
ABSTRACT
We report stem group Galliformes from the lower Eocene (Ypresian) London Clay of Walton-on-the-Naze (Essex, U.K.), which are among the oldest known fossils of galliform birds. The specimