
ABSTRACT
The Madtsoiidae are an extinct lineage of snakes known from the Late Cretaceous to the Late Pleistocene, with a rich fossil record distributed mainly across Gondwanan landmasses. However, only a few taxa are represented by cranial or articulated remains, and most madtsoiids are known only by isolated vertebrae. The unambiguous record of Madtsoiidae from the Cenozoic in South America had been restricted to the genus Madtsoia from Eocene and Oligocene deposits of Patagonia and Brazil. Here, we describe a new madtsoiid taxon, Powellophis andina gen. et sp. nov., based on an articulated postcranial skeleton from the Mealla Formation (middle–late Paleocene) in northwestern Argentina. The new taxon is estimated to be around 3 meters long, with a vertebral morphology sharing similar features with other mid-to-large forms. Its inclusion in a recent analysis of madtsoiid relationships recovers Powellophis as an early member of a clade formed by mostly large bodied and gigantic taxa. Its presence in the Paleocene of northwestern Argentina fills the gap between the diverse Late Cretaceous and Eocene–Oligocene records of madtsoiids in South America, confirms their presence in northern Gondwana by the early Paleogene, and expands the diversity of the group.
INTRODUCTION
Madtsoiidae are an extinct group of snakes known from the Late Cretaceous to the Late Pleistocene (LaDuke et al., Citation2010; Rage et al., Citation2014). Their monophyly and placement among snakes are highly debated, but many recent phylogenetic analyses have retrieved madtsoiids as an early diverging lineage of snakes outside crown group Serpentes, indicating their importance for understanding the early evolution of the group (e.g., Caldwell et al., Citation2015; Garberoglio et al., Citation2019a, Citation2019b; Gómez et al., Citation2019; Fachini et al., Citation2020). Alternatively, other recent hypotheses have placed madtsoiids inside crown group Serpentes, either as an early radiation of non-alethinophidian snakes (e.g., Longrich et al., Citation2012; Hsiang et al., Citation2015; Harrington and Reeder, Citation2017; Zaher et al., Citation2022) or more deeply nested within Alethinophidia (e.g., Wilson et al., Citation2010; Zaher and Scanferla, Citation2012).
The combination of vertebral features commonly considered as diagnostic of Madtsoiidae include: presence of parazygantral foramina; lack of prezygapophyseal processes; presence of paracotylar foramina; interdiapophyseal width approaching or exceeding the width between prezygapophyses (Rage, Citation1998; Scanlon, Citation2005). At least 24 species in 15 genera have been described as belonging to this clade, with a widespread but mainly Gondwanan distribution (LaDuke et al., Citation2010; Rage et al., Citation2014; Rio and Mannion, Citation2017; Gómez et al., Citation2019). They are recorded after the Eocene only in Australia, Argentina, and India (Scanlon, Citation2004; Albino and Brizuela, Citation2014; Wasim et al., Citation2022).
The Late Cretaceous South American record of madtsoiids is known mostly from isolated vertebrae, and includes a rich diversity of species of different size classes from Patagonia. Taxa considered to be small-sized, i.e., less than 2 meters in estimated length, include Alamitophis argentinus, Alamitophis elongatus, and Patagoniophis parvus (Albino, Citation1986, Citation1994, Citation2000, Citation2007). Rionegrophis madtsoioides (Albino, Citation1986, Citation2007), with an estimated length between 2 and 4 meters, represents a mid-sized form, and Eomadtsoia ragei, with an estimated length surpassing 4 meters is considered a large-sized form (Gómez et al., Citation2019). In contrast, South American Cenozoic records are less diverse and only represented by the mid-to-large form Madtsoia (Simpson, Citation1933; Albino, Citation1993; Albino and Brizuela, Citation2014; Rage, Citation1998).
Madtsoiids are tentatively known from Paleocene deposits in South America from a possible specimen from Bolivia (Rage, Citation1991), but are well represented from Eocene deposits, with two species being currently described: Madtsoia bai Simpson, Citation1933 from Patagonia, Argentina and Madtsoia camposi Rage, Citation1998 from Brazil. A vertebral centrum tentatively referred to the genus Madtsoia from the Oligocene of Patagonia represents the youngest record of madtsoiids outside Australia and India (Albino and Brizuela, Citation2014; Wasim et al., Citation2022).
Here, we describe a new Paleocene aged, mid-sized madtsoiid snake from the Mealla Formation of Jujuy Province, northwestern Argentina. This is based on an articulated postcranial skeleton. We include this new taxon in a recent comprehensive phylogenetic study of madtsoiid snakes and discuss the implications of this new record, which expands the diversity of madtsoiids from the poorly sampled Paleocene of South America and represents the first madtsoiid from a non-Patagonian site in Argentina.
GEOLOGICAL SETTING
The Mealla Formation corresponds to the inferior unit of the Santa Bárbara Subgroup (Salta Group) which is a Paleogene fluvial and lacustrine sequence typical of NW Argentina. Based on the first record of henricosborniid mammals, the outcrops of the Mealla Formation SSW from the Tres Cruces sub-basin were correlated with the Río Chico Formation of Patagonia and assigned to the Riochican South American Land Mammal Age (SALMA) (Pascual et al., Citation1978; Quattrocchio et al., Citation1997), which currently is considered early Eocene in age (Gelfo et al., Citation2009). Meanwhile, Marshall et al. (Citation1997) correlated this unit with the earlier Peligran SALMA (Selandian, mid-Paleocene), although this cannot be supported on its mammal content (Gelfo et al., Citation2009). Later, Woodburne et al. (Citation2014) assigned the Mealla Formation to the Tanethian (late Paleocene) and Hyland et al. (Citation2015) calculated an age around 59 Ma (Selandian–Tanethian) based on magnetostratigraphic methods, also supporting a late Paleocene age for this formation. Nevertheless, Zimicz et al. (Citation2020) questioned the age model of Hyland et al. (Citation2015) and the previously proposed correlations based on the mammalian content, concluding that the age of the Mealla Fm. should extend from the late Danian to the Selandian (early–middle Paleocene). Recently, del Papa et al. (Citation2022) provided for the first time an absolute date for the upper levels of the Mealla Fm., giving a U–Pb zircon age of 58 Ma. Combined with previously estimated ages, along sedimentological and stratigraphic approaches, del Papa et al. (Citation2022) propose a temporal range of 61–57 Ma for the Mealla Fm.
The Mealla Formation is 250 m thick and characterized by medium to fine grained sandstones intercalated with fine grained mudstone deposits, which are interpreted as fluvial, floodplain, and lacustrine facies (del Papa et al., Citation2022). The environmental interpretation indicates the presence of a sandy low energy and low sinuosity meandering fluvial system, mud dominated floodplains with paleosol development, and shallow, freshwater clastic lakes with periods of subaerial exposure (del Papa et al., Citation2022).
The vertebrate fauna from the Mealla Fm. found around the locality of Mina Aguilar includes teleostean fishes, pelomedusid turtles, a sebecid crocodile, and notoungulate mammals (Pascual et al., Citation1978; Zimicz et al., Citation2020; Bravo et al., Citation2021).
MATERIALS AND METHODS
Materials
The fossil specimen described in this contribution was found and collected by Dr. Jaime Powell (†) in sediments corresponding to the Mealla Formation near Mina Aguilar, Jujuy Province, northwestern Argentina (). This material is deposited in the Vertebrate Paleontological Collection of the Instituto Miguel Lillo (PVL) in San Miguel de Tucumán, Argentina. The original specimen, PVL 4714, consisted of an articulated skeleton with crania and postcrania (precloacal vertebrae and ribs) preserved within the same block of sediment and corresponding to a single individual. After its extraction from the field, a cast was made of the block containing PVL 4714 before being sectioned into nine successive slabs during preparation. The skull and vertebral column of the specimen were dorsoventrally oriented relative to the bedding plane in the first five slabs, and on the first string of vertebrae of slab 6, whereas vertebrae from the second string of slab 6 and those on slabs 7, 8, and 9 were disposed in a lateral orientation relative to the bedding plane. The first slab (PVL 4714-1), containing the skull articulated with the atlas, axis, and 14 anteriormost vertebrae, as well as several vertebrae that were separated from other slabs, are not available in the PVL collection and are considered lost. Thus, the material described here includes eight slabs containing part of the postcranial skeleton of a single individual (PVL 4714-2 to PVL 4714-9), and the cast made before the loss of several elements was used for reference. All the slabs containing the specimen were prepared both dorsally and ventrally, and four vertebrae were removed and prepared separately to reveal the whole morphology.
FIGURE 1. Detail of the study area (square in map of Argentina) and location of Mina Aguilar (star).
