
ABSTRACT
The Eocene taxon Masillaraptoridae includes long-legged, raptorial birds, the phylogenetic affinities of which are poorly resolved. Here, fossils from the London Clay of Walton-on-the-Naze (Essex, U.K.) are described, which corroborate the hypothesis that masillaraptorids are stem group representatives of the Falconiformes (falcons). Two partial skeletons are assigned to a new genus and species, Danielsraptor phorusrhacoides, gen. et sp. nov., whereas a smaller species closely resembles Masillaraptor parvunguis from the German fossil site Messel. The new fossils show previously unrecognized derived characters shared by masillaraptorids and falconiform birds, and a phylogenetic analysis also supported a sister group relationship between the Masillaraptoridae and the Falconiformes. By contrast, the long and deep beak of masillaraptorids is remarkably similar to that of the cariamiform Phorusrhacidae and the tarsometatarsus also shows a resemblance to that of some Cariamiformes. The similarities between masillaraptorids and the Cariamiformes are notable, because most sequence-based analyses show the Falconiformes and Cariamiformes to be closely related, and a few studies even suggest a sister group relationship between both taxa. These similarities may be plesiomorphic for a more inclusive clade containing falconiforms and cariamiforms, but it is also possible that masillaraptorids represent a direct evolutionary link between the Falconiformes and Cariamiformes.
INTRODUCTION
Falcons (Falconiformes) were long considered to be closely related to other diurnal birds of prey. However, most current molecular analyses recover them in a clade together with seriemas (Cariamiformes), parrots (Psittaciformes), and passerines (Passeriformes; Ericson et al., Citation2006; Hackett et al., Citation2008; Prum et al., Citation2015; Kuhl et al., Citation2021). These taxa exhibit very disparate morphologies and none is particularly close to falcons in anatomy, behavior, or ecology.
The fossil record of the Falconiformes is very poor. The earliest modern-type falconiform is from Miocene deposits (Li et al., Citation2014; Mayr, Citation2017), and the scanty Paleogene fossil record does not shed much light on the morphology of early stem group representatives. Even though a few fragmentary fossils from the early Eocene of Europe and Antarctica were assigned to the Falconiformes (Harrison, Citation1984; Cenizo et al., Citation2016; Mayr, Citation2022), these are only based on distal tarsometatarsi and therefore do not elucidate the evolutionary history of falconiform birds. However, there is one better represented Eocene taxon, the Masillaraptoridae, for which affinities to the Falconiformes were proposed.
Masillaraptorids are known from various European localities and the taxon was first established for Masillaraptor parvunguis Mayr, Citation2006 from the latest early or earliest middle Eocene of Messel in Germany (Mayr, Citation2006, Citation2009). From this fossil site, two badly preserved skeletons and a skull of M. parvunguis are known. A partial tarsometatarsus of Masillaraptor was also identified in the middle Eocene strata of the Geisel Valley in Germany (Mayr, Citation2002, Citation2020), and a distal tarsometatarsus and a partial beak of a masillaraptorid were found at the lower Eocene fossil site Egem in Belgium (Mayr and Smith, Citation2019). An as yet undescribed skull of a masillaraptorid also exists from the lower Eocene North American Green River Formation (Grande, Citation2013:fig. 143A, top left figure panel).
Arguably the best-preserved fossils of Masillaraptor-like birds come from the lower Eocene London Clay of Walton-on-the-Naze (Essex, U.K.). The specimens were found more than two decades ago by the late Michael Daniels, but until recently they remained in private hands (Daniels, Citation1992, Citation1995). Owing to the bequest of this highly prolific and successful collector, these fossils and all other items of the Daniels collection were recently donated to National Museums Scotland, now allowing their formal description.
The Daniels collection includes at least two species of the Masillaraptoridae, the close affinities of which were first recognized by Daniels (Citation1995). The main specimen of the larger species comprises substantial portions of the skull and sheds light on the phylogenetic affinities of masillaraptorids. A smaller species appears to be closely related to, or even conspecific with, Masillaraptor parvunguis from Messel.
Masillaraptor parvunguis was considered to be of uncertain affinities when first described (Mayr, Citation2006), but subsequently masillaraptorids were regarded as stem group representatives of the Falconiformes (Mayr, Citation2009, Citation2022). The published character evidence for this placement is still weak but the new fossils exhibit previously unknown characters, which further support close affinities between masillaraptorids and falconiforms. Daniels (Citation1995) proposed that the larger species from the London Clay, which is described in the present study, shows affinities to the cariamiform Phorusrhacidae. Even though this hypothesis is not confirmed herein, the similarities between masillaraptorids and phorusrhacids are of potential significance with regard to current hypotheses on the phylogenetic affinities of the Falconiformes.
MATERIAL AND METHODS
To assess the affinities of the fossils, a phylogenetic analysis was performed based on the revised and emended character matrix of Mayr (Citation2016) including nine ingroup taxa and 42 characters (see Supplemental Data for character descriptions and character matrix). The analysis was run with the heuristic search modus of NONA 2.0 (Goloboff, Citation1993) through the WINCLADA 1.00.08 interface (Nixon, Citation2002), using the commands hold 10000, mult*1000, hold/10, and max*. Bootstrap support values were calculated with 1000 replicates, 10 searches holding 10 trees per replicate, and TBR branch swapping without max*. Tree length (L), consistency index (CI), and retention index (RI) were calculated. The trees were rooted with the anseriform Anhimidae (Chauna torquata). Caracara plancus was included as a representative as crown group Falconiformes. Following Mayr (Citation2016), the stem group cariamiform taxon Dynamopterus (Idiornithidae) was scored after D. gallicus (Milne-Edwards, Citation1892), the best represented species, with additional character scorings being based on D. minor (Milne-Edwards, Citation1892) and D. cf. itardiensis (Mourer-Chauviré, Citation1983). Scoring of Bathornis is based on B. grallator (Wetmore, Citation1944) with scorings for tarsometatarsus characters added after B. celeripes Wetmore, Citation1933 (Mayr, Citation2016). The Phorusrhacidae are represented by the species Llallawavis scagliai Degrange et al., Citation2015, Psilopterus lemoinei (Moreno and Mercerat, Citation1891), and Patagornis marshi Moreno and Mercerat, Citation1891.
Institutional Abbreviations—AMNH, American Museum of Natural History, New York, U.S.A.; GMH, Geiseltalsammlung, Martin-Luther Universität of Halle-Wittenberg, Germany; IRSNB, Royal Belgian Institute of Natural Sciences, Brussels, Belgium; NHMUK, Natural History Museum, London, U.K.; NMS, National Museums Scotland, Edinburgh, U.K.; SMF, Senckenberg Research Institute, Frankfurt, Germany.
SYSTEMATIC PALEONTOLOGY
Emended Diagnosis—Upper beak long, mediolaterally compressed, and dorsoventrally deep, with straight culmen (dorsal ridge) and deeply hooked tip; tomia just caudal of tip of upper beak not forming sharp cutting edges but bearing “tomial grooves”; processus zygomaticus well developed; mandible with deep rami, long and narrow mandibular symphysis, and upcurved rostral portion; lacrimal bone with long and tapering processus supraorbitalis; one caudal vertebra with very long processus ventralis; pygostyle with large lamina pygostyli; coracoid with long processus procoracoideus and medial projection on extremitas sternalis; sternum with two pairs of caudal incisions; legs long; femur equaling tarsometatarsus in length; hypotarsus block-like; trochlea metatarsi IV bearing wing-like plantar flange; first phalanx of second toe and second and third phalanges of fourth toe shortened.
