
ABSTRACT
Among non-avian dinosaurs, troodontids are relatively rare but diverse. The Nemegt Basin in the Mongolian Gobi Desert, which incorporates three of the most fossiliferous beds in the world, is one such region with high troodontid diversity and has also produced eight troodontid taxa until now. The diversity of troodontids is biased towards the two formations, Nemegt and Djadochta. Despite its rich vertebrate fossil record, no troodontids have been described from the Baruungoyot Formation. This faunal absence is perplexing as the Baruungoyot Formation has often been considered intermediate between the other two units in stratigraphy and paleoenvironment, but there are no definite troodontid materials that could fill this ‘gap.’ Here, we report a new troodontid, Harenadraco prima gen. et sp. nov., from the Baruungoyot Formation in Hermiin Tsav, Mongolia. It is represented by an incomplete skeleton that mainly consists of partially articulated left hind limb elements. Harenadraco is small and lightly built like other troodontids, but its tarsometatarsus exhibits extreme slenderness comparable only to Philovenator among cursorial non-avian maniraptorans. It also implies high cursoriality that could be an adaptation of Harenadraco to its environment where potential prey animals like mammals and predators such as eudromaeosaurs were likely to be agile. The discovery of Harenadraco confirms the presence of troodontids in all three formations in the Nemegt Basin.
INTRODUCTION
Troodontids are a diverse group of theropod dinosaurs, generally characterized by large orbits, long hind limbs with asymmetrical metatarsi, and an enlarged ungual on the second pedal digit as well as many bird-like features (Makovicky & Norell, Citation2004). They have been found in the sedimentary deposits that range from the Late Jurassic to the latest Cretaceous of Asia and North America (Hartman et al., Citation2019; Makovicky & Norell, Citation2004). The Upper Cretaceous beds in the Gobi Desert have been a rich source of many troodontid taxa, especially the Nemegt and Djadochta formations in the Nemegt Basin, Mongolia (Jerzykiewicz et al., Citation2021) as well as the Wulansuhai Formation in Bayan Mandahu, China (Pei et al., Citation2017, Citation2022; Xu et al., Citation2011). From the former, eight troodontid taxa are currently known (Jerzykiewicz et al., Citation2021). The Baruungoyot Formation, however, has been a puzzling exception because no troodontids have been found even though it is one of the major sedimentary formations in the Nemegt Basin and interpreted to be intermediate to the Djadochta and Nemegt formations in terms of age and/or environment (Gradzínski et al., Citation1977; Jerzykiewicz et al., Citation2021). As the faunal compositions of the Djadochta and Baruungoyot formations are very similar, the absence of troodontids in the latter is even more puzzling.
In 2018, a crew of the SNU-IP Expedition discovered a small theropod partial skeleton in the Baruungoyot Formation at Hermiin Tsav in the Mongolian Gobi Desert (Fig. S1). It includes fragmentary pelvic bones and partially articulated hind limb elements and is identified as a new troodontid. The new taxon is the first troodontid recognized from the Baruungoyot Formation (). Although the specimen mainly consists of hind limb bones, it provides meaningful information on the vertebrate evolution of the Baruungoyot Formation. Additionally, the morphology of its tarsometatarsus is quite peculiar and stands out among theropods with arctometatarsalian conditions due to its extreme slenderness.
FIGURE 1. Distribution of troodontids in the Nemegt Basin. The map is based on the image of Google Earth.
METHODS
Phylogenetic Analysis
In this study, the character list in Wang et al. (Citation2022), which is based on that of Pei et al. (Citation2020), was used to score the characters of Harenadraco. The scorings were added to the most recent Theropod Working Group (TWiG) data matrix from Wang et al. (Citation2022), which resulted in a total of 171 taxa and 855 characters. LH PV39, an unidentified troodontid represented mainly from vertebrae from the Wulansuhai Formation was treated as an independent operational taxonomic unit (OTU) in our analysis. The same methods (except for the max ram increase from 100 to 1000) of the parsimony analysis in Wang et al. (Citation2022), which followed Pei et al. (Citation2020) (initial Wagner trees with five random addition sequences followed by TBR, sectorial searches with CSS, RSS, and XSS, and five cycles of tree drifting that was followed by tree fusing), were used with TNT version 1.6 (Goloboff & Morales, Citation2023). Our analysis was done under both extended implied weighting (XIW) and equal weighting (EW), and unstable taxa were identified with heuristic and new techniques and all pruned automatically using the scripts from Pei et al. (Citation2020). All the used scripts (.run files) and data matrix (datamatrix.tnt), the produced trees (.svg files), and the batch file (winanalyze.bat) that can run the analysis are included in Supplementary File 1.
Institutional Abbreviations—MPC, Institute of Paleontology, Mongolian Academy of Sciences, Ulaanbaatar, Mongolia; SNU, Seoul National University, Seoul, Republic of Korea.
SYSTEMATIC PALEONTOLOGY
DINOSAURIA Owen, Citation1842
THEROPODA Marsh, Citation1881
TROODONTIDAE Gilmore, Citation1924
HARENADRACO PRIMA, gen. et sp. nov.
()
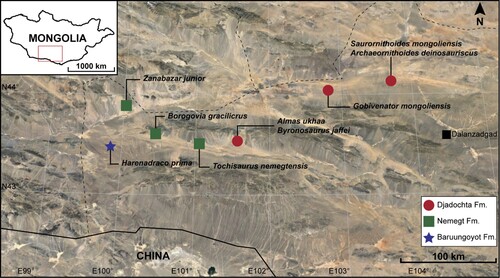
FIGURE 2. Femora of Harenadraco prima (MPC-D 110/119, holotype). A–D, left femur in A, anterior, B, posterior, C, medial, and D, lateral views. E–H, right femur in E, anterior, F, posterior, G, medial, and H, lateral views. Abbreviations: ft: fourth trochanter; pf, popliteal fossa; pt, posterior trochanter.
