropoda, Ailurarctos, and Agriarctos (including Agriarctos depereti), as well as their newly erected Kretzoiarctos in Ailuropodini. However, the early evolution of Ailuropodini is still poorly known due to scant materials (Abella et al., Citation2012). Phylogeny of Ailuropodini has been proposed by several authors, but most studies were only concentrated on proving certain taxa as Ailuropodini, not intra-tribe relationships (Abella et al., Citation2012), or briefly summarize the evolution without deep analyses (Hunt, Citation2004). None has concentrated in solving the evolution of members within Ailuropodini. While several lower dentitions of Turolian Ailuropodini have been known in Europe, few upper dentitions, especially the most characteristic P4, are known.
In this work, we describe two isolated teeth found in Bulgaria. One of the two is a well-preserved, unworn P4. Even though the material is scant, it brings important knowledge about the evolution of giant panda. All known members of early Ailuropodini are reviewed, with emphasis on the Tortonian–Messinian (Vallesian–late Turolian) and equivalent late Bahean and Baodean in eastern Asia species.
GEOLOGICAL BACKGROUND
Both teeth are from the old collections of the NMNHS and were probably received by the paleontologist of the Museum, Dr. Ivan Nikolov, before the 1980s. They were labeled as Guredjia without other indication. Guredjia is the old name of Ognyanovo village (Elin Pelin Municipality), situated in NW Bulgaria on the northern foothills of Ihtimanska Sredna Gora Mountains (). The coal-bearing sediments of the Novihan member of the Lozenets svita (= Lozenets formation) are revealed in several sites of the region, for example, at 1 km to the east from Novi Han (the holostratotype of the Novihan member of the svita, which is at about 10 km to the west from Ognyanovo, and also near Grigorevo, Elin Pelin, Kostinbrod, and Ravno Pole). The member is characterized by gray to gray-greenish sandy clays and several seams of black clays and also by several layers of lignite at the base of the member. The mollusks, macroflora, and pollen found give grounds to assume a late Pontian–early Dacian age of this member (Kamenov and Kojumdgieva, Citation1983; M. Yaneva, pers. comm.; Popov, Citation1993). The teeth are shiny (especially the enamel) and colored black from contact with coal, which would belong to the Novihan member. Given that the coal seams form the basis of the Novihan member, it is possible to assume a probable late Pontian (i.e., late Messinian) age of the fossil teeth. Such an age seems to be supported by the evolved morphology and size of the tooth.
FIGURE 1. Location of Ognyanovo in Bulgaria.
MATERIALS AND METHODS
The terminology and measurements of the teeth follow Jiangzuo et al. (Citation2019) and are shown in Fig. S1.
The following specimens have been examined for this study:
Ailurarctos yuanmouensis: FYV2509.1-5, from Yuanmou, Yunnan Province, China.
Ailurarctos lufengensis: IVPP V6892, IVPP V25032, KIZ ???, from Lufeng, Yunnan Province, China.
Ailuropoda melanoleuca, IOZ 32756, IOZ 26449, IOZ 32755, KIZ 011454, from Sichuan Province, China.
Ailuropoda microta IVPP V14564, from Jinyin Cave, Guangxi Province, China.
Agriarctos vighi, HNM Ob/5691, from Rózsaszentmárton, Heves County, Hungary.
Agriarctos gaali HNM V.60.1751, from Hatvan, Heves County, Hungary.
cf. Agriarctos depereti NMNHS FM-C3555 cast of MHNL AA52bis (P4), from Soblay, Auvergne-Rhône-Alpes, France; uncatalogued cast of MHNL AA52bis (P4) in the collection of the University of Vienna;
cf. Agriarctos depereti, NMNHS FM-C3556 cast of P4 of MHNL AA96 (P4-M1), from Soblay, Auvergne-Rhône-Alpes, France; NMNHS FM-C3557 cast of M1 of MHNL AA96 (P4-M1) from Soblay, France;
cf. Agriarctos depereti: uncatalogued cast of MHNL AA50-51 (M1-M2) from Soblay, Auvergne-Rhône-Alpes, France, in the collection of the University of Vienna;
?Agriarctos nikolovi sp. nov. NMNHS FM3546-a, b, from Ognyanovo, Sofia county, Elin Pelin municipality, Bulgaria.
Abbreviation of generic names—A., Ailuropoda; Ai., Ailurarctos; Ag., Agriarctos.
Institutional Abbreviations—HNM, Hungarian Natural History Museum, Budapest, Hungary; IOZ, Institute of Zoology, Chinese Academy of Sciences, Beijing, China; IVPP, Institute of Vertebrate Paleontology and Paleoanthropology, Beijing, China, collection catalog with Vas the initial character; KIZ, Kunming Institute of Zoology, Chinese Academy of Sciences, Kunming, China; MHNL, Museum d'histoire naturelle de Lyon, Lyon, France; NMNHS, National Museum of Natural History at the Bulgarian Academy of Sciences, Sofia, Bulgaria; FYV, collection from Yuanmou, now housed in Yunnan Institute of Cultural Relics and Archaeology, Kunming, China.
SYSTEMATIC PALEONTOLOGY
Order CARNIVORA Bowdich, Citation1821
Family URSIDAE Batsch, Citation1788
Subfamily AILUROPODINAE Grevé, Citation1892
Tribe AILUROPODINI Grevé, Citation1892
?AGRIARCTOS NIKOLOVI sp. nov.
()
Etymology—Named in honor of Dr. Ivan Nikolov, who has preserved the fossil teeth in the collections of the NMNHS and for his contributions to the study of the fossil mammal fauna of Bulgaria.
FIGURE 2. Teeth of ?Ag. nikolovi: A, NMNHS FM3546-b, upper canine in labial and lingual views; B, NMNHS FM3546-a, P4 in buccal, occlusal, and lingual views.
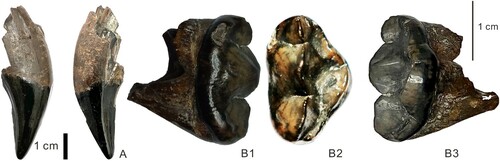
Holotype—Upper right carnassial tooth (NMNHS FM3546-a) and upper right canine (NMNHS FM3546-b; both numbers from the collection of the NMNHS), belonging to the same individual.
Age—Estimated late Messinian (i.e., latest Turolian) age (MN13).
Diagnosis—Derived Ailuropodini of large size. Canines proportionally very large. P4 with distinct parastyle, but not enlarged to metacone size. Inner lobe expanded with well-developed protocone and especially hypocone. Protocone and hypocone subdivided. Paracone and metacone under very slight angle. Buccal wall straight, without concavity in the middle.
Differential Diagnosis—Differs from Kretzoiarctos and Miomaci in having much stronger P4 parastyle and more complicated P4 inner lobe; differs from Ag. depereti in having larger P4 inner lobe and more developed small cusps in the inner lobe; differs from Ailurarctos in having smaller P4 parastyle.
