
ABSTRACT
The lungfish, the extant sister group of the tetrapods, have an evolutionary history illustrated by a fossil record extending for ∼420 million years. The post-Paleozoic fossil record of the clade is almost exclusively restricted to sediments deposited in freshwater paleoenvironments and is characterized by an abundance of highly mineralized tooth plates, whereas cranial and postcranial remains are scarce. Here, we report a sample of isolated tooth plates found in the Upper Triassic Pebbly Arkose Formation of the Mid-Zambezi Basin, Zimbabwe. It consists of pterygoid and prearticular tooth plates from adult individuals, plus some dental plates referred to juvenile individuals, which we refer to a new species of Ferganoceratodus. This discovery provides an opportunity to review briefly the tooth plates of the ‘ptychoceratodontid morphotype’ reported from around the world. We discuss how various occurrences previously referred to Ptychoceratodus may be more appropriately referred, with caution, to Ferganoceratodus. We also describe the histology of the tooth plates of the new species and note similarities with other Mesozoic taxa. The scarcity of histological data for Mesozoic lungfish tooth plates compounds the problem of assigning isolated tooth plates to genus and species level. Ferganoceratodus and closely related taxa arose in the Early Triassic in southern Gondwana and diversified worldwide in the Late Triassic. The genus then became more common in Laurasia during the Jurassic and Early Cretaceous and declined thereafter with relict Late Cretaceous occurrences in Madagascar and South America.
INTRODUCTION
The distribution of lungfish is now restricted to the former landmasses of Gondwana, i.e., Australia with Neoceratodus, Africa with Protopterus, and South America with Lepidosiren (Brownstein et al., Citation2023). The latter two are sister genera and result from a vicariant event associated with the opening of the South Atlantic (Cavin et al., Citation2007). The Neoceratodus lineage split is more obscure, and the inclusion of extant taxa in the evolutionary framework is still unable to provide a clear picture of the phylogenetic relationships of post-Paleozoic lungfishes, including extant lineages (e.g., Schultze, Citation2004; Cavin et al., Citation2007).
The Triassic Period was a time of both change and stasis with respect to lungfish evolution (Schultze, Citation2004). Although no substantial changes in gross tooth plate morphology and phylogenetic diversity were observed in Triassic lungfishes (Kemp et al., Citation2017), this is the time when we first see changes in lungfish behavior, with abundant evidence for burrowing and aestivation as exemplified by the preservation of numerous burrows in the Chinle Formation of the southwestern U.S.A. (Dubiel et al., Citation1988; Gobetz et al., Citation2006).
Early researchers included Triassic lungfish species in the ‘wastebasket’ genus Ceratodus (Agassiz Citation1835) but many of these specimens have now been referred to a variety of genera and many species. However, the post-Paleozoic lungfish fossil record, which is almost exclusively restricted to freshwater paleoenvironments, consists mostly of isolated tooth plates (Kemp et al., Citation2017). Although varying in size and shape between species, lungfish tooth plates exhibit characteristics that vary during ontogeny, and many of these characteristics are easily erased by wear during life or by the process of fossilization, making it difficult to identify characters for phylogenetic analyses.
Here, we report new lungfish tooth plate material from the Pebbly Arkose Formation of the Mid-Zambezi Basin, Zimbabwe. This provides only the second record of a fossil lungfish from Zimbabwe and one of a handful of such occurrences from the Triassic of southern and eastern Africa (from Angola, South Africa, Tanzania, and Zambia), most of which have been mentioned in faunal lists without detailed description or illustration (e.g., Antunes et al., Citation1990; Bender & Hancox, Citation2004; Broom, Citation1913; Crompton, Citation1955; Jubb & Gardiner, Citation1975; Raath et al., Citation1992). Lungfishes from penecontemporaneous units in the main Karoo Basin of South Africa, from the Burgersdorp Formation and Stormberg Group, are either taxonomically indeterminate (e.g., Kitching & Raath, Citation1984) or have been referred to two of the four Mesozoic dipnoan families, Ceratodontidae and Ptychoceratodontidae (Bender & Hancox, Citation2004). Our new material provides a valuable new datum for understanding the distribution and morphology of Triassic Gondwanan lungfish.
GEOLOGIC AND FAUNAL SETTING
The material described herein was discovered at a field site nicknamed ‘The Dentist’ (16°48′33.79″S, 28°28′5.71″E) that is situated on the southern shoreline of Lake Kariba, within the boundaries of Matusadona National Park, approximately 50 km west of Kariba, Zimbabwe. For detailed site descriptions, maps, and stratigraphic logs see Barrett et al. (Citation2020) and Sciscio et al. (Citation2021b). It was found in the Pebbly Arkose Formation (also referred to, in error, as the Tashinga Formation; Barrett et al., Citation2020), which forms part of the Triassic–Lower Jurassic aged upper Karoo Group in the Mid-Zambezi Basin (e.g., Ait-Kaci Ahmed, Citation2018; Bond & Falcon, Citation1973; Sciscio et al., Citation2021b). In the Mid-Zambezi Basin, the Pebbly Arkose Formation reaches thicknesses of 79–137 m (Bond & Falcon, Citation1973) and consists of fine- and coarse-grained sandstone units with interbedded conglomerates and paleosol horizons (Sciscio et al., Citation2021b). At ‘The Dentist’ site, the fossiliferous sediments consist of a pedogenically overprinted red to grey muddy-siltstone, which includes rhizocretions, coprolites, small pedogenic nodules, sandstone-filled desiccation cracks (Barrett et al., Citation2020; Sciscio et al., Citation2021b), and trace fossils (Sciscio et al., Citation2021a). Several very fine to fine-grained, thin, and discontinuous sandstone layers occur within the siltstone. Historically, the Pebbly Arkose Formation within the Mid-Zambezi Basin was considered Upper Triassic in age on the basis of paleobotanical evidence, due to the occurrence of several fossil tree taxa (e.g., Dadoxylon, Rhexoxylon) known also from the more securely dated Elliot Formation of the main Karoo Basin (Bond, Citation1972; Bond & Falcon, Citation1973). An Upper Triassic age for the Pebbly Arkose Formation in the Kariba area has been supported by a U-Pb date of 209.2 ± 4.5 Ma (Norian) derived from detrital zircons (Barrett et al., Citation2020). However, it should be noted that the Pebbly Arkose Formation has been mapped in other Karoo-aged basins in the region, notably the Cabora Bassa Basin, where biostratigraphic evidence suggests an earlier—?late Carnian—age (Griffin et al., Citation2022; Raath, Citation1996). It seems likely that the formation was deposited diachronously, with its lithological similarities allowing it to be mapped over a considerable distance (Griffin et al., Citation2022).
The Pebbly Arkose Formation of the Mid-Zambezi Basin has yielded a mixed freshwater/terrestrial assemblage, including temnospondyls, phytosaurs, and theropod and sauropodomorph dinosaurs. To date, only freshwater taxa have been recovered from ‘The Dentist’ site, suggesting local-scale environmental heterogeneity (Barrett et al., Citation2020; Sciscio et al., Citation2021b). Lungfish tooth plates were reported previously from the Pebbly Arkose Formation of an island in the estuary of the Bumi River (Raath et al., Citation1992) but were not described or figured. These specimens were relocated in the NHMZ and consist of nine fragments (QG 142 and NHMB 11685) that are too incomplete to refer to the new species described herein, or any other taxon, and are considered Dipnoi indet. herein (see Supplementary information).
MATERIALS AND METHODS
Two petrological thin sections were made of NHMZ 2432(8) in vertical planes to a thickness of 30 μm. One vertical section passes through the length of the fourth tooth ridge while the second section passes through the heel of the inner angle of the tooth plate. Petrological thin sections were studied using a Leica DMLP reflected/transmitted light polarizing microscope with a DFC 420C camera with 10× and 40× objective lenses and images captured using Leica Application Suite v. 4.00. Micro-CT sections were also made through NHMZ 2432(8) for digital histology at the Natural History Museum, London using a Nikon Metrology HMX ST 225 micro-CT scanner with the following parameters: 150 kV; 67 μA; 0.5 mm copper filter; x-offset = 0.000; y-offset = −3.951; z-offset = 0.007; source to object = 80.806 mm; source to detector = 1173.8 mm; and mask radius = 13.573, 3142 projections with an angular step of 0.115°. Projections were constructed using the CTPro package with Beam hardening type = simple. Voxel size is 0.014 mm in X, Y, and Z. Reconstructed thin sections were viewed in Fiji image analysis software (Schindelin et al., Citation2012). All reconstructed micro-CT data can be found in the Morphosource online repository at https://www.morphosource.org, project ID: 000547763.
