
INTRODUCTION
The occurrence of Middle Miocene fossil mammals at Napudet, Kenya, was first documented in the 1990s by members of the Koobi Fora Research Project (KFRP). Fieldwork efforts were renewed in 2013 by Isaiah Nengo, and have since been continued by researchers involved in the Napudet Research Project (NRP). Work by the NRP has resulted in the recovery of numerous faunal remains (Rae et al., 2017), including an infant ape skull belonging to Nyanzapithecus alesi (Nengo et al., Citation2017), a small sample of hippopotamid specimens (Boisserie et al., Citation2017), small-bodied lorisoid-like primates (Myers et al., Citation2020), a partial ape skeleton (Russo et al., Citation2019), and at least eight specimens of carnivores. At an estimated age of roughly 13 Ma (Nengo et al., Citation2017), the fossils from Napudet provide a rare window into the African Middle Miocene, being younger than the faunas from Kalodirr (∼17 Ma, Boschetto et al., Citation1992) and Buluk (∼16 Ma, Lukens et al., Citation2017) and older than the fauna from nearby Lothagam (down to 7.25 Ma, Werdelin, Citation2010). As the only roughly contemporary African carnivores thus come from Member A of the Ngorora Formation, Kenya (12.5 Ma, see Morales and Pickford, Citation2005), the material from Napudet contributes important information for interpreting the evolutionary history of African carnivores.
GEOLOGICAL SETTING
Napudet is located in South Turkwel, west of Lake Turkana, northern Kenya (2°57′N, 35°52′E) (). The Middle Miocene site lays south of the Turkwel River, across from the older localities of Kalodirr and west of the younger site of Lothagam (McDougall and Brown, Citation2009; Brown and McDougall, Citation2011). Sedimentary strata at Napudet were first identified by the Kenya Geological Survey (Dodson, Citation1971). The stratigraphy at Napudet consists of four primary successive units: the Emunyan Beds, the red series, the Nachukui Formation, and the Galana Boi Formation (Nengo et al., Citation2017). All the material described here was retrieved from the Emunyan Beds, which contains both abundant fossil wood and vertebrate fossils. These sedimentary strata are comprised of a series of bedded tuffs, paleosols, and altered pyroclastics, which are underlain by a columnar basalt layer dated to approximately 13.3 Myr (Nengo et al., Citation2017), and the Emunyan Beds are capped by another, as yet undated, basalt (Nengo et al., Citation2017; Rae et al., Citation2017).
FIGURE 1. Geographic map of Kenya showing the position of the Napudet (Middle Miocene) fossil locality.
MATERIALS AND METHODS
Materials
Fossil specimens collected at Napudet are curated at the Turkana Basin Institute (TBI), Turkwel. Locations for the curation of comparative materials are provided in the text.
Terminology and measurements
Dental terminology is derived from Smith and Dodson (Citation2003), and dental measurements follow Peigné and Heizmann (Citation2003). Specimens were measured with calipers to the nearest 0.1 mm. Length of teeth refers to the greatest anterior-posterior dimension of the tooth crown, and width to the greatest bucco-lingual dimension.
Institutional Abbreviations—KNM, National Museums of Kenya, Nairobi, Kenya; OCO, Orrorin Community Organization, Kenya; KP, Kipsaraman, Kipsaraman Community Museum, Kenya; TBI, Turkana Basin Institute, Kenya.
Anatomical Abbreviations—An upper-case letter indicates a tooth in the maxillary series, a lower-case letter refers to a tooth in the mandibular series, e.g., P1, upper first premolar; m2, lower second molar.
SYSTEMATIC PALEONTOLOGY
Order CARNIVORA Bowdich, Citation1821
Family AMPHICYONIDAE Trouessart, Citation1885
Subfamily AMPHICYONINAE Hunt, Citation1998
Genus MYACYON Sudré & Hartenberger, Citation1992
MYACYON sp.
(, , )
FIGURE 2. Carnivora of Napudet (Middle Miocene of Kenya, Africa). Myacyon sp., KNM NP-64518, isolated left m2 in A, occlusal, B, buccal, C, lingual view. Small Amphicyonidae indet., KNM NP-102409, isolated left P3 in D, occlusal, E, lingual, F, buccal view; KNM NP-102410, isolated left p2 in G, lingual, H, occlusal, I, buccal view; KNM NP-102530, posterior half of right p4 in J, lingual, K, occlusal, L, buccal view. Both scale bars are 20 mm, top scale bar is for A, all other figures are scaled to the bottom scale bar.