DANIELSRAPTOR, gen. nov.
Type Species—Danielsraptor phorusrhacoides, sp. nov.
Diagnosis—Characterized by ventral portion of cotyla of second phalanx of third toe forming a marked, lip-like projection; the new taxon is masculine in gender.
Differential Diagnosis—Danielsraptor, gen. nov. differs from the distinctly smaller Masillaraptor in having pygostyle with curved caudoventral margin (straighter in Masillaraptor); coracoid stouter, with proportionally longer omal extremity; scapula with proportionally longer acromion; humerus without projected processus flexorius; tarsometatarsus somewhat stouter, with trochlea metatarsi II reaching farther distally and trochlea metatarsi IV bearing a better developed wing-like flange. Danielsraptor differs from Stintonornis Harrison, Citation1984 in smaller dorsal and plantar openings of foramen vasculare distale and proportionally smaller trochlea metatarsi II that reaches less far distally; distinguished from Antarctoboenus Cenizo et al., Citation2016 in that foramen vascular distale proportionally larger and trochlea metatarsi IV having a larger plantar flange.
Etymology—The taxon is named after the late Michael Daniels, who collected and prepared the holotype and the tentatively referred specimen as well as numerous other bird fossils from Walton-on-the-Naze; raptor (Lat.: ravisher, thief) is the vernacular English term for a bird of prey and refers to the presumed feeding ecology of the new taxon.
DANIELSRAPTOR PHORUSRHACOIDES, sp. nov.
Holotype—NMS.Z.2021.40.12 (, S1; partial skeleton including the skull, vertebrae, fragmentary wing bones, elements of the pectoral girdle, pelvis, substantial portions of both legs, as well as various bone fragments that are still partially enclosed in matrix), collected in 1991 by M. Daniels (original collector’s number WN 91696).
FIGURE 1. Holotype of Danielsraptor phorusrhacoides, gen. et sp. nov. from the lower Eocene London Clay of Walton-on-the-Naze, Essex, U.K. (NMS.Z.2021.40.12), skull elements. A, B, upper beak in A, lateral and B, ventral view. C, dorsal nasal bar in ventral view. D, jugal bars. E, F, dorsal portion of neurocranium in dorsal (E) and ventral (F) view. G, H, mandible in dorsal (G) and lateral (H) view; the arrow denotes an enlarged view of the caudal end. I–L, left quadrate in caudal (I), medial (J), lateral (K), and ventral (L) view. M, skull of Masillaraptor parvunguis from the Messel fossil site in Germany (IRSNB Av 83). N, O, left os lacrimale in dorsal (N) and lateral (O) view. P, two unidentified ossicles, which may represent the caudal portions of the pterygoids. Abbreviations: cdl, condylus lateralis; cdm, condylus medialis; cdp, condylus pterygoideus; cpo, capitulum oticum; cps, capitulum squamosum; ctq, cotyla quadratojugalis; ctl, cotyla lateralis; dcp, descending process of lacrimal; fpn, foramen pneumaticum; pmd, processus medialis; ppo, processus postorbitalis; prj, dorsal projection; pso, processus supraorbitalis; tgr, tomial groove. Scale bars equal 10 mm; same scale for A–H.

FIGURE 2. Comparison of skull elements of the Masillaraptoridae, crown group Falconiformes, and the Cariamiformes. A, digitally assembled skull bones and mandible of A, Danielsraptor phorusrhacoides (holotype, NMS.Z.2021.40.12). B, skull of Caracara plancus (Falconiformes; SMF 3462). C, skull of Psilopterus lemoinei (Cariamiformes, Phorusrhacidae; AMNH 9257, the dotted line indicates the shape of the missing tip of the beak). D–L, left quadrate of D–F, D. phorusrhacoides (holotype), G–I, C. plancus (SMF 6441), and J–L, Cariama cristata (Cariamiformes; SMF 1862; right quadrate, mirrored) in caudal (D, G, J), medial (E, H, K), and lateral (F, I, L) view. M–O, caudal end of mandible (lateral view) of M, D. phorusrhacoides (holotype), N, Micrastur ruficollis (Falconiformes; SMF 9812), and O, C. cristata (SMF 1862). Abbreviations: cdl, condylus lateralis; cdm, condylus medialis; cdp, condylus pterygoideus; cpo, capitulum oticum; cps, capitulum squamosum; iic, incisura intercapitularis; orb, processus orbitalis; prj, dorsal projection; tgr, tomial groove. Scale bars equal 10 mm.

FIGURE 3. Holotype of Danielsraptor phorusrhacoides, gen. et sp. nov. from the lower Eocene London Clay of Walton-on-the-Naze, Essex, U.K. (NMS.Z.2021.40.12), vertebrae and elements of the pectoral girdle and wing. A–D, four thoracic vertebrae in dorsal and lateral view. E, a caudal vertebra with a long processus ventralis in cranial, ventral, and lateral view. F, five further caudal vertebrae in different views. G, pygostyle in lateral view. H, I, left coracoid in dorsal (H) and ventral (I) view. J–L, right coracoid in ventral (J), dorsal (K), and medial (L) view. M, N, right scapula in medial (M) and lateral (N) view. O, extremitas sternalis of furcula. P, left extremitas omalis of furcula. Q, R, cranialmost portion of sternum in lateral (Q) and ventral (R) view. S, proximal portion of left humerus in cranial view. T, U, distal end of left (T) and right (U) humerus in cranial view. V, distal end of left ulna in ventral view. Abbreviations: cdd, condylus dorsalis; cdv, condylus ventralis; csc, cotyla scapularis; fac, facies articularis clavicularis; fns, foramen nervi supracoracoidei; fos, fossa in sulcus supracoracoideus; pcv, processus ventralis; sac, sulcus articularis coracoideus; spe, spina externa. Scale bar equals 10 mm.

FIGURE 4. Danielsraptor phorusrhacoides, gen. et sp. nov., bones of the tentatively referred specimen NMS.Z.2021.40.13 in comparison to the holotype and Masillaraptor cf. parvunguis. A, three thoracic vertebrae of NMS.Z.2021.40.13 in lateral and—regarding a fragmentary specimen—dorsal view. B, seven caudal vertebrae of NMS.Z.2021.40.13 in different views. C, furcula of NMS.Z.2021.40.13. D–F, pygostyle of D, the holotype of D. phorusrhacoides (NMS.Z.2021.40.12), E, the tentatively referred specimen NMS.Z.2021.40.13, and F, M. cf. parvunguis (NMS.Z.2021.40.14); the dotted lines indicate the reconstructed hypothetical outlines of the bones. G–I, coracoid (dorsal view) of G, the holotype of D. phorusrhacoides, H, the tentatively referred specimen NMS.Z.2021.40.13, and I, M. cf. parvunguis (NMS.Z.2021.40.14). J, K, right scapula (lateral view) of J, the holotype of D. phorusrhacoides and K, the tentatively referred specimen NMS.Z.2021.40.13. L–N, sternum fragments (ventral view) of L, the holotype of D. phorusrhacoides and M, N, the tentatively referred specimen NMS.Z.2021.40.13 (M: corpus in ventral view, N: caudolateral portion of left side); the arrows denote details of the spina externa. O, P, distal end of the humerus (cranial view) of O, M. cf. parvunguis (left side; NMS.Z.2021.40.14) and P, the holotype of D. phorusrhacoides (right side). Abbreviations: acr, acromion; fah, facies articularis humeralis; flx, processus flexorius; icl, incisura lateralis; icm, incisura medialis; mpr, medial projection; pcc, processus costales; pcv, processus ventralis; per, perforation in caudoventral portion of pygostyle; ppc, processus procoracoideus; spe, spina externa; spr, sternal projection on tip of processus procoracoideus; tbi, trabecula intermedia; tbl, trabecula lateralis. Scale bars equal 10 cm.