Description—The canine is large and robust. A wear facet from the contact with the lower canine is seen on the base of the mesial surface of the tooth. The mesial and distal ridges are well marked: A well expressed sharp ridge is present on the distal surface of the C1. A similar, but slightly less pronounced ridge is present on the mesial surface, displaced slightly lingually. The labial surface of the tooth is the largest one. It is slightly convex. The lingual one is relatively flat. A visible rim separates the lingual surface from a third, short, flat to slightly concave postero-lingual surface, reaching distally the distal ridge.
The P4 is unworn, with a triangular shape. There is no distinct concavity in both antero- or posterior border of the inner lobe. The buccal wall is straight, not concave in the middle. Three large buccal cusps are present. The paracone is the dominant cusp, followed by metacone and then parastyle. The parastyle practically reaches the metacone in height, but is significantly shorter medio-distally than the metacone, which is rather elongated. The cutting edge of the three cusps is well developed. The paracone and metacone are oriented in a very slight angle. The medial faces of these three cusps are rather flat, whereas the lateral faces of parastyle and paracone are convex. The inner lobe is greatly extended to most of the medial border. There are two large cusps, representing protocone on the anterior side and hypocone on the posterior side. They are relatively mesially located on the inner lobe. The hypocone is clearly larger than the protocone. It is more distally placed (close to the paracone/metacone boundary) and presents several, weakly expressed corrugations. Both the protocone and hypocone are subdivided around their middle part.
COMPARISON
The P4 of ?Agriarctos nikolovi sp. nov. shows clear Ailuropodinae affinity, and in particular Ailuropodini affinity. The large and ridge-like parastyle, and greatly enlarged inner lobe with multiple cusps point to an affinity to either Agriarctos, Ailurarctos, or Ailuropoda. It differs from Indarctos and Agriotherium by its much larger and blade-like parastyle and more expanded and complex inner lobe.
Comparison with Kretzoiarctos and Miomaci
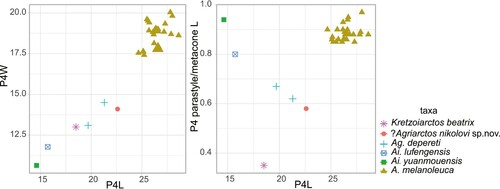
The tooth of ?Ag. nikolovi sp. nov. differs significantly from Kretzoiarctos: The P4 of Kretzoiarctos is smaller (). The parastyle is distinct in Kretzoiarctos (Abella et al., Citation2011) but it is relatively small, much smaller (shorter and lower) than that of the Bulgarian one. The inner lobe in Kretzoiarctos is smaller, with the only hypocone developed. Similar differences can be applied to Miomaci, which has a smaller parastyle and simpler inner lobe in a similar way (de Bonis et al., Citation2017).
TABLE 1. Measurements (in mm) of upper teeth of Ailuropodini. Measurements of K. beatrix are from Abella et al. (Citation2011, Citation2012); metric data for Ag. depereti from Soblay are from Crusafont and Kurtén (Citation1976), and others were measured by authors. Abbreviations: L, length; Max, maximum; Min, minimum; MN, Mammal Neogene zone; n, sample size; W, width.
Comparison with cf. Agriarctos depereti
Two P4s of cf. Agriarctos depereti (see below Discussion point 1–3 about the taxonomic status and the scope of this taxon) are known from Soblay (Viret, Citation1949). The Bulgarian P4 is similar to the two P4 of cf. Ag. depereti in having a similarly enlarged blade-like parastyle (). But the Bulgarian P4 has a parastyle nearly in line with the paracone, whereas the parastyle in the two P4s from Soblay turns slightly buccally. The paracone and metacone in Bulgarian P4 are nearly in line. The situation in cf. Ag. depereti is variable. There is no buccal concavity in the Bulgarian P4, which is present in both teeth of cf. Ag. depereti. The buccal cingulum is nearly absent in the Bulgarian P4, but well developed in cf. Ag. depereti.
FIGURE 3. Upper dentition (PA) of?Agriarctos nikolovi in comparison with other representatives of Ailuropodini. A, cf. Ag. depereti, uncatalogued cast of MHNL AA52bis in the collection of the University of Vienna; B, ?Ag. nikolovi NMNHS FM3546-a (reversed for comparison); C, Ai. yuanmouensis, YICRA YV2509.1 (reversed); D, Ai. lufengensis, IVPP V6892.12; E, A. microta, IVPP V14564; F1-3, A. melanoleuca, IOZ32756, IOZ26449, IOZ32755. Not to scale.
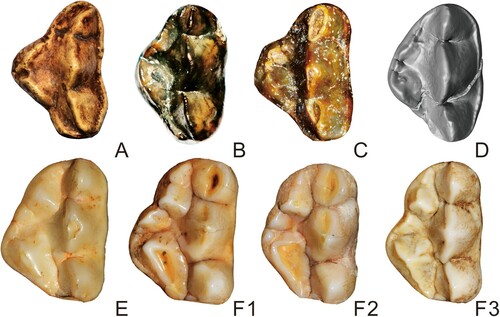
The inner lobe of the Bulgarian P4 is clearly more derived than that of cf. Ag. depereti. The Bulgarian P4 has a distinctly more expanded inner lobe, with clearly developed protocone and hypocone, both subdivided. In the P4 of cf. Ag. depereti, the inner lobe only expends around half of the tooth length, with the protocone located more posteriorly, and both protocone and hypocone are not subdivided. The Bulgarian tooth therefore is more derived than cf. Ag. depereti and also larger.
Comparison with Ailurarctos
Two species of Ailurarctos are known. The Bulgarian tooth is similar to the P4s of both species in having well-developed inner lobe cusps, with both protocone and hypocone subdivided. The parastyle and metacone also form a very small angle or in line in all three species, and the buccal concavity is indistinct or absent. The buccal wall of ?Agriarctos nikolovi is straight, not concave, in which it is close to the more derived Ailurarctos lufengensis.
However, the Bulgarian tooth is different from the two species of Ailurarctos in several aspects. The parastyle is distinctly smaller than the metacone in length in the Bulgarian tooth, whereas in both species of Ailurarctos, the parastyle is close to the metacone in length (). The Bulgarian tooth has no cingulum, which is present in both species of Ailurarctos. A small cusp is present between protocone and hypocone in both species of Ailurarctos, which is absent in the Bulgarian tooth, though the latter trait could be variable, as is seen in the situation of Ailuropoda.
FIGURE 4. Dental ratio of P4 of Ailuropodini. Abbreviations: L, length; W, width, in mm.
Comparison with Ailuropoda
The P4 of ?Agriarctos nikolovi is slightly larger than in Ailuropoda microta and smaller than other species of Ailuropoda (Pei, Citation1987), whereas its canine size is comparable to that of the largest individuals of Ailuropoda melanoleuca and larger than that of other species. The P4 is similar to that of Ailuropoda in the absence of the cingulum. The differences between the Bulgarian tooth and Ailuropoda can largely be referred to the differences between the Bulgarian tooth and Ailurarctos. The Bulgarian tooth further differs from Ailuropoda by a less enlarged hypocone.
The canine of ?A. nikolovi is very robust, comparable in size (in spite of the smaller dimensions of P4) to those of Ailuropoda melanoleuca.