TABLE 1. Measurements of carnivores from Napudet (Middle Miocene). ‘—‘ indicates missing measurements.
New material from Napudet—KNM NP-64518, isolated left m2 ().
Distribution—Middle Miocene of East Africa.
Description—The m2 KNM NP-64518 is larger than that of most other African amphicyonids except Cynelos jitu from Buluk, Kenya (Morlo et al., Citation2021) and Amphicyon giganteus from Arrisdrift (Morales et al., Citation2003) and Moghra (Morlo et al., Citation2019), although its maximum width approaches that of both taxa (; ). The Napudet specimen differs from C. jitu, however, in having a talonid that is much narrower than the trigonid, although less so than in other Middle Miocene amphicyonids attributed to Myacyon. The protoconid, metaconid, and hypoconid of the m2 are heavily abraded. Unlike C. jitu, no notch separates the highest cusp, the protoconid from the anterior cingulid. The trigonid is wider than the talonid. Between the metaconid and hypoconid there is a small conulid, and this cusp is separated from the hypoconid by a notch. The lingual tooth base is enlarged, but there is no clear enlargement at the posterobuccal corner.
FIGURE 3. Bivariate plot of comparative m2 sizes of large African Cynelos Jourdan, Citation1862, Amphicyon Lartet in Michelin, Citation1836, Myacyon Sudre and Hartenberger, Citation1992, and Namibiocyon Morales and Pickford, Citation2022. Data of Cynelos and Namibiocyon were taken from Morlo et al. (Citation2021) and other measurements are taken from the literature. Black symbols indicate measurements of Middle Miocene specimens, white symbols indicate measurements of Early Miocene specimens. KNM NP-64518, Myacyon sp. from Napudet measurements indicated by a star (*); Myacyon kiptalami from Ngorora Formation and Myacyon cf. kiptalami from Samburu Hills measurements indicated by black squares (▪); Myacyon dojambir indicated by white triangles (△); Amphicyon giganteus indicated by black circles (●); Cynelos anubisi measurements indicated by white squares (□); Cynelos macrodon (including Cynelos cf. macrodon from Muruyur) measurements indicated by white circles (○); Namibiocyon ginsburgi indicated by a white diamond (◇); Cynelos jitu measurements indicated by inverted white triangles (▽). Axis numbers in mm.
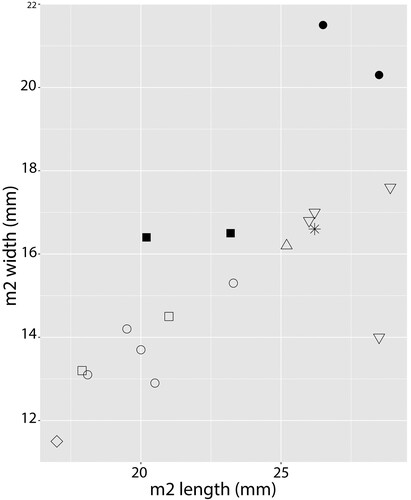
Comparisons—The much larger size of KNM-NP 64518 (), combined with the presence of a lingual conulid anterior to the hypoconid, and the lack of a posterobuccal enlargement, differentiates the Napudet m2 from the other three amphicyonid m2s known from the Middle Miocene of Kenya: KNM-SH 37907, Myacyon cf. kiptalami, from Samburu Hills, Narumungule Formation, locality SH 76 (ca. 9.5 Ma) (Tsujikawa, Citation2005:fig. 1; Morales et al., Citation2016); OCO 3’09, Myacyon kiptalami from Ngorora (ca. 12.5 Ma) (Morales et al., Citation2010:fig. 6; Morales et al. Citation2016); and KNM-MY 89, Cynelos cf. macrodon, from Muruyur (up to 14.5 Ma, Werdelin, Citation2010) (Morlo et al., Citation2019:fig. 3). KNM NP-64518 is most similar to KNM-SH 37907 but differs in having a wider talonid. It differs in the same way from OCO 3’09 and also differs from this specimen in having a taller metaconid. The Napudet M2 differs from KNM-MY 89 in having a trigonid that is much wider than the talonid, and differs from the similarly sized m2 of C. jitu in having a much narrower talonid than trigonid.