FIGURE 5. Holotype of Danielsraptor phorusrhacoides, gen. et sp. nov. from the lower Eocene London Clay of Walton-on-the-Naze, Essex, U.K. (NMS.Z.2021.40.12), pelvis and hindlimb elements. A, B, pelvis (dorsal view) and left femur in a block of matrix in dorsal (A) and lateral (B) view; C, D, right femur in cranial (C) and caudal (D) view. E, left tibiotarsus lacking distal end. F–H, left tarsometatarsus in dorsolateral (F), dorsal (G), and plantar (H) view. I, J, right tarsometatarsus in dorsal (I) and medial (J) view. K, distal end of right tarsometatarsus in distal view. L, proximal end of right tarsometatarsus in proximal view. M, os metatarsale I and pedal phalanges of the left foot. N, second phalanges of third toe (left and right). O, os metatarsale I and pedal phalanges of the extant Caracara plancus (Falconiformes, SMF 4092). P, os metatarsale I and pedal phalanges of the extant Cariama cristata (Cariamiformes; SMF 2462, the arrows delimit the borders between articulated phalanges of the fourth toe). The identity of the phalanges in M, O, and P is indicated by Roman (toes) and Arabic (phalanges) numerals. Abbreviations: ais, ala ischii; att, antitrochanter; cst, constriction; ctr, crista trochanteris; fac, foramen acetabuli; fii, foramen ilioischiadicum; flg, plantarly directed flange of trochlea metatarsi IV; fob, foramen obturatum; lfe, left femur; lip, lip-like projection formed by cotyla; mtI, os metatarsale I; pub, pubis; sul, hypotarsal sulcus for tendon of musculus flexor digitorum longus. Scale bars equal 10 mm (same scale for A–J).

Diagnosis—As for genus.
Etymology—The species epithet refers to the bill shape of the new species being similar to that of the South American Phorusrhacidae.
Type Locality and Horizon—Walton-on-the-Naze, Essex, U.K.; Walton Member of the London Clay Formation (previously Division A2; Jolley, Citation1996; Rayner et al., Citation2009; Aldiss, Citation2012); early Eocene (early Ypresian, 54.6–55 million years ago; Collinson et al., Citation2016).
Tentatively Referred Specimen—NMS.Z.2021.40.13 (, S1; partial sternum, right coracoid and scapula, partial furcula, partial pygostyle), collected in 1986 in the type locality by M. Daniels (original collector’s number WN 86549).
Measurements (in mm)—NMS.Z.2021.40.12: estimated skull length, >87; mandible length, 89.2; coracoid length, 35.4 (right), >33.4 (left); humerus distal width, >13.6 (left), 14.4 (right); femur length, 77.1 (left); femur proximal width (right), 14.1; femur distal width (right), 13.7; tibiotarsus (left) length as preserved, 85.7; tarsometatarsus length as preserved, 76.0 (right), >74.4 (left); tarsometatarsus proximal width, 12.0 (left), 11.6 (right); tarsometatarsus distal width, 12.0 (right). Pedal phalanges, I1, 14.2; II1, 11.2; II2, 15.7; III1, 20.0; III2, 19.0; IV1, 12.0; IV2, 8.2; IV3, 7.9; IV4, 12.2. NMS.Z.2021.40.13: coracoid length, 37.0 (right); scapula length as preserved, 44.0 (right).
Remarks—The tentatively referred specimen NMS.Z.2021.40.13 differs from the holotype in some features, including a somewhat straighter caudal margin of the pygostyle, a spina externa of the sternum that is not bifurcated, and a wider extremitas cranialis of the scapula with a more protruding facies articularis humeralis. These differences may eventually support an assignment of NMS.Z.2021.40.13 to a new species, but at present the basis for the distinction of two co-occurring and similar-sized Danielsraptor species in the London Clay of Walton-on-the-Naze is weak.
Description and Comparison—In the holotype, substantial portions of the skull are preserved (). The long and mediolaterally compressed beak (, B) exhibits a distinctive shape and closely resembles that of Masillaraptor parvunguis (see Mayr, Citation2009). The rostrum has a sharply hooked tip. The tomia (cutting edges) are sigmoidally curved and just caudal of the tip of beak they form “tomial grooves” (sensu Jollie, Citation1977) rather than sharp cutting edges (these grooves are also present in crown group Falconiformes). The nostrils appear to have been fairly large and dorsally positioned, but their exact shape cannot be determined, since the surrounding portions of the beak are broken (the dorsal nasal bar is preserved as an isolated element; ).
The lacrimal () has a well-developed, tapering processus supraorbitalis and a long descending process, which appears to have contacted the jugal, so that an os lacrimale communicans is likely to have been absent (this peculiar ossicle represents an ossified ligamentum jugolacrimale and is a derived characteristic of the Cariamiformes; Mayr, Citationin press). Both jugal bars are preserved and are comparatively stout (). The interorbital section of the frontal bones is fairly long and moderately wide (). The base of the right processus postorbitalis is preserved and suggests that this process was well developed. This also appears to be true for the fossae temporales, of which only the dorsal portion of the right one is preserved. The caudal and ventral portions of the neurocranium are broken. Two unidentified ossicles may represent the caudal portions of the pterygoids (); however, the wide and irregularly shaped end of the bones is unlike the pterygoid of both crown group Falconiformes and crown group Cariamiformes.
The holotype includes the left quadrate, which is largely complete except for the broken processus orbitalis (). The processus oticus lacks an incisura intercapitularis and its dorsal margin has a subtriangular outline with a raised central portion. There are no pneumatic foramina on the caudal surface of the processus oticus, and the foramen pneumaticum is situated in the caudomedial portion of the quadrate. The bone resembles the quadrate of crown group Cariamiformes () and differs from the quadrate of crown group Falconiformes () in the lack of an incisura intercapitularis, the rostrocaudally shorter condylus medialis, and the less laterally projecting condylus lateralis.
The mandible () corresponds to that of Masillaraptor parvunguis in its shape. The tip of the long and mediolaterally narrow symphysis is upcurved and broadly rounded. The rami are dorsoventrally deep. A distinctive character of the caudal articular end is a marked dorsal projection, which borders the cotyla lateralis rostrally (, ); this projection is also found in crown group Falconiformes () but is absent in extant and fossil Cariamiformes (). The processus medialis is short.
Even though the holotype specimen comprises various fragments of the vertebrae (, ), these show only a few characters of potential taxonomic or phylogenetic significance. Cranial cervical vertebrae are absent in both specimens, and the caudal cervical and thoracic vertebrae are unremarkable (, ). A notarium appears to have been absent. One of the caudal vertebrae is notable in that it exhibits a very long processus ventralis (, ). The pygostyle is large and has a wide lamina pygostyli. The pygostyle of the tentatively referred specimen NMS.Z.2021.40.13 is craniocaudally narrower than that of the holotype and has a somewhat straighter caudal margin ().