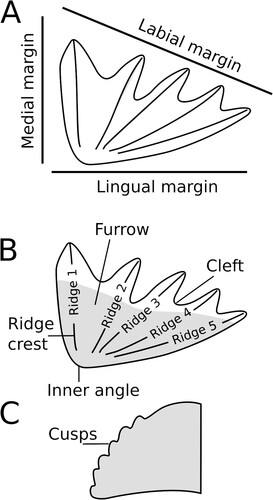
Morphological terminology—Following Romer and Smith (Citation1934) we refer to the bone that carries the upper tooth plate as the pterygoid and that carrying the lower tooth plate the prearticular. The long, narrow crests emanating laterally from the occlusal surface are referred to herein as tooth ridges following the terminology of Ahlberg et al. (2007). Tooth rows are defined as individual aligned dentine cusp elements (Ahlberg et al., 2007). Denticulations are numerous small toothlets/odontodes scattered over dermal bone surfaces in the mouth and pharynx (Ahlberg et al., 2007), whereas we follow the terminology of Kemp (Citation2002) in using the term cusps for small projections fused on the labial edge of the tooth ridges ().
FIGURE 1. Tooth plate terminology used in this paper. A, orientation of medial, labial, and lingual margins of a tooth plate. B, terminology of parts of a tooth plate. Gray area represents approximate region of occlusal surface. C, lateral view of the end (at the labial edge) of a single tooth ridge showing unworn cusps.
Institutional abbreviations—MPEF, Museo Paleontológico Egidio Feruglio, Chubut, Argentina; NHMZ, Natural History Museum of Zimbabwe, Bulawayo, Zimbabwe (formerly QG, The Queen Victoria Museum, Salisbury [now Harare] and NHMB, Natural History Museum of Bulawayo, no longer used). See Supplemental Information for further explanation of the history of these institutional names.
SYSTEMATIC PALEONTOLOGY
Subclass SARCOPTERYGII Romer, Citation1955
Order DIPNOI Müller, Citation1845
Suborder CERATODONTOIDEI Nikolski, Citation1954 sensu Kemp et al., Citation2017
Family incertae sedis
Genus FERGANOCERATODUS Nessov and Kaznyshkin, Citation1985
Diagnosis (after Cavin et al., Citation2007)—Skull roof with two unpaired bones in the median series, two paired bones on the mediolateral series, and two bones on the lateral series; bones of the median series suture through a short contact; dental plates of cutting type, with five radiating ridges on the pterygoid plates and four radiating ridges on the prearticular plates separated by deep notches.
FERGANOCERATODUS EDWARDSI sp. nov.
()
FIGURE 2. Ferganoceratodus edwardsi. A-E, upper tooth plates; F-J, lower tooth plates. A-B, NHMZ 2432(9), holotype in labial and occlusal views. Arrowhead indicates the surface for a potential symphyseal contact; C, NHMZ 2432(6); D, NHMZ 2432(13); E, NHMZ 2432(5); F, NHMZ 2432(1), juvenile; G, NHMZ 2432(2), juvenile; H, NHMZ 2432(12); I, NMHZ 2432(11); J, NHMZ 2432(14); K, NHMZ 2432(4), juvenile. Scale bars equal 10 mm.
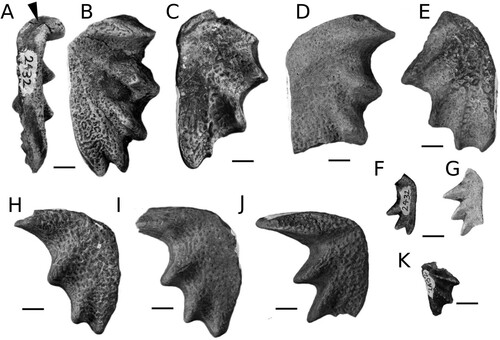
FIGURE 3. Petrological micrographs of Ferganoceratodus edwardsi (NHMZ 2432[8], prearticular tooth plate). A, plane-polarized light (PPL) coronal section through heel of tooth plate showing pulp canals and dentinal tubules below the occlusal surface surrounded by circumdenteonal dentine. Interdenteonal dentine is present between denteons and a thin layer of probable marginal dentine lies just below the occlusal surface; B, section in ‘A’ in crossed-polarized light (XPL) showing inclined orientation of interdenteonal dentine crystals and monorefringent layer of marginal dentine below occlusal surface; C, median section below occlusal surface in PPL through fourth tooth ridge showing inverted ‘cones’ predominantly of circumdenteonal dentine with branching denteons and regions of interdenteonal dentine between cones; D, ‘C’ in XPL; E, ridge of harder interdenteonal dentine with softer ‘cones’ of circumdenteonal dentine on either side. The denteons in the interdenteonal dentine converge towards the occlusal surface; F, ‘D’ in XPL; G, median section through end of tooth ridge showing thin layer of enamel on surface of tooth ridge and complex association of layers of monorefringent and birefringent interdenteonal dentine among a cone of circumdenteonal dentine. Note how denteons are subparallel to the surface of the tooth ridge below the region covered with enamel; H, XPL of ‘G’ overlain with transparency of ‘G’ to highlight relationship between birefringent interdenteonal dentine and regions of darker dentine seen in PPL. Abbreviations: cd, circumdenteonal dentine; e, enamel; id, interdenteonal dentine; md, marginal dentine; t, dentinal tubules; pc, pulp canal. Scale bars for A, B, E, F equal 500 μm. Scale for C, D, G, H equal 1 mm.
![FIGURE 3. Petrological micrographs of Ferganoceratodus edwardsi (NHMZ 2432[8], prearticular tooth plate). A, plane-polarized light (PPL) coronal section through heel of tooth plate showing pulp canals and dentinal tubules below the occlusal surface surrounded by circumdenteonal dentine. Interdenteonal dentine is present between denteons and a thin layer of probable marginal dentine lies just below the occlusal surface; B, section in ‘A’ in crossed-polarized light (XPL) showing inclined orientation of interdenteonal dentine crystals and monorefringent layer of marginal dentine below occlusal surface; C, median section below occlusal surface in PPL through fourth tooth ridge showing inverted ‘cones’ predominantly of circumdenteonal dentine with branching denteons and regions of interdenteonal dentine between cones; D, ‘C’ in XPL; E, ridge of harder interdenteonal dentine with softer ‘cones’ of circumdenteonal dentine on either side. The denteons in the interdenteonal dentine converge towards the occlusal surface; F, ‘D’ in XPL; G, median section through end of tooth ridge showing thin layer of enamel on surface of tooth ridge and complex association of layers of monorefringent and birefringent interdenteonal dentine among a cone of circumdenteonal dentine. Note how denteons are subparallel to the surface of the tooth ridge below the region covered with enamel; H, XPL of ‘G’ overlain with transparency of ‘G’ to highlight relationship between birefringent interdenteonal dentine and regions of darker dentine seen in PPL. Abbreviations: cd, circumdenteonal dentine; e, enamel; id, interdenteonal dentine; md, marginal dentine; t, dentinal tubules; pc, pulp canal. Scale bars for A, B, E, F equal 500 μm. Scale for C, D, G, H equal 1 mm.](/cms/asset/4c1537c2-6b98-4362-a722-3c256fe2336a/ujvp_a_2365391_f0003_oc.jpg)
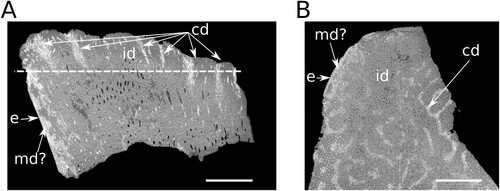
FIGURE 4. A, vertical micro-CT section though the heel of prearticular tooth plate NHMZ 2432(8). Medial margin is to the left showing high X-ray opacity circumdenteonal dentine and a thin layer of enamel on the surface. ‘Cones’ of higher X-ray opacity with well-developed circumdenteonal dentine coincide with wear pits in the occlusal surface (top) while the main body of the pulp canal is composed of interdenteonal dentine with denteons dispersed throughout. Line represents section in ‘B’; B, horizontal micro-CT section through NHMZ 2432(8) showing regions of pits, and depressions corresponding to circumdenteonal dentine (high X-ray opacity). Abbreviations: cd, circumdenteonal dentine; e, enamel; id, interdenteonal dentine; md, marginal dentine. Scale bars equal 2 mm.
Holotype—NHMZ 2432(9). A pterygoid tooth plate separated from the pterygoid.
Etymology—Named in honor of Mr. Steve Edwards, discoverer of ‘The Dentists’ and other Late Triassic field sites in Matusadona National Park, who collected the specimens described herein.