Remarks—KNM-NP 64518 establishes the presence of another enormous amphicyonid in Africa, the size of the Cynelos jitu (238.3–305.4 kg; Morlo et al., Citation2021) from Buluk (). With the exception of KNM-MY 89, all of the very large Middle Miocene amphicyonid specimens have been assigned to Myacyon kiptalami Morales and Pickford, Citation2005 (Morales et al., Citation2016). The m2 from Napudet is much larger than the other Middle Miocene specimens (). It also differs in its length and width proportions from the m2s assigned to Cynelos macrodon and C. jitu (Morlo et al., Citation2021). The fact that the talonid of the Napudet specimen is much narrower than the trigonid, coupled with its much larger size, suggests that the giant amphicyonid from Napudet is morphologically distinct enough to be assigned to a new, as yet unidentified, species of Myacyon.
AMPHICYONIDAE indet.
(, )
New material from Napudet—KNM NP-102409, isolated left P3 (); KNM NP-102410, isolated left p2 (), KNM NP-102530, right p4 fragment ().
Distribution—Middle Miocene of East Africa.
Description—KNM NP-102409 is a left upper premolar, presumably P3 (). The low, unicuspid tooth is slightly bowed, with a concave outline lingually and a convex outline buccally. The tooth is much wider posteriorly than anteriorly. The preparacrista runs anterolingually, while the postparacrista runs straight posteriorly and shows clear wear marks.
KNM NP-102410 is a lower left premolar, presumably p2 (). The tooth is very simply built, with a low protocone as the only cusp. The preprotocristid curves only very slightly lingually. A thick cingulid surrounds the tooth. No abrasion marks are visible.
KNM NP-102530 is the posterior half of a right p4 (). The specimen preserves the protoconid and its posterior wall. A centrally placed posterior accessory cuspid that is not clearly separated by a notch from the protoconid is also present, and the base of the tooth is enlarged at its posterobuccal corner.
Comparisons and Remarks—We interpret KNM NP-102409 as a P3 of a small amphicyonid (Cynelos or Myacyon) because it is morphologically close to the P3 of Cynelos lemanensis (Peigné and Heizman, Citation2003:fig. 5), but it is about 10% smaller. Moreover, of the three premolars present in amphicyonids, only P3 occludes with the lower teeth, in this case it would be the preprotocristid of p4. This kind of abrasion also differentiates KNM NP-102409 from the P3 of hyainailourids, which typically show horizontal abrasion. KNM NP-102409 is either too large, too low crowned, or differs in lacking accessory cusps to be assigned to another of the known Middle Miocene carnivores.
KNM NP-102409 is slightly smaller than the P3 of Namibiocyon ginsburgi from Arrisdrift (Morales and Pickford, Citation2022) and C. lemanensis. The specimen thus falls in the size range of C. euryodon (Savage, Citation1965), a taxon so far only reported from the Early Miocene of Africa (Morales et al., Citation2016). An m2 of the Middle Miocene Cynelos minor (Morales and Pickford, Citation2008) from Kipsaramon is morphologically identical to C. euryodon, but is 16% smaller. Given the 10% size variability in m2-length of C. lemanensis (Peigné and Heizmann Citation2003:table 2) and values of up to 25% in other amphicyonids (Peigné and Heizmann Citation2003:table 8), it seems possible that C. euryodon persisted into the Middle Miocene and that the P3 KNM NP-102409 belongs to this taxon. However, as the P3 of C. euryodon is unknown, we assign the tooth to a small Amphicyonidae indet.
KNM NP-102410, fits well in size and morphology with KNM NP-102409 if interpreted as a p2. We therefore assign this specimen to the same small amphicyonid taxon as the P3 KNM NP-102409.
KNM NP-102530 is a p4 fragment, and the low tooth crown with a centrally located posterior accessory cuspid is not typical for a feliform. Lutrines lack a posterior accessory cuspid on p4, and large mellivorines have the accessory cuspid shifted buccally. Only very few hyaenodonts are known from the African Middle Miocene (Lewis and Morlo, Citation2010), and none of these are in the size range of KNM NP-102530. Members of Hyainailouros, Dissopsalis, and Anasinopa are all larger. We tentatively assign KNM-NP 102530 to C. euryodon, due to the presence of the unseparated posterior accessory cuspid, and the distinct basal posterior enlargement, which are well known features of amphicyonids. The width of the tooth also falls within the size range expected for a small amphicyonid.