The coracoid (, ) resembles that of crown group Falconiformes in its proportions and morphological features and is clearly distinguished from the coracoid of fossil and extant Cariamiformes. The sulcus supracoracoideus exhibits a fossa, which undercuts the facies articularis clavicularis (). The ventral portion of the facies articularis clavicularis is elongated in sternal direction of the bone (). The long processus procoracoideus is visible in the tentatively referred specimen NMS.Z.2021.40.13 (), whereas the process is broken in both coracoids of the holotype. Unlike in crown group Falconiformes, there is a deeply excavated cotyla scapularis (), which is a plesiomorphic characteristic of neornithine birds (Mayr, Citation2021). The foramen nervi supracoracoidei () is small. The medial margin of the sternal extremity forms a distinct convexity in the holotype and a small projection in the tentatively referred specimen NMS.Z.2021.40.13 (); the angulus medialis is pointed in both specimens.
The scapula (, ) is a stout bone with a curved corpus and a short acromion. It has similar proportions to the scapula of the late Eocene North American cariamiform species Bathornis grallator (Wetmore, Citation1944) (Bathornithidae; Mayr, Citation2016). In the tentatively referred specimen NMS.Z.2021.40.13, the extremitas cranialis is wider than in the holotype and has a more protruding facies articularis humeralis ().
The furcula (, ) has a wide sternal extremity and strap-like shafts. A well-developed apophysis furculae is absent, with the extremitas sternalis exhibiting only a low ridge. The omal extremity is poorly differentiated and—unlike in crown group Falconiformes—it lacks a pronounced facies articularis acrocoracoidea.
The holotype includes only the cranialmost portion of the sternum and the short spina externa has a slightly bifurcated tip (). The sternum of the tentatively referred specimen NMS.Z.2021.40.13 is more complete and exhibits fragments of the caudolateral section (). Unlike in the holotype, the tip of the spina externa is not bifurcated in NMS.Z.2021.40.13 (). The sulci articulares coracoidei are overlapping. The caudal margin appears to have featured two pairs of incisions, even though the trabecula intermedia is broken.
Only fragments of the proximal and distal ends of the humerus are present in the holotype (). On the distal end of the bone, there is no processus flexorius. A fragment of the distal end of the ulna shows an unspecific morphology ().
The pelvis () has a long preacetabular portion of similar overall proportions to the preacetabular portion of the pelvis of crown group Falconiformes (in the Cariamiformes, the preacetabular portion is mediolaterally narrower). As in crown group Falconiformes, the cristae iliacae dorsales are co-ossified with the crista spinosa of the synsacrum. The ala ischii is dorsoventrally deep as it is in crown group Falconiformes (in crown group Cariamiformes, the ala is less deep). Unlike in crown group Cariamiformes, there is not laterally projecting process dorsal of the antitrochanter. Well-defined tubercula praeacetabularia are absent.
The left femur is nearly complete and preserved in articulation with the pelvis (). The bone is fairly long. As in crown group Falconiformes, the proximal end exhibits a well-developed crista trochanteris (), but unlike in extant Falconiformes there is no pneumatic foramen on its cranial surface. Also as in crown group Falconiformes, the caput femoris bears a deep fovea ligamenti capitis; the impressiones obturatoriae form two marked depressions. There is a distinct linea intermuscularis along the cranial surface of the shaft. The sulcus patellaris on the distal end is narrow. In its overall proportions and morphology, the bone more closely resembles the femur of crown group Falconiformes than the stouter femur of the Cariamiformes.
The holotype comprises the proximal portion of the left tibiotarsus (), which resembles the tibiotarsus of crown group Falconiformes but does not exhibit notable features of phylogenetic significance. Of the distal end, only a fragment of the condylus lateralis is preserved.
The tarsometatarsus () is a fairly long bone and shows a superficial resemblance to the tarsometatarsus of the North American Bathornithidae (Mayr, Citation2016) and the European Idiornithidae (Dynamopterus and Ibidopodia; Mourer-Chauviré, Citation2013; De Pietri and Mayr, Citation2014), both being stem group representatives of the Cariamiformes. However, the eminentia intercotylaris is not as prominent as in cariamiform birds and the trochlea metatarsi II is shaped differently, forming a plantar projection; furthermore, the bone is not as elongated as the tarsometatarsus of cariamiform birds. The hypotarsus is block-like with a shallow, medially situated sulcus for the tendon of the flexor digitorum longus muscle (); hypotarsal canals are absent. This morphology is more similar to the hypotarsus of the Cariamiformes, whereas the hypotarsus of the Falconiformes forms a prominent crest. A canalis interosseus distalis is present. The trochlea metatarsi II forms a plantarly directed projection. The lateral surface of the trochlea metatarsi III bears a notch. The trochlea metatarsi IV exhibits a plantarly directed, wing-like projection.
The os metatarsale I () has a relatively short processus tarsometatarsalis with a broadly rounded tip. Altogether, 16 non-ungual pedal phalanges of both feet are preserved in the holotype (, S1). The first phalanx of the hallux is moderately long, whereas it is strongly abbreviated in crown group Cariamiformes (). The first phalanx of the second toe is shorter than the second (penultimate) one. The second phalanx of the third toe exhibits an unusual derived morphology in that the ventral portion of the cotyla forms a marked lip-like projection and in that the distal end of the phalanx bears a constriction on its plantar surface (). The second and third phalanges of the fourth toe are abbreviated, as they are in crown group Falconiformes and Cariamiformes (); unlike in the Cariamiformes, the third phalanx is not shortened. The ungual phalanges are long and sharply pointed. However, unlike in crown group falconiforms, the tuberculum flexorium is only weakly developed and the sulcus neurovascularis is laterally open.
MASILLARAPTOR Mayr, Citation2006
MASILLARAPTOR cf. M. PARVUNGUIS Mayr, Citation2006
Referred Specimens—NMS.Z.2021.40.14 (, S1; partial skeleton including most major limb bones), collected in 1993 by M. Daniels (original collector’s number WN 93777). NMS.Z.2021.40.15 (, S1; partial wing and pectoral girdle bones), collected in 1993 by M. Daniels (original collector’s number WN 93780A).
FIGURE 6. Leg bones of Masillaraptor cf. parvunguis from the lower Eocene London Clay of Walton-on-the-Naze, Essex, U.K. (A–I) compared with other fossils of Masillaraptor (J, K), as well as London Clay specimens of Coturnipes (L, M), Danielsraptor, gen. nov. (N, O), and Stintonornis (P, Q). A, Masillaraptor cf. parvunguis, proximal end of left femur in cranial view (NMS.Z.2021.40.14). B, M. cf. parvunguis, distal end of right femur in caudal view (NMS.Z.2021.40.14). C, M. cf. parvunguis, distal end of left tibiotarsus in cranial view (NMS.Z.2021.40.14). D, M. cf. parvunguis, distal end of right tibiotarsus in cranial view (NMS.Z.2021.40.14). E, F, M. cf. parvunguis, left tarsometatarsus in dorsal (E) and plantar (F) view (NMS.Z.2021.40.14). G–I, M. cf. parvunguis, right tarsometatarsus in dorsal (G), plantar (H), and distal (I) view (NMS.Z.2021.40.14). J, left tarsometatarsus of Masillaraptor sp. from Messel in plantar view (SMF-ME 1068). K, left tarsometatarsus of Masillaraptor sp. from the middle Eocene of the Geisel Valley in plantar view (GMH NW XIV). L, M, distal end of the left tarsometatarsus of Coturnipes cooperi from the London Clay of the Isle of Sheppey in L, dorsal and M, plantar view (holotype, NHMUK A 3706). N, O, right tarsometatarsus of Danielsraptor phorusrhacoides, gen. et sp. nov. (NMS.Z.2021.40.12) in N, plantar and O, dorsal view. P, Q, distal end of the right tarsometatarsus of Stintonornis mitchelli from the London Clay of the Isle of Sheppey in P, plantar and Q, dorsal view (holotype, NHMUK A 5284). R, pedal phalanges of Masillaraptor cf. parvunguis (NMS.Z.2021.40.14); the identity of some phalanges is indicated by Roman (toes) and Arabic (phalanges) numerals. For the tarsometatarsi, the trochleae are numbered. Abbreviations: fvd, foramen vasculare distale; mtI, fossa metatarsi I; pst, pons supratendineus. Scale bars equal 5 mm.