DISCUSSION
Review of Ailuropodini and Definition of a New Subtribe Ailuropodina
Two tribes could be recognized in Ailuropodinae, i.e., Ailuropodini and Agriotheriini. Ailuropodini differs from Agriotheriini, according to our view, by a more enlarged and crest-like (not cusp-like) parastyle, which reaches the height of the metacone; and by a centrally situated (in Miocene forms) and complex inner lobe. The earliest possible member of Ailuropodini is Kretzoiarctos beatrix from Nombrevilla 2 and Abocador de Can Mata in Spain, with an age of MN8 (Abella et al., Citation2011, Citation2012). The materials from Nombrevilla 2 were reported by Abella et al. (Citation2011), including a P4 and a M1. The P4 develops a small but distinct cusp-like parastyle, which is characteristic of Ailuropodinae. In the following year, Abella et al. (Citation2012) described a broken mandible from Abocador de Can Mata with c-m2. The phylogenetic analysis of Abella et al. (Citation2012) suggested Kretzoiarctos was the sister group of Agriarctos, but as Qiu et al. (Citation2014) pointed out, the interrelationship of taxa within the Ailuropodini in their phylogeny is problematic, since Ailuropoda is located at the base of Ailuropodini in their phylogeny, which seems unlikely. Compared with Indarctos vireti, Kretzoiarctos had already developed the parastyle of P4, indicating that Kretzoiarctos probably had already diverged from the Indarctos lineage and evolved towards Ailuropodini; however, the discovery of early Agriotheriini Miomaci found in Rudabánya (Hungary, MN9), which has a weak but clearly defined parastyle in P4 (de Bonis et al., Citation2017), suggests that this character could be variable in early Ailuropodinae. The other teeth of Kretzoiarctos mostly remain conservative without typical Ailuropodini characters. These characters led Qiu et al. (Citation2014) to include Kretzoiarctos in Agriotheriini rather than Ailuropodini. Therefore, the evolutionary position of Kretzoiarctos is still unclear.
Similar to the situation of Kretzoiarctos, the status of Agriarctos within Ailuropodini is also controversial. Questionable is also the species content of this genus (see the analysis in point 3 of the Discussion). Agriarctos was established by Kretzoi (Citation1942) based on p3-m2 from Hatvan, Hungary, whose age is usually attributed to MN12, but which is controversial and may be a little younger (see below). The type species Agriarctos gaali was erected from Hatvan, whereas a m1 from Rózsaszentmárton, Hungary (MN12) was described by Kretzoi (Citation1942) as A. vighi. Kretzoi also assigned Ursavus depereti established by Schlosser (Citation1902) on the basis of m1 and m2 from Melchingen, Germany (MN10), and Ursavus ehrenbergi from Halmyropotamos, Greece (MN12) to Agriarctos. Kretzoi (Citation1942) thought Agriarctos was close to Agriotherium, and he mentioned the similarity of premolar morphology with Ailuropoda, but supposed that otherwise the taxa are different. Thenius (Citation1947) later reassigned Halmyropotamos materials back to Ursavus, which was accepted by later authors (i.e., Qiu et al., Citation2014). Apart from the material from the type locality, some other remains from Europe were also assigned to U. depereti: Depéret and Llueca (Citation1928) assigned an incomplete mandible with m1 and m2 from Luzinay, France to this species; Viret and Mazenot (Citation1948) and later Viret (Citation1949) also assigned some isolated teeth (including P4, M1, M2) and a mandible fragment with m2 from Soblay, France (MN10) to Ursavus depereti, and these materials were redescribed by Ménouret and Mein (Citation2008) under Agriarctos depereti; Weitzel and Tobien (Citation1952) described one M1 and two m1 from Gau-Weinheim, Germany as Ursavus depereti and later assigned to this species another m2 (Tobien, Citation1955); Solounias (Citation1981) mentioned Ursavus cf. depereti in the fauna list of Samos, but he did not give any description. Roth and Morlo (Citation1997) assigned some isolated teeth to Ursavus depereti when they described the bear materials from Dorn-Dürkheim 1 (Rheinhessen, Germany); Koufos (Citation2006) described a mandible from Perivolaki, Thessaly, Greece (early MN12) as Ursavus depereti. As reflected above, all these authors probably neglected Agriarctos and still regard depereti as a species of Ursavus. Thenius (Citation1979) regarded Agriarctos (represented by Ag. gaali and Ag. depereti) as a direct ancestor of Ailuropoda in his review of the evolution of Ailuropodini. This view was supported by Hendey (Citation1980a, Citationb), though he neglected Ag. gaali. Qiu and Qi (Citation1989) made an adjustment to this view when they described Ailurarctos. They thought Ag. gaali is indeed closely related to Ailuropoda, but it is not its direct ancestor. However, some authors still hold a different opinion. Bernor et al. (Citation2003) redetermined Ag. gaali as Indarctos cf. vireti. Viranta (Citation2004) included Hatvan material in I. arctoides. Hunt (Citation2004) did not include Agriarctos in Ailuropodini, whereas Baryshnikov (Citation2007) assigned Ag. gaali to Indarctos arctoides as a subspecies of the latter, and reassigned Ag. depereti back to Ursavus. Therefore, the taxonomy of Agriarctos is still controversial (see point 3 in the Discussion).
Compared with the ambiguous Kretzoiarctos and Agriarctos, the evolutionary status of Ailurarctos as the ancestor of Ailuropoda is widely accepted (Hunt, Citation2004; Baryshnikov, Citation2007; Abella et al., Citation2012). This bear was first described by Qi (Citation1984). She distinguished isolated teeth of bears from Lufeng into two taxa, one as Ursavus depereti with a distinct wrinkle in the occlusal surface and another as Ursidae gen. indet. with a smooth occlusal surface. Later on, Qi (Citation1985) added three other teeth to Ursavus depereti. Qiu and Qi (Citation1989) restudied these materials and thought the wrinkled teeth were different from Ursavus depereti and assigned the materials to a new taxon, Ailurarctos lufengensis. The premolars of Ai. lufengensis are already specialized in developing large accessory cusps. The inner lobe of P4 develops several cusps like Ailuropoda, whereas the molars are relatively conservative and not much different from Ursavus. The material of Ailurarctos from Yuanmou were initially briefly described as Ailurarctos sp. nov. by Zong and Jiang (Citation1991). Later this material was formally described as Ai. yuanmouensis (Zong, Citation1997). The material includes a maxilla fragment with P4-M2, one M2, one p2, one m1, and four m2s. Zong (Citation1997) thought Ai. yuanmouensis was slightly smaller than Ai. lufengensis and differed from the latter species in shorter talons of M2 and the m2 with the width across the trigonid similar to the width across the talonid. Zong (Citation1997) noticed these characters and the simpler occlusal surface of the teeth of Ai. yuanmouensis, but he did not comment on the evolutionary status of the two species, probably influenced by the fact that the age of the Yuanmou fauna was thought to be Pliocene and younger than the Lufeng fauna at that time (Qian, Citation1997). Later, more comprehensive study suggested that the Yuanmou fauna was around 8–7 Ma, and Lufeng was 6.9–6.2 Ma (Dong and Qi, Citation2013).