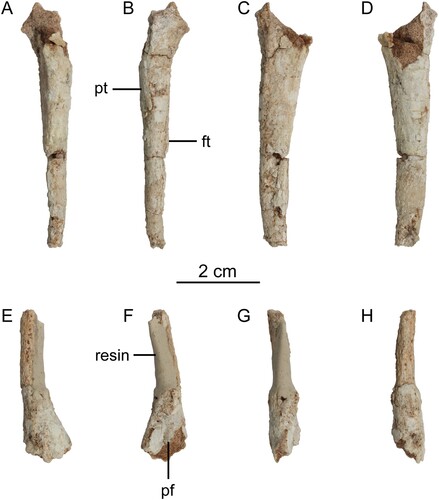
FIGURE 3. Tibiotarsi of Harenadraco prima (MPC-D 110/119, holotype). A–D, proximal left tibiotarsus in A, anterior, B, posterior, C, medial, and D, lateral views. E–I, distal left tibiotarsus in E, anterior, F, posterior, G, medial, H, lateral, and I, distal views. Abbreviations: ap, ascending process of astragalus; cc, cnemial crest; fc, fibular crest; ig, intercondylar groove; lc, lateral condyle; msmc, medial margin of the medial condyle.
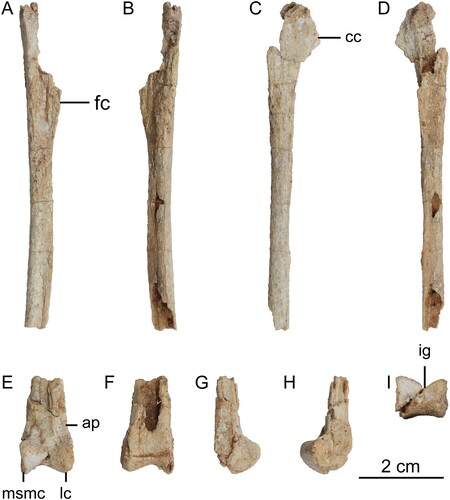
FIGURE 4. Left tarsometatarsus of Harenadraco prima (MPC-D 110/119, holotype). A–F, left tarsometatarsus in A, dorsal, B, ventral with a line drawing of the proximal end on the right, C, medial, D, lateral, E, proximal, and F, distal views. The dashed lines in the line drawing indicate eroded margins. Abbreviations: mt II, metatarsal II; mt III, metatarsal III; mt IV, metatarsal IV; vf, ventral flange.
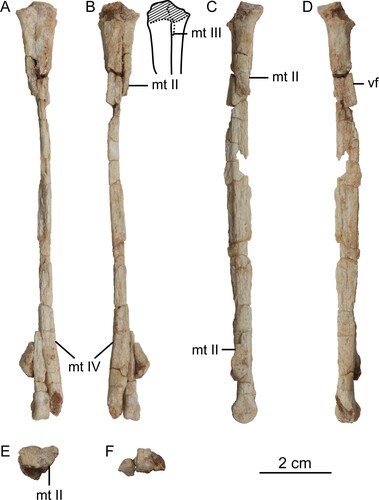
FIGURE 5. Left pedal phalanges and the skeletal reconstruction of the preserved parts in Harenadraco prima (MPC-D 110/119, holotype). A, left pedal digits II–IV in lateral view. B, left pedal phalanx III-2 in proximal view. C–F, left pedal phalanx III-3 in C, dorsal, D, ventral, E, proximal, and F, distal views. G–L, left pedal phalanx IV-1 in G, dorsal, H, ventral, I, medial, J, lateral, K, proximal, and L, distal views. M, skeletal reconstruction of Harenadraco prima (MPC-D 110/119, holotype) with missing parts in gray. Abbreviations: II-3, pedal phalanx II-3; III-3, pedal phalanx III-3; IV-2, pedal phalanx IV-2.
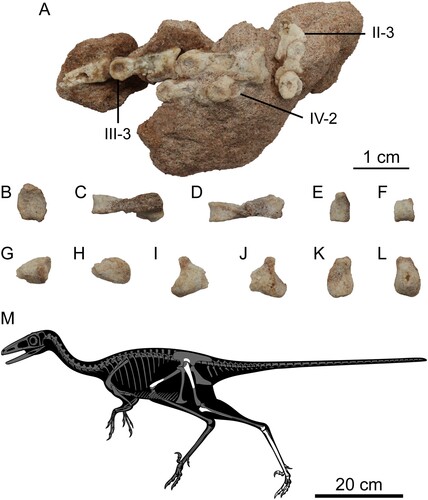
Holotype—MPC-D 110/119, a partial postcranial skeleton that consists of two fragments of the right ilium, incomplete left hind limb elements including the proximal femur, distal tibiotarsus, metatarsals II–IV, and pedal phalanges, a distal portion of the right femur, and unidentified bone fragments.
Locality and Horizon—Baruungoyot Formation (Upper Cretaceous) at Hermiin Tsav, Ömnögovi Province, Mongolia (Eberth, Citation2018).
Etymology—The name of the genus is a composition of the Latin words harena (sand) and draco (dragon). The species name “prima” means first in Latin, referring to the taxon being the first troodontid from the Baruungoyot Formation.
Diagnosis—A small-sized troodontid diagnosed by the following unique combination of characters (autapomorphies indicated with an asterisk): tall S-shaped medial margin of medial condyle of astragalocalcaneum in anterior view*, poorly developed flexor sulcus on the distal articular end of metatarsal II, which also lacks collateral ligament fossae*, metatarsal III excluded from the proximal end of the metatarsus in dorsal view (shared with Tochisaurus, Zanabazar, and Philovenator), metatarsal IV only slightly more robust than metatarsal II at the proximal end in ventral view (shared with Daliansaurus and Philovenator), pronounced ventral protrusion near the proximal end of metatarsal IV (shared with Liaoningvenator), extremely narrow proximal shaft of metatarsal IV*, greatly elongated flexor tubercle of pedal phalanx II-3 that reaches the shaft of the preceding phalanx when in articulation*, and the distal articular surface of pedal phalanx III-3 not ginglymoid*.