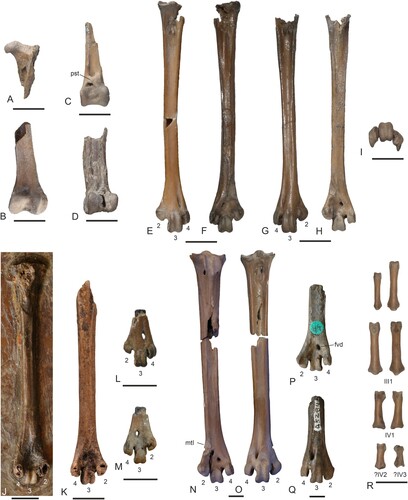
FIGURE 7. Vertebrae as well as pectoral girdle and wing elements of Masillaraptor cf. parvunguis from the lower Eocene London Clay of Walton-on-the-Naze, Essex, U.K. A, three cervical vertebrae in different views (NMS.Z.2021.40.14). B, three thoracic vertebrae in lateral view (NMS.Z.2021.40.14). C, caudal vertebra in dorsal and lateral view (NMS.Z.2021.40.14). D, three caudal vertebrae in different views (NMS.Z.2021.40.14). E, pygostyle in lateral view (NMS.Z.2021.40.14). F, G, right coracoid in ventral (F) and dorsal (G) view (NMS.Z.2021.40.14). H, I, left coracoid in dorsal (H) and ventral (I) view (NMS.Z.2021.40.14). J, left coracoid in dorsal view (NMS.Z.2021.40.15). K, L, cranial extremities of left (K) and right (L) scapula (NMS.Z.2021.40.15). M, N, furcula (M: NMS.Z.2021.40.14; N: NMS.Z.2021.40.15). O, sternum (ventral view) and proximal portion of left carpometacarpus (ventral view) in a small block of matrix (NMS.Z.2021.40.14). P, Q, partial right humerus in caudal and cranial view (NMS.Z.2021.40.14). R, distal end of left humerus in cranial view (NMS.Z.2021.40.14). S, T, distal ends of right (S) and left (T) humerus in cranial view (NMS.Z.2021.40.15). U, proximal portion of right ulna in cranial view (NMS.Z.2021.40.15). V, partial left carpometacarpus in ventral view (NMS.Z.2021.40.14). W, proximal portion of right carpometacarpus in ventral view (NMS.Z.2021.40.14). X, left phalanx proximalis digiti majoris (NMS.Z.2021.40.14). Y, left and right os carpi ulnare (NMS.Z.2021.40.15). Z, os carpi radiale and wing phalanges (NMS.Z.2021.40.14). Abbreviations: cmc, carpometacarpus; ila, incisura lateralis; imd, incisura medialis; ote, ossified tendons; pcl, processus craniolateralis; pcv, processus ventralis; pla, processus lateralis. Scale bar equals 10 mm.

FIGURE 8. Comparison of selected postcranial elements of the Masillaraptoridae, crown group Falconiformes, and crown group Cariamiformes. A–D, coracoid (dorsal view) of A, Masillaraptor cf. parvunguis (NMS.Z.2021.40.14), B, Danielsraptor phorusrhacoides (NMS.Z.2021.40.13), C, Caracara plancus (Falconiformes; SMF 3462), and D, Cariama cristata (Cariamiformes; SMF 1862). E–G, right humerus (caudal view) of E, Ma. cf. parvunguis (NMS.Z.2021.40.14), F, Micrastur ruficollis (Falconiformes; SMF 9812), and G, C. cristata (SMF 1862). H, I, distal end of left humerus (cranial view) of H, Ma. cf. parvunguis (NMS.Z.2021.40.14) and I, Mi. ruficollis (SMF 9812). J–L, proximal end of right ulna (cranial view) of J, Ma. cf. parvunguis (NMS.Z.2021.40.14), K, Mi. ruficollis (SMF 9812), and L, C. cristata (SMF 1862). M–O, left carpometacarpus of M, Ma. cf. parvunguis (NMS.Z.2021.40.15), N, Mi. ruficollis (SMF 9812; right side, mirrored), and O, C. cristata (SMF 1862; right side, mirrored). P–R, pelvis (dorsal view) of P, D. phorusrhacoides (holotype, NMS.Z.2021.40.12; surrounding matrix digitally brightened), Q, Mi. ruficollis (SMF 9812), and R, C. cristata (SMF 1862). S, T, left femur (cranial view) of S, D. phorusrhacoides (holotype, NMS.Z.2021.40.12; surrounding matrix digitally brightened) and T, Mi. ruficollis (SMF 9812). U–AA, proximal (V, X, Z) and distal (U, W, Y, AA) ends of the right tarsometatarsus of U, Ma. cf. parvunguis (NMS.Z.2021.40.14), V, W, D. phorusrhacoides (holotype, NMS.Z.2021.40.12), X, Y, C. plancus (SMF 6441), and Z, AA, C. cristata (SMF 1862). Abbreviations: ctd, cotyla dorsalis, ctr, crista trochanteris; ctv, cotyla ventralis; ecr, insertion scar for musculus extensor carpi radialis; flg, wing-like flange; flx, processus flexorius, fns, foramen nervi supracoracoidei; pex, processus extensorius; pnf, pneumatic foramen; sul, hypotarsal sulcus. Scale bars equal 10 mm.

Locality and Horizon—Walton-on-the-Naze, Essex, U.K.; Walton Member of the London Clay Formation; early Eocene (early Ypresian).
Measurements (in mm)—NMS.Z.2021.40.14: Coracoid length, 20.6 (left)/21.8 (right); humerus length as preserved, 37.9 (right); estimated total humerus length, ∼42–43 [∼46]; ulna length as preserved, 42.1 (right); estimated total ulna length, ∼46 [∼49]; tarsometatarsus length, 37.0 (left)/>35.0 (right) [∼34.7/35.2]; tarsometatarsus distal width, 5.5 (left)/5.5 (right). NMS.Z.2021.40.15: Carpometacarpus length, 25.6 (left). The values in brackets indicate the dimensions of the holotype of Masillaraptor parvunguis (from Mayr, Citation2006).
Remarks—NMS.Z.2021.40.14 was previously considered to be conspecific with Coturnipes cooperi, a species described by Harrison and Walker (Citation1977) on the basis of a distal tarsometatarsus from the London Clay of the Isle of Sheppey (Daniels, Citation1995; Olson, Citation1999; Mayr, Citation2006). However, NMS.Z.2021.40.14 has a proportionally wider trochlea metatarsi II and a more distally situated foramen vasculare distale than C. cooperi (), and close affinities––let alone an identity at species level––are no longer maintained (Mayr and Smith, Citation2019; Mayr, Citation2022).