The fossil is articulated and relatively well preserved, although it shows breakage and weathering, particularly on distal surfaces such as the neural spines and hypapophyses. The bones were covered by a thin layer of gypsum (pers. observ., LNT). Photographs of the specimen were taken with a Nikon D90 digital camera and a Dino-Lite Digital Microscope.
Measurements and Nomenclature
Measurements in were taken with a manual digital caliper (0.01 mm error), and followed those defined by LaDuke (Citation1991), Albino (Citation2011a), and subsequently used by other authors (e.g., Vasile et al., Citation2013; Rio and Mannion, Citation2017; Gómez et al., Citation2019). Osteological nomenclature followed mainly Rage (Citation1984, Citation1998, Citation2001), Rage and Albino (Citation1989), and Albino (Citation2011a).
TABLE 1. Vertebral measurements (see Fig. S11) in mm of selected vertebrae from PVL 4714. Abbreviations: APV, anterior precloacal vertebra; CL, centrum length; CNW, condyle width; COW; cotyle width; DW, width across diapophyses; MPV, middle precloacal vertebrae; NAW, neural arch width; NSH, neural spine height; POW, width across postzygapophyses; PR-PO, length from the anterior edge of the prezygapophyseal facet to the posterior edge of the postzygapophyseal facet; PRW, width across prezygapophyses; TVH, total vertebral height; ZSH, zygosphene height; ZSW, zygosphene width.
Phylogenetic Analyses
In order to assess the position of the new taxon among snakes and its affinities with Madtsoiidae, it was included in the data matrix from Gómez et al. (Citation2019), with the addition of a few scoring changes and a new character: the absence/presence of distinct fossae on the dorsolateral surface of the neural arch (Supplemental Data). The character scorings of the new taxon were based only on anatomical information observed on the available blocks. The resulting matrix of 265 characters and 55 taxa was analyzed by maximum parsimony in TNT v.1.5-beta (Goloboff and Catalano, Citation2016), following the same procedures as in Gómez et al. (Citation2019) for tree search. The analysis was performed under equal weight and consisted of 500 tree replicates obtained by random addition sequence (RAS), searching for new tree topologies with tree bisection and reconnection (TBR), saving 100 trees per replication and collapsing branches of zero length after tree search. A final round of TBR branch swapping was performed to the trees obtained, and the outgroup Varanus was used for rooting the optimal topologies. Most characters were treated as unordered during tree search, except a few multi-state characters based on continuous data (Supplemental Data), which were treated as ordered following Gómez et al. (Citation2019). Node support was calculated by the Bremer index and under absolute frequencies by jackknifing (p = 0.36, 1000 pseudo-replicates).
Body Length Estimation
The total body length of the new taxon was estimated quantitatively, based on a previous sample of vertebral measures and measures of total length taken from extant snakes (McCartney et al., Citation2018). Total length was regressed on two different vertebral measures using a reduced major axis (RMA) regression with a forced zero intercept in the statistical package PAST (Hammer et al., Citation2001). Then, the slopes (130.04, 417.93) of each of the regressions (r2 = 0.95026, r2 = 0.90843; P [corrected] = 0.0001) were used to estimate the total length of the new taxon based on both condylar width and prezygapophyseal width (, Figs. S1, S2).
Institutional Abbreviations—PVL, Paleontología de Vertebrados, Instituto “Miguel Lillo”, San Miguel de Tucumán, Argentina.
SYSTEMATIC PALEONTOLOGY
SQUAMATA Oppel, Citation1811
OPHIDIA Brongniart, Citation1800
MADTSOIIDAE (Hoffstetter, Citation1961) McDowell, Citation1987
POWELLOPHIS ANDINA gen. et sp. nov.
(, S3–S9)
FIGURE 2. Powellophis andina gen. et sp. nov. (PVL 4714). A, cast of the original specimen as collected from the field; B, correspondence of each of the sectioned slabs after preparation.
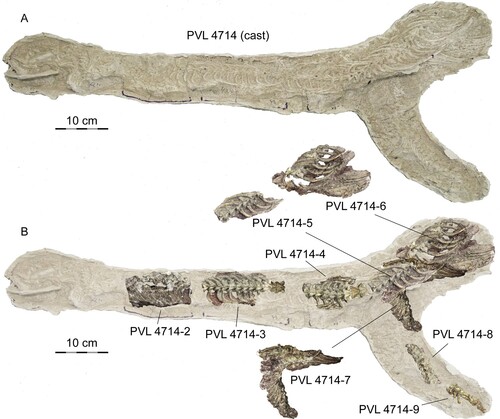
FIGURE 3. PVL 4714-3, anterior precloacal vertebrae (38th and 39th) and interpretative drawing. A, anterior view; B, posterior view; C, left lateral view; D, dorsal view, E, ventral view. Abbreviations: con, condyle; cot, cotyle; hp, hypophysis; izc, interzygapophyseal constriction; izr, interzygapophyseal ridge; lc, laminar crest; lf, lateral foramen; naf, neural arch fossa; nc, neural canal; ns, neural spine; pcf, paracotylar foramen; pzf, parazygantral foramen; prz, prezygapophysis; scf, subcentral foramen; scr, subcentral ridge; syn, synapophysis; zp, zygosphene; zt, zygantrum.
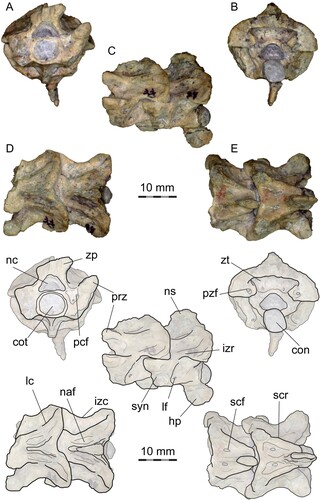
FIGURE 4. PVL 4714-4, middle precloacal vertebra (49th) and interpretative drawing. A, anterior view; B, posterior view; C, left lateral view; D, dorsal view, E, ventral view. Abbreviations: con, condyle; cot, cotyle; hk, hemal keel; izc; interzygapophyseal constriction; izr, interzygapophyseal ridge; lc, laminar crest; lf, lateral foramen; naf, neural arch fossa; nc, neural canal; ns, neural spine; pcf, paracotylar foramen; pe, posterior embayment; pzf, parazygantral foramen; prz, prezygapophysis; ptz, postzygapophysis; scf, subcentral foramen; scr, subcentral ridge; syn, synapophysis; zp, zygosphene; zt, zygantrum.
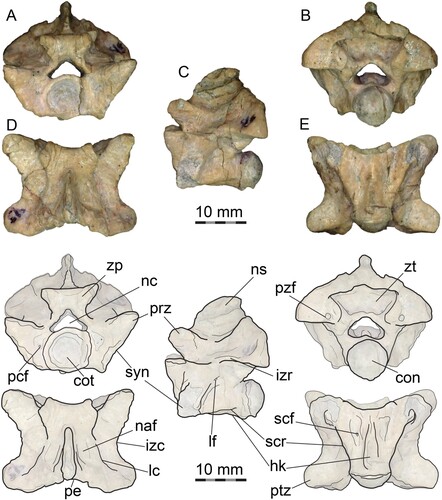
FIGURE 5. PVL 4714-4, middle precloacal vertebra (50th) and articulated left rib fragment. A, anterior view; B, posterior view; C, right lateral view; D, dorsal view, E, ventral view. Abbreviations: r, rib fragment; zp, zygosphene from succeeding vertebra.

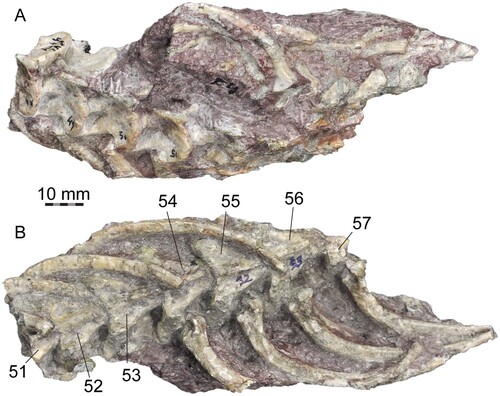
FIGURE 6. PVL 4714-5, middle precloacal vertebrae and ribs (51st to 57th). A, dorsal view. B, ventral view. Numbers indicate position in the column.
Etymology—The generic name comes from Powell, in honor of Dr. Jaime Powell (†), collector of the studied specimen, and in recognition of his contribution to the vertebrate paleontology of Argentina, and ophis, Greek for “snake." The specific epithet andina is after the Andes region in northwestern Argentina, where the fossil was found.
Holotype—PVL 4714, a nearly complete skeleton preserved articulated in nine continuous siltstone blocks and a cast of the original specimen before preparation. The only available blocks for study were PVL 4714-2 to PVL 4714-9.