Both species of Ailurarctos show clear affinity to Ailuropoda. Compared with European members of the Ailuropodini, Ailurarctos has an enlarged P4 parastyle, approaching that of the metacone, which is not seen in any European species. The inner lobe of the P4 of Ailurarctos is also very complicated. The M2 has a connected metacone and protocone posterior ridge, which is retained in living Ailuropoda, but this connection has been lost in cf. Ag. depereti. Ailurarctos is most probably the direct ancestor of Ailuropoda as Qiu and Qi (Citation1989) suggested.
On the basis of the notes above we propose a subtribe, Ailuropodina, which includes Ailurarctos and Ailuropoda. Notably, Ai. yuanmouensis is clearly more primitive than Ai. lufengensis because of: the less expanded P4 inner lobe, the lack of M2 entmetacone, the narrower m1 without RMed4 (see ). At the same time both species are in a single line towards the recent Ailuropoda. That is to say, that the type species Ai. lufengensis is closer to Ailuropoda than to Ai. yuanmouensis, and Ailurarctos is a paraphyletic group. See and for dentition of Ailuropodini.
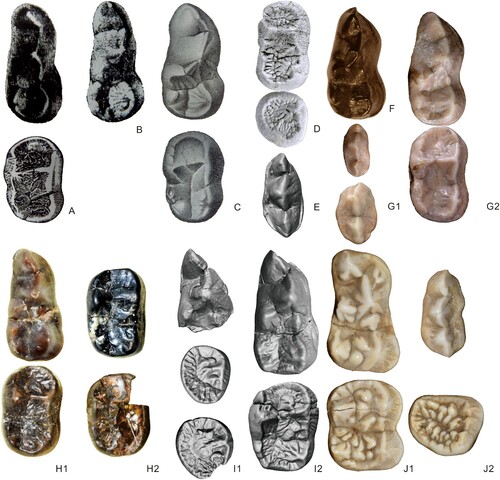
FIGURE 5. Upper teeth of Agriarctos and Ailurarctos. A1, cf. Ag. depereti from Soblay, from top to bottom P4, M1, and M2, from Viret (Citation1949); A2, cf. Ag. depereti from Soblay, from top to bottom P4, M1, and M2, cast from University of Vienna, MC AA 52bis; B, cf. Ag. depereti from Dorn-Dürkheim 1, from top to bottom P4 uncatalogued cast of MHNL AA52bis (reversed), M1 (reversed), and M2, uncatalogued casts of MHNL AA50-51 (reversed), all in the collection of the University of Vienna, modified from Roth and Morlo (Citation1997); C, Ai. yuanmouensis, from top to bottom P4, M1, and M2 (YICRA YV2509.1, reversed); D1, Ai. lufengensis, from top to bottom P4 (IVPP V6892.12), M1 (IVPP V6892.2, reversed), and M2 (IVPP V6892.6, reversed); D2, Ai. lufengensis, from top to bottom P4 (IVPP V6892.1, reversed), M1 (IVPP V6892.3), and M2 (IVPP V6892.6); D3, Ai. lufengensis, from top to bottom P4 (IVPP V6892.8), P2 (left, IVPP V6892.7) and P3 (right, IVPP V6892.7), and M2 (IVPP V6892.5, reversed); E, A. melanoleuca P4, M1 and M2 (IOZ32755, reversed). Not to scale.
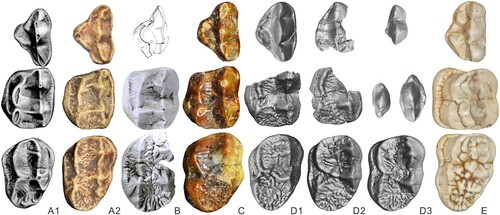
FIGURE 6. Lower teeth of Agriarctos and Ailurarctos. A, cf. Agriarctos sp. from Gau-Weinheim, m1 and m2 (reversed), from Weitzel and Tobien (Citation1952) and Tobien (Citation1955); B, cf. Ag. depereti from Gau-Weinheim, m1, from Weitzel and Tobien (Citation1952); C, Agriarctos depereti from Melchingen, from top to bottom m1 and m2 (both reversed), from Schlosser (Citation1902), Depéret and Llueca (Citation1928); D, cf. Ag. ?depereti from Dorn-Dürkheim 1, from top to bottom m1 and m2 (both reversed), from Roth and Morlo (Citation1997); E, Ai. lufengensis p4, V6892.13 (reversed); F, Ag. vighi, HSGI Ob/5691 from Rózsaszentmárton, m1 (reversed), photos courtesy of J. Wagner; G1, Ag. gaali V.60.1751 from Hatvan, from top to bottom p3 and p4, photos courtesy of J. Wagner; G2, Ag. gaali V.60.1751 from Hatvan, from top to bottom m1 and m2, photos courtesy of J. Wagner; H1, Ai. yuanmouensis, from top to bottom m1 (YICRA YV2509.5, reversed) and m2 (YICRA YV2509.4, reversed); H2, Ai. yuanmouensis, from top to bottom m2 (YICRA YV2509.2) and m2 (YICRA YV2509.3, reversed); I1, Ai. lufengensis, from top to bottom m1 (IVPP V6892.9), m3 (IVPP V6892.11, reversed), and m3 (IVPP V6892.10); I2, Ai. lufengensis, from top to bottom m1 (IVPP V25032), m2 (IVPP V6892.10); J1, A. melanoleuca from top to bottom m1 and m2 (IOZ32755, reversed); J2, A. melanoleuca from top to bottom p4 and m3 (IOZ32755, reversed). Not to scale.
Polyphyly of “Agriarctos” depereti
The poor preservation and scanty remains (one m1 and one m2) from the type locality Melchingen, Germany, led to a vague and insufficient diagnosis for the species, and the fossils later assigned to “Agriarctos” depereti must be considered as definitely polyphyletic. So far, the best representatives of Ag. depereti are actually from Soblay of France. This is because these materials include the only upper dentition of Ag. depereti, especially P4, which is diagnostic for Ailuropodini. Only one lower m2 is preserved in Soblay. It seems identical to the m2 from Melchingen: the tooth is relatively wide with middle constriction, and the cusp arrangement is also similar (Qiu and Qi, Citation1989; Ménouret and Mein, Citation2008). This makes it possible to see the fossil remains from Soblay conspecific with the ones from Melchingen. The Late Miocene Ursavus (e.g., U. tedfordi and Ursavus sp. V6894.B from Lufeng) often has m2 with no middle constriction. Therefore, the bears from Melchingen and Soblay probably belong to the same species, forming the major representatives of cf. Ag. depereti. The material from Luzinay is represented by an incomplete mandible with m1 and m2 (Depéret and Llueca, Citation1928). The size of the teeth is larger than those from Melchingen and Soblay, but the morphology is not different significantly. Materials from Gau-Weinheim are represented by one M1, two m1s, and one m2 (Weitzel and Tobien, Citation1952; Tobien, Citation1955). These teeth are smaller than corresponding teeth from Melchingen, Soblay, and Luzinay. The lingual border of M1 forms a distinct angle, like the case of the M1 from Soblay (especially MC AA96). m2 is also very similar to Melchingen and Soblay materials in having middle constriction. However, both m1s from Gau-Weinheim (= Wisseberg) are different from the m1 from Melchingen. The tooth is distinctly more slender than Melchingen m1, especially in the trigonid part. The fossils from Gau-Weinheim are from two layers, with the majority from MN9/10 but also including middle Miocene components (Pickford and Pourabrishami, Citation2013). Therefore, the m1s and m2 attributed to Ag. depereti from Gau-Weinheim are not necessarily from the same species; in any case the scarcity of materials does not allow deeper research for now.