Referred Specimens—Thirteen isolated tooth plates consisting of: pterygoid tooth plates, NHMZ 2432(5), NHMZ 2432(6), NHMZ 2432(7), NHMZ 2432(8), NHMZ 2432(8), NHMZ 2432(13); prearticular tooth plates, NHMZ 2432(11), NHMZ 2432(12), NHMZ 2432(14); pterygoid tooth plates of juvenile individuals; NHMZ 2432(1), NHMZ 2432(2), NHMZ 2432(3); and a prearticular tooth plate of juvenile individual NHMZ 2432(4).
Diagnosis—Ferganoceratodus edwardsi is diagnosed by the following combination of characters: tooth plates with concave pterygoid tooth plate occlusal surface (“plateau area” of Smith and Campbell [Citation1987] and Panzeri [Citation2020, Citation2022]) and convex prearticular tooth plate occlusal surface; occlusal pits present on both the occlusal surface and on inter-ridge furrows; pterygoid tooth plates possess five tooth ridges, prearticular tooth plates have four tooth ridges; pterygoid and prearticular tooth plates have a posteriorly curved first ridge with the pterygoid tooth plate first ridge displaying an abrupt angle whereby the distal portion of the ridge turns posteriorly; all ridges from the pterygoid tooth plates of approximately similar lengths; first ridge of the prearticular tooth plates significantly longer than the following second and third ridges, fourth ridge poorly developed.
Locality, Horizon, and Age—‘The Dentist’ site, Matusadona National Park, southern shore of Lake Kariba, Mid-Zambezi Basin, Zimbabwe; Pebbly Arkose Formation (Upper Triassic: Norian; see Barrett et al. [Citation2020]; Sciscio et al. [Citation2021a]).
Notes—F. edwardsi is reminiscent of F. acutus, which is probably from the Upper Triassic of Morafenobe, Madagascar. This species was described by Priem (Citation1924) based on a single tooth plate considered as either a pterygoid tooth plate (Martin, Citation1981) or from an undetermined position (Martin et al., Citation1999). Here, we consider the tooth plate described by Priem (Citation1924) to be a prearticular one, and we associate it with pterygoid tooth plates that are probably from the same locality and were previously associated with F. cf. hislopianus (Martin, Citation1981; Martin et al., Citation1999; see below). It is possible that future work will indicate that F. edwardsi is synonymous with F. acutus, but the absence of the first ridge in the holotype of F. acutus, and in the two pterygoid dental plates mentioned by Martin (Citation1981) and Martin et al. (Citation1999), due to breakage, makes comparisons difficult. A revision of the Malagasy material is required to solve this taxonomic problem.
DESCRIPTION
The upper (pterygoid) tooth plates have five blunt ridges oriented towards the mediolingual junction, with the fifth ridge forming the lingual margin (). The inner angle (alpha) measures circa 115°. The ridges are approximately straight. The inter-ridge furrows are wide, decrease in width posteriorly, and are quite deep, especially in the smallest specimens. A distinct buttress is present along the lingual ridge. It is difficult to discern if a symphyseal contact was present along the mesial edge, but a short contact seems to have been present on the holotype at least (, arrowhead).
The prearticular tooth plates have four ridges, with the first one by far the longest and the fourth one shorter than the second and third. The ridges are blunt and oriented towards the mediolingual junction, but the fourth ridge does not form the lingual margin. The inner angle (alpha) measures circa 80°. The mesial and lingual margins are slightly convex in dorsal view. The shape of the mesial margin indicates that the tooth plates were likely not in contact with each other. The inter-ridge furrows are wide, decreasing in width posteriorly, but are not especially deep even in small (young) individuals.
Four smaller, incomplete tooth plates belong to juvenile specimens. Three (2432[1], 2432[2], 2432[3]) are pterygoid tooth plates. Numerous small, densely packed rounded pits are present on their occlusal surface and in the inter-ridge furrows. These form larger shallower excavations or a reticulated pattern of shallow grooves in larger-sized teeth due to wear. The inter-ridge furrows are deeper than on the large plates. On a small prearticular tooth plate (2432[4]), a series of aligned cusps extends along the ridge. All the features distinguishing adult tooth plates from juveniles—shape and location of the pits, depth of the inter-ridge furrows, presence of cusps—have been previously identified as characters of juvenile specimens in populations of European Triassic lungfishes (Skrzycki, Citation2015).
Tooth plate histology
Petrographic thin-sections and micro-CT scan data allow the tooth plate histology of Ferganoceratodus edwardsi to be investigated. We employ the histological terminology of Kemp (Citation2001, Citation2002) herein. In plane-polarized light, features typical of Mesozoic dipnoan tooth plates are clearly seen. Denteons (pulp cavities plus circumdenteonal dentine) are pervasive throughout the tooth plate in both vertical and horizontal sections from which fine dentinal tubules project (). Circumdenteonal dentine surrounding pulp cavities is finely crystalline (). Between denteons the tooth plate is composed of crystalline interdenteonal dentine, which forms the major part of the tooth plate. Darker areas of interdenteonal dentine are visible between denteons in plane-polarized light in vertical section corresponding to areas of monorefringent interdenteonal dentine crystals in crossed-polarized light.
The pitted texture of the occlusal surface of the tooth plate results from the distributions of different dentine tissues (Kemp, Citation1993). Deeper convex pits represent regions composed primarily of closely spaced denteons with extensively developed circumdenteonal dentine () where branching denteons are densely clustered into cones. The arrangement of denteons in interdenteonal regions varies depending on the vertical position in the tooth plate. Towards the pulp canal they are perpendicular to the lingual margin of the tooth plate. In the central region of the tooth plate, the denteons are primarily vertical, and as they approach the occlusal surface they converge towards the harder-wearing ridges in the manner of a stacked cone (). At the labial edge below the occlusal surface of the tooth ridge the denteons are arranged subparallel to the occlusal surface of the tooth ridge below a layer of enamel on the surface ().
Micro-CT scan data from NHMZ 2432(8) reveal a high density of denteons running vertically throughout the tooth plate, as is typical of Mesozoic dipnoan tooth plates (). Pits in the occlusal surface of the tooth plate correspond to regions of higher X-ray opacity, densely packed denteons with well-developed circumdenteonal dentine while ridges correspond to lower X-ray opacity interdenteonal dentine (). The regions of higher opacity do not penetrate to the base of the tooth plate. A layer of higher X-ray opacity is also present along the lingual margin of the tooth plate which is, in turn, covered in a thin layer of enamel. The high opacity layer beneath the enamel is not identified as petrodentine as it still contains denteons. No petrodentine (sensu Lison, Citation1941) is observed in either the thin-sections or micro-CT data.
DISCUSSION
Histology
The arrangement of dentine tissues seen in Ferganoceratodus edwardsi exhibits similarities with other Mesozoic lungfish tooth plates. The structure of wear pits being associated with regions of circumdenteonal dentine is also seen in Ceratodus latissimus, but in C. pectinatus tooth ridges composed of interdenteonal dentine stand proud of troughs of circumdenteonal dentine rather than individual pits (Kemp, Citation2001). Further similarities are shared with the Late Cretaceous taxon Metaceratodus baibianorum (Panzeri et al., Citation2022). In particular, Panzeri et al. (Citation2022) noted regions of darker interdenteonal dentine in thin-section plane-polarized light which, when viewed in crossed polarized light, appears monorefringent. This is not a feature of interdenteonal dentine noted by Kemp (Citation2001) or Kemp and Barry (Citation2006) but has been noted by Smith (Citation1984) in Neoceratodus. The significance of the similarity in dentine tissues between Ferganoceratodus edwardsi and Metaceratodus baibianorum is uncertain. The lack of thin-section analyses of post-Paleozoic lungfish tooth plates does not allow easy distinctions to be made between phylogenetic relationships in dipnoan dental histology or convergence in Mesozoic dipnoan dental tissues. The greater degree of mineralization seen in Mesozoic dipnoan tooth plates compared with Paleozoic forms has been noted previously (Reisz et al., Citation2004) but the only histological character in recent phylogenetic analysis of Dipnoi (Kemp et al., Citation2017) is the presence/absence of petrodentine. Whereas morphological features of lungfish tooth plates can be unreliable indicators of phylogenetic relationships, further study of the dental histology of post-Paleozoic taxa might provide further characters that can delineate evolutionary changes within the group (Kemp, Citation2001). With no thin-section data available for Ptychoceratodus, besides the absence of petrodentine, it is not possible to state whether or not there are histological differences between this taxon and Ferganoceratodus that could be used to distinguish the isolated tooth plates of these genera.
Ptychoceratodus versus Ferganoceratodus
Taxonomic recognition of lungfish genera and species based solely on the characters provided by isolated dental plates is a difficult task that has been undertaken by some authors (e.g., Skrzycki, Citation2015) but rejected by others (e.g., Schultze, Citation1981) who consider dental plates as unreliable for taxonomy. We consider that it is possible to distinguish between species since each tooth plate population shares a combination of generally unique characters. However, we consider genera based solely on tooth plates to be doubtful and prefer to refer this kind of material to genera known from both cranial and dental material.