Type Locality and Horizon—Surroundings of Mina Aguilar (3,980 m asl, 23°12′59″S, 65°40′49″W), 29 kilometers south to the village of Tres Cruces, Humahuaca Department, Jujuy Province, northwestern Argentina. Mealla Formation (middle–late Paleocene).
Diagnosis—Mid-sized madtsoiid snake (estimated body length around 3 meters) characterized by a unique combination of features: wide and short vertebrae (differs from Alamitophis, Nanowana, Nidophis, and Patagoniophis); moderately constricted vertebrae (differs from Alamitophis, Gigantophis, Nidophis, and Patagoniophis); a single large parazygantral foramen on each side of the zygantrum; paracotylar foramina and paracotylar fossae on each side of the cotyle; absence of prezygapophyseal processes; prezygapophyses with their main axis anterolaterally oriented (differs from Alamitophis tingamarra, Eomadtsoia, Rionegrophis, Sanajeh, Wonambi, Yurlunggur, and all species of Madtsoia except Madtsoia camposi); tall and only moderately vaulted neural arch in middle precloacal vertebrae (distinct from all mid-to-large madtsoiids except Madtsoia camposi), with demarcated dorsolateral ridges (differing from Adinophis and Eomadtsoia); absence of arqual ridges (Scanferla and Canale, Citation2007) (differs with most madtsoiids, except Menarana laurasiae); shallow depressions on the dorsal surface of the neural arch, between neural spine and dorsolateral ridges (differs with most mid-to-large madtsoiids except Sanajeh, Menarana, Rionegrophis, Madtsoia pisdurensis, and Madtsoia madagascariensis); moderately high neural spine (distinct from madtsoiids with low neural spines, such as Adinophis, Eomadtsoia, and Yurlunggur); posterior margin of neural arch shallowly concave in dorsal view (differs from most mid-to-large madtsoiids, except Eomadtsoia and Menarana nosymena); thick zygosphene with deeply concave anterior margin in middle precloacal vertebrae (differs from all madtsoiids except Madtsoia bai, Madtsoia madagascariensis, and Menarana nosymena); broad subtriangular centrum in ventral view; anterior precloacal vertebrae with posteriorly oriented, paddle-shaped (wide and laterally compressed) hypapophyses; middle precloacal vertebrae with well-demarcated and narrow hemal keel (differs from most mid-to-large madtsoiids, except Eomadtsoia and Rionegrophis); lateral edge of the diapophyses medial to lateral edge of the prezygapophyses (differs from all madtsoiids except Eomadtsoia). The following characters and characters states are recovered as autapomorphies of Powellophis andina in the phylogenetic analysis: 231:0; 238:0; and 244:0.
DESCRIPTION
The total length of the preserved parts of the specimen is approximately 1290 mm (). The head length is best assessed by the length of the well-preserved right mandibular ramus, which measures 95 mm. Some skull elements preserved in the lost block PVL 4714-1 are visible in the cast (, S3) and include the complete right lower mandible exposed in ventral view, right maxilla and left maxilla in ventral view, the broken right pterygoid in ventral view, the left dentary in lateral view, and a toothed fragment probably corresponding to the left palatine, along other unidentified elements. The dentary is robust and with an apparently almost straight medial margin. Visible teeth on the left dentary and right maxilla are needle-shaped, elongate and distinctly recurved. The compound bone bears a well-developed retroarticular process. The transverse (lateral) process of the pterygoid is present, but the degree of lateral expansion is not clear. The quadrate ramus of the pterygoid is markedly curved and with a distinct longitudinal groove. We do not attempt to describe in detail the skull elements based on the cast, since the proper shape and condition of the bones are not entirely visible, and sutural contacts between elements forming the right lower mandible are not discernible.
Along with the skull, the first slab contained 16 vertebrae: the atlas, the axis, and 14 anterior precloacals (J. Powell, pers. comm. to AA). The material described here includes the postcranial skeleton preserved in the eight slabs numbered as PVL 4714-2 to 4714-9: PVL 4714-2 (Fig. S4), contained a string of 10 anterior precloacal vertebrae continuous to the first slab (17th–26th), but only nine are preserved because the third vertebrae in the section (19th) is missing; PVL 4174-3 (, S5), separated from the previous slab by one missing vertebrae (27th), bears an articulated string of 12 anterior precloacals (28th to 39th), from which the last two were separated and prepared (); PVL 4714-4 (, , S6), separated by two missing vertebrae (40th and 41st) from PVL 4714-3, contains nine anterior to middle precloacal vertebrae (42nd to 50th), from which the last two vertebrae were each one separated and prepared (, ); PVL 4714-5 (), continuous to PVL 4714-4, contains seven articulated middle precloacal vertebrae (51st to 57th); PVL 4714-6 (Fig. S7), a block containing the posterior half of the last vertebra from PVL 4714-5 (57th) and other 12 middle precloacal vertebrae in two different sections that point to opposite directions: the first section includes a string of six consecutive vertebrae (59th–64th), and the second section contains a string of six articulated middle precloacal vertebrae; the position of these sections in the column is unknown because there is a discontinuity of missing vertebrae that were already eroded when the specimen was collected; PVL 4714-7 (Fig. S8), comprises at least 18 articulated middle to posterior precloacal vertebrae; PVL 4714-8 (Fig. S9), a string of 10 articulated posterior precloacal vertebrae; and PVL 4714-9 (Fig. S9), seven articulated posterior precloacal vertebrae.
Originally, the vertebral column comprised a total of 105 preserved vertebrae, of which 21 are now lost and only present in the cast. The total number was likely much higher, as the specimen lacks a portion of the mid-trunk region located between the two strings of precloacal vertebrae preserved on PVL 4714-6 and both cloacal and caudal regions, as the last slab bears only precloacals. Thus, the specimen mainly consists of two series of vertebrae interrupted by a hiatus on slab 6. The first string of vertebrae contains the skull, atlas, axis, and anteriormost 64 precloacal vertebrae (including slabs 1 to 5, and the first string of slab 6), whereas the second series retains 41 precloacal vertebrae (including the second string of vertebrae of slab 6 and slabs 7, 8, and 9).
General Vertebral Features
Within the nine articulated vertebrae preserved in PVL 4714-4, the first four are badly preserved on their ventral surface, but the fifth vertebra (46th) shows the presence of a well-defined hypapophysis (Fig. S6). The sixth (47th) is not easily determined but seems to have the first hemal keel. The last three vertebrae in PVL 714-4 (48th–50th) show well-developed hemal keels across their centra and are identified as among the anteriormost middle precloacals (Fig. S6). The last two vertebrae preserved in PVL 4714-4 (49th and 50th) were separated from the block and prepared from all sides to serve as the primary elements to base the description of the new taxon, and will be first described in detail, whereas the remaining sections will be used to assess the intracolumnar variation.
The first one of these two (49th) is nearly complete (), whereas the second one (50th), which is articulated with a fragment of the corresponding left rib and the zygosphene of the succeeding vertebra, lacks the zygosphene and condyle (), although the shape of the synapophyses is better preserved than in the preceding one.
Anterior View—The cotyle is big relative to the total width of the vertebra and circular. One or two small paracotylar foramina are present at each side of the cotyle, located in shallow fossae (). The zygosphene is robust, slightly wider than the cotyle and relatively thick, with a low width to height ratio (). The zygosphene articular surfaces are moderately inclined, around 30° from the vertical (sagittal) plane, and the zygosphene dorsal margin is slightly concave. The neural canal is low, with distinct arterial grooves, trifoliate in cross section and relatively small (about half the area of the cotyle). The neural arch laminae are relatively low anteriorly and high posteriorly, rising sharply (). The prezygapophyses are well projected laterally and moderately inclined dorsally, between 20 and 30° from horizontal. Prezygapophyseal processes are absent. The synapophyses are badly preserved, but they are clearly differentiated into distinct parapophyses and diapophyses (, ). The lateral edge of the diapophyses is only slightly projected, remaining medial to the lateral edge of the prezygapohyses. The ventral edge of the parapophyses projects ventrally beyond the ventral margin of the cotyle.
Lateral View—The vertebrae are higher than long (, ). The neural spine is well-developed and moderately high (representing less than 30% of the total height of the vertebrae, although the dorsal margin is weathered), extending from the base of the zygosphene to the posterior edge of the neural arch (). The zygosphenal articular surfaces are oval in shape. Posterior to the zygosphene, shallow fossae are present on the dorsolateral surface of the neural arch, to each side lateral to the neural spine (). The interzygapophyseal ridge is prominent, short and sinuous. One small lateral foramen is present, below the interzygapophyseal ridge and positioned close to the posterior edge of the synapophyses (). The synapophyses are prominent and anteroventrally elongated. The condyle is posterodorsally oriented and well demarcated from the centrum by a marked and deep precondylar constriction (). The subcentral ridge is almost straight. The hemal keel is well developed, ventrally prominent.