Differential Diagnosis—Harenadraco differs from Borogovia gracilicrus (Cau & Madzia, Citation2021; Osmólska, Citation1987) in the following characters: both condyles of the astragalocalcaneum comparably extending proximally (in Borogovia, the lateral condyle extends farther proximally), narrower distal end of metatarsal II than those of metatarsals III and IV (in Borogovia, they are subequal in width), absence of a flexor groove on the distal end of metatarsal II (present in Borogovia), quadrangular ventral surface of the distal end of metatarsal III bearing a longitudinal sulcus (triangular without a sulcus in Borogovia), presence of a distinct diaphysis of pedal phalanx II-2 (absent in Borogovia), enlarged and strongly recurved ungual of the second pedal digit (not recurved in Borogovia), much shorter pedal phalanx III-3 relative to phalanx III-2 (subequal in length in Borogovia), and slenderer phalanges of the fourth pedal digit than those of the third digit (more robust in Borogovia).
Harenadraco differs from Tochisaurus nemegtensis (Kurzanov and Osmólska, Citation1991) in having a relatively narrower proximal shaft of metatarsal IV with a sharp triangular cross section (more robust and round subtriangular cross section in Tochisaurus).
Harenadraco is distinguished from Zanabazar junior (Barsbold, Citation1974; Norell et al., Citation2009) by the absence of a swollen area supporting the medial condyle of the astragalocalcaneum (present in Zanabazar), the anteroposteriorly elongated medial fossa on the medial condyle of the astragalocalcaneum (proximodistally elongated in Zanabazar), and metatarsal II only slightly narrower than metatarsal IV in ventral view (much narrower in Zanabazar).
Harenadraco is different from Almas ukhaa (Pei et al., Citation2017) in having anteriorly oriented condyles of the astragalocalcaneum (anteroventrally oriented in Almas) and lacking a prominent ridge on the dorsal surface of the distal shaft of metatarsal IV (present in Almas).
Harenadraco differs from Byronosaurus jaffei (Makovicky et al., Citation2003; Norell et al., Citation2000) in the absence of collateral ligament fossae on the distal articular end of metatarsal II (present in Byronosaurus) and pedal phalanx II-2 with a relatively reduced and dorsoventrally low shaft (robust and dorsoventrally high in Byronosaurus).
Harenadraco is differentiated from Gobivenator mongoliensis (Tsuihiji et al., Citation2014) by having both condyles of astragalus with comparable proximodistal heights (in Gobivenator, the lateral condyle is higher than the medial one), metatarsal IV that is slightly more robust than metatarsal II at the proximal end (much more robust in Gobivenator), and a mediolaterally narrowed proximal shaft of metatarsal IV (in Gobivenator, the proximal shaft does not drastically change in width).
Harenadraco is different from Saurornithoides mongoliensis (Norell et al., Citation2009; Osborn, Citation1924) in the following characters: slightly ginglymoid metatarsal III (not ginglymoid in Saurornithoides), the subrectangular cross section of the distal shaft of metatarsal IV (round in Saurornithoides), robust distal articular end of pedal phalanx II-2 that dorsally extends above the level of the shaft (in Saurornithoides, the distal articular end does not extend above the shaft), elongated flexor tubercle of pedal phalanx II-3 (ungual) that proximally extends beyond the distal articular end of the preceding phalanx (relatively short in Saurornithoides), and flat ventral surface of pedal phalanx IV-1 near the proximal end (rounded in Saurornithoides).
Harenadraco is distinguished from Philovenator curriei (Currie and Peng, Citation1993; Xu et al., Citation2012) by the presence of an S-shaped medial margin of the medial condyle of the astragalocalcaneum (smooth in Philovenator), a shallow and wide intercondylar groove on the astragalocalcaneum (deep and narrow in Philovenator), metatarsal II becoming more extensive dorsoventrally at the proximal end (absent in Philovenator), a dorsal protrusion of the proximal ends of metatarsals II and IV (absent in Philovenator), a nearly nonexistent exposure of the proximal end of metatarsal III in ventral view (limited but relatively well exposed in Philovenator), and the modest ventral flange of metatarsal IV with a smooth proximal margin (very pronounced ventral flange with an abrupt and stiff proximal margin in Philovenator).
Harenadraco is differentiated from Papiliovenator neimengguensis (Pei et al., Citation2022) by having metatarsal III with a narrower distal shaft compared with metatarsal IV in ventral view (comparable in Papiliovenator).
Harenadraco also differs from Linhevenator tani (Xu et al., Citation2011) in lacking a longitudinal groove on the dorsal surface of the distal shaft of metatarsal III (present in Linhevenator) and having a transversely narrow and ridge-like dorsal surface of the proximal shaft of metatarsal IV (flat in Linhevenator).
DESCRIPTION
The holotype of Harenadraco prima (MPC-D 110/119) consists of hind limb elements and several fragmentary postcranial bones (see Table S1 for measurements). The pelvic girdle of MPC-D 110/119 includes two small fragments of the right ilium. Besides the well-developed brevis shelf and accompanied brevis fossa visible from the partial right postacetabular process, very little anatomical information can be obtained from them.
Both femora are partially preserved (). The left femur is represented by its proximal part but without the femoral head. The shaft is only slightly curved anteriorly. Most of the greater and lesser (anterior) trochanters are missing, but the raised mound-like posterior trochanter is observed on the posterolateral surface. The fourth trochanter is low and weakly developed. In most troodontids, the fourth trochanter is absent except for in a few taxa, including Linhevenator (Xu et al., Citation2011) and Liaoningvenator (Shen et al., Citation2017). Distally, the shaft markedly narrows, although it is broken at the mid-shaft region. The right femur preserves a small portion of the distal shaft and extremity except for the condyles. A short ridge is developed on the posterior surface proximal to the shallow popliteal fossa. Near the distal end of the medial surface is a robust process that may be homologous to the one in Philovenator (Xu et al., Citation2012).