Although the skull is not preserved in the fossils from Walton-on-the-Naze, they can be unambiguously assigned to the Masillaraptoridae on the basis of the distinctive morphology of the postcranial elements. In addition to a similar overall morphology of all the major bones, the London Clay fossils (NMS.Z.2021.40.14 and NMS.Z.2021.40.15) correspond with Danielsraptor phorusrhacoides, gen. et sp. nov. in, e.g., the shape of the coracoid (which exhibits a foramen nervi supracoracoidei and a medial projection on the sternal end); the very large pygostyle; the fact that one of the caudal vertebrae bears a very long processus ventralis; the shape of the long tarsometatarsus; the abbreviated second and third phalanges of the fourth toe; and the raptor-like shape of the ungual phalanges.
Description and Comparisons—As far as comparisons with the compression fossils of Masillaraptor parvunguis from Messel are possible, NMS.Z.2021.40.14 and NMS.Z.2021.40.15 show a close resemblance to this species in size and morphology. Apart from their significantly smaller size, the specimens also correspond well to Danielsraptor phorusrhacoides, gen. et sp. nov. in the morphology of most bones, even though there are some salient differences. The following description focuses on features that have not yet been described for the M. parvunguis fossils from Messel, or in which M. parvunguis is distinguishable from D. phorusrhacoides.
NMS.Z.2021.40.14 includes some cervical, thoracic, and caudal vertebrae (). Two of the thoracic vertebrae exhibit ossified tendons that are co-ossified with the processus spinosus (); this observation is notable, because Mayr (Citation2009) noted that in a specimen of M. parvunguis from Messel the processus spinosi appear to be conjoined. One of the caudal vertebrae has a very long processus ventralis. The pygostyle () is of similar shape to that of D. phorusrhacoides, but the dorsal portion is craniocaudally wider and the bone exhibits a perforation in its caudoventral section ().
Details of the coracoid cannot be discerned in the specimens of M. parvunguis from Messel. In the fossils from the London Clay, the bone is well preserved (). The extremitas omalis is proportionally shorter than in D. phorusrhacoides, and the long processus procoracoideus is more strongly bent towards the omal extremity and its tip lacks a sternally directed projection (). The shaft of the bone exhibits a very small foramen nervi supracoracoidei. The processus lateralis, which is not preserved in the fossils of D. phorusrhacoides, forms a pointed projection.
As in D. phorusrhacoides, the U-shaped furcula has craniocaudally compressed, strap-like shafts and a wide sternal extremity without a well-developed apophysis furculae (). The omal extremity of the bone is simple and terminates with a straight end.
The corpus of the sternum () has a subrectangular outline, being much longer than wide. The processus craniolaterales are short. The caudal margin exhibits four incisions, with the lateral ones being considerably deeper than the medial ones.
The wing bones of NMS.Z.2021.40.14 show a close resemblance to those of crown group Falconiformes and are very different from the corresponding elements of the Cariamiformes, both fossil and extant (). The humerus () in particular has a similar overall shape to that of crown group Falconiformes (). The tuberculum dorsale is small. The crista deltopectoralis is dorsally prominent and has a concave outline in its proximal section. The condylus dorsalis, on the distal end of the bone, is narrow. The processus flexorius is better developed than in D. phorusrhacoides. The insertion scar for the musculus extensor carpi radialis forms a distinct, transverse ridge ().
NMS.Z.2021.40.14 includes a partial ulna, which lacks the distal end (). The proximal end is similar to the proximal ulna of the Falconiformes (); the cotyla dorsalis is large and reaches farther distally than the cotyla ventralis.
The carpometacarpus () also resembles the corresponding bone of crown group Falconiformes. The proximal end features a deep fossa infratrochlearis, which is situated caudal to the processus pisiformis; the processus extensorius is moderately developed. The spatium intermetacarpale is narrow, whereas it is very wide in extant Cariamiformes and fossil stem group cariamiforms, such as the Idiornithidae (Dynamopterus) and Bathornithidae.
The phalanx proximalis digiti majoris () exhibits a processus internus indicis, which was—probably erroneously owing to the poor preservation of the fossils—considered to be absent in the specimens of M. parvunguis from Messel (Mayr, Citation2006).
As far as comparisons are possible, the femur () is similar to that of D. phorusrhacoides. As in the latter species, the proximal end has a well-developed crista trochanteris, and, unlike in crown group Falconiformes (), there is no pneumatic foramen on its cranial surface.
The distal ends of both tibiotarsi are preserved in NMS.Z.2021.40.14 (), but the right one is distorted and does not allow the recognition of osteological details. The bone exhibits a medially situated sulcus extensorius and a proximodistally broad pons supratendineus. The condyles are widely separated and the condylus medialis is narrower than the condylus lateralis. The distal end of the bone is very different from the distal tibiotarsus of extant Falconiformes, which has a unique apomorphic morphology with an additional opening lateral of the canalis extensorius.
The tarsometatarsus () resembles that of D. phorusrhacoides in its proportions and overall morphology, but has a somewhat narrower shaft, a proportionally shorter trochlea metatarsi II, and a less developed plantar flange on the trochlea metatarsi IV. The hypotarsus is not preserved in the fossils.
NMS.Z.2021.40.14 includes several pedal phalanges, including abbreviated phalanges of the fourth toe (; the complete set of phalanges is preserved in the Messel specimens of M. parvunguis; Mayr, Citation2006).
DISCUSSION
Masillaraptorids resemble cariamiform birds in the block-like hypotarsus (), and their long, deep, and mediolaterally compressed beak shows a resemblance to that of the cariamiform Phorusrhacidae (). However, even though Daniels (Citation1995) considered the holotype specimen of Danielsraptor phorusrhacoides, gen. et sp. nov. to be from a volant representative of the Phorusrhacidae, masillaraptorids lack many synapomorphies uniting the Phorusrhacidae and crown group Cariamiformes (Cariamidae) and are outside a clade formed by the latter two taxa. In our analysis, four characters were optimized as synapomorphies of a clade including the Phorusrhacidae and Cariamidae: a very short ulna, which measures only about 3/4 of the humerus length (ch. 22), a mediolaterally compressed pelvis (ch. 28), a femur without a well-developed crista trochanteris (ch. 31), and a tibiotarsus with a prominent tubercle distal of the pons supratendineus (ch. 33). Derived characters shared by the Phorusrhacidae and Cariamidae, which are absent in the Masillaraptoridae, furthermore include a reduced acrocoracoid process of the coracoid, a carpometacarpus with a wide intermetacarpal space (), a short hallux, and an abbreviated penultimate phalanx of the fourth toe (). In addition, the long descending process of the lacrimal bone of masillaraptorids indicates the absence of an os lacrimale communicans, the presence of which is a derived characteristic of the Cariamiformes (Mayr, Citationin press).
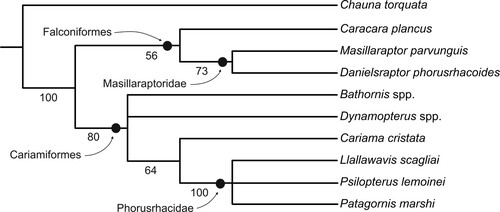
Our phylogenetic analysis also corroborated a position of masillaraptorids outside the Cariamiformes. This analysis resulted in two most parsimonious trees (; L = 64, CI = 0.67, RI = 0.73) and supported a sister group relationship between Danielsraptor phorusrhacoides and Masillaraptor parvunguis as well as a sister group relationship between the Masillaraptoridae and crown group Falconiformes. Four characters were optimized as synapomorphies of the Masillaraptoridae and Falconiformes: a beak with a sharply hooked tip (ch. 8) and distinct tomial grooves (ch. 9), a mandible with a marked dorsal projection rostral of the cotyla lateralis (ch. 13), and an abbreviated first phalanx of the second toe (ch. 38). The second and third of these characters (tomial groves and mandibular projection) are diagnostic apomorphies of Pan-Falconiformes, the clade that includes the Masillaraptoridae and crown group Falconiformes.