Posterior View—The neural arch is wide and slightly vaulted. The condyle is circular. The zygantrum is spacious and deep, with large zygantral facets. A small zygantral foramen is located on each side, close to the ventral edge of the zygantral cavity, in rounded shallow fossae (). The zygantral roof is thick and almost straight on its ventral margin. A single large parazygantral foramen is present on each side of the zygantrum, ventromedially positioned, close to the postzygapophyseal facet (, ). The postzygapophyses are large and robust, and the articular facets are almost straight, not inclined from the horizontal plane.
Dorsal View—The vertebrae are wider than long (, ). The neural arch is constricted but not strongly marked, as the prezygapophyseal width is not significantly greater than the interzygapophyseal width (minimal neural arch width). The prezygapophyseal facets are oval in outline (although the margins are slightly eroded), and are anterolaterally projected (angle around 50° from the sagittal plane). The anterior margin of the zygosphene is deeply concave (). The neural spine is well-developed, extending from the base of the zygosphene to the posterior border of the neural arch, where it is broader than anteriorly and overhangs the posterior neural arch notch to a small degree (). To each side of the neural spine, a shallow fossa is present on the dorsolateral surface of the neural arch, as a longitudinal depression delimited by faintly marked ridges (, ). These structures have been described in other madtsoiids, named as parazygosphenal fossae (e.g., LaDuke et al., Citation2010). There are no foramina located on the fossae. Lateral to these depressions, laminar crests or parasagittal (= dorsolateral) ridges are present, but weakly developed on the neural arch, extending from the base of the zygosphene to the postzygapophyseal lappet. Arqual ridges—prominent crests on each side of the neural arch, allocated above the zygantral fossae and subparallel to the neural spine (Scanferla and Canale, Citation2007)—are absent. On its posterior margin, the neural arch is shallowly notched.
Ventral View—The centrum is short and subtriangular, much broader anteriorly than posteriorly (, ). Subcentral ridges are prominent and almost straight, extending from the base of the parapophyses to the condylar constriction (). A well-defined, narrow hemal keel is developed along most of the ventral surface of the centrum, reaching the precondylar constriction. Although it is not completely preserved, the hemal keel is lower anteriorly, but rises and broadens posteriorly (). The posterior end of the hemal keel is simple, lacking a conspicuous structure such as tubercles or posterior processes. Lateral to the hemal keel, shallow depressions, bearing small subcentral foramina, are present anteriorly. The precondylar constriction is well marked and separates the hemal keel from the condyle, which is well exposed ventrally (). The postzygapophyses are broad and laterally oriented, with oval articulatory facets and an almost straight posterior margin.
Intracolumnar Variation
Anterior Precloacal Vertebrae—The anterior precloacals, identified by the presence of hypapophyses, are present in PVL 4714-2 (10 vertebrae), PVL 4714-3 (12 vertebrae), and PVL 4714-4 (which contains the transition from the anterior to the middle precloacal region of the column, with at least five anterior precloacal vertebrae). Thus, at least 27 anterior precloacals are preserved in PVL 4714. Based on the cast of the original specimen (), one vertebra is missing between PVL 4714-2 and PVL 4714-3, and two more vertebrae are missing between PVL 4714-3 and PVL 4714-4, indicating a minimum of 30 anterior precloacals in the available material. Additionally, 16 vertebrae were present in the first slab with the skull, indicating the presence of at least 46 anterior precloacal vertebrae with hypapophyses (‘cervical vertebrae’) in Powellophis.
The last two vertebrae from PVL 4714-3 (38th and 39th), articulated to each other and corresponding to anterior precloacals (Fig. S5), were separated and prepared from all sides (). The second one (39th) is mostly complete, but the first vertebra (38th) lacks the right prezygapophysis and right zygosphene, whereas the hypapophysis and the neural spine are both broken. These two vertebrae are slightly smaller than the middle precloacals described above, but very similar in proportions, as expected since they are located close to the transition from anterior to middle precloacals. The neural canal is slightly larger, relative to the size of the cotyle (), in comparison to the middle precloacals. Paracotylar fossae and paracotylar foramina are more conspicuous and deeper. Prezygapophyses are less projected laterally and more steeply inclined, around 30° from the horizontal plane (). The fossae on the dorsolateral surface of the neural arch, lateral to the neural spine, are more conspicuous and deeper, and delimited by marked ridges extending posteriorly from the back of the zygosphene; these ridges are distinct from the laminar crest and positioned medially to the latter (). The synapophyses are less ventrally projected, but the parapophyses still extend ventrally beyond the ventral margin of the cotyle. On the left side, the lateral foramina are larger and more ventrally positioned, above the subcentral ridges and close to the distal margin of the synapophyses (). On the right side, the lateral foramen is placed in a central position (). The subcentral ridge is more prominent than in the middle precloacals. The neural spine is squared in lateral view, and around the same height as in the middle precloacals, lacking a posterior tubercle (). The neural arch is higher and more vaulted (). The medial depressions on the ventral surface of the centrum are deeper and the subcentral foramina are more conspicuously marked. The most notable difference with the middle precloacals is the presence of a well-developed and robust hypapophysis (), instead of the hemal keel. The hypapophysis is elongate (as long as the height of the centrum), wide anteroposteriorly and laterally compressed. It is a paddle-shaped process, with a rounded distal edge, posteroventrally directed from the posterior half of the centrum, and extending beyond the posterior margin of the respective condyle (). The remaining anterior precloacal vertebrae in PVL 4714-3 do not differ further from the two already described (Fig. S5), and the more anterior vertebrae from PVL 4714-2 are not preserved well enough to be properly described (Fig. S4).
Middle Precloacal Vertebrae—The middle precloacal region is preserved in PVL 4714-4 (by the last three or four vertebrae of the section), PVL 4714-5 (seven vertebrae), PVL 4714-6 (13 preserved vertebrae), and PVL 4714-7. PVL 4714-4, PVL 4714-5, and PVL 4714-6 are continuous to each other (). The first section of vertebrae from PVL 4714-6 is constituted of the posterior half of the last vertebrae in PVL 4714-5 and six articulated vertebrae that are separated from each other by only one missing (Fig. S7). The next section in PVL 4714-6 is another string of six vertebrae, but this section is separated by a discontinuity of an unknown number of elements. Nevertheless, the two strings of vertebrae separated from this discontinuity are not markedly different in size and morphology. PVL 4714-7 is continuous to PVL 4714-6 and preserves at least 18 articulated vertebrae that decrease in size posteriorly (Fig. S8). The limit between middle precloacals and posterior precloacals in this section is not clear, since these vertebrae are obscured by sediment and not properly visible. Thus, somewhat around ∼40 middle precloacals can be counted in the available material, but the number was surely higher. The neural spines are high and squared (, S7, S8), not markedly inclined or canted, as the posterior margin is almost straight in lateral view. The neural spines lack posterior tubercles in all middle precloacals. The hemal keel is tall and narrow, well-demarcated in all middle precloacals (, S6, S7), but in more distal ones the hemal keel is lower, and as wide anteriorly as posteriorly (Fig. S7).
Posterior Precloacal Vertebrae—The posterior precloacal region is preserved in PVL 4714-7 (the limit between middle and posterior precloacals is not clear), PVL 4714-8 (nine vertebrae), and PVL 4714-9 (seven vertebrae). These sections are not continuous with each other () and there are no isolated and prepared vertebrae available from this region of the column. The morphology of these vertebrae varies mainly as expected for posterior precloacals. They are smaller in overall size, and have more depressed neural arches with less demarcated laminar or dorsolateral crests (Fig. S9). The dorsolateral surface of the neural arch lacks marked depressions or fossae lateral to the neural spine. The neural spine decreases in height posteriorly along the column and it is lower in these vertebrae than in the middle precloacals. The diapophyses are more laterally projected, and their ventral margin is positioned below the level of the neural canal floor. The parapophyses are more ventrally projected. The hemal keel is broad and delimited by subcentral paralymphatic fossae, present as deep depressions separating the centrum anteriorly from the parapophyses (Fig. S9).
Ribs
At least 110 ribs are preserved and articulated with their respective vertebra (, , S4–S9). The ribs of Powellophis are robust compared with the overall size of their corresponding vertebrae and relatively long. In the middle trunk region (, S7), the ribs are evenly curved whereas in the anterior trunk region are bowed, markedly curved (Fig. S5). The proximal head is divided in two distinct facets, a dorsal diapophyseal facet and a ventral parapophyseal facet (). The diapophyseal facet is large and concave, the parapophyseal facet is flat to slightly convex, and broader than the diapophyseal facet. A marked notch and a low ridge separate both facets from one another. Dorsally, a low and blunt tuberculiform process (tuber costae) is present (). The ventral margin is more anteriorly projected than the dorsal margin and a well-developed ventral process is present, rounded to subtriangular in shape ().