Only the left tibiotarsus is preserved (). It is broken into two pieces and missing the proximal articular surface and the distal part of the shaft. The cnemial crest is straight and anteriorly oriented as in Philovenator (Xu et al., Citation2012), although its shape is uncertain due to the eroded margin. The fibular crest is thin and extends anterolaterally. Distal to the fibular crest, a low ridge defines the lateral margin of the tibial shaft. The shaft is almost straight but slightly curves medially near the presumed mid-shaft region. It is also very slender and subtriangular in cross section. Distally, the tibia is flattened and mediolaterally expands to have a subrectangular cross section. On the posterolateral margin, a prominent crest marks the posterior margin of a fossa that might have received the fibula like that of Zanabazar (Norell et al., Citation2009).
The left astragalus and calcaneum are firmly articulated but not co-ossified with the distal end of the tibia (). However, these two proximal tarsals are fused to form a single astragalocalcaneum, which does not exhibit any suture. Complete fusion of the astragalus and calcaneum is also known in several other troodontids such as Sinornithoides (Russell & Dong, Citation1993), Zanabazar (Norell et al., Citation2009), Philovenator (Xu et al., Citation2012), Almas (Pei et al., Citation2017), and Borogovia (Cau & Madzia, Citation2021), whereas they are incompletely fused in Gobivenator (Tsuihiji et al., Citation2014) and not fused at all in Talos (Zanno et al., Citation2011). The ascending process is heavily damaged and does not retain its proximal end. Distally, it covers the entire width of the anterior surface of the tibia. A distinct groove separates the distal condyles from the ascending process, similar to other troodontids (Norell et al., Citation2009; Zanno et al., Citation2011). They are also separated by the intercondylar groove extending to the posterior surface of the astragalar body. It is shallow and wide unlike the deep and narrow groove in Philovenator (Xu et al., Citation2012). The two condyles are subtriangular and similar in proximodistal height, although the medial one extends further anteriorly and distally. In addition, it has a steeper distal margin than the lateral condyle. The tall S-shaped medial margin of the medial condyle makes the distal part of the medial surface visible in anterior view. The anteroposterior length of the medial condyle is about 1.1 times greater than the proximodistal height, unlike Philovenator, in which this ratio is 1.6 (Xu et al., Citation2012). The astragalar body caps the entire distal surface of the tibia but does not reach its posterior surface.
The left tarsometatarsus was preserved in articulation with the tibia and astragalocalcaneum (). There is no sign of any distal tarsals, but the contact between the tarsometatarsus and the astragalocalcaneum when they were discovered suggests the distal tarsals are likely to be co-ossified with the metatarsus. Metatarsals I and V are not preserved, and the rest are incomplete. The tarsometatarsus exhibits a typical troodontid arctometatarsalian condition involving asymmetry and a pinched metatarsal III (Makovicky & Norell, Citation2004). It is also very elongated and slender. Proximally, metatarsal II expands mediolaterally to have a broad proximal articular surface, which also becomes more extensive dorsoventrally to produce a curved dorsal margin that leads to a dorsal projection unlike a flat dorsal margin in Philovenator (Xu et al., Citation2012). It is missing most of its shaft, and only a fraction of the proximal shaft remains. Nonetheless, the location of the distal contact between metatarsals II and III implies that metatarsal II must have been much shorter than metatarsal IV. The small broken surface of the distal part indicates that the shaft must have narrowed towards the distal end to become a very thin bone as in other derived troodontids (Currie & Peng, Citation1993; Kurzanov & Osmólska, Citation1991; Osmólska, Citation1987; Tsuihiji et al., Citation2014; Zanno et al., Citation2011). The distal articular surface of metatarsal II is simple and subrectangular in shape due to reduced condyles.
Metatarsal III preserves the distal shaft and articular end as well as a part of the proximal shaft. There is no trace of metatarsal III at the proximal end of the tarsometatarsus in dorsal view, and its exposure in ventral view is greatly reduced unlike in Philovenator (Currie & Peng, Citation1993; Xu et al., Citation2012). This ventral exposure is limited to near the proximal end and almost invisible. It must have been even less exposed originally as the eroded ventral surface of metatarsal II caused more exposure. The distal shaft of metatarsal III tapers proximally and ventrally, resulting in a pointed ventral surface and a triangular cross section. This is also a common feature among troodontids (Kurzanov & Osmólska, Citation1991; Norell et al., Citation2009; Zanno et al., Citation2011) and other theropods with an arctometatarsalian condition (Snively et al., Citation2004). In dorsal view, metatarsal III does not overlap metatarsal IV, unlike in Borogovia, in which metatarsal III mediolaterally broadens to extend over metatarsal IV (Cau & Madzia, Citation2021). The distal articular end of metatarsal III is mediolaterally narrow but dorsoventrally expanded. It is marginally ginglymoid with a shallow longitudinal groove that extends to the ventral surface. Dorsally, a deep extensor fossa is developed proximal to the articular end. The distal condyles of metatarsal III differ, the medial one being larger. Such a condition is also known in Talos (Zanno et al., Citation2011). The ligament fossae are also asymmetrical, as the medial fossa is much smaller.