We focus our comparisons on larger tooth plates referred to adult specimens. Based on isolated tooth plates alone, two genera share similar characteristics, Ptychoceratodus Jaekel, Citation1926 and Ferganoceratodus Kaznyshkin and Nessov (in Nessov & Kaznyshkin, Citation1985). In his comparisons of dental characters, Skrzycki (Citation2015:table 5) listed no differences between these two genera in the main characteristics of their pterygoid and prearticular plates. Moreover, several species have been referred to each genus, widening the range of diagnostic characters and causing systematic confusion. For example, some species referred to Ptychoceratodus have the same number of crests on their tooth plates as Ferganoceratodus while morphometric data demonstrate that only Ptychoceratodus serratus consistently forms an isolated population (Skrzycki, Citation2015:figs. 6, 7). Therefore, we focus our discussion on the type species of each genus, namely Ptychoceratodus serratus (Agassiz, 1838) and Ferganoceratodus jurassicus Kaznyshkin and Nessov (in Nessov & Kaznyshkin, Citation1985).
P. serratus was based on a Late Triassic specimen from Switzerland consisting of a pterygoid tooth plate with six ridges separated by deep furrows (). This species was later identified in connection with skull bones found in the Ladinian (upper Middle Triassic) of Kupferzell-Bauersbach, Germany (Kemp, Citation1998; Schultze, Citation1981; Skrzycki, Citation2015) (). In this specimen, the pterygoid and prearticular tooth plates have six and five ridges, respectively. F. jurassicus, originally described from the Middle Jurassic of Kyrgyzstan, is known from various isolated elements of the skull roof associated with tooth plates with five ridges on the pterygoid and four on the prearticular tooth plates (; Nessov & Kaznyshkin, Citation1985).
FIGURE 5. Comparison of Ferganoceratodus erdwardsi tooth plates with the type species of Ptychoceratodus, P. serratus. A, holotype of P. serratus (redrawn from Agassiz, Citation1833–43:vol. 3, pl. 19, fig. 18). B, material referred to P. serratus by Schultze (Citation1981). C, the new material of F. edwardsi. D, the type series of Ferganoceratodus, F. jurassicus (redrawn from Nessov & Kaznyshkin, Citation1985). E, type specimen of F. phillipsi upper row, pterygoid tooth plates; lower row, prearticular tooth plates. Scale bars equal 10 mm.
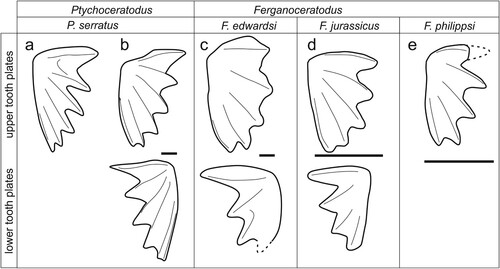
It should be noted that material referred to Ptychoceratodus serratus and the type specimen of Ferganoceratodus jurassicus include their calvaria. In the material referred to by Schultze (Citation1981) as P. serratus, as in F. jurassicus, the skull roof is almost square in shape and characterized by few ossifications, some of which are shared by both species, such as the paired anterior mediolateral bones (KLMN in Schultze, Citation1981; KLMN + 3 in Kemp, Citation1998), and the posterior mediolateral bones (bone I or IJ in previous studies). The main difference is in the medial series, with a single anterior bone in both (the E bone) and two posterior bones in P. serratus (C in the center and AB behind) and a single bone (the ABC bone) occupying the same position in F. jurassicus (ABC bone). Additionally, in P. serratus a small bone is wedged between the posterior medial series (AB) and the anterior and posterior medial bones (KLMN and I), without reaching the lateral margin of the series (the J bone), while this bone is absent (or considered fused to I) in Ferganoceratodus (Cavin et al., Citation2007; Nessov & Kaznyshkin, Citation1985).
Based on these comparisons, we refer to Ptychoceratodus those species whose pterygoid tooth plates have six ridges and prearticular tooth plates have five ridges and that are associated with a medial skull roof series consisting of three bones. We refer to Ferganoceratodus those species whose pterygoid tooth plates have five ridges and prearticular tooth plates have four ridges and that are associated with a medial calvarium series consisting of two bones. Mesozoic species based on tooth plate material have been assigned to genera other than Ptychoceratodus and Ferganoceratodus although they exhibit the general characteristics present in both taxa, particularly the number of ridges present in the pterygoid and prearticular plates. In the absence of cranial material associated with this dental material, in the following discussion we provisionally and cautiously refer these species to one of these two genera pending the discovery of more complete material.
Compared with these two type species, our material is closer to F. jurassicus, with five and four ridges in the pterygoid and prearticular tooth plates, respectively (compare with ). Several species previously referred to Ptychoceratodus, however, have the same number of crests as Ferganoceratodus (see below for discussion).
Gondwanan ‘ptychoceratodonts’
‘Ptychoceratodont’ occurrences that are geographically or stratigraphically close to F. edwardsi are known from the Early Triassic of South Africa, Australia, and Tasmania (Kemp, Citation1996), the Upper Triassic of South America (Agnolin et al., Citation2017; Richter & Toledo, Citation2008), and the Late Triassic of Madagascar and India (Martin, Citation1982a; Martin et al., Citation1999).
P. phillipsi (Agassiz, 1838) is a species originally described from the Middle Jurassic oolitic limestones of Stonesfield, Oxfordshire, U.K., and was described based on an isolated pterygoid tooth plate bearing five ridges. The type material was considered lost by Woodward (Citation1890) and subsequent authors, but is preserved in the collection of the Natural History Museum of Neuchâtel, Switzerland, as mentioned by Martin et al. (Citation1999) and Skrzycki (Citation2015) (). This species has been reported from the Carnian of Germany (Seegis, Citation1997), possibly the Carnian of Brazil (Richter & Toledo, Citation2008; as Ptychoceratodus cf. phillipsi), from the Norian of Germany (Martin et al., Citation1981), the Rhaetian of France (Martin et al., Citation1981), and the Early Triassic of South Africa and Australia (Kemp, Citation1996). Additional material from the ?Middle Triassic of Zambia was considered by Peecock et al. (Citation2017) to exhibit a high degree of similarity with Ptychoceratodus phillipsi but they were reluctant to definitively assign these specimens to Ptychoceratodus because they lacked cranial material. The specimens of Peecock et al. (Citation2017) possess five ridges on the pterygoid tooth plates and four ridges on the prearticular tooth plates with deep furrows between the tooth ridges, matching the original diagnosis of Ferganoceratodus by Nessov and Kaznyshkin (Citation1985) but not the emended diagnosis for Ptychoceratodus by Skrzycki (Citation2015). The material from Australia and South Africa is associated with incomplete cranial elements. Kemp (Citation1996) proposed a reconstruction of the skull roof, with the presence of two large bones along the mediolateral series (KLM in front and I behind), an anterior median bone (bone E), and an incomplete posterior median bone (possibly bone C). The tooth plate specimen from Australia referred to ‘P’. phillipsi has five and four ridges in the pterygoid and prearticular plates, respectively (Kemp, Citation1996: this difference is not always seen in European material referring to this species (e.g., Martin et al., Citation1981). Although the middle bone series of the calvaria in these specimens is incompletely known, the dental characters of this material justify assignment of this specimen to Ferganoceratodus, as F. phillipsi comb. nov. Kemp (Citation1996) referred other isolated tooth plates to this species, including Early Triassic fossils from near Hobart, Tasmania and Burgersdorp, South Africa. The holotype of ‘Ceratodus’ ornatus Broom, Citation1909, also from Burgersdorp, is a prearticular tooth plate bearing four ridges that was assigned to Pytchoceratodus by Martin (Citation1982) and Martin et al. (Citation1999), whereas Kemp (Citation1996) considered ‘C.’ ornatus to closely resemble P. phillipsi and so assigned the material described by Broom (Citation1913) to the latter taxon.