HYAENODONTA or CARNIVORA
(Fig. S1, )
Several specimens from Napudet possibly represent members of Hyaenodonta or Carnivora, but the remains are too fragmentary to be attributed to a specific taxon.
Dental material— KNM NP-64543 (Fig. S1A–B) is the root of a large, indeterminable tooth.
KNM NP-102528 (Fig. S1C–D) is an upper left canine, possibly of a small carnivore, although its tip is rather blunt.
KNM NP-102529 (Fig. S1E–G) is a single rooted, low, but very slender tooth with a main cusp and a posterior smaller cusp. It may represent p1 or P1 or M2 of a feliform or an anterior premolar of an amphicyonid.
Postcranial material—KNM NP-102552 (Fig. S1H–K) is a partial distal phalanx of a carnivoran. The specimen is unfissured, while distal phalanges of hyainailourids are fissured. The phalanx is simply built and is clearly not that of a felid. The size of the specimen cannot be estimated because the distal end is broken, but judging from the remaining part it is about the size expected for a small amphicyonid.
DISCUSSION
Though small, the collection of eight carnivore specimens from Napudet contributes to understanding the evolutionary history of these animals, especially the evolution of amphicyonids. The African fossil record documents a period of faunal turnover at the Early–Middle Miocene boundary (ca. 16 Ma), likely due to a decline in temperature following the mid-miocene Climatic Optimum (e.g., Zachos et al., Citation2001). This turnover event has been reported to include amphicyonid genera and species (Morales et al., Citation2016), however, both amphicyonid taxa from Napudet suggest a different possible scenario, in which several species of Cynelos may have evolved and persisted independently from each other into the Middle Miocene. Such a scenario was previously proposed by Morlo et al. (Citation2019) who reported a derived specimen of possibly Cynelos macrodon from the Middle Miocene.
For the large m2 from Napudet, two different scenarios are possible. It may belong to a new species of Myacyon, a strictly Middle Miocene genus that rapidly diversified after the Early–Middle Miocene boundary (Morales et al., Citation2016), including the giant amphicyonid from Napudet. In this scenario, Myacyon may have descended from one of the large Early Miocene amphicyonid species (see Tsujikawa, Citation2005; Morales and Pickford, Citation2005, Citation2008). A possible candidate is Namibiocyon ginsburgi, as the morphology of the Napudet m2 indicates a highly carnivorous adaptation. This taxon is among the most carnivorous of the Early Miocene amphicyonids as evidenced by the presence of its narrow m2 talonid, an m1 metaconid that is more reduced than in other species, and a more anteriorly pointing m1 paracristid (see Morlo et al. Citation2019, Citation2021; Morales and Pickford, Citation2022). The alternative scenario for the giant Napudet amphicyonid is that it represents a derived Cynelos jitu and evolved separately into Myacyon sp. from Napudet. This would render the genus Myacyon polyphyletic with the talonid reduction in m2 occurring separately in at least two genera. Though a rapid evolution towards gigantism in a monophyletic Myacyon independently to C. jitu (scenario 1) appears maybe less likely than a parallel evolving C. jitu (scenario 2), the latter would necessitate a complete revision of Myacyon, a topic that cannot be done based on the m2 from Napudet. We therefore refer the large m2 to Myacyon sp. instead of C. jitu.
Another piece of evidence for the hypothesis that different species of Early Miocene amphicyonids survived the Early–Middle Miocene boundary is provided by the three specimens of a small amphicyonid at Napudet that are close to the Early Miocene Cynelos euryodon. The taxon possibly also includes the Middle Miocene “Hecubides minor” (=Cynelos minor) as a junior synonym, as the size difference between C. euryodon and “C. minor” does not largely exceed the size variation of other amphicyonid species (see Peigné and Heizmann, Citation2003). However, more diagnostically relevant material (e.g., m1, P4, M1, M2) will be required to support such a scenario.
The fragmentary nature of the non-amphicyonid material from Napudet prevents the material from being identified with confidence, though the occurrence of these four specimens hints that the richness of the carnivore fauna may increase with additional fieldwork.