Materials from Dorn-Dürkheim 1 (the age is unknown because of mixture of the material, but probably older than MN12: Spassov et al., Citation2012) represent the most abundant materials assigned to cf. Ag. depereti. P4 (fragment), M1, M2, m2, m3 are present in these materials. One m2 (SM DD3491) assigned to Ursavus sp. highly likely belongs to the same species. The inner lobe of P4 is preserved. It is very similar to one P4 (MC AA96) from Soblay: The protocone is very small whereas the hypocone is much larger. On the other hand, other teeth from Dorn-Dürkheim 1 are different from those in Melchingen and Soblay. The lingual corner of M1 (SM DD 3477) is present but weaker than in Dorn-Dürkheim 1 material, and the protocone is clearly subdivided into two parts. The protocone of M2 (SM DD 3482) is also subdivided, and the distal tip of the protocone is no longer connected with Rme3 but instead with a metaconule. The tooth occlusal surface is more complicated than in M2 from Soblay and Rpa1.2 is also present. The m2 (SM DD 3491) and m3 also have complicated occlusal surfaces. Rend3 is present in one m2 (SM DD3491). The other four m2s from this locality have a simpler occlusive surface. This is probably due to its higher abrasion degree. These characters suggest the materials from Dorn-Dürkheim 1 are different from typical Ag. depereti from Melchingen and Soblay which we will note here as cf. Ag. depereti (see point 3 in the Discussion). Several characters that distinguish Ag. depereti from Ailuropodina are further developed in bears from Dorn-Dürkheim 1: Posterior tips of protocone in M1 and M2 not medially shifted and mesocones are separated. This indicates such morphology is not plesiomorphic states in Ailuropodini, but autapomorphy for Agriarctos. The age of Dorn-Dürkheim 1 (see Spassov et al., Citation2012) is also probably younger than those of Melchingen and Soblay. The bear from Dorn-Dürkheim 1 should be considered a separate species of Ailuropodini, distinct from cf. Ag. depereti from Melchingen and Soblay.
Ursavus cf. depereti from Samos mentioned by Solounias (Citation1981) is unavailable to us and not discussed here. Ursavus depereti from Perivolaki, Thessaly, Greece is represented by a pair of the mandible with an age of MN12 (Koufos, Citation2006; Koufos, Citation2011). Both premolars and molars of this bear are much slenderer than those assigned to cf. Ag. depereti. The p1 and p2 are single-rooted, whereas the p3 and p4 have two roots. The premolars are all small without distinct accessory cusps, which is sharply different from the general morphology of Ailuropodini (though the premolars of cf. Ag. depereti are unknown). The molars are also different from cf. Ag. depereti in small and posterior located metaconid of m1 and absence of middle constriction in m2. In fact, this specimen is rather close to U. tedfordi from Northern China (Qiu et al., Citation2014). Both specimens show the reduction of premolars and m1 metaconid. RPrd3 is lingual to mesoconid, which is characteristic of Ursinae. But the Perivolaki material is also different from U. tedfordi in a larger size, shallower mandible, longer rostrum with distinct diastemata between premolars, more elongated teeth, less reduced p3 with two separated roots (fused in U. tedfordi), lower p4 main cusp, and m1 trigonid. These differences suggest Perivolaki material is a different species from U. tedfordi with a less carnivorous diet. In fact, the size, age, and geography of Perivolaki material fit U. ehrenbergi from Halmyropotamos, Greece, but U. ehrenbergi is so far only represented by the holotype, which is a maxilla fragment and not comparable to Perivolaki material. Therefore Perivolaki material is viewed as Ursavus sp. here.
In summary, the materials previously assigned to Agriarctos depereti actually belong to different lineages of bears. Only materials from Melchingen, Soblay, and Luzinay can be viewed as cf. Ag. depereti (see below our discussion on the problematic consideration of this species to Agriarctos, which makes the use of this genus name somewhat conditional). The status of materials from Gau-Weinheim and Samos is not solved. The sample from Dorn-Dürkheim 1 represent a more derived species of Ailuropodini, than the typical cf. Ag. depereti, whereas material from Perivolaki belongs to Ursavus with a similar evolution grade as U. ehrenbergi and probably belongs to this species.
Status of the Bulgarian Taxon and its Taxonomic and Nomenclatorial Problems
The P4 of ?Agriarctos nikolovi seems to be closer to European forms than to the genus Ailurarctos. The expanded inner lobe with several cusps and the large size demonstrates a gradual evolution from the much older (late Vallesian) cf. Ag. depereti (see below) from Soblay to the late Pontian Bulgarian taxon, which demonstrates several apomorphies. The resemblance in some features (strongly plicated cusps of the inner lobe, a very slight angle between paracone and metacone, the straight labial wall) to Ailurarctos and to Ailuropoda in size is probably not evidence of close relationship but parallelism.
It is necessary to note in this respect that the scarce remains, often incomparable to each other, make the nomenclature and taxonomic issues related to the European Ailuropodini rather confusing and contradictory. That is why we dwell on them in more detail here, taking into account that the evolved European forms are only tentatively grouped in Agriarctos in the recent literature (Abella et al., Citation2012; de Bonis et al., Citation2017). Thenius (Citation1979) analyzed the lower teeth of Agriarctos gaali Kretzoi Citation1942 and concluded that the upper P4 of this taxon must have a centrally located protocone (inner lobe) and therefore classifies the Soblay upper teeth, which have this characteristic morphology, to the genus Agriarctos as Ag. depereti. This conclusion related to the position of the inner lobe of the unknown P4 of Ag. gaali seems logical, but it still does not mean that the taxon from Soblay (late Vallesian: see Abella et al., Citation2012 and references herein) and that from Hatvan belong to the same genus. The age of Hatvan is debatable. Mein (Citation1990) correlated this locality to the latest Turolian (MN13). Fejfar et al. (Citation1997) and then Bernor et al. (Citation2003) included the locality in the MN12, seeing a parallel between the hipparions from this locality and the ones from Baltavar (Hungary), Vienna basin (Austria), and Baccinello V3 (Italy), believing that these hipparions have a late MN12 age. But Baccinello V3 has a Middle Messinian age (Rook et al., Citation2011), so the age of Hatvan is uncertain, possibly very close to the MN12/13 boundary, or early MN13 in age. Even if the Hatvan ursid has an Ailuropodina P4 morphology (which is only supposed), the small taxon from Soblay and the large one from Hatvan could have several significant differences, moreover, they are distant from each other in time by at least two million years. After some opinions (Bernor et al., Citation2003; Baryshnikov, Citation2007) the Hatvan ursid represents an Indarctos species, although this opinion is disputed in later work (Abella et al., Citation2012). Having in mind all these obscurities in the relation of the species from Hatvan to the line of the giant panda and of the taxon from Soblay to Agriarctos, here we prefer to note the Ailuropodini teeth of Soblay as cf. Agriarctos depereti. See for the distribution map of Ailuropodini.