With the exception of a fragmentary tooth plate from the Upper Triassic of Brazil referred to F. cf. phillipsi (Richter & Toledo, Citation2008), the only other occurrence of a ‘ptychoceratodont’ from the Triassic of South America is a pterygoid right tooth plate (not lower [prearticular] left: contra Agnolin et al., Citation2017:fig. 2) from the Upper Triassic of Mendoza Province, Argentina, which was referred to a new species, Ptychoceratodus cuyanus (Agnolin et al., Citation2017). The presence of five ridges indicates affinities with the genus Ferganoceratodus: however, the very open internal angle and the first elongated ridge of this dental plate may indicate that it belongs to another genus. Nevertheless, differentiation between Ptychoceratodus and Ferganoceratodus based on the relative sharpness of tooth ridges as proposed by Agnolin et al. (Citation2017), can be discounted for reasons of differential wear, a feature that is clearly seen even intraspecifically. Tooth plates from the Cenomanian–Maastrichtian of Argentina, originally attributed to Ceratodus iheringi, were transferred with caution to Metaceratodus (Kemp, Citation1997), Ptychoceratodus (Martin, Citation1982c), Altantaceratodus (Cione et al., Citation2007), or Ameghinoceratodus (Apesteguia et al., Citation2007). Ameghinoceratodus should be considered a junior synonym of Atlantoceratodus (Agnolin, Citation2010). Atlantoceratodus is close to Ptychoceratodus and Ferganoceratodus, differing from them only by the more rounded inner angle and the thinner and more acute ridges (Apesteguia et al., 2007; Cione et al., Citation2007). Because these differences are minimal and are not associated with preserved cranial characters, and because the number of crests is similar to that of Ferganoceratodus, with five and four ridges in the pterygoid and prearticular plates, respectively, we tentatively consider this species can be referred to this genus, pending the discovery of more complete material. For the same reasons, Atlantoceratodus patagonicus from the Campanian-Maastrichtian of the Río Negro province, Argentina (Agnolin, Citation2010), is also provisionally referred to Ferganoceratodus. Metaceratodus is represented by several species in the Upper Cretaceous of Argentina (Apesteguía et al., 2007; Cione & Gouiric-Cavalli, Citation2012; Panzeri et al., Citation2020). Most of the tooth plates referred to this genus have five ridges and four ridges in the pterygoid and prearticular tooth plates, respectively, indicating that these species might belong to Ferganoceratodus. However, in Metaceratodus, the inner angle is located in a posteromedial position, regardless of the growth stage, whereas it is located in an anteromedial position in Ptychoceratodus and Ferganoceratodus (Panzeri et al., Citation2020). This difference might indicate that this Argentinean genus could be distinct from the two ‘ptychoceratodonts’.
Lungfish tooth plates referred to four species of Ceratodus from the Upper Triassic Maleri Formation of India (Oldham, Citation1859) were restricted by Martin et al. (Citation1999) to two species of Ptychoceratodus, P. hislopianus and P. virapa. The latter authors also referred pterygoid tooth plates from the Upper Triassic of the Morafenobe region, Madagascar, to P. cf. hislopianus, and a prearticular tooth plate from the same region, originally described by Priem (Citation1924) as Ceratodus acutus, to the genus Ptychoceratodus, as P. acutus. We note that the large pterygoid tooth plates of P. cf. hislopianus bear five ridges while the large prearticular tooth plate of P. acutus bears four ridges. We suggest that this material belongs to a single taxon showing the characteristics of Ferganoceratodus that is named here F. acutus nov. comb. This species is morphologically similar to F. edwardsi (see above).
The youngest Ferganoceratodus occurrence is F. madagascarensis from the Upper Cretaceous of the Mahajanga Basin in northwest Madagascar (Martin, 1982; Martin et al., Citation1999). It is worth noting that the prearticular tooth plate of this species shares with F. edwardsi an elongated first ridge with a gently convex mesial margin.
European ‘ptychoceratodonts’
Other Ptychoceratodus and Ferganoceratodus species are known from more-or-less complete skull roofs. P. rectangulus is represented by skull elements from the Norian of Germany, whose bone pattern was reconstructed by Schultze (Citation1981), and from the Norian of Greenland, whose bone pattern was reconstructed by Pawlak et al. (Citation2020). Unfortunately, the arrangements of the central and posterior bones of the medial series are unknown in both cases. In Schultze (Citation1981: fig. 15), this part of the calvaria is reconstructed by comparison with P. serratus, resulting in a central C bone, a posterior bone AB, and a mediolateral bone J, all of which are hypothetical and separated by dotted lines on the reconstruction. In the specimen from Greenland, the presence of a J bone is considered uncertain, and more likely fused with the I bone (Pawlak et al., Citation2020). Pawlak et al. (Citation2020) also suggested that only two bones were present in the medial series (the E bone and the ABC bone) a hypothesis we support based on the margin of the void zone of the calvarium of this specimen (Pawlak et al., Citation2020:fig. 8). The pterygoid and prearticular tooth plates of P. rectangulus bear five and four ridges, respectively. Cranial and dental features of this species allow us to refer it to Ferganoceratodus, as F. rectangulus comb. nov.
Based on the diagnostic characters proposed above for Ptychoceratodus and Ferganoceratodus, the generic attributions of several Mesozoic species should be reinvestigated. Indeed, consideration should be given to referring most of the Triassic lungfishes known only from tooth plates to Ferganoceratodus based on the number of ridges present, i.e., pterygoid and prearticular tooth plates with five and four ridges, respectively. Other tooth plate characters used by Skrzycki (Citation2015) to diagnose species are also present in Ferganoceratodus. For example, P. concinnus and P. roemeri have prearticular tooth plates that are close to each other, as in Ferganoceratodus (contact lacking in P. serratus); and in these species the first ridge is curved more-or-less posteriorly, as in Ferganoceratodus (whereas it is curved sinusoidally in P. serratus). The other characters used by Skrzycki (Citation2015) are poorly defined and are not assessed here. Consequently, we suggest referring these two species to Ferganoceratodus (as F. concinnus and F. roemeri), and keep the question open for other species previously referred to Ptychoceratodus (P. silesiacus, P. donensis, P. madelungi).
Asian ‘ptychoceratodonts’
In addition to the first discovered occurrence of Ferganoceratodus from the Middle Jurassic of Kyrgyzstan (Nessov & Kaznyshkin, Citation1985), Asian countries have yielded a rich fossil record of this genus. ‘Ceratodus’ szechuanensis and ‘Ceratodus’ youngi were described from the Lower Jurassic Kyangyan site near Weiyuan, Sichuan (Liu & Yeh, Citation1957; Young, Citation1942). Martin et al. (Citation1997) considered ‘C’. youngi to be a junior synonym of ‘C.’ szechuanensis and included this species in the genus Ferganoceratodus. In Thailand, Martin and Ingavat (Citation1982) and Martin et al. (Citation1999) referred isolated dental plates from the Upper Triassic Hai Hin Lat Formation, the Middle Jurassic Khlong Min Formation, and the lower Upper Jurassic Phu Kradung Formation to F. szechuanensis. In the upper part of the Phu Kradung Formation, considered to be basal Cretaceous in age, the Phu Nam Jun site has yielded a sub-complete lungfish skull with its tooth plates still attached, which Cavin et al. (Citation2007) referred to the new species F. martini. They suggested that the isolated Late Triassic and Jurassic tooth plates from Thailand could all be referred to Ferganoceratodus. Recently, a separate species, F. annekempae, was recognized from the Phu Noi site based on a partially articulated skull with associated tooth plates. This locality is situated in the Phu Kradung Formation but is stratigraphically lower than the Phu Nam Jun site and is considered to be Upper Jurassic in age (Cavin et al., Citation2020). Cranial material from F. martini and F. annekempae show that the median series is composed of two ossifications only, as in the type species F. jurassicus.
North American ‘ptychoceratodonts’
Ferganoceratodus was present on almost all landmasses during the first part of the Mesozoic, with the conspicuous exception of North America. This absence might have been caused by a bias linked to the “traditional practice of assigning isolated lungfish tooth plates from the Cretaceous of North America to Ceratodus” (Parris et al., Citation2014:278). Indeed, several species of ‘Ceratodus’ from the Late Jurassic and Early Cretaceous were included by Parris et al. (2014) and Frederickson and Cifelli (Citation2017) in the ‘ptychoceratodont morphotype,’ due to the presence of well-developed ridges. Additionally, these species have plates with ridge numbers that correspond to the genus Ferganoceratodus, with five and four ridges on the pterygoid and prearticular dental plates, respectively. These are Potamoceratodus guentheri, Ceratodus fossanovum, C. texanus, C. kirklandi, and C. carteri (Frederickson & Cifelli, Citation2017; Main et al., Citation2014; Pardo et al., Citation2010; Parris et al., 2014). Potamoceratodus guentheri, from the Upper Jurassic Morrison Formation, is one of the few North American Mesozoic lungfish associated with a fairly well-preserved skull roof. The calvarium is characterized by an anterior rostral extension formed by the anterior median bone, and by a posterolateral extension of the posterior bone of the medial series (Pardo et al., Citation2010). The last characteristic recalls the pattern present in Tellerodus sturii from the Upper Triassic of Lunz, Austria (Martin, Citation1982b). Schultze (Citation1981) considered this bone to be the fusion of the ABIJ bones in Tellerodus, and Pardo et al. (Citation2010) as the fusion of the ABCI bones in Potamoceratodus. However, the homologies between these bones have been questioned (e.g., Cavin et al., Citation2007), and we consider here that the bone patterns of the two taxa are very similar to each other, and distinct from Ferganoceratodus. This distinction is reinforced by the Tellerodus ceratodontid-like tooth plate possessed by Potomoceratodus that is very different from the Ferganoceratodus–Ptychoceratodus pattern, indicating that the North American ‘ptychoceratodont morphotype’ tooth plates might represent a separate lineage of North American lungfish, as already suggested by Pardo et al. (Citation2010) and Frederickson and Cifelli (Citation2017), a lineage that shows some convergence in the morphology of their dental plates with Ferganoceratodus.