Supplemental Material
Download MS Word (5.6 MB)ACKNOWLEDGEMENTS
Isaiah O. Nengo was passionately dedicated to the work at Napudet. The Napudet Research Project is grateful for his leadership, generosity, and inspiration, and we will warmly and fondly remember his contributions to our lives, the lives of his students and other collaborators, and the fields of paleontology and paleoanthropology, as we honor and carry on his legacy at Napudet. Our friend and collaborator will be sorely missed. We thank the National Museums of Kenya for permission to work at Napudet, R., M. and L. Leakey, L. Martin, and the Turkana Basin Institute for encouragement and logistical support. Special thanks are due to the fossil hunters of Team Napudet, including Foothills-DeAnza students: J. Agamata, J. Contreras, P. Zamora, and J. Wang. Fieldwork at Napudet is funded by: The Leakey Foundation, Wenner-Gren Foundation, Foothill-De Anza Foundation, National Science Foundation, and National Geographic Society.
LITERATURE CITED
- Boisserie, J.-R., C. Kiarie, F. Lihoreau, & I. O. Nengo, I.O. (2017). Middle Miocene Kenyapotamus (Cetartiodactyla, Hippopotamidae) from Napudet, Turkana Basin, Kenya. Journal of Vertebrate Paleontology, 37(2):e1272055. DOI: 10.1080/02710252834.2017.1272055
- Boschetto, H. B., Brown, F. H., McDougall, I. (1992). Stratigraphy of the Lothidok range, Northern Kenya, and K/Ar ages of its Miocene primates. Journal of Human Evolution, 22, 47–71.
- Bowdich, T. E. (1821). An analysis of the natural classifications of Mammalia, for the use of students and travellers. J. Smith.
- Brown, F., & McDougall, I. (2011). Geochronology of the Turkana depression of northern Kenya and southern Ethiopia. Evolutionary Anthropology, 20, 217–227.
- Dodson, R. G., (1971). Geology of the Area South of Lodwar: Degree Sheet 18, NE Quarter, with Coloured Geological Map. Republic of Kenya, Ministry of Natural Resources.
- Hunt, R. M. Jr. (1998). Amphicyonidae: in C. M. Janis, K. M. Scott & L. L. Jacobs (Eds.). Evolution of tertiary mammals of North America Terrestrial carnivores, ungulates, and ungulatelike mammals (pp. 196–227). Volume 1. Cambridge University Press.
- Jourdan, C. (1862). Des terrains sidérolitiques. Revue des sociétés savantes. Sciences mathématiques, physiques et naturelles Paris, 1, 130–133.
- Lewis, M. E., & M. Morlo. (2010). Chapter 7: ‘Creodonta’: in L. Werdelin, & W. J. Sanders (Eds.), Cenozoic Mammals of Africa (pp. 543–560). Cambridge University Press, Cambridge; p.
- Lukens, W. E., D. J. Peppe, E. Locke, E. R. Miller, A. L. Deino, K. O. Oginga, & I. O. Nengo. (2017). Paleoenvironments and mammalian fauna of the Early Miocene fossil site at Buluk, Kenya. American Journal of Physical Anthropology, 162(S64), 268–269.
- McDougall, I., & Brown, F. H. (2009). Timing of volcanism and evolution of the northern Kenya Rift. Geological Magazine, 146, 34–47.
- Michelin, J. L. H. (1836). Notes sur les ossemens et coquilles fossiles trouvés dans un terrain d’eau douce à Simorre et Sansan. Bulletin de la Société géologique de France, 7, 217–220.
- Morales, J., Brewer, P., & Pickford M. (2010). Carnivores (Creodonta and Carnivora) from the basal middle Miocene of Gebel Zelten, Libya, with a note on a large amphicyonid from the middle Miocene of Ngorora, Kenya. Bulletin of the Tethys Geological Society, Cairo, 5, 43–54.
- Morales, J., & Pickford, M. (2005). Carnivores from the Middle Miocene Ngorora Formation (13–12 Ma), Kenya. Estudios Geologicos, 61, 271–284.
- Morales, J., & Pickford, M. (2008). Creodonts and carnivores from the Middle Miocene Muruyur Formation, Kipsaramon and Cheparawa, Baringo District, Kenya. Comptes Rendus Palevol, 7, 487–497.