Metatarsal IV is nearly complete and preserves most of its length. Proximally, it is only slightly more robust than metatarsal II. Both metatarsals II and IV have similar mediolateral widths at the proximal end in dorsal view. In other troodontids, the proximal end of metatarsal IV tends to be much wider (Currie & Peng, Citation1993; Kurzanov & Osmólska, Citation1991; Tsuihiji et al., Citation2014; Zanno et al., Citation2011). On the other hand, the ventral surface of metatarsal IV is conspicuously broader than that of metatarsal II. There is a robust dorsal protrusion at the proximal end, which is absent in Philovenator (Xu et al., Citation2012). Metatarsal IV has an extremely thin and mediolaterally compressed shaft. The shaft bears a proximodistally long ventral flange, but it is modestly developed with a smooth proximal margin unlike the relatively more pronounced tall flange with an abrupt and stiff proximal margin in Philovenator (Xu et al., Citation2012). The medial surface of the ventral flange is offset from the rest of the shaft, presumably to accommodate metatarsal III. It suggests that metatarsal III proximally extended at least to the level of the proximal shaft of metatarsal IV. Distal to the flange, metatarsal IV mediolaterally expands and has a subrectangular cross section. The preserved distal articular end of metatarsal IV occupies only a small ventral part, which is comparable to the distal end of metatarsal III.
Most left pedal phalanges are preserved (, S2). Pedal phalanx I-1 is the only preserved phalanx from the first pedal digit, although it only retains the proximal end. It has a concave proximal articular surface, which is subtriangular in shape and slightly wider than tall. Dorsally, a pointed tip overhangs the proximal articular surface. On the ventral surface, there is a prominent tubercle on each side.
The phalanges from the second pedal digit are in articulation with each other. Phalanx II-1 is incomplete, with only the distal articular end being preserved. It is large and bears a deep groove that extends throughout the entire articular surface, separating the two distal condyles. The lateral condyle is larger than the medial one, as in Borogovia (Osmólska, Citation1987). On the other hand, the lateral collateral ligament fossa is smaller in opposition to the condition in Borogovia (Osmólska, Citation1987) or Talos (Zanno et al., Citation2011). Only the lateral surfaces of II-2 and II-3 are exposed. Phalanx II-2 is highly modified with a robust proximoventral heel to receive the enlarged distal articular end of II-1 and a shortened shaft, which are shared with other derived troodontids (Makovicky & Norell, Citation2004). The proximal articular surface is also the tallest part in II-1 and is marked by a central ridge that articulates with the groove of the preceding phalanx. The shaft is short but not to the extent that several other troodontids exhibit (Osmólska, Citation1987; Xu et al., Citation2012; Zanno et al., Citation2011). Distally, the shaft becomes lower to form a weakly developed neck. The distal articular end of II-2 is smaller than that of II-1 but has a larger lateral collateral ligament fossa. Phalanx II-3 is missing its distal half, but it is enlarged and recurved as in most troodontids (Currie & Peng, Citation1993) except for Borogovia (Cau & Madzia, Citation2021; Osmólska, Citation1987). In lateral view, it has a short dorsal lip at the proximal articular end. The flexor tubercle, on the other hand, is much elongated and extends proximally beyond the level of the distal articular end of II-2, which distinguishes it from the relatively short and more bulbous ones of other troodontids (Currie & Peng, Citation1993; Zanno et al., Citation2011). The lateral surface of II-3 is poorly preserved, but a groove that extends to the flexor tubercle can be recognized.
Pedal digit III is only missing phalanx III-1. Phalanx III-2 is the longest and most robust element among the preserved pedal phalanges. Its proximal articular surface is characterized by a ventrally positioned slight convexity and a concave ventral margin, which is more sharply curved medially. It also has an elongated shaft that is dorsoventrally not much shallower than the articular ends. At the distal end of the shaft, a deep extensor fossa is present on the dorsal surface. The distal articular end is not ginglymoid and has a pronounced collateral ligament fossa on each side. Phalanx III-3 is much shorter than III-2 but still more robust than the preserved phalanges from the fourth digit. Its proximal articular surface is markedly concave and has a flat ventral margin. The shaft is mediolaterally compressed to a much greater extent than that of III-2. Like III-2, however, the distal articular end is not ginglymoid. The collateral ligament fossae of III-3 are asymmetrical, the lateral one being much larger and deeper. Phalanx III-4 is missing the distal tip, and most of its lateral surface is damaged. It is dorsoventrally low and somewhat straight, partly due to the highly reduced flexor tubercle, unlike in Talos (Zanno et al., Citation2011). The dorsal and ventral margins of the proximal articular surface are nearly flat.
Pedal digit IV is missing the distalmost phalanx. Only the proximal half of phalanx IV-1 is preserved. It is mediolaterally narrow and has an asymmetrical articular surface, which is uniformly concave and subtrapezoidal. The ventral heel is pronounced and medially more robust as in Talos, but no distinct pit is present on each side unlike in this taxon (Zanno et al., Citation2011). The preserved proximal shaft sharply tapers distally. Phalanx IV-2 preserves the distal end and a small portion of the distal shaft. The distal articular end has a deep lateral collateral ligament fossa. Phalanx IV-3 is slender, having a long shaft and low articular ends. The ventral heel at the proximal end is relatively robust. It protrudes ventrally more than the distal articular end, which has a large and deep lateral collateral ligament fossa that occupies most of its lateral surface. Phalanx IV-4 is short and slender. The proximal articular surface has a proximodistally tall central ridge. It is dorsally pointed, whereas its ventral margin is wide and flat. The ventral heel is robust like that of IV-3 but laterally more pronounced, and the ventral protrusion is limited. A shallow but well-defined extensor fossa is developed just proximal to the distal articular end. The distal articular surface has a wide groove that does not extend to the dorsal surface. The lateral condyle is slightly larger than the medial one. It also has a deeper and larger collateral ligament fossa.