In the Late Triassic and Early Jurassic of North America, other non-ceratodontid lungfish have been recorded. The Chinle Formation has yielded remains referred to Arganodus dorotheae (Martin, Citation1982a), a tooth plate with six sharp ridges that differs from Ferganoceratodus. In the Lower Jurassic Moenave Formation, Ceratodus stewarti is based on a single prearticular dental plate with four fairly sharp ridges that is somewhat similar to Ferganoceratodus (Milner & Kirkland, Citation2006), but which might suggest it belongs to the Potomoceratodus lineage mentioned above.
Paleobiogeography and Paleoecology
Based on this short review, Ferganoceratodus first appeared during the Early Triassic in southern Gondwana, while in Europe another genus is present, Ptychoceratodus (). During the Middle and Late Triassic, the geographic distribution of Ferganoceratodus appears to expand, with occurrences in western Gondwana (Africa and South America), including the Zimbabwean occurrence of F. edwardsi described here, and occurrences in Laurasia, i.e., Europe, Greenland, and central and eastern Asia. In the Late Jurassic, the Laurasian distribution persists, but there are no further occurrences in Gondwana. During the Early Cretaceous, only one occurrence is noted, in Thailand, and in the Late Cretaceous, only Madagascar and Argentina have yielded remains referable to this genus.
FIGURE 6. Geographic and stratigraphic distributions of Ferganoceratodus spp. and Ptychoceratodus serratus. See text for details.
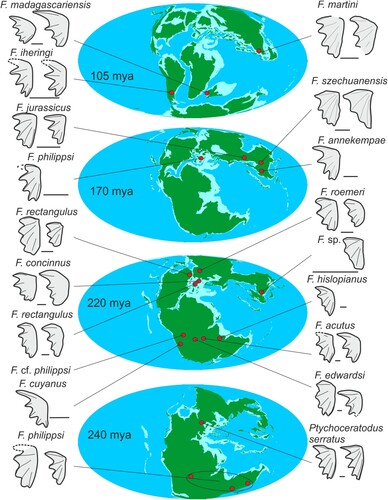
As noted elsewhere, the Pebbly Arkose Formation of the Kariba region is unusual in comparison with other Karoo-aged basins in southern and eastern Africa in preserving abundant remains of freshwater taxa, including lungfish, temnospondyl amphibians, and phytosaurs (Barrett et al., Citation2020). Lungfish tooth plates are very abundant at several sites (Sciscio et al., Citation2021b), especially at ‘The Dentists’ and other nearby localities, but they are usually incomplete, preventing their definitive referral to Ferganoceratodus edwardsi, although their preserved portions are morphologically similar (see Supplementary Information; Raath et al., Citation1992). This indicates that they were probably more important faunal components of Karoo-aged ecosystems than their poor record in other similarly aged basins, such as the main Karoo Basin (e.g., Kitching & Raath, Citation1984), would suggest.
CONCLUSIONS
The discovery of numerous lungfish tooth plates from the Upper Triassic Pebbly Arkose Formation of northern Zimbabwe indicates the presence of a new species of ‘ptychoceratodontid’ in Gondwana, Ferganoceratodus edwardsi. This new taxon resembles another species from approximately contemporary sites in Madagascar, F. acutus. A brief review of the global record of ‘ptychoceratodontids’ and comparisons of these taxa with the holotype specimens of Ptychoceratodus and Ferganoceratodus suggest that most of the species previously referred to Ptychoceratodus should be referred to Ferganoceratodus. This would indicate the presence of a group of lungfish species with few cutting ridges, distinct from the ceratodontids with crushing-tooth plates and also from neoceratodontids, which have tooth plates with numerous parallel ridges. The interrelationships within this species-group are unclear due to difficulties in defining characters based solely on tooth plates for phylogenetic analysis. Histological data from a prearticular tooth plate of Ferganoceratodus edwardsi exhibits similarities with the Mesozoic taxa Ceratodus latissimus and Metaceratodus baibianorum. It is not clear if ‘ptychoceratodontid’ lungfishes form a clade (Agnolin, Citation2010), if they correspond to stem members of Lepidosirenidae (Cavin et al., Citation2007), are stem neoceratodontids (Apesteguía et al., 2007), or stem Ceratodontoidei (Kemp et al., Citation2017). This uncertainty, and the lack of published histological data from most Mesozoic lungfish tooth plates, underscores the difficulty of constructing a useful phylogenetic framework from fossil material with broad spatial and temporal distributions but few characters to work with.
AUTHOR CONTRIBUTIONS
JNC, TJC, LC and PMB designed the project; DM, MZ, JNC and PMB conducted fieldwork and collected specimens; TJC, LC, and PMB analysed the specimens and collected data; TJC, LC, and PMB drafted the manuscript; all authors edited the manuscript.
SUPPLEMENTARY FILES
Supplementary File 1.docx: Other lungfish material.
Supplemental Material
Download MS Word (7.2 MB)ACKNOWLEDGMENTS
We thank S. Edwards for his tireless work in bringing the paleontology of the Kariba region to light and for his help, knowledge, and hospitality during our fieldwork in the region. We also thank other members of our extended Kariba fieldwork family for discussion and support: T., P., and L. Broderick, K. Chapelle, K. Dollman, W. Edwards, D. and J. Glynn, E. Mbambo, R. Niven, L. Sciscio, S. Tolan, and P. Viglietti. The crew of the ‘Musankwa,’ our home during our time on Kariba—C. Katupu, G. Swalika, S. Mangoroma, and N. Mapira—kept us all safe and well looked-after. V. Fernandez (formerly NHM, London) is thanked for scanning the tooth plate material. I. Febbrari (University of Edinburgh) is thanked for preparing thin-sections. The National Museums and Monuments of Zimbabwe enabled the loan of this material to PMB. This research was made possible by the continued support of Earth Sciences Departmental Investment Fund (NHM, London: to PMB), the Palaeontological Scientific Trust (to JNC), the National Research Foundation of South Africa (NRF) African Origins Platform (grants 98800 and 118794 to JNC), and GENUS: the DSI-NRF Centre of Excellence in Palaeosciences (to JNC). We thank Matsuadona National Park for permission to work in the area, under a permit obtained from the Research Council of Zimbabwe (RCZ 02849). Two anonymous reviewers made valuable suggestions that greatly contributed to the quality of the manuscript.
DISCLOSURE STATEMENT
No potential conflict of interest was reported by the author(s).
DATA AVAILABILITY STATEMENT
The data that support the findings of this study are openly available in Morphosource at https://www.morphosource.org, project ID: 000547763 and https://doi.org/10.17602/M2/M547805
.LITERATURE CITED
- Agassiz, L. (1833–1843). Recherches sur les Poissons Fossiles. Volumes 1– 5. Petitpierre.
- Agnolin, F. (2010). A new species of the genus Atlantoceratodus (Dipnoiformes: Ceratodontoidei) from the uppermost Cretaceous of Patagonia and a brief overview of fossil dipnoans from the Cretaceous and Paleogene of South America. Brazilian Geographical Journal: Geosciences and Humanities Research Medium, 1(2), 162–210.
- Agnolín, F. L., Bogan, S., Egli, F. B., Novas, F. E., Isasi, M. P., Marsicano, C., & Mancuso, A. (2017). A new lungfish (Dipnoi) from the Late Triassic of South America. Journal of Vertebrate Paleontology, 37(1), e1245665. https://doi.org/10.1080/02724634.2017.1245665
- Ahlberg, P. E., Smith, M. M., & Johanson, Z. (2007). Developmental plasticity and disparity in early dipnoan (lungfish) dentitions (vol 8, pg 331, 2006). Evolution & Development, 9(3), 306–306. doi:10.1111/j.1525-142X.2007.00163_1.x
- Ait-Kaci Ahmed, A. (2018). The geology of the country between Chireya and Goredema, Gokwe North and Gokwe South Districts. Zimbabwe Geological Survey Bulletin, 105, 1–86.
- Antunes, M. T., Maisey, J. G., Marques, M. M., Schaeffer, B., & Thomson, K. S. (1990). Triassic fishes from the Cassange Depression (R. P. de Angola). Ciencias da Terra (Número especial), 1–64.