DISCUSSION
Taxonomy and Anatomical Comparisons
The new articulated postcranial skeleton from the Mealla Formation described above shows a unique combination of features that permits the erection of a new snake taxon, Powellophis andina gen. et sp. nov. The new species is assigned to Madtsoiidae based on the general configuration of the vertebrae, along with the presence of distinct parazygantral and paracotylar foramina, and the absence of prezygapophyseal processes (Rage, Citation1998; Scanlon, Citation2005), which clearly distinguish it from known extinct and extant boas and pythons (Constrictores, sensu Georgalis and Smith, Citation2020) that have a similar overall vertebral morphology.
Among Madtsoiidae, small Late Cretaceous and Paleogene taxa such as Adinophis, Alamitophis, Australophis, Patagoniophis, Herensugea, Nanowana, and Nidophis (Albino, Citation1986, Citation1994, Citation2000, Citation2007; Rage, Citation1996; Scanlon, Citation1997, Citation2005; Vasile et al., Citation2013; Pritchard et al., Citation2014), are markedly smaller, with an estimated body length of less than 2 meters, and most of them have more elongate vertebrae. In contrast, the wide and short, large-sized vertebrae of Powellophis resembles medium-sized taxa such as Rionegrophis, Madtsoia camposi, Menarana nosymena, and Sanajeh (Albino, Citation1986, Citation2007; Rage, Citation1998; LaDuke et al., Citation2010; Wilson et al., Citation2010), which are estimated between 2 and 4 meters in length, and large-sized to gigantic taxa that exceed 4 meters in length such as Gigantophis, Wonambi naracoortensis, Menarana laurasiae, Yurlunggur, Eomadtsoia, Platyspondylophis, and remaining species of Madtsoia (Simpson, Citation1933; Smith, Citation1976; Scanlon, Citation1992; Rage, Citation1996; Scanlon and Lee, Citation2000; LaDuke et al., Citation2010; Mohabey et al., Citation2011; Rio and Mannion, Citation2017; Smith et al., Citation2016; Gómez et al., Citation2019). The narrow and well-demarcated hemal keel of Powellophis and the lack of diapophyses extending laterally beyond the limit of the prezygapophyses, clearly distinguishes it from those taxa, except the older Eomadtsoia (Late Cretaceous of Patagonia). It differs clearly from the latter by the more constricted neural arch, higher neural spine, developed parasagittal ridges/laminar crests on the neural arch, absence of arqual ridges, a dorsally concave zygosphenal roof, a more deeply concave anterior margin of the zygosphene, and neural arch with distinct fossae on its dorsolateral surface, lacking foramina. The only madtsoiids previously described from the Paleogene of South America belong to the genus Madtsoia: M. bai from Patagonia and M. camposi from Brazil (Simpson Citation1933; Rage Citation1998), both from deposits currently assigned to the Eocene (Gelfo et al., Citation2009). Besides the features mentioned above that distinguish the new taxon from Madtsoia and other mid- to large-size madtsoiids, Powellophis is notably smaller than M. bai and is differentiated further from both South American species assigned to this genus by the following characters: a markedly less vaulted neural arch (strongly vaulted in M. bai), neural arch lacking foramina (present in M. camposi), neural arch only shallowly notched posteriorly (strongly notched in M. bai and M. camposi), zygosphene wider than the cotyle (narrower than the cotyle in M. bai), concave anterior margin of the zygosphene (straight anterior margin in M. camposi), more steeply inclined prezygapophyses (inclined less than 15° in M. bai), prezygapophyses anterolaterally projected (laterally everted in M. bai), parapophyses ventral margin below ventral margin of the cotyle (clearly above ventral margin of the cotyle in M. bai), relatively lower neural spine (higher neural spine in M. bai), and longer hypapophyses on anterior precloacals (shorter in M. bai).
The continuous anterior section of the column shows that hypapophyses were present in the first 46 vertebrae in Powellophis. The presence of hypapophyses is restricted only to the anterior precloacal region in madtsoiids, but the total distribution is not known because most taxa are known from incomplete specimens or isolated vertebrae. In modern snakes where hypapophyses are restricted to the anterior region, the total number ranges from only 4–6 in scolecophidians and up to 40–70 in most snakes (List, Citation1966; Hoffstetter and Gasc, Citation1969; Scanlon, Citation2004). Among fossil snakes, 45 vertebrae with hypapophyses are present in Haasiophis (Palci et al., Citation2013), and at least 18 in Pachyrhachis (Lee and Caldwell, Citation1998). In Dinilysia and Najash, retrieved in the analysis closely related to taxa traditionally considered as madtsoiids, at least 6–10 vertebrae bearing hypapophysis are recorded, but the total number is unknown as the cervical region is incompletely known in both (Rage and Albino, Citation1989; Caldwell and Albino, Citation2002; Caldwell and Calvo, Citation2008; Garberoglio et al., Citation2019c).
The neural arch fossae seen in Powellophis (, )—distinct depressions on the dorsolateral surface of the neural arch located posterior to the zygosphene—are also present in other madtsoiids, especially medium- to large-size forms. These fossae were first described in Madtsoia madagascariensis and Menarana nosymena (and also probably in Alamitophis argentinus) by LaDuke et al. (Citation2010), as ‘parazygosphenal fossae’ that bear ‘parazygosphenal foramina.’ Distinct fossae on the neural arch were subsequently described in other medium- and large-sized madtsoiids, such as Sanajeh and Madtsoia pisdurensis (Wilson et al., Citation2010; Mohabey et al., Citation2011). Nevertheless, the typical parazygosphenal foramina present in other extant and fossil snakes, such as Acrochordidae, Colubridae, Lapparentophiidae, and Palaeophiidae (e.g., Rage, Citation1983; Head, Citation2005; McCartney and Seiffert, Citation2016; McCartney et al., Citation2018; Vullo, Citation2019) are located in a very different position, between the zygosphene and the prezygapophyses. The foramina on the dorsal surface of the neural arch in madtsoiids, when present, are located posterior to the zygosphene, either with or without the development of distinct fossae. Thus, these foramina seen in some madtsoiids are not considered homologous to the proper parazygosphenal foramina of other snakes.
Body Size and Paleoecology
Previous studies have estimated the body size of extinct snakes known from isolated vertebrae (e.g., Head et al., Citation2009; LaDuke et al., Citation2010; McCartney et al., Citation2018; Calvert et al., Citation2022). Here, based on the vertebral measures and the known total body length (including tail) from a sample of extant snakes studied by McCartney et al. (Citation2018), two different regression models were performed to estimate the body length of Powellophis based on measurements from middle precloacal vertebrae (45th). The estimated total body length ranges from 3378.4 mm using the prezygapophyseal width to 3205.5 mm using the condylar width. These estimations are consistent with a body size that could be qualitatively assessed using the preserved elements of PVL 4714, as the head length is estimated around 100 mm, and the precloacal postcranial skeleton preserved articulated is approximately 1290 mm. However, we do note that a significant portion of the middle precloacal region is not preserved, as well as part of the posterior precloacal region and the total of cloacal and tail regions and all estimations are thus tentative statements.
Without information on the cranial morphology, a robust interpretation of the ecological habit of the new taxon is limited, as the postcranial morphology of Powellophis resembles those of generalist terrestrial to arboreal and semi-aquatic extant snakes such as Boa or Eunectes. We do however confidently exclude extreme ecological habits such as those of fossorial or obligatorily aquatic snakes (Albino and Caldwell, Citation2003). The vertebrae of Powellophis, with a tall neural arch, well-developed hemal keel and a moderately high neural spine in the middle precloacal region, contrast with the small-sized vertebrae usually found in extant fossorial snakes, with a depressed neural arch, without hemal keel and with a low or absent neural spine in the middle precloacal region (Hoffstetter and Gasc, Citation1969). Also, the vertebrae from Powellophis are wide with high synapophyses that articulate with laterally directed ellipsoidal ribs, producing what we interpret as a cylindrical body shape that clearly differs from the laterally compressed body of obligatorily aquatic snakes. This latter group usually have narrow vertebrae and low synapophyses that articulate with ventrally directed ribs.
The palynomorph content and the paleoenvironmental reconstruction of the Mealla Formation as a fluvial plain environment with ephemeral lakes drained by meandering channels suggests a humid tropical to intertropical forest with seasonal dryness (Quatrocchio et al., Citation1997; Zimicz et al., Citation2020; del Papa et al., Citation2022). In this context, and taking together the body size (estimated around 3 meters) and the vertebral morphology of Powellophis, a terrestrial-semiaquatic-arboreal mode of life analog to extant boids is suggested.