- Morales, J., & Pickford, M. (2022). The taxonomic status of “Ysengrinia” ginsburgi (Morales et al. 1998 (Amphicyonidae, Carnivora) from the basal Middle Miocene of Arrisdrift, Namibia. Communications of the Geological Survey of Namibia, 24, 1–16.
- Morales, J., Pickford, M., Fraile, S, Salesa, M., & Soria, D. 2003. Creodonta and Carnivora from Arrisdrift, Early Middle Miocene of southern Namibia. Memoirs of the Geological Survey of Namibia, 19, 177–194.
- Morales, J., Pickford, M. & Valenciano, A. (2016). Systematics of African Amphicyonidae, with descriptions of new material from Napak (Uganda) and Grillental (Namibia). Journal of Iberian Geology, 42(2), 131–150.
- Morlo, M., Miller, E. R., Bastl, K., Gawad, M. A., Hamdan, M., El-Barkooky, A. N., & Nagel, D. (2019). New amphicyonids (Mammalia, Carnivora) from Moghra, Early Miocene, Egypt. Geodiversitas, 41(2), 731–745.
- Morlo, M., Friscia, A., Miller, E. R., Locke, E., & Nengo, I. O. (2021). Systematics and paleobiology of Carnivora and Hyaenodonta from Buluk, Early Miocene, Kenya. Acta Palaeontologica Polonica, 46(2), 465–484. DOI: 10.4202/app.00794.2020
- Myers, L. C., Russo, G. A., & Nengo, I. O. (2020). A lorisoid-like distal humerus from the Middle Miocene at Napudet, Kenya. American Journal of Physical Anthropology, 171, 195.
- Nengo, I. O., Tafforeau, P., Gilbert, C. C., Fleagle, J., Miller, E. R., Feibel, C., Fox, D. L., Feinberg, J., Pugh, K. D., Berruyer, C., Mana, S., Engle, Z., & Spoor, F. (2017). New infant cranium from the African Miocene sheds light on ape evolution. Nature, 548(7666), 169–174.
- Peigné, S., & Heizmann, E. P. J. (2003). The Amphicyonidae (Mammalia: Carnivora) from Ulm-Westtangente (MN 2, Early Miocene), Baden-Württemberg, Germany: Systematics and Ecomorphology. Staatliches Museum für Naturkunde, Serie B, 343, 1–133.
- Rae, T. C., Nengo, I. O., Currano, E. D., Feibel, C., Fox, D. L., & Miller, E. R. (2017). New palaeontological discoveries from the Middle Miocene site of Napudet, west Turkana, Kenya. Conference: American Association of Physical Anthropologists annual meeting 2016.
- Russo, G. A., Ward, C. V., Kuo S., McGechie, F. R., & Nengo, I. O. (2019). An ape partial postcranial skeleton from the Middle Miocene of Napudet, Turkana Basin, Kenya. American Journal of Physical Anthropology, 168, 211.
- Savage, R. J. G. (1965). Fossil mammals of Africa: 19. The Miocene Carnivora of East Africa. Bulletin of the British Museum of Natural History (Geology), 10(8), 239–316.
- Sudre, J., & Hartenberger, J.-L. (1992). Oued Mya 1, nouveau gisement de mammifères du Miocène supérieur dans le sud algérien. Géobios, 25, 553–565.
- Smith, J. B., & Dodson. P. (2003). A proposal for a standard terminology of anatomical notation and orientation in fossil vertebrate dentitions. Journal of Vertebrate Paleontology, 23, 1–12.
- Trouessart, E. L. (1885). Catalogue des mammiferes vivants et fossiles (carnivores). Germain et G. Grassin, Angers.
- Tsujikawa, H. (2005). The Updated Late Miocene Large Mammal Fauna from Samburu Hills, Northern Kenya. African study monographs. Supplementary issue, 32, 1–50.
- Werdelin, L. (2010). Chronology of Neogene mammal localities: in L. Werdelin & W. J. Sanders (Eds.), Cenozoic Mammals of Africa (pp. 27–43). Cambridge University Press.
- Zachos, J., Pagani, M., Sloan, L., Thomas, E., & Billups, K. (2001). Trends, rhythms and aberrations in global climate 65 Ma to present. Science, 292, 686–693.