PHYLOGENETIC ANALYSIS
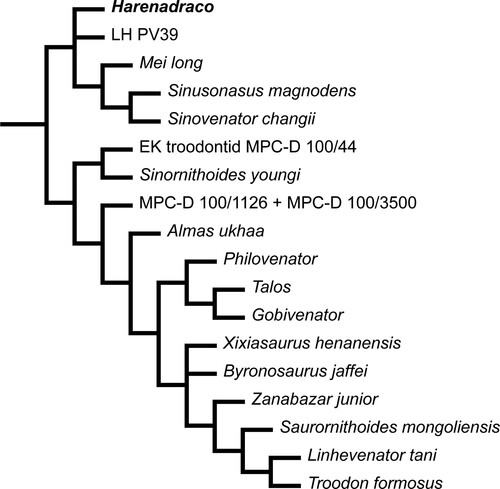
A total of four reduced consensus trees (under EW and XIW, with and without pruning of unstable taxa) were obtained from our phylogenetic analysis (Figs. S3–S6). They all have similar overall topology to those of Wang et al. (Citation2022). Under EW, the monophyly of Deinonychosauria is supported, but Dromaeosauridae and Troodontidae are not supported without pruning as in Wang et al. (Citation2022). Furthermore, Harenadraco and several other troodontid OTUs such as Philovenator are regarded unstable under EW. On the other hand, under XIW, the monophyletic Dromaeosauridae and Troodontidae are relatively well resolved within Deinonychosauria even without pruning. The topology of both reduced consensus trees under XIW is also similar to that of Wang et al. (Citation2022). It is notable that the troodontid affinity of Harenadraco is supported and not pruned under XIW. However, the position of Harenadraco is somewhat peculiar because it forms a separate clade with several Early Cretaceous troodontids from China, although LH PV39 from the Wulansuhai Formation is also included in this clade (). The phylogenetic relationship between Harenadraco and LH PV39 is not resolved, and it is currently not possible to directly compare their morphology because there are no overlapping elements. More complete specimens are thus essential in order to fully understand the relationship and trait evolution within Late Cretaceous troodontids from the Gobi Desert.
FIGURE 6. Phylogeny of Troodontidae in the reduced consensus tree under XIW. Pruning of unstable taxa resulted in the exact same topology.
DISCUSSION
Metatarsal Morphology of Harenadraco prima
One of the most prominent characteristics of Harenadraco is its remarkably slender tarsometatarsus. Although most of the shaft of metatarsal II is missing, the cross sections of both proximal and distal ends suggest a markedly narrow shaft, which is common in derived troodontids (Currie & Peng, Citation1993; Kurzanov & Osmólska, Citation1991; Osmólska, Citation1987; Tsuihiji et al., Citation2014; Zanno et al., Citation2011). Although metatarsal IV is more robust than metatarsal II, it is still very thin and slender. Based on the preserved elements, the length of the tarsometatarsus is about 22 times its estimated mediolateral width, which is an extraordinarily high ratio among small- to medium-sized theropods with an arctometatarsalian condition and also compared with palaeognaths (). Only Philovenator, another small-sized troodontid, and Rhea, a modern palaeognath, have comparably slender metatarsals with similar ratios. Unfortunately, the lengths of other hind limb bones are unknown in Harenadraco, but the preserved shafts of the femur and the tibia suggest they are also very slender (, ). It cannot be ruled out that the holotype of Harenadraco is a juvenile, and the proportion might have been different in adults. However, the extremely thin nature of the compact bones from the hind limb and their severe microbial erosion prevented proper osteohistological analysis. Determining the exact ontogenetic stage of MPC-D 110/119 is thus currently not feasible, but it seems unlikely that the slenderness of the metatarsus is transient in the life cycle of Harenadraco as troodontids generally have gracile metatarsi (e.g., Gobivenator, Tochisaurus, Philovenator, and Talos). Slender and elongated limbs and arctometatarsalian conditions in theropods are commonly associated with cursoriality (Holtz, Citation1995). The morphology of the tarsometatarsus and other hind limbs of Harenadraco thus indicates its highly cursorial lifestyle (). Nevertheless, the extreme slenderness of the tarsometatarsus is peculiar even compared with other cursorial theropods with arctometatarsalian conditions. Moreover, the slenderness of the metatarsus does not appear to be size dependent in troodontids, although the sample size is small (Table S2). It is also notable that Philovenator is the only other troodontid with a similar metatarsal proportion. Whether the extreme slenderness of the metatarsus was independently acquired by each taxon is unclear, but there is no evidence of a close relationship between the two taxa in our phylogenetic analysis (). As they are both represented mostly by their hind limb elements, more specimens of these two taxa will be helpful in understanding their relationship.
FIGURE 7. Metatarsal proportions of arctometatarsalian theropods and palaeognaths. Omitted taxa from left to right: Archaeornithomimus asiaticus, Harpymimus okladnikovi, Ornithomimus sp., Ceratonykus oculatus, Linhenykus monodactylus, Nemegtonykus citus, Citipes elegans, Avimimus portentosus, Daliansaurus liaoningensis, Liaoningvenator curriei, Linhevenator tani, Tochisaurus nemegtensis, Aptornis otidiformis, and Rhynchotus rufescens. The y-axis indicates the length/width ratio of metatarsus (metatarsus maximum length / metatarsus transverse width at mid-shaft). The metatarsal proportions of Harenadraco prima, Philovenator curriei, and one specimen of Rhea americana only exceed 20. Please see Table S2 for the metatarsal proportion of each taxon.
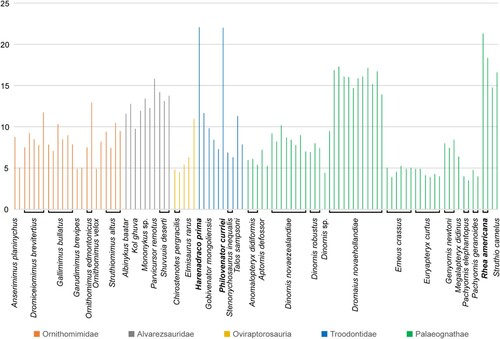
FIGURE 8. Life reconstruction of Harenadraco prima (artwork by Yusik Choi).