FIGURE 9. Strict consensus tree of the two most parsimonious trees (L = 64, CI = 0.67, RI = 0.73) resulting from the phylogenetic analysis; unsupported nodes were collapsed. Bootstrap support values are indicated next to the internodes.
Falconiform affinities of masillaraptorids are also suggested by the fact that similar tarsometatarsi were previously assigned to the Falconiformes. This is true for Stintonornis mitchelli Harrison, Citation1984 from the London Clay of the Isle of Sheppey (; Harrison, Citation1984) and for Antarctoboenus carlinii Cenizo et al., Citation2016 from the early Eocene of Antarctica (Cenizo et al., Citation2016), of which only distal ends of the tarsometatarsi are known.
The strata of the London Clay at Walton-on-the-Naze are from division A2 of the London Clay (Rayner et al., Citation2009) and date back to 55–54.6 Ma (Collinson et al., Citation2016). The London Clay exposed at the Isle of Sheppey, which yielded the holotype of Stintonornis mitchelli, is from the younger divisions C–F (51.5–53.5 Ma; Collinson et al., Citation2016). The holotype of Antarctoboenus carlinii has an age of 52.8–49 million years (Cenizo et al., Citation2016). Therefore, both species are younger than the masillaraptorids from Walton-on-the-Naze. The proportionally larger trochlea metatarsi II may suggest that Stintonornis mitchelli is more closely related to crown group Falconiformes, but more data on the osteology of this taxon are needed for a well-founded phylogenetic placement. Likewise, the exact affinities between Antarctoboenus carlinii and masillaraptorids cannot be established without more information on the skeletal morphology of the former taxon.
Even though masillaraptorids are here considered to be stem group representatives of the Falconiformes, the last common ancestor of crown group Falconiformes is likely to have lived in the New World, because the South American Herpetotherinae and Polyborinae are successive sister taxa of other crown group Falconiformes (Fuchs et al., Citation2015). Molecular data yielded a 34.2-million-year maximum time estimate for the earliest divergences within crown group Falconiformes, in the earliest Oligocene (Fuchs et al., Citation2015). This date is much younger than the latest (middle Eocene) occurrence of the Masillaraptoridae. The Antarctic taxon Antarctoboenus is likewise at least 15 million years older than the presumed split of crown group Falconiformes, and this taxon is more likely to have had a masillaraptorid-like morphology than showing a close resemblance to extant falcons.
Most differences between the Masillaraptoridae and crown group Falconiformes are due to the retention of plesiomorphic characters in masillaraptorids. For example, this is true for the cup-like cotyla scapularis of the coracoid (see Mayr, Citation2021), the morphology of the distal end of the tibiotarsus, the block-like hypotarsus, the proportionally smaller and shorter trochlea metatarsi II, and the different morphology of the ungual phalanges (which have a more pronounced tuberculum flexorium and a closed sulcus neurovascularis in extant Falconiformes).
The long legs of masillaraptorids are probably also plesiomorphic for the Falconiformes, because early diverging taxa of crown group Falconiformes have proportionally longer legs than typical falcons (Falconinae). The leg proportions of masillaraptorids may indicate a predominantly terrestrial way of living, and these birds may have searched for food (such as larger arthropods or small vertebrates) on the ground, as do extant caracaras (Caracara spp.). On the other hand, there are some comparatively long-legged crown group Falconiformes, which do not forage on the ground (e.g., the species of the South American Herpetotherinae), so that the exact foraging habits of masillaraptorids remain elusive. The large pygostyle suggests the presence of long tail feathers, which, together with the long ulna, indicate well-developed flight capabilities. The peculiar shape of the second phalanx of the third toe is notable, but the functional significance of this feature remains elusive. The lip-like projection formed by the cotyla of this phalanx may indicate that masillaraptorids had a grasping foot, but this assumption contrasts with the fact that the tubercula flexoria of the ungual phalanges of masillaraptorids are rather poorly developed (by contrast, in crown Falconiformes the ungual phalanges have prominent flexor tubercles, whereas the tuberculum flexorium of the second phalanx of the third toe is weakly developed).
The beak of masillaraptorids is reminiscent of that of the cariamiform Phorusrhacidae () but also shows a resemblance to the beak of the falconiform caracaras (Polyborinae; ). Extant Cariamiformes and Falconiformes share only a few derived characters, including a lacrimal bone with a long processus supraorbitalis as well as abbreviated central phalanges of the fourth toe and raptor-like ungual phalanges. In most current sequence-based analyses, the Cariamiformes and Falconiformes are successive sister taxa of the clade including the Psittaciformes and Passeriformes (Ericson et al., Citation2006; Hackett et al., Citation2008; Prum et al., Citation2015; Kuhl et al., Citation2021). By contrast, unpartitioned coding exons support a sister group relationship between the Cariamiformes and the Falconiformes (Braun and Kimball, Citation2021:fig. 4D). The mosaic of derived falconiform features and presumably plesiomorphic cariamiform characters (e.g., a block-like hypotarsus) present in masillaraptorids potentially shows that the latter represents an evolutionary link between the Falconiformes and the Cariamiformes and lends some support to a sister group relationship between the Cariamiformes and Falconiformes (Braun and Kimball, Citation2021:fig. 4D). Alternatively, the similarities between masillaraptorids and cariamiform birds may represent parallelisms in closely related taxa, or they may be plesiomorphic for the clade including the Cariamiformes, Falconiformes, Psittaciformes, and Passeriformes, the stem species of which would then most likely have been a long-legged bird with a raptorial way of living.
Supplemental Material
Download Zip (1.5 MB)ACKNOWLEDGMENTS
We thank S. Tränkner for taking most of the photographs (additional images were taken by GM). ACK thanks D. Bain for his considerable assistance in the transfer of the Daniels collection to NMS. G. Hantke helped with the packing and transport of the specimens. Comments from the Associate Editor V. De Pietri and two anonymous reviewers improved the manuscript.
LITERATURE CITED
- Aldiss, D. T. 2012. The stratigraphical framework for the Palaeogene successions of the London Basin, UK. British Geological Survey Open Report OR/12/004:1–87.
- Braun, E. L., and R. T. Kimball. 2021. Data types and the phylogeny of Neoaves. Birds 2:1–22.
- Cenizo, M., J. I. Noriega, and M. A. Reguero. 2016. A stem falconid bird from the lower Eocene of Antarctica and the early southern radiation of the falcons. Journal of Ornithology 157:885–894.
- Collinson, M. E., N. F. Adams, S. R. Manchester, G. W. Stull, F. Herrera, S. Y. Smith, M. J. Andrew, P. Kenrick, and D. Sykes. 2016. X-ray micro-computed tomography (micro-CT) of pyrite-permineralized fruits and seeds from the London Clay Formation (Ypresian) conserved in silicone oil: a critical evaluation. Botany 94:697–711.
- Daniels, M. 1992. Untitled report; in C. Mourer-Chauviré (ed.), Society of Avian Paleontology and Evolution, information letter n° 6, October 1992. http://www.sapesociety.org/wp-content/uploads/2014/10/newsletter1992.pdf. Accessed 11 Jan. 2021.
- Daniels, M. 1995. Untitled report; in C. Mourer-Chauviré (ed.), Society of Avian Paleontology and Evolution, information letter n° 9, October 1992. http://www.sapesociety.org/wp-content/uploads/2014/10/newsletter1995.pdf. Accessed 11 Jan. 2021.