A direct comparison of the Bulgarian P4 with the same tooth of the Hatvan taxon is also not possible and the Bulgarian taxon is most probably at least ca. 1 Ma younger (see: Boehme et al. [Citation2017] and Spassov et al. [Citation2017] for the age of the middle Turolian, Tortonian/Messinian boundary). Having in mind all these taxonomic uncertainties and considerations we still think that the most parsimonious taxonomic decision is to describe here the Bulgarian taxon as a new species under the name ?Agriarctos nikolovi sp. nov. (and not as Ailurarctos nikolovi) noting its large size, relative age, geographic location, and morphological features (see reconstruction of ?Agriarctos nikolovi in .).
FIGURE 7. Proposed dispersal route of Ailuropodini. The red area indicates the possible distribution of the ancestor of Ailuropodini. Background from Geomapapp.
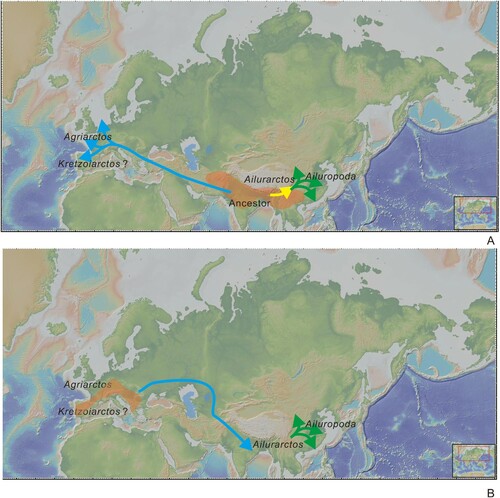
FIGURE 8. Reconstruction of ?A. nikolovi sp. nov. from Bulgaria. Artwork by Velizar Simeonovski, Chicago. The reconstruction was developed by a regressive estimate of the coloration of the modern A. melanoleuca, based on the variation of the coloration of Ursus arctos. Ursus arctos is used for calibration because the variation of its coloration (geographic, ecological, and individual) provides the most comprehensive model for the evolution of the coloration in Ursidae genera.

Ecology of ?Agriarctos nikolovi and related forms
Though with a simpler occlusive surface, the Turolian Agriarctos is similar to Ailurarctos or even Ailuropoda in many aspects. ?Agriarctos nikolovi, even though less derived than Ailurarctos (if the characters of Ailuropoda are viewed as derived characters) in having a less elongated P4 parastyle, it has a quite expanded inner lobe, similar to that of Ai. lufengensis, thus more derived than the situation of both cf. Ag. depereti and Ai. yuanmouensis in this aspect, reflecting the increasing masticatory ability evolved in both genera. ?Agriarctos nikolovi is also more derived than Ailurarctos in the absence of cingulum, though the function of this structure is unclear. Having in mind the taphonomic features related to the fossils of ?Ag. nikolovi, and in particular that they were found in coal-bearing deposits, we can assume that this ailuropodine inhabited humid forests associated with river floods and swampy areas. This type of landscape is not at all dominant, but it must have been present in the environment of the forested savannah of the middle and late Turolian “Pikermian paleobiome” of the Balkans (see Fortelius et al., Citation2019) and may have had some territorial expansion in local episodes (see Böhme et al., Citation2017) of increased humidity in the second half of the Messinian. It is possible that the Messinian salinity crisis (5.96–5.33 Ma) contributed mainly to the disappearance of this latest European Ailuropodini. The very robust and disproportionately large canines of ?Ag. nikolovi, comparable in size with the ones of A. melanoleuca, give some additional data on the ecology of the species. Most likely, the canines indicate a rather large body size of the animal. Judging by the features of the upper carnassial (see Discussion above), we can hardly assume that the canines were related to particularly active predation. It seems more likely that they gave advantage in competition and defence with the numerous predators known from the hipparion fauna of the Pikermian and post-Pikermian Turolian biomes of the Balkans, or related with sexual dimorphism.
The surfaces of the lower teeth are weakly wrinkled in cf. Ag. depereti and Ag. gaali, whereas the m2 from Dorn-Dürkheim 1 is rather complicated, like that of Ailurarctos. This indicates the bear from Dorn-Dürkheim 1 is probably not a direct ancestor of Ag. gaali, and complicated m2 shared with Ailurarctos can be explained as parallelism. Ag. gaali is similar to Ailurarctos or Ailuropoda in many ways, and this species is even more derived than Ai. yuanmouensis in: larger size (comparable to A. microta); wider m1 with a convex buccal side of the trigonid; elongated m2 with a distinct middle constriction. Ag. gaali is also more derived than Ai. lufengensis in a larger size; stronger accessory cusps of premolars with distinct inner ridge; convex buccal side of trigonid of m1 and elongated m2. These characters shared by Ag. gaali and Ailuropoda are interpreted as parallelism, since it lacks synapomorphy defining Ailuropodine mentioned above. Such paralleled characters in European and Asian lineage are related to increasing masseteric ability (enhancing the premolar size/strength and enlarging the molar occlusive area), indicating both Ailuropodina and Ag. gaali evolved towards an herbivorous diet. The simpler occlusive surface of Ag. gaali suggests it fed on softer food than Ailurarctos did. Both the European and the Chinese Miocene lineages were unlikely to mainly feed on bamboo, such as the living Ailuropoda, since the cusps and ridges of the former two genera are not strong enough to sustain the pressure from hard bamboo, and bamboo fossils are rare in the European Late Miocene (D. Ivanov, Bulgarian Acad. Sci., pers. comm.).
An important find was described very recently: the so-called “false thumb”, known in the recent giant panda, was also discovered in Ailurarctos lufengensis (Wang et al., Citation2022). This morphofunctional peculiarity is interpreted by the authors, judging after recent Ailuropoda, as an adaptation above all for bamboo food consumption, but also in connection with the plantigrade posture. As noted above, the teeth of Ailurarctos and ?Agriarctos nikolovi were probably not adapted yet to hard food like bamboo. We could suppose that in the early (Ailurarctos) stage of evolution of the radial sesamoid in giant pandas as extra “thumb”, it represented an adaptation mainly for gripping and holding thin twigs with other, softer than bamboo, plant food. In this case, having in mind the derived evolutionary stage of the contemporary to Ailurarctos ?Agriarctos nikolovi, we cannot rule out the possibility that a similar adaptation was also developed in this ailuropodin.
Dispersal of Early Ailuropodini
The European late Miocene giant panda, which roughly can be assigned to Agriarctos, represent a lineage evolved independently from Ailuropodina. The early member cf. Ag. depereti already shows several derived characters (subdivided M2 protocone, loss contact of M2 metacone and protocone), which indicates the different trends from Ailuropodina. Therefore, Agriarctos might indicate an early dispersal of Ailuropodini into Europe around the middle/late Miocene boundary. The position of Kretzoiarctos is not solved due to poor preservation. Whether it represents an early member of Ailuropodini as proposed by Abella et al. (Citation2012) or an early member of Agriotheriini as proposed by Qiu et al. (Citation2014) needs more material to answer. When Ai. yuanmouensis appeared, it was already more derived than contemporary ?Agriarctos from Dorn-Dürkheim 1, suggesting a deep divergence between the two lineages.