Phylogeny
The phylogenetic analysis found six most parsimonious tress (MPTs) of 810 steps each (consistency index [CI] = 0.392; retention index [RI] = 0.688), of which the strict consensus is well resolved (, S10). Our analysis obtained the same basic topology previously obtained by Gómez et al. (Citation2019): most taxa traditionally considered as madtsoiids are recovered outside crown group Serpentes, within a monophyletic Madtsoiidae with the exclusion of Herensugea (). The lack of cranial information for most known madtsoiid taxa restricts confidence in the monophyly of the Madtsoiidae. Most broad-based studies that assess the phylogenetic relationships of the snake total group (stem- and crown) among squamates do recover a monophyletic Madtsoiidae, but only include as tip taxa those madtsoiids that include skull material: Sanajeh indicus, Wonambi, and Yurlunggur (e.g., Wilson et al., Citation2010; Longrich et al., Citation2012; Zaher and Scanferla, Citation2012; Caldwell et al., Citation2015; Hsiang et al., Citation2015; Harrington and Reeder, Citation2017; Garberoglio et al., Citation2019b). However, even with such taxonomic restriction a monophyletic Madstoiidae is not uniformly accepted. For example, the analysis of Zaher et al. (Citation2022) recovered Sanajeh indicus as a stem snake but well away from the Australian madtsoiids Yurlunggur and Wonambi. Madstoiid monophyly was also not uniformly recovered in the Garberoglio et al. (Citation2019a) study, which analyzed fragmentary cranial material assigned to Madtsoia camposi, Menarana nosymena, and Nanowana (see Scanlon, Citation1997; Rage, Citation1998; LaDuke et al., Citation2010 for description of that cranial material).
FIGURE 7. Phylogenetic relationships of Powellophis andina gen. et sp. nov. Strict consensus of six most parsimonious trees of 810 steps each. Numbers above and below branches indicate Bremer support and jackknife absolute values, respectively. Crown group Serpentes is shown as a solid triangle, for complete tree see Fig. S10.
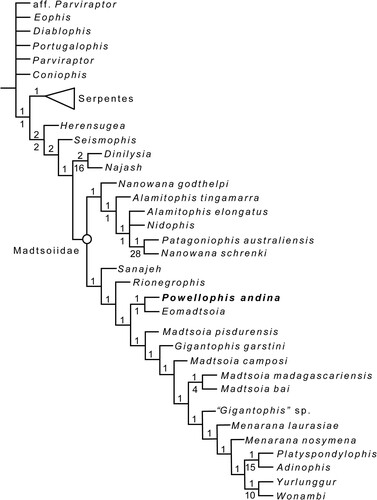
Similar to previous studies focusing on the ingroup relationships of Madtsoiidae (Vasile et al., Citation2013; Rio and Mannion, Citation2017; Gómez et al., Citation2019), two size-related clades of madtsoiids are retrieved here as well, with Powellophis as an early evolving form of the clade that includes mostly large-sized forms, as the sister taxon to Eomadtsoia, both crownward to the mid-sized forms Sanajeh and Rionegrophis (). In contrast with Gómez et al. (Citation2019), the clade that includes large-bodied madtsoiids is here fully resolved. Nevertheless, still none of the genera represented by more than one species is recovered as monophyletic, with only two of the four species assigned to the genus Madtsoia grouped together, M. bai and M. madagascariensis. The mid-sized vertebrae of Powellophis share some features commonly present in small taxa (such as a sharp hemal keel and anterolaterally projected prezygapophyses), that contrast with younger large to gigantic taxa, such as Madtsoia bai. The vertebral morphology of the new taxon links the derived morphology of younger gigantic madtsoiids from South America with the ancestral madtsoiid condition, according to the obtained topology. This fits well with the general trend of size increase around the Late Cretaceous previously observed for the clade of mostly large-sized madtsoiids (Gómez et al., Citation2019). We do not rule out the possibility that some vertebral features common to large-sized madtsoiids could be correlated with size, affecting the phylogenetic result. However, not all members of this clade are large-bodied, since mid-sized taxa such as Menarana nosymena and even the small-sized Adinophis are nested within, indicating the presence of vertebral characters supporting this group that are not size-correlated, and that independent trends towards gigantism and shifts to small size likely occurred within this lineage.
Paleobiogeographic Implications
Given their known temporal distribution in South America from the Late Cretaceous to the Oligocene (Albino and Brizuela, Citation2014), the presence of madtsoiids in Paleocene deposits was expected, but had remained unrecorded. The previous record of Madtsoiidae from the Paleocene was restricted to a fragmentary vertebra of a possible madtsoiid from the Tiupampan Area of Bolivia (Rage, Citation1991). This was included within a rich squamate assemblage with a diverse snake fauna that includes an anilioiid, boiids, a tropidophiid, and the derived macrostomatan Kataria anisodonta (Rage, Citation1991; Scanferla et al., Citation2013). In contrast, the poor Paleocene record of snakes and the absence of madtsoiids from other localities in South America likely indicates a sampling bias. The new taxon described here fills this previous gap between the diverse Late Cretaceous taxa from Patagonia and younger taxa from the Eocene and Oligocene of Patagonia and Brazil. Moreover, Powellophis is the first madtsoiid described from a non-Patagonian site in Argentina, indicating a wider distribution for madtsoiids in South America. It also indicates that they were present in northern South America by the early Paleogene, and likely even earlier, as the Late Cretaceous vertebrate assemblages from northwestern Argentina are still poorly known (e.g., Agnolin et al., Citation2022). The new taxon described here, and the Eocene macrostomatan Amaru scagliai (Albino, Citation2018), are together the only known Paleogene snake taxa from northwestern Argentina. Comparisons with the best known and more diverse Paleogene snake taxa from Patagonia, that include the genus Madtsoia and the boid genera Boa, Chubutophis, and Waincophis (Albino, Citation2011b) are still preliminary.
Powellophis expands the diversity of madtsoiids from the Paleogene of South America, previously known only from the genus Madtsoia, including the species Madtsoia bai (Simpson, Citation1933), Madtsoia camposi (Rage, Citation1998), and other material possibly referred to the same genus (Rage, Citation1991; Albino, Citation1993; Albino and Brizuela, Citation2014). All these forms are mid to large snakes, and small madtsoiids seem to be restricted to the Cretaceous in South America, although they still remained diverse after the K/Pg boundary in other landmasses such as Australia (Scanlon, Citation1997, Citation2005). This pattern could reflect the fact that only large-bodied madtsoiids survived into the Cenozoic in South America, occupying the niche of important predators while the climate was still tropical to subtropical. However, as the record of snakes outside Patagonia is still patchy, especially for the early Paleogene, future discoveries will determine if small-bodied madtsoiids persisted into the Cenozoic in South America or if they were replaced by crown group snakes (Serpentes).
Supplemental Material
Download Zip (18 MB)ACKNOWLEDGMENTS
AA especially thanks the paleontologist J. Powell (†), collector of the studied specimen, for the unconditional loan of the material and for his sincere friendship during more than three decades. Special thanks to the curators and personnel of the Instituto Miguel Lillo. We also thank J. Posik and L. Toledo from the Museo de la Plata for preparation of the specimen, M. Velasco for assisting with photographs, and F. Riguetti, R. O. Gómez, and M. W. Caldwell for their help. Thanks are extended to an anonymous reviewer, A. C. Pritchard and the editors P. L. Godoy and G. Bever who made constructive suggestions that greatly improved the quality of the manuscript. We thank G. Bever also for the help during the processing of the manuscript. We are also grateful to the Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) of Argentina for support. This work was partially funded by the grant PIP-CONICET 112-201501-00065 to AA.
REFERENCES
- Agnolín, F. D, M. A. Cerroni, A. Scanferla, A. Goswami, A. Paulina-Carabajal, T. Halliday, A. R. Cuffa, and S. Reuil. 2022. First definitive abelisaurid theropod from the Late Cretaceous of Northwestern Argentina, Journal of Vertebrate Paleontology 41: e2002348.
- Albino, A. M. 1986. Nuevos Boidae Madtsoiinae en el Cretácico tardío de Patagonia (Formación Los Alamitos, Río Negro, Argentina); pp. 15–21 in J. F. Bonaparte (ed.), Simposio Evolución de los Vertebrados Mesozoicos. Actas IV Congreso Argentino de Paleontología y Bioestratigrafía, Mendoza.
- Albino, A. M. 1993. Snakes from the Paleocene and Eocene of Patagonia, Argentina: paleoecology and coevolution with mammals. Historical Biology 7:51–69.