- Degrange, F. J., C. P. Tambussi, M. L. Taglioretti, A., Dondas, and F. Scaglia. 2015. A new Mesembriornithinae (Aves, Phorusrhacidae) provides new insights into the phylogeny and sensory capabilities of terror birds. Journal of Vertebrate Paleontology 35:e912656.
- De Pietri, V. L., and G. Mayr. 2014. The enigmatic Ibidopodia palustris from the early Miocene of France – the first Neogene record of Cariamiformes (Aves) in Europe. Journal of Vertebrate Paleontology 34:1470–1475.
- Ericson, P. G. P., C. L. Anderson, T. Britton, A. Elzanowski, U. S. Johansson, M. Källersjö, J. I. Ohlson, T. J. Parsons, D. Zuccon, and G. Mayr. 2006. Diversification of Neoaves: integration of molecular sequence data and fossils. Biology Letters 2:543–547.
- Fuchs, J., J. A. Johnson, and D. P. Mindell. 2015. Rapid diversification of falcons (Aves: Falconidae) due to expansion of open habitats in the late Miocene. Molecular Phylogenetics and Evolution 82:166–182.
- Goloboff, P. A. 1993. NONA version 2.0 [Computer software]. Published by the author, S. M. de Tucumán.
- Grande, L. 2013. The lost world of Fossil Lake: Snapshots from deep time. University of Chicago Press, Chicago, 425 pp.
- Hackett, S. J., R. T. Kimball, S. Reddy, R. C. K. Bowie, E. L. Braun, M. J. Braun, J. L. Chojnowski, W. A. Cox, K.-L. Han, J. Harshman, C. J. Huddleston, B. D. Marks, K. J. Miglia, W. S. Moore, F. H. Sheldon, D. W. Steadman, C. C. Witt, and T. Yuri. 2008. A phylogenomic study of birds reveals their evolutionary history. Science 320:1763–1767.
- Harrison, C. J. O. 1984. Further additions to the fossil birds of Sheppey: A new falconid and three small rails. Tertiary Research 5:179–187
- Harrison, C. J. O., and C. A. Walker. 1977. Birds of the British lower Eocene. Tertiary Research, Special Paper 3:1–52.
- Jolley, D. W. 1996. The earliest Eocene sediments of eastern England: an ultra-high resolution palynological correlation. Geological Society, London, Special Publications 101:219–254.
- Jollie, M. 1977. A contribution to the morphology and phylogeny of the Falconiformes (part II). Evolutionary Theory 2:115–208.
- Kuhl, H., C. Frankl-Vilches, A. Bakker, G. Mayr, G. Nikolaus, S. T. Boerno, S. Klages, B. Timmermann, and M. Gahr. 2021. An unbiased molecular approach using 3’UTRs resolves the avian family-level tree of life. Molecular Biology and Evolution 38:108–127.
- Li, Z., Z. Zhou, T. Deng, Q. Li, and J. A. Clarke. 2014. A falconid from the late Miocene of northwestern China yields further evidence of transition in late Neogene steppe communities. The Auk: Ornithological Advances 131:335–350.
- Linnaeus, C. 1758. Systema naturae per regna tria naturae. 10th edition, 2 vols. L. Salmii, Holmiae, 824 pp.
- Mayr, G. 2002. Avian remains from the middle Eocene of the Geiseltal (Sachsen-Anhalt, Germany); pp. 77–96 in Z. Zhou and F. Zhang (eds.), Proceedings of the 5th Symposium of the Society of Avian Paleontology and Evolution, Beijing, 1-4 June 2000. Science Press, Beijing.
- Mayr, G. 2006. A new raptorial bird from the middle Eocene of Messel, Germany. Historical Biology 18:99–106.
- Mayr, G. 2009. A well-preserved skull of the “falconiform” bird Masillaraptor from the middle Eocene of Messel (Germany). Palaeodiversity 2:315–320.
- Mayr, G. 2016. Osteology and phylogenetic affinities of the middle Eocene North American Bathornis grallator - one of the best represented, albeit least known Paleogene cariamiform birds (seriemas and allies). Journal of Paleontology 90:357–374.
- Mayr, G. 2017. Avian evolution: the fossil record of birds and its paleobiological significance. Wiley-Blackwell, Chichester, 293 pp.
- Mayr, G. 2020. An updated review of the middle Eocene avifauna from the Geiseltal (Germany), with comments on the unusual taphonomy of some bird remains. Geobios 62:45–59.
- Mayr, G. 2021. The coracoscapular joint of neornithine birds––extensive homoplasy in a widely neglected articular surface of the pectoral girdle and its functional correlates. Zoomorphology 140:217–228.
- Mayr, G. 2022. Paleogene fossil birds, second edition. Springer, Cham, 239 pp.
- Mayr, G. In press. A survey of the uncinate bone and other poorly known ossicles associated with the lacrimal/ectethmoid complex of the avian skull. The Anatomical Record.
- Mayr, G., and T. Smith. 2019. A diverse bird assemblage from the Ypresian of Belgium furthers knowledge of early Eocene avifaunas of the North Sea Basin. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 291:253–281.
- Milne-Edwards, A. 1892. Sur les oiseaux fossiles des dépôts éocènes de phosphate de chaux du Sud de la France. Comptes Rendus du second Congrès Ornithologique International, Budapest, 1891:60–80.
- Moreno, F. P., and A. Mercerat. 1891. Catálogo de los pájaros fósiles de la República Argentina conservados en el Museo de La Plata. Anales del Museo de La Plata 1:7–71.
- Mourer-Chauviré, C. 1983. Les Gruiformes (Aves) des Phosphorites du Quercy (France). 1. Sous-ordre Cariamae (Cariamidae et Phorusrhacidae). Systématique et biostratigraphie. Palaeovertebrata 13:83–143.
- Mourer-Chauviré, C. 2013. Idiornis Oberholser, 1899 (Aves, Gruiformes, Cariamae, Idiornithidae): A junior synonym of Dynamopterus Milne-Edwards, 1892 (Paleogene, Phosphorites du Quercy, France). Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 270:13–22.
- Nixon, K. C. 2002. WinClada, version 1.00.08 [Computer software]. Published by the author, Ithaca, NY.
- Olson, S. L. 1999. Early Eocene birds from eastern North America: a faunule from the Nanjemoy Formation of Virginia; pp. 123–132 in R. E. Weems and G. J. Grimsley (eds.), Early Eocene vertebrates and plants from the Fisher/Sullivan site (Nanjemoy Formation) Stafford County, Virginia. Virginia Division of Mineral Resources Publication 152.
- Prum, R. O., J. S. Berv, A. Dornburg, D. J. Field, J. P. Townsend, E. M. Lemmon, and A. R. Lemmon. 2015. A comprehensive phylogeny of birds (Aves) using targeted next-generation DNA sequencing. Nature 526:569–573.
- Rayner, D., T. Mitchell, M. Rayner, and F. Clouter. 2009. London Clay fossils of Kent and Essex. Medway Fossil and Mineral Society, Rochester, Kent, 228 pp.
- Sharpe, R. B. 1874. Catalogue of the Accipitres, or diurnal birds of prey, in the collection of the British Museum (volume 1). Trustees of the British Museum, London, 479 pp.
- Wetmore, A. 1933. Bird remains from the Oligocene deposits of Torrington, Wyoming. Bulletin of the Museum of Comparative Zoology 75:297–311.
- Wetmore, A. 1944. A new terrestrial vulture from the Upper Eocene deposits of Wyoming. Annals of Carnegie Museum 30:57–69.