- Apesteguía, S., Agnolin, F., & Claeson, K. (2007). Review of Cretaceous dipnoans from Argentina (Sarcopterygii: Dipnoi) with descriptions of new species. Revista del Museo Argentino de CIencias Naturales, 9 (1), 27–40. doi:10.22179/REVMACN.9.362
- Barrett, P. M., Sciscio, L., Viglietti, P. A., Broderick, T. J., Suarez, C. A., Sharman, G. R., Jones, A. S., Munyikwa, D., Edwards, S. F., Chapelle, K. E. J., Dollman, K. N., Zondo, M., & Choiniere, J. N. (2020). The age of the Tashinga Formation (Karoo Supergroup) in the Mid-Zambezi Basin, Zimbabwe and the first phytosaur from mainland sub-Saharan Africa. Gondwana Research, 81, 445–460. https://doi.org/10.1016/j.gr.2019.12.008
- Bender, P. A., & Hancox, P. J. (2004). Newly discovered fish faunas from the Early Triassic, Karoo Basin, South Africa, and their implications. Gondwana Research, 7(1), 185–192. https://doi.org/10.1016/S1342-937X(05)70317-8
- Bond, G. (1972). Milestones in Rhodesian palaeontology (1901–1971). South African Journal of Geology, 75, 149–58. https://hdl.handle.net/10520/AJA10120750_2736
- Bond, G., & Falcon, R. (1973). The palaeontology of Rhodesia, with a section on the palynology of the Middle Zambezi Valley. Rhodesia Geological Survey Bulletin, 70, 1–121.
- Broom, R. (1909). The fossil fishes of the Upper Karoo beds of South Africa. Annals of the South African Museum, 7, 251-269.
- Broom, R. (1913). The fossil fishes of the upper Karoo Beds of South Africa. Annals of the South African Museum, 7, 251–268. + pls. XII–XIII.
- Brownstein, C. D., Harrington, R. C., & Near, T. J. (2023). The biogeography of extant lungfishes traces the breakup of Gondwana. Journal of Biogeography, 50(7), 1191–1198. https://doi.org/10.1111/jbi.14609
- Cavin, L., Suteethorn, V., Buffetaut, E., & Tong, H. (2007). A new Thai Mesozoic lungfish (Sarcopterygii, Dipnoi) with an insight into post-Palaeozoic dipnoan evolution. Zoological Journal of the Linnean Society, 149(2), 141–177. https://doi.org/10.1111/j.1096-3642.2007.00238.x
- Cavin, L., Deesri, U., & Chanthasit, P. (2020). A new lungfish from the Jurassic of Thailand. Journal of Vertebrate Paleontology, 40(4), e1791895. https://doi.org/10.1080/02724634.2020.1791895
- Cione, A. L., & Gouiric-Cavalli, S. (2012). Metaceratodus kaopen comb. nov. and M. wichmanni comb. nov., two Late Cretaceous South American species of an austral lungfish genus (Dipnoi). Alcheringa: An Australasian Journal of Palaeontology, 36(2), 203–216. doi:10.1080/03115518.2012.621804
- Cione, A. L., Gouiric Cavalli, S., Goin, F. J., & Poiré, D. G. (2007). Atlantoceratodus a new genus of lungfish from the upper Cretaceous of South America and Africa. Revista del Museo de La Plata, 10(62), 1–12.
- Crompton, A. W. (1955). On some Triassic cynodonts from Tanganyika. Proceedings of the Zoological Society of London, 125(3-4), 617–669. doi:10.1111/j.1096-3642.1955.tb00620.x
- Dubiel, R.F., Blodgett, R.H. and Bown, T.M. (1987) Lungfish burrows in the Upper Triassic Chinle and Dolores Formations, Colorado Plateau. Journal of Sedimentary Research, 57(3), 512-521.
- Frederickson, J. A., & Cifelli, R. L. (2017). New Cretaceous lungfishes (Dipnoi, Ceratodontidae) from western North America. Journal of Paleontology, 91(1), 146–161. https://doi.org/10.1017/jpa.2016.131
- Gobetz, K., & Andallanjlerner, S. L. (2006). Lungfish burrows of varying morphology from the Upper Triassic. New Mexico Museum of Natural History and Science, Bulletin, 37, 140–146.
- Griffin, C. T., Wynd, B. M., Munyikwa, D., Broderick, T. J., Zondo, M., Tolan, S., Langer, M. C., Nesbitt, S. J., & Taruvinga, H. R. (2022). Africa’s oldest dinosaurs reveal early suppression of dinosaur distribution. Nature, 609(7926), 313–319. doi:10.1038/s41586-022-05133-x
- Jaekel, O. (1926). Zur Morphogenie der Gebisse und Zähne. Vierteljahrsschrift Zahnheilkunde, 42, 217–242.
- Jubb, R. A., & Gardiner, B. G. (1975). A preliminary catalogue of identifiable fossil fish material from Southern Africa. Annals of the South African Museum, 67(11), 381–440.
- Kemp, A. (1993). Ceratodus diutinus, a new ceratodont from Cretaceous and Late Oligocene–Medial Miocene deposits in Australia. Journal of Paleontology, 67(5), 883–888. doi:10.1017/S0022336000037148
- Kemp, A. (1996). Triassic lungfish from Gondwana. In G. Arratia, & G. Viohl (Eds.), Mesozoic fishes: Systematics and paleoecology (pp. 409–416). Verlag Dr Friedrich Pfeil.
- Kemp, A. (1997). Four species of Metaceratodus (Osteichthyes: Dipnoi, Family Ceratodontidae) from Australian Mesozoic and Cenozoic deposits. Journal of Vertebrate Paleontology, 17(1), 26–33. https://doi.org/10.1080/02724634.1997.10010949
- Kemp, A. (1998). Skull structure in post-Paleozoic lungfish. Journal of Vertebrate Paleontology, 18(1), 43–63. https://doi.org/10.1080/02724634.1998.10011033
- Kemp, A. (2001). Petrodentine in derived dipnoan tooth plates. Journal of Vertebrate Paleontology, 21(3), 422–437. https://doi.org/10.1671/0272-4634(2001)021[0422:PIDDTP]2.0.CO;2
- Kemp, A. (2002). Growth and hard tissue remodelling in the dentition of the Australian lungfish, Neoceratodus forsteri (Osteichthyes: Dipnoi). Journal of Zoology, 257(2), 219–235. doi:10.1017/S0952836902000821
- Kemp, A., & Barry, J. C. (2006). Prismatic dentine in the Australian lungfish, Neoceratodus forsteri (Osteichthyes: Dipnoi). Tissue and Cell, 38(2), 127–140. https://doi.org/10.1016/j.tice.2006.01.001
- Kemp, A., Cavin, L., & Guinot, G. (2017). Evolutionary history of lungfishes with a new phylogeny of post-Devonian genera. Palaeogeography, Palaeoclimatology, Palaeoecology, 471, 209–219. https://doi.org/10.1016/j.palaeo.2016.12.051
- Kitching, J. W., & Raath, M. A. (1984). Fossils from the Elliot and Clarens formations (Karoo sequence) of the northeastern Cape, Orange Free State, and Lesotho, and a suggested biozonation based on tetrapods. Palaeontologia Africana, 25, 111–25. http://hdl.handle.net/10539/16130
- Lison, L. (1941). The structure of teeth of lungfish. Petrodentine. Comptes Rendus des Séances de la Société de Biologie et de ses Filiales, 135, 431.
- Liu, H.-T., & Yeh, H.-K. (1957). Two new species of Ceratodus from Szechuan, China. Vertebrata PalAsiatica, 4(1), 310–311.
- Main, D. J., D. C. Parris, B. S. Grandstaff, & B. Carter. (2014). A new lungfish (Dipnoi: Ceratodontidae) from the Cretaceous Woodbine Formation, Arlington Archosaur Site, north Texas. Texas Journal of Science 63, 283–298.
- Martin, M. (1981). Les dipneustes méozoïques malgaches, leurs affinités et leur intérêt paléogéographique. Bulletin de la Société Géologique de France, S7-XXIII(6), 579–585. doi:10.2113/gssgfbull.S7-XXIII.6.579
- Martin, M., Sigogneau-Russel, D., Coupatez, P., & Wouters, G. (1981). Les cératodontidés (Dipnoi) du Rhétien de Saint-Nicolas-de-Port (Meurthe-et-Moselle). Geobios, 14(6), 773–791. doi:10.1016/S0016-6995(81)80151-9
- Martin, M. (1982a). Nouvelles données sur la phylogénie et la systématique des Dipnoi postpaléozoïques. Comptes Rendus de l’Académie des Sciences, Série II, 294, 611–614.