Vertebrate Fauna of the Baruungoyot Formation
The Baruungoyot Formation exhibits a diverse vertebrate fauna that is rich in dinosaurs. Theropods, especially maniraptorans, are the most diverse among them, similar to the faunas of the Nemegt and Djadochta formations (Jerzykiewicz et al., Citation2021). Most major maniraptoran clades, such as alvarezsaurids, oviraptorids, dromaeosaurids, and even avialans, are known from all three formations, although only a few species overlap across two of them (Chiappe et al., Citation2001; Fanti et al., Citation2012). One exception to this has been Troodontidae, as Harenadraco is the first reported troodontid from the Baruungoyot Formation, and each of the other two formations has yielded multiple troodontid taxa (). The lack of troodontids from the Baruungoyot Formation thus far is very peculiar because it is generally considered to be intermediate between the Djadochta and Nemegt formations in regards to age and/or environment (Gradzínski et al., Citation1977; Jerzykiewicz et al., Citation2021). With the discovery of Harenadraco, it is now apparent that this “gap” in the troodontid record was a sampling artifact. The fragile nature of the holotype of Harenadraco further supports this. The presence of Harenadraco in the Baruungoyot Formation also demonstrates that troodontids of the Nemegt Basin were diverse enough to adapt to various environments.
AUTHOR CONTRIBUTIONS
SL gathered and analyzed the data and drafted the manuscript. Y-NL designed and supervised the project. S-HK and J-YP produced the figures. ZB, DI, and KT provided resources. All authors contributed to the discussion and editing of the manuscript.
SUPPLEMENTARY FILES
Supplementary File 1. Figs. S1–S6, Tables S1, S2, and Literature Cited.
Supplementary File 2. Scripts and data matrix used in our phylogenetic analysis and the produced trees.
Supplemental Material
Download Zip (13.8 MB)ACKNOWLEDGMENTS
We want to thank all the field crew of the SNU-IP Expedition in 2018. M. Son provided valuable comments on the preparation of the specimen. We are also grateful to the editor A. Balanoff and senior editor M. D'Emic as well as three anonymous reviewers for their comments that improved the quality of this manuscript. This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF), funded by the Ministry of Education (grant number 2022R1I1A2060919) to Y.-N.L.
DISCLOSURE STATEMENT
No potential conflict of interest was reported by the author(s).
DATA AVAILABILITY STATEMENT
The data that support the conclusions of this study are available in the manuscript and its supplementary files.
Correction Statement
This article has been corrected with minor changes. These changes do not impact the academic content of the article.
LITERATURE CITED
- Barsbold, R. (1974). Saurornithoididae, a new family of small theropod dinosaurs from Central Asia and North America. Palaeontologia Polonica, 30, 5–22.
- Cau, A., & Madzia, D. (2021). The phylogenetic affinities and morphological peculiarities of the bird-like dinosaur Borogovia gracilicrus from the Upper Cretaceous of Mongolia. PeerJ, 9, e12640. https://doi.org/10.7717/peerj.12640
- Chiappe, L. M., Norell, M. A., & Clark, J. M. (2001). A new skull of Gobipteryx minuta (Aves; Enantiornithes) from the Cretaceous of the Gobi Desert. American Museum Novitates, 2001(3346), 1–15. https://doi.org/10.1206/0003-0082(2001)346<0001:ANSOGM>2.0.CO;2
- Currie, P. J., & Peng, J.-H. (1993). A juvenile specimen of Saurornithoides mongoliensis from the Upper Cretaceous of northern China. Canadian Journal of Earth Sciences, 30(10), 2224–2230. https://doi.org/10.1139/e93-193
- Eberth, D. A. (2018). Stratigraphy and paleoenvironmental evolution of the dinosaur-rich Baruungoyot-Nemegt succession (Upper Cretaceous), Nemegt Basin, southern Mongolia. Palaeogeography, Palaeoclimatology, Palaeoecology, 494, 29–50. https://doi.org/10.1016/j.palaeo.2017.11.018
- Fanti, F., Currie, P. J., & Badamgarav, D. (2012). New specimens of Nemegtomaia from the Baruungoyot and Nemegt Formations (Late Cretaceous) of Mongolia. PLoS ONE, 7(2), e31330. https://doi.org/10.1371/journal.pone.0031330
- Gilmore, C. W. (1924). On Troodon validus, an orthopodous dinosaur from the Belly River Cretaceous of Alberta, Canada. Bulletin of the Department of Geology, University of Alberta, 1, 1–43.
- Goloboff, P. A., & Morales, M. E. (2023). TNT version 1.6, with a graphical interface for MacOS and Linux, including new routines in parallel. Cladistics, 39(2), 144–153. https://doi.org/10.1111/cla.12524
- Gradziński, R., Kielan-Jaworowska, Z., & Maryańska, T. (1977). Upper Cretaceous Djadokhta, Barun Goyot and Nemegt formations of Mongolia, including remarks on previous subdivisions. Acta Geologica Polonica, 27(3), 281–318.
- Hartman, S., Mortimer, M., Wahl, W. R., Lomax, D. R., Lippincott, J., & Lovelace, D. M. (2019). A new paravian dinosaur from the Late Jurassic of North America supports a late acquisition of avian flight. PeerJ, 7, e7247. https://doi.org/10.7717/peerj.7247
- HoltzJr.T. R. (1995). The arctometatarsalian pes, an unusual structure of the metatarsus of Cretaceous Theropoda (Dinosauria: Saurischia). Journal of Vertebrate Paleontology, 14(4), 480–519. doi:10.1080/02724634.1995.10011574
- Jerzykiewicz, T., Currie, P. J., Fanti, F., & Lefeld, J. (2021). Lithobiotopes of the Nemegt Gobi Basin. Canadian Journal of Earth Sciences, 58(9), 829–851. https://doi.org/10.1139/cjes-2020-0148
- Kurzanov, S. M., & Osmólska, H. (1991). Tochisaurus nemegtensis gen. et sp. n., a new troodontid (Dinosauria, Theropoda) from Mongolia. Acta Palaeontologica Polonica, 36(1), 69–76.