The common ancestor of Agriarctos and Ailuropodina is unknown, probably with roots in Kretzoiarctos or a related taxon. Note that Kretzoiarctos already has a relatively larger body size, suggesting a potential early divergence of this genus as the only ancestor of the European Agriarctos. We suppose South Asia–Southeastern Asia–Southern China, which has a rather poor fossil record during the middle and early late Miocene, is more likely to be the origination area for the tribe. In Northern China and Europe, the fossil records are much more complete, but no such representatives were present. On the other hand, the so far earliest representative of the tribe is still found in Europe, and having in mind the data about the wide distribution of the humid forests in Western Europe in the middle Miocene and the beginning of the late Miocene, the hypothesis of European origin cannot be excluded and new material of this group will be needed for further analysis. We present both hypotheses of dispersal in . The endemic evolution of Ai. yuanmouensis and Ai. lufengensis suggests Yunnan could be within the center of Ailuropodina evolution.
CONCLUSION
The find of isolated teeth from Bulgaria is identified as a new, rather young in age and very evolved species of giant panda lineage, ?Agriarctos nikolovi sp. nov. Our analysis specifies the frames of the tribe Ailuropodini, and shows that the taxon mentioned generally in the literature as Agriarctos depereti is a polyphyletic group which includes various taxa from which not all represent members of Ailuropodini. Only the fossils from Melchingen, Soblay, and Luzinay could be attributed to this species noted here as cf. Agriarctos depereti, because its generic appurtenance to Agriarctos is impossible to prove without new fossil discoveries. ?Agriarctos nikolovi sp. nov. demonstrates more affinities with Agriarctos than with Ailurarctos, but at the same time shows some more advanced traits than the more primitive Ailurarctos, Ai. yuanmouensis, suggesting a parallelism with Ailurarctos in several features, specifically related to an increase in specific herbivory. European late Miocene species seem to form an independent lineage of the giant panda lineage, in which ?Ag. nikolovi is the most evolved member of this lineage, whereas the Asian genus Ailurarctos must be the direct ancestor to Ailuropoda. The subtribe Ailuropodina is proposed for these two Chinese genera.
Supplemental Material
Download MS Word (375.2 KB)ACKNOWLEDGMENTS
The authors would like to express their deep gratitude to Z. Qiu, who supported the authors to study the material of Ailurarctos lufengensis and provided useful literature. The authors would also like to show thanks to G. Rabeder, as well as to H. Thomas (Paris) who allowed the authors to study the cast of Agriarctos depereti from Soblay. Besides, the authors thank Y. Hou for help in CT scanning, J. Chen for help in the observation of fossil specimens in IVPP, and X. Zhu and Y. Yang for help in the observation of living Ailuropoda in IOZ and A. Ignatov (NMNHS) who prepared the photos of the new Bulgarian taxon. This work was supported by the Strategic Priority Research Program of Chinese Academy of Sciences (Grant No. XDB26000000 and XDA20070203), Chinese Natural Science Foundation Program (grant number 42102001 and 41772018), Key Frontier Science Research Program of the Chinese Academy of Sciences (Grant Nos. QYZDY-SSW-DQC-22 and GJHZ1885), Second Tibetan Plateau Scientific Expedition and Research (Grant 2019QZKK0705).
LITERATURE CITED
- Abella, J., P. Montoya, and J. Morales. 2011. A new species of Agriarctos (Ailuropodinae, Ursidae, Carnivora) in the locality of Nombrevilla 2 (Zaragoza, Spain). Estudios Geológicos 67:187–191.
- Abella, J., D. M. Alba, J. M. Robles, A. Valenciano, C. Rotgers, R. Carmona, P. Montoya, and J. Morales. 2012. Kretzoiarctos gen. nov., the oldest member of the giant panda clade. PLoS One 7, e48985.
- Baryshnikov, G. 2007. Fauna of Russia and neighbouring countries. Mammals. Ursidae. Nauka, Saint Petersburg, 542 pp.
- Batsch, A. J. G. C. 1788. Versuch einer Anleitung, zur Kenntniβ und Geschichte der Thiere und Mineralien, für akademische Vorlesungen entworfen und mit den nöthigsten Abbildungen versehen Erster Theil. Academischen Buchhandlung, Jena.
- Bernor, R. L., C. Feibel, and S. Viranta. 2003. The Vertebrate Locality of Hatvan, Late Miocene (Middle Turolian, MN12), Hungary. Advances in Vertebrate Paleontology Hen to Panta. Bucharest: Romanian Academy Emil Racovita Institute of Speleology:105–112.
- Böhme, M., N. Spassov, M. Ebner, D. Geraads, L. Hristova, U. Kirscher, S. Kötter, U. Linnemann, J. Prieto, S. Roussiakis, G. Theodorou, G. Uhlig, and M. Winklhofer. 2017. Messinian age and savannah environment of the possible hominin Graecopithecus from Europe. PLOS ONE 12, e0177347.
- Bonis, L. de, J. Abella, G. Merceron, and D. R. Begun. 2017. A new late Miocene ailuropodine (giant panda) from Rudabánya (North-central Hungary). Geobios 50:413–421.
- Bowdich, T. W. 1821. An Analysis of the Natural Classification of Mammalia, for the Use of Students and Travelers. J. Smith, Paris, 115 pp.
- Crusafont Pairó, M. and B. Kurtén, 1976. Bears and bear-dogs from the Vallesian of the Vallés-Penedés basin, Spain. Acta Zoologica Fennica 144:1–30.
- Depéret, C., and F. G. Llueca. 1928. Sur l’ Indarctos arctoides et la phylogénie des Ursidés. Bulletin de la Société Géologique de France 28, 149–160.
- Dong, W., and G.-Q. Qi. 2013. Hominoid-producing localities and biostratigraphy in Yunnan; pp. 293–313 in X. M. Wang, L. J. Flynn, and M. Fortelius (eds.), Fossil Mammals of Asia. Columbia University Press, New York.
- Fejfar, O., W. D. Heinrich, M. A. Pevzner, and E. A. Vangengeim. 1997. Late Cenozoic sequences of mammalian sites in Eurasia: an updated correlation. Palaeogeography Palaeoclimatology Palaeoecology 133:259–288.
- Fortelius, M., F. Bibi, H. Tang, I. Žliobaitė, J. T. Eronen, and F. Kaya. 2019. The nature of the Old World savannah palaeobiome. Nature Ecology & Evolution 3:504–504.
- Grevé, C. 1892. Zoologischen Jahrbücher, Abtheilung für Systematik, Geographie und Biologie der Thiere, 6:589–616
- Hendey, Q. B. 1980a. Agriotherium (Mammalia, Ursidae) from Langebaanweg, South Africa, and relationships of the genus. Annals of the South African Museum 81, 1–109.
- Hendey, Q. B. 1980b. Origin of the giant panda. South African Journal of Science 76:179–180.