- Martin, M. (1982b). Revision von Tellerodus sturii (Teller) 1891. Verhandlen Geologi, 1982, 21–31.
- Martin, M., & Ingavat, R. (1982). First record of an Upper Triassic ceratodontid (Dipnoi, Ceratodontiformes) in Thailand and its paleogeographical significance. Geobios, 15(1), 111–114. doi:10.1016/S0016-6995(82)80063-6
- Martin, M. (1982c). Nouvelles données sur la phylogénie et la systématique des Dipneustes postpaléozoïques, conséquences stratigraphiques et paléogéographiques. Geobios, 15, 53-64.
- Martin, M., Barbieri, L., & Cuny, G. (1999). The Madagascan Mesozoic ptychoceratodontids (Dipnoi) systematic relationships and paleobiogeographical significance. Oryctos, 2, 3–16.
- Martin, M., Buffetaut, E. Tong, H., & Suteethorn, V. (1997). New Jurassic dipnoans from Thailand. Geological Society of Denmark, Online Series, 1. https://2dgf.dk/dgf-online-series/new-jurassic-dipnoans-from-thailand/
- Milner, A. R., & Kirkland, J. I. (2006). Preliminary review of the Early Jurassic (Hettangian) freshwater Lake Dixie fish fauna in the Whitmore Point Member, Moenave Formation in southwest Utah. New Mexico Museum of Natural History and Science, Bulletin, 37, 510–521.
- Müller, J. (1845). Über den Bau und die Granzen der Ganoiden, und über das natürliche System der Fische. Archiv für Naturgeschichte, 1845, 91–142.
- Nessov, L. A., & Kaznyshkin, M. N. (1985). A lungfish and turtles from Upper Jurassic of northern Fergana, Kirghiz SSR. Vestnik Zoologii, 1, 33–39.
- Nikolski, G. W. (1954). Castnaja ikhtiologija. Sovetskaja Nauka. [In Russian].
- Oldham, T. (1859). On some fossil teeth of the genus Ceratodus from Maleri, south of Nagpur. Memoirs of the Geological Survey of India, 1, 300–309.
- Panzeri, K. M., Pereyra, M. E., & Cione, A. L. (2022). The South American dipnoan Metaceratodus baibianorum (Dipnoi, Ceratodontidae) from the Upper Cretaceous La Colonia Formation, Patagonia, Argentina: an approach from the histology of the tooth plates. Cretaceous Research, 133, 105144. https://doi.org/10.1016/j.cretres.2022.105144
- Panzeri, K. M., Cavalli, S. G., Muñoz, N. A., & Cione, A. L. (2020). Metaceratodus baibianorum, a new dipnoan species from the Upper Cretaceous of southern South America supported by traditional and geometric morphometric analyses. Journal of Vertebrate Paleontology, 40(2), e1769640. https://doi.org/10.1080/02724634.2020.1769640
- Pardo, J. D., Huttenlocker, A. K., Small, B. J., & Gorman, M. A. (2010). The cranial morphology of a new genus of lungfish (Osteichthyes: Dipnoi) from the Upper Jurassic Morrison Formation of North America. Journal of Vertebrate Paleontology, 30(5), 1352–1359. https://doi.org/10.1080/02724634.2010.501430
- Parris, D. C., Grandstaff, B. S., and Banks, N. T. (2014). Lungfishes from the Trinity Group (Cretaceous) of North Texas. Texas Journal of Science, 63, 267–282.
- Pawlak, W., Talanda, M., Sulej, T., & Niedzwiedzki, G. (2020). Dipnoan from the Upper Triassic of East Greenland and remarks about palaeobiogeography of Ptychoceratodus. Acta Palaeontologica Polonica, 65(3), 561–574. https://doi.org/10.4202/app.00679.2019
- Peecook, B. R., Steyer, J. S., Tabor, N. J., & Smith, R. M. (2017). Updated geology and vertebrate paleontology of the Triassic Ntawere Formation of northeastern Zambia, with special emphasis on the archosauromorphs. Journal of Vertebrate Paleontology, 37(sup1), 8–38. doi:10.1080/02724634.2017.1410484
- Priem, F. (1924). Paléontologie de Madagascar XII: les poissons fossiles. Annales de Paléontologie, 23, 83–178.
- Raath, M. A. (1996). The earliest evidence of dinosaurs from central Gondwana. Memoirs of the Queensland Museum, 39, 703–709.
- Raath, M. A., Oesterlen, P. M., and Kitching, J. W. (1992). First record of Triassic Rhynchosauria (Reptilia: Diapsida) from the Lower Zambezi Valley, Zimbabwe. Palaeontologica Africana, 29, 1–10. http://hdl.handle.net/10539/16176
- Reisz, R. R., Krupina, N. I., & Smith, M. M. (2004). Dental histology in Ichnomylax karatajae sp. nov., an early Devonian dipnoan from the Taymir Peninsula, Siberia, with a discussion on petrodentine. Journal of Vertebrate Paleontology, 24(1), 18–25. doi:10.1671/1824-2
- Richter, M., & Toledo, C. E. V. (2008). The first Triassic lungfish from South America (Santa Maria Formation, Paraná Basin) and its bearing on geological correlations within Pangaea. Geological Society, London, Special Publications, 295(1), 43–54. https://doi.org/10.1144/SP295.4
- Romer, A. S. (1955). Herpetichthyes, Amphibioidei, Choanichthyes, or Sarcopterygii? Nature, 176(4472), 126. https://doi.org/10.1038/176126a0
- Romer, A. S. & Smith, H. J. (1934). American Carboniferous dipnoans. The Journal of Geology, 42(7), 700-719.
- Schindelin, J., Arganda-Carreras, I., Frise, E., Kaynig, V., Longair, M., Pietzsch, T., Preibisch, S., Rueden, C., Saalfeld, S., Schmid, B., & Tinevez, J. Y. (2012). Fiji: an open-source platform for biological-image analysis. Nature methods, 9(7), 676–682. doi:10.1038/nmeth.2019
- Schultze, H. P. (1981). Das Schädeldach eines ceratodontiden Lungenfisches aus der Trias Süddeutschlands (Dipnoi, Pisces). Stuttgarter Beiträge für Naturkunde, Serie B, 70, 1–31.
- Schultze, H. P. (2004). Mesozoic sarcopterygians. In G. Arratia, & A. Tintori (Eds.), Mesozoic fishes 3 – systematics, paleoenvironments and biodiversity (pp. 463–492). Verlag Dr Friedrich Pfeil.
- Sciscio, L., Broderick, T. J., Barrett, P. M., Munyikwa, D., Zondo, M., & Choiniere, J. N. (2021a). Invertebrate and plant trace fossils from the terrestrial Late Triassic of Zimbabwe. Palaios, 36(4), 129–140. https://doi.org/10.2110/palo.2020.071
- Sciscio, L., Viglietti, P. A., Barrett, P. M., Broderick, T. J., Munyikwa, D., Chapelle, K. E. J., Dollman, K. N., Edwards, S. F., Zondo, M., & Choiniere, J. N. (2021b). Sedimentology and palaeontology of the Upper Karoo Group in the Mid-Zambezi Basin, Zimbabwe: new localities and their implications for interbasinal correlation. Geological Magazine, 158(6), 1035–1058. https://doi.org/10.1017/S0016756820001089
- Seegis, D. (1997). Die Lehrbergschichten im Mittleren Keuper von Süddeutschland – Stratigraphie, Petrographie, Paläontologie, Genese. Verlag Manfred Hennecke.
- Skrzycki, P. (2015). New species of lungfish (Sarcopterygii, Dipnoi) from the Late Triassic Krasiejów site in Poland, with remarks on the ontogeny of Triassic dipnoan tooth plates. Journal of Vertebrate Paleontology, 35(5), e964357. https://doi.org/10.1080/02724634.2015.964357
- Smith, M. M. (1984). Petrodentine in extant and fossil dipnoan dentitions: microstructure, histogenesis and growth. In Proceedings of the Linnean Society of New South Wales, 107, 367-407.
- Smith, M. & Campbell, K. S. W. (1987). Comparative morphology, histology and growth of the dental plates of the Devonian dipnoan Chirodipterus. Philosophical Transactions of the Royal Society of London. B, Biological Sciences, 317(1185), 329-363.
- Young, C.-C. (1942). Fossil vertebrates from Kuangyuan, N. Szechuan, China. Bulletin of the Geological Society of China, 22(3-4), 293–309. doi:10.1111/j.1755-6724.1942.mp223-4016.x
- Woodward, A. S. (1890). A synopsis of the fossil fishes of the English Lower Oolites. Proceedings of the Geologists' Association, 11(6), 285–306. doi:10.1016/S0016-7878(90)80030-4