- Albino, A. M. 1994. Una nueva serpiente del Cretácico de Patagonia. Pesquisas 21:58–63.
- Albino, A. M. 2000. New record of snakes from the Cretaceous of Patagonia (Argentina). Geodiversitas 22:247–253.
- Albino, A. M. 2007. Lepidosauria; pp. 87–115 in Z. Gasparini, L. Salgado and R. A. Coria (eds.), Patagonian Mesozoic Reptiles. Indiana University Press, Bloomington & Indianapolis, Indiana.
- Albino, A. M. 2011a. Morfología vertebral de Boa constrictor (Serpentes: Boidae) y la validez del género mioceno Pseudoepicrates Auffenberg, 1923. Ameghiniana 48:53–62.
- Albino, A. M. 2011b. Evolution of Squamata Reptiles in Patagonia based on the fossil record. Biological Journal of the Linnean Society 103:441–457.
- Albino, A. M. 2018. New macrostomatan snake from the Paleogene of northwestern Argentina. Geobios 51:175–179.
- Albino, A. M., and S. Brizuela. 2014. First record of squamate reptiles from the Oligocene of South America. Alcheringa: An Australasian Journal of Palaeontology 38:412–421.
- Albino, A. M., and M. W. Caldwell. 2003. Hábitos de vida de la serpiente cretácica Dinilysia patagonica Woodward. Ameghiniana 40:407–414.
- Bravo, G.G., D. Pol, and D. A. García-López. 2021. A new sebecid mesoeucrocodylian from the Paleocene of northwestern Argentina. Journal of Vertebrate Paleontology 41:e1979020.
- Brongniart, A. 1800. Essai d’une classification naturelle des reptiles. Primere partie. Establissement des Orders. Bulletin des Sciences par la Société Philomatique, Paris 11:81–82.
- Caldwell, M. W., and A. Albino. 2002. Exceptionally preserved skeletons of the Cretaceous snake Dinilysia patagonica Woodward, 1901. Journal of Vertebrate Paleontology, 22:861–866.
- Caldwell, M. W., and J. Calvo. 2008. Details of a new skull and articulated cervical column of Dinilysia patagonica Woodward, 1901. Journal of Vertebrate Paleontology 28:349–362.
- Caldwell, M. W., R. L. Nydam, A. Palci, and S. Apesteguía. 2015. The oldest known snakes from the Middle Jurassic-Lower Cretaceous provide insights in snake evolution. Nature Communications 6:5996.
- Calvert, C. J., A. J. Mead, and D. Parmley. 2022. Size estimates of the extinct marine snake Pterosphenus schucherti from Eocene-aged sediments of Central Georgia. Georgia Journal of Science 80:Article 1.
- del Papa, C., J. Babot, J. Dahlquist, D. G. López, V. Deraco, C. Herrera, S. Bertelli, G. W. Rougier, and N. P. Giannini. 2022. Toward a chronostratigraphy of the Paleocene-Eocene sedimentary record in northwestern Argentina. Journal of South American Earth Sciences 113:103677.
- Fachini, T. S., S. Onary, A. Palci, M. S. Y. Lee, M. Bronzati, and A. S. Hsiou. 2020. Cretaceous blind snake from Brazil fills major gap in snake evolution. Iscience 23:101834.
- Garberoglio, F. F., S. Apesteguía, T. R. Simões, A. Palci, R. O. Gómez, R. L. Nydam, H. C. E. Larsson, M. S. Y Lee, and M. W. Caldwell. 2019a. New skulls and skeletons of the Cretaceous legged snake Najash, and the evolution of the modern snake body plan. Science advances 5:eaax5833.
- Garberoglio, F. F., R. O. Gómez, S. Apesteguía, M. W. Caldwell, M. L. Sánchez, and G. Veiga. 2019b. A new specimen with skull and vertebrae of Najash rionegrina (Lepidosauria: Ophidia) from the early Late Cretaceous of Patagonia. Journal of Systematic Palaeontology 17:1533–1550.
- Garberoglio, F. F., R. O. Gómez, T. R. Simões, M. W. Caldwell, and S. Apesteguía, S. 2019c. The evolution of the axial skeleton intercentrum system in snakes revealed by new data from the Cretaceous snakes Dinilysia and Najash. Scientific reports 9(1):1–10.
- Gelfo, J. N., F. J. Goin, M. Woodburne, and C. Muizon. 2009. Biochronological relationships of the earliest South American Paleogene mammalian faunas. Palaeontology 52:251–269.
- Georgalis, G. L., and K. T. Smith. 2020. Constrictores Oppel, 1811–the available name for the taxonomic group uniting boas and pythons. Vertebrate Zoology 702:91–304.
- Goloboff, P. A., and S. A. Catalano. 2016. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 32:221–238.
- Gómez, R. O., F. F. Garberoglio, and G. W. Rougier. 2019. A new Late Cretaceous snake from Patagonia: Phylogeny and trends in body size evolution of madtsoiid snakes. Comptes Rendus Palevol 18:771–781.
- Hammer, Ø., D. A. Harper, and P. D. Ryan. 2001. PAST: Paleontological statistics software package for education and data analysis. Palaeontologia electronica 4(1): 4.
- Harrington, S. M., and T. W. Reeder. 2017. Phylogenetic inference and divergence dating of snakes using molecules, morphology and fossils: new insights into convergent evolution of feeding morphology and limb reduction. Biological Journal of the Linnean Society 121:379–394.
- Head, J. J. 2005. Snakes of the Siwalik Group (Miocene of Pakistan): systematics and relationship to environmental change. Palaeontologia Electronica 8.1.18A:1–33.
- Head, J. J., J. I. Bloch, A. K. Hastings, J. R. Bourque, E. A. Cadena, F. A. Herrera, D. P. Polly, and C. A. Jaramilo. 2009. Giant boid snake from the Paleocene neotropics reveals hotter past equatorial temperatures. Nature 457:715–718.
- Hoffstetter, R. 1961. Noveaux restes d’un serpent Boidé (Madtsoia madagascariensis nov. sp.) dans le Crétacé Supérieur de Madagascar. Bulletin du Muséum National d’Histoire naturelle, Série 2, Paris 33:152–160.
- Hoffstetter, R., and J.-P. Gasc. 1969. Vertebrae and ribs of modern reptiles; pp. 201–310 in C. Gans, A. d. A. Bellairs, and T. S. Parsons (eds.), Biology of the Reptilia, Volume 1, Morphology A. Academic Press, London.
- Hsiang, A. Y., D. J. Field, T. H. Webster, A. D. Behlke, M. B. Davis, R. A. Racicot, and J. A. Gauthier. 2015. The origin of snakes: revealing the ecology, behavior, and evolutionary history of early snakes using genomics, phenomics, and the fossil record. BMC evolutionary biology 15:1–22.
- Hyland, E., N. Sheldon, and J. Cotton. 2015. Terrestrial evidence for a two-stage mid-Paleocene biotic event. Palaeogeography Palaeoclimatology, Palaeoecology 417:371–378.
- LaDuke, T. C. 1991. The fossil snakes of Pit 91, Rancho La Brea, California. Contributions in Science, Natural History Museum of Los Angeles County 424:1–28.
- LaDuke, T. C., D. W. Krause, J. D. Scanlon, and N. J. Kley. 2010. A Late Cretaceous (Maastrichtian) snake assemblage from the Maevarano Formation, Mahajanga basin, Madagascar. Journal of Vertebrate Paleontology 30:109–138.
- Lee, M. S. Y., and M. W. Caldwell. 1998. Anatomy and relationships of Pachyrhachis problematicus, a primitive snake with hindlimbs. Philosophical Transactions of the Royal Society of London, Series B353:1521–1552.
- List, J. C. 1966. Comparative osteology of the snake families Typhlopidae and Leptotyphlopidae. Illinois Biological Monographs 36:1–112.
- Longrich, N. L., B.-A. S. Bhullar, and J. A. Gauthier. 2012. A transitional snake from the Late Cretaceous period of North America. Nature 488:205–208.
- Marshall, L.G., T. Sempere, and R. Butler. 1997. Chronostratigraphy of the mammal-bearing Paleocene of South America. Journal of South American Earth Sciences 10:49–70.
- McCartney, J. A., and E. R. Seiffert. 2016. A late Eocene snake fauna from the Fayum Depression, Egypt. Journal of Vertebrate Paleontology 36:e1029580.
- McCartney, J. A., E. M. Roberts, L. Tapanila, and M. A. O'Leary. 2018. Large palaeophiid and nigerophiid snakes from Paleogene Trans-Saharan Seaway deposits of Mali. Acta Palaeontologica Polonica 63:207–220.