- Hunt, R. M. Jr. 2004. A paleontologist’s perspective on the origin and relationships of the giant panda’ pp. 45–52 in D. Lindburg and K. Baragona (eds.), Giant pandas: biology and conservation. University of California Press.
- Jiangzuo, Q., J. Liu, and J. Chen. 2019. Morphological homology, evolution, and proposed nomenclature for bear dentition. Acta Palaeontologica Polonica 64:693–710.
- Kamenov, B., and E. Kojumdgieva. 1983. Stratigraphy of the Neogene of the Sofia basin. Palaeontology, Stratigraphy & Lithology 18:9–85.
- Koufos, G. D. 2006. The late Miocene vertebrate locality of Perivolaki, Thessaly, Greece. 4. Carnivora. Palaeontographica, Abt. A 276:9–74.
- Koufos, G. D. 2011. The Miocene carnivore assemblage of Greece. Estudios Geológicos 67:91–320.
- Kretzoi, M. 1942. Zwei neue Agriotheriiden asu dem ungarischen Pannon. Földtani Közlöny 72:50–353.
- Mein, P. 1990. Updating of MN zones; pp. 73–90 in: E. Lindsay, V. Falbusch, and P. Mein, (eds.), European Neogene mammal chronology. Plenum Press, New York.
- Ménouret, B., and P. Mein. 2008. Les vertébrés du Miocène supérieur de Soblay (Ain, France). Documents des Laboratoires de géologie Lyon, 1665:1–97.
- Pei, W. 1987. Canivora, Proboscidea and Rodentia from Liucheng Gigantopithecus Cave and other caves in Guangxi. Memoirs of Institute of Vertebrate Palaeontology and Palaeoanthropology, Academia Sinica 18:1–119.
- Pickford, M., and Z. Pourabrishami. 2013. Deciphering Dinotheriensande deinotheriid diversity. Palaeobiodiversity and Palaeoenvironments 93:121–150.
- Popov, N. 1993. Novihan Member of the Lozenets Formation; p. 233 in: Y. Tenchov (ed.), Glossary of the Formal Lithostratigraphic units in Bulgaria (1882–1992). Publishing House of the Bulgarian Academy of Sciences, Sofia.
- Qi, G. Q. 1984. First discovery of Ursavus in China and note on other Ursidae specimens from the Ramapithecus fossil site of Lufeng. Acta Anthropologica Sinica 3:53–61.
- Qi, G. Q. 1985. A preliminary report on Carnivora from the Ramapithecus locality, Lufeng, Yunnan. Acta Anthropologica Sinica 4:33–43.
- Qian, F. 1997. 7 Determination of geological age; pp. 161–178 in Z. Q. He (ed.), Yuanmou Hominoid Fauna. Yunnan Science and Technology Press, Kunming.
- Qiu, Z. X., and G. Q. Qi. 1989. Ailuropod found from the Late Miocene deposits in Lufeng, Yunnan. Vertebrata PalAsiatica 27:153–169.
- Qiu, Z. X., T. Deng, and B. Y. Wang. 2014. A Late Miocene Ursavus skull from Guanghe, Gansu, China. Vertebrata PalAsiatica 52:265–302.
- Rook, L., O. Oms, M. G. Benvenuti, and M. Papini. 2011. Magnetostratigraphy of the Late Miocene Baccinello–Cinigiano basin (Tuscany, Italy) and the age of Oreopithecus bambolii faunal assemblages. Palaeogeography, Palaeoclimatology, Palaeoecology 305:286–294.
- Roth, C. H., and M. Morlo, 1997. Die Raubtiere (Mammalia, Carnivora) aus dem Turolium von Dorn-Dürkheim 1 (Rheinhessen). Teil 2: Ursidae. Courier-Forschungsinstitut Senckenberg 197:49–71.
- Schlosser, M. 1902. Beiträge zur Kenntniss der Säugethierreste aus den süddeutschen Bohnerzen. Geologische und Palaeontologische Abhandlungen N. F. 5:117–258.
- Solounias, N. 1981. The Turolian fauna from the Island of Samos, Greece. With special emphasis on the hyaenids and the bovids. Contributions to Vertebrate Evolution 6:1–232.
- Spassov, N., M. Böhme, D. Geraads, S. Kötter, and C. van Baak. 2017. Pikermian mammal event, post-Pikermian mammal turnover and appearance of Graecopithecus, Book of Abstracts of the 15 th Congress of the RCMNS, p. 29.
- Spassov, N., D. Geraads, L. Hristova, G. Markov, G. Merceron, T. Tzankov, K. Stoyanov, M. Böhme, and A. Dimitrova. 2012. A hominid tooth from Bulgaria: the last pre-human hominid of continental Europe. Journal of human evolution 62:138–145.
- Thenius, E. 1947. Ursavus ehrenbergi aus dem Pont von Euböa (Griechenland). Sitzungsberichte der Österreichischen Akademie der Wissenschaften, Matematisch-naturwissenschaftliche Klasse 156:225–249.
- Thenius, E. 1979. Zur systematischen und phylogenetischen Stellung des Bambusbären: Ailuropoda melanoleuca David (Carnivora, Mammalia). Zeitschrift für Säugetierkunde 44:286–305.
- Tobien, H. 1955. Neue und wenig bekannte Carnivoren aus den Unterpliozänen Dinotheriensanden Rheinhessens. Notizblatt des Hessischen Landesamtes für Bodenforschung zu Wiesbaden 83(6. Folge, H. 6):7–31.
- Viranta, S. 2004. Habitat preferences of European Middle Miocene omnivorous ursids. Acta Palaeontologica Polonica 49:325–327.
- Viret, J. 1949. Observations complémentaires sur quelques mammifères fossiles de Soblay. Eclogae Geologicae Helvetiae 42:469–476.
- Viret, J., and G. Mazenot. 1948. Nouveaux restes de mammifères dans le gisement de lignite pontien de Soblay (Ain). Annales de Paléontologie 34:19–56.
- Wang X., D. Su, N. Jablonski, X. Ji, J. Kelley, L. Flynn, and T. Deng. 2022. Earliest giant panda false thumb suggests conflicting demands for locomotion and feeding. Scientific Reports. 12:10538.
- Weitzel, K.A., and H. Tobien. 1952. Indarctos und Ursavus (Carnivora, Mamm.) aus den Unterpliozänen Dinotheriensanden Rheinhessens. Notizblatt des Hessischen Landesamtes für Bodenforschung zu Wiesbaden 80(6. Folge, H. 3):7–14.
- Zhou, C. Q., J. C. Hu, C. G. Yuan, and F. W. Wei. 1997. Giant pandas’ food habits and feeding behaviour in Mabian Dafengding Natural reserve. Journal of Sichuan Teachers College (Natural Science) 18:3–7.
- Zong, G. F., 1997. 5.3.4 Carnivora; pp. 69–88 in Z.Q. He (ed.), Yuanmou Hominoid Fauna. Yunnan Science and Technology Press, Kunming,
- Zong, G. F., and C. Jiang. 1991. A preliminary observation on Carnivora from the Neogene locality from Yuanmou, Yuannan. Vertebrata PalAsiatica 29:136–142.