- Makovicky, P. J., & Norell, M. A. (2004). Troodontidae. In D. B. Weishampel, P. Dodson, & H. Osmólska (Eds.), The Dinosauria (2nd ed (pp. 184–195). University of California Press.
- Makovicky, P. J., Norell, M. A., Clark, J. M., & Rowe, T. (2003). Osteology and relationships of Byronosaurus jaffei (Theropoda: Troodontidae). American Museum Novitates, 2003(3402), 1–32. https://doi.org/10.1206/0003-0082(2003)402<0001:OAROBJ>2.0.CO;2
- Marsh, O. C. (1881). Classification of the Dinosauria. American Journal of Science–Third Series, 23(133), 81–86.
- Norell, M. A., Makovicky, P. J., Bever, G., Balanoff, A., Clark, J. M., Barsbold, R., & Rowe, T. (2009). A review of the Mongolian Cretaceous dinosaur Saurornithoides (Troodontidae, Theropoda). American Museum Novitates, 2009(3654), 1–63. https://doi.org/10.1206/648.1
- Norell, M. A., Makovicky, P. J., & Clark, J. M. (2000). A new troodontid theropod from Ukhaa Tolgod, Mongolia. Journal of Vertebrate Paleontology, 20(1), 7–11. https://doi.org/10.1671/0272-4634(2000)020[0007:ANTTFU]2.0.CO;2
- Osborn, H. F. (1924). Three new Theropoda, Protoceratops zone, central Mongolia. American Museum Novitates, 144, 1–12.
- Osmólska, H. (1987). Borogovia gracilicrus gen. et. sp. n., a new troodontid dinosaur from the Late Cretaceous of Mongolia. Acta Palaeontologica Polonica, 32(1–2), 133–150.
- Owen, R. (1842). Report on British fossil reptiles. Part II. Report for the British Association for the Advancement of Science, 1841, 60–204.
- Pei, R., Pittman, M., Goloboff, P. A., Dececchi, T. A., Habib, M. B., Kaye, T. G., Larsson, H. C. E., Norell, M. A., Brusatte, S. L., & Xu, X. (2020). Potential for powered flight neared by most close avialan relatives, but few crossed its thresholds. Current Biology, 30(20), 4033–4046. https://doi.org/10.1016/j.cub.2020.06.105
- Pei, R., Norell, M. A., Barta, D. E., Bever, G. S., Pittman, M., & Xu, X. (2017). Osteology of a new Late Cretaceous troodontid specimen from Ukhaa Tolgod, Ömnögovi Aimag, Mongolia. American Museum Novitates, 2017(3889), 1–47. https://doi.org/10.1206/3889.1
- Pei, R., Qin, Y., Wen, A., Zhao, Q., Wang, Z., Liu, Z., Guo, W., Liu, P., Ye, W., Wang, L., Yin, Z., Dai, R., & Xu, X. (2022). A new troodontid from the Upper Cretaceous Gobi Basin of inner Mongolia, China. Cretaceous Research, 130, 105052. https://doi.org/10.1016/j.cretres.2021.105052
- Russell, D. A., & Dong, Z.-M. (1993). A nearly complete skeleton of a new troodontid dinosaur from the Early Cretaceous of the Ordos Basin, Inner Mongolia, People’s Republic of China. Canadian Journal of Earth Sciences, 30(10), 2163–2173. https://doi.org/10.1139/e93-187
- Shen, C.-Z., Zhao, B., Gao, C.-L., Lü, J., & Kundrát, M. (2017). A new troodontid dinosaur (Liaoningvenator curriei gen. et sp. nov.) from the Early Cretaceous Yixian Formation in Western Liaoning Province. Acta Geoscientica Sinica, 38(3), 359–371. https://doi.org/10.3975/cagsb.2017.03.06
- Snively, E., Russell, A. P., & Powell, G. L. (2004). Evolutionary morphology of the coelurosaurian arctometatarsus, descriptive, morphometric and phylogenetic approaches. Zoological Journal of the Linnean Society, 142(4), 525–553. https://doi.org/10.1111/j.1096-3642.2004.00137.x
- Tsuihiji, T., Barsbold, R., Watabe, M., Tsogtbaatar, K., Chinzorig, T., Fujiyama, Y., & Suzuki, S. (2014). An exquisitely preserved troodontid theropod with new information on the palatal structure from the Upper Cretaceous of Mongolia. Naturwissenschaften, 101(2), 131–142. https://doi.org/10.1007/s00114-014-1143-9
- Wang, S., Zhang, Q., Tan, Q., Jiangzuo, Q., Zhang, H., & Tan, L. (2022). New troodontid theropod specimen from Inner Mongolia, China clarifies phylogenetic relationships of later-diverging small-bodied troodontids and paravian body size evolution. Cladistics, 38(1), 59–82. https://doi.org/10.1111/cla.12467
- Xu, X., Tan, Q., Sullivan, C., Han, F., & Xiao, D. (2011). A short-armed troodontid dinosaur from the Upper Cretaceous of Inner Mongolia and its implications for troodontid evolution. PLoS ONE, 6(9), e22916. https://doi.org/10.1371/journal.pone.0022916
- Xu, X., Zhao, Q., Sullivan, C., Tan, Q.-W., Sander, M., & Ma, Q.-Y. (2012). The taxonomy of the troodontid IVPP V 10597 reconsidered. Vertebrata PalAsiatica, 50(2), 140–150.
- Zanno, L., Varricchio, D. J., O’Connor, P. M., Titus, A. L., & Knell, M. J. (2011). A new troodontid theropod, Talos sampsoni gen. et sp. nov., from the Upper Cretaceous Western Interior Basin of North America. PLoS ONE, 6(9), e24487. https://doi.org/10.1371/journal.pone.0024487