- McDowell, S. B. 1987. Systematics; pp. 3–50 in R. A. Seigel, J. T. Collins, and S. S. Novak (eds.), Snakes: Ecology and Evolutionary History. MacGraw-Hill Publications, New York.
- Mohabey, D. M., J. J. Head, and J. A. Wilson. 2011. A new species of the snake Madtsoia from the Upper Cretaceous of India and its paleobiogeographic implications. Journal of Vertebrate Paleontology 31:588–595.
- Oppel, M. 1811. Die Ordungen, Familien, und Gattungen der Reptilien, als Prodrom einer Naturgeschichte derselben. Joseph Lindauer, Munich, 87 pp.
- Palci, A., M. W. Caldwell, and R. L. Nydam. 2013. Reevaluation of the anatomy of the Cenomanian (Upper Cretaceous) hind-limbed marine fossil snakes Pachyrhachis, Haasiophis, and Eupodophis. Journal of Vertebrate Paleontology 33:1328–1342.
- Pascual, R., M. R. Vucetich, and J. Fernández. 1978. Los primeros mamíferos (Notoungulata, Henricosborniidae) de la Formación Mealla (grupo Salta, subgrupo Santa Bárbara). Sus implicancias filogenéticas, taxonómicas y cronológicas. Ameghiniana 15:367–390.
- Pritchard, A. C., J. A. McCartney, D. W. Krause, and N. J. Kley. 2014. New snakes from the Upper Cretaceous (Maastrichtian) Maevarano Formation, Mahajanga Basin, Madagascar. Journal of Vertebrate Paleontology 34:1080–1093.
- Quattrocchio, M., W. Volkheimer, and C. E. del Papa. 1997. Palinology and paleoenvironment of the “Faja Gris” Mealla Formation (Salta group) at garabatal creek (NW Argentina). Palynology 21:231–247.
- Rage, J.-C. 1983. Palaeophis colossaeus nov. sp. (le plus grand Serpent connu?) de l’Eocène du Mali et le problème du genre chez les Palaeopheinae. Comptes Rendus des Séances de l’Académie des Sciences de Paris 296:1029–1032.
- Rage, J.-C. 1984. Serpentes; in Handbuch der Paläoherpetologie, Volume 11. Gustav Fischer Verlag, Stuttgart, Germany, 80 pp.
- Rage, J.-C. 1991. Squamate reptiles from the early Paleocene of the Tiupampa area (Santa Lucia Formation), Bolivia; pp. 503–508 in R. Suarez-Soruco (ed.), Fósiles y Facies de Bolivia—Volume 1 Vertebrados. Revista Técnica de YPFB 12.
- Rage, J.-C. 1996. Les Madtsoiidae (Reptilia, Serpentes) du Crétacé supérieur d’Europe: témoins gondwaniens d’une dispersion transtéthysienne. Comptes Rendus de l’Académie de Sciences, Série IIa 322:603–608.
- Rage, J.-C. 1998. Fossil snakes from the Palaeocene of São José de Itaboraí, Brazil. Part 1. Madtsoiidae, Aniliidae. Palaeovertebrata, 27:109–144.
- Rage, J.-C., 2001. Fossil snakes from the Paleocene of São José de Itaboraí, Brazil. Part II. Boidae. Palaeovertebrata 30, 111–150.
- Rage, J.-C., and A. M. Albino. 1989. Dinilysia patagonica (Reptilia, Serpentes): matériel vertébral additionnel du Crétacé supérieur d'Argentine. Étude complémentaire des vertèbres, variations intraspécifiques et intracolumnaires. Neues Jahrbuch für Geologie und Paläontologie-Monatshefte:433–447.
- Rage, J.-C., G. Métais, A. Bartolini, I. A. Brohi, R. A. Lashari, L. Marivaux, D. Merle, and S. H. Solangi. 2014. First report of the giant snake Gigantophis (Madtsoiidae) from the Paleocene of Pakistan: paleobiogeographic implications. Geobios 47:147–153.
- Rio, J. P., and P. D. Mannion. 2017. The osteology of the giant snake Gigantophis garstini from the upper Eocene of North Africa and its bearing on the phylogenetic relationships and biogeography of Madtsoiidae. Journal of Vertebrate Paleontology 37:e1347179.
- Scanferla, C. A., and J. I. Canale. 2007. The youngest record of the Cretaceous snake genus Dinilysia (Squamata, Serpentes). South American Journal of Herpetology 2:76–81.
- Scanferla, C. A, H. Zaher, F. E. Novas, C. de Muizon, and R. Céspedes. 2013. A new snake skull from the Paleocene of Bolivia sheds light on the evolution of macrostomatans. PLoS ONE 8(3):e57583.
- Scanlon, J. D. 1992. A new large madtsoiid snake from the Miocene of the Northern Territory. The Beagle, Records of the Northern Territory Museum of Arts and Sciences 9:49–60.
- Scanlon, J. D. 1997. Nanowana gen. nov., small madtsoiid snakes from the Miocene of Riversleigh: sympatric species with divergently specialised dentition. Memoirs of the Queensland Museum 41:393–412.
- Scanlon, J. D. 2004. First known axis vertebra of a madtsoiid snake (Yurlunggur camfieldensis) and remarks on the neck of snakes. The Beagle: Records of the Museums and Art Galleries of the Northern Territory 20:207–215.
- Scanlon, J. D. 2005. Australia’s oldest known snakes: Patagoniophis, Alamitophis, and cf. Madtsoia (Squamata: Madtsoiidae) from the Eocene of Queensland. Memoirs of the Queensland Museum 51:215–235.
- Scanlon, J. D., and M. S. Y. Lee. 2000. The Pleistocene serpent Wonambi and the early evolution of snakes. Nature 403:416–420.
- Simpson, G. G. 1933. A new fossil snake from the Notostylops beds of Patagonia. Bulletin of the American Museum of Natural History 67:1–22.
- Smith, M. J. 1976. Small fossil vertebrates from Victoria Cave, Naracoorte, South Australia IV. Reptiles. Transactions of the Royal Society of South Australia 100:39–51.
- Smith, T., K. Kumar, R. S. Rana, A. Folie, F. Solé, C. Noiret, T. Steeman, A. Sahni, and K. D. Rose. 2016. New early Eocene vertebrate assemblage from western India reveals a mixed fauna of European and Gondwana affinities. Geoscience Frontiers 7:969–1001.
- Vasile, Ș., Z. Csiki-Sava, and M. Venczel. 2013. A new madtsoiid snake from the Upper Cretaceous of the Haţeg Basin, western Romania. Journal of Vertebrate Paleontology 33:1100–1119.
- Vullo, R. 2019. A new species of Lapparentophis from the mid-Cretaceous Kem Kem beds, Morocco, with remarks on the distribution of lapparentophiid snakes. Comptes Rendus Palevol 18:765–770.
- Wasim A. W., R. K. Sehgal, A. Čerňanský, R. Patnaik, N. Kumar, A. P. Singh, P. Uniyal, and N. P. Singh. 2022. A find from the Ladakh Himalaya reveals a survival of madtsoiid snakes (Serpentes, Madtsoiidae) in India through the late Oligocene, Journal of Vertebrate Paleontology:e2058401.
- Wilson, J. A., D. M. Mohabey, S. E. Peters, and J. J. Head. 2010. Predation upon hatchling dinosaurs by a new snake from the Late Cretaceous of India. PLoS Biology 8:e1000322.
- Woodburne, M. O., F. J. Goin, M. Bond, A. A. Carlini, J. N. Gelfo, G. M. López, A. Iglesias, and A. N. Zimicz. 2014. Paleogene land mammal faunas of South America; a response to global climatic changes and indigenous floral diversity. Journal of Mammalian Evolution 21:1–73.
- Zaher, H., and C. A. Scanferla. 2012. The skull of the Upper Cretaceous snake Dinilysia patagonica Smith-Woodward, 1901, and its phylogenetic position revisited. Zoological Journal of the Linnean Society 164:194–238.
- Zaher, H., D. M. Mohabey, F. G. Grazziotin, and J. A. Wilson Mantilla. 2022. The skull of Sanajeh indicus, a Cretaceous snake with an upper temporal bar, and the origin of ophidian wide-gaped feeding. Zoological Journal of the Linnean Society:zlac001.
- Zimicz, A. N., M. Fernández, M. Bond, L. Chornogubsky, M. Arnal, M. Cárdenas, J. C. Fernicola. 2020. Archaeogaia macachaae gen. et sp. nov., one of the oldest Notoungulata Roth, 1903 from the early-middle Paleocene Mealla Formation (Central Andes, Argentina) with insights into the Paleocene-Eocene South American biochronology. Journal of South American Earth Sciences 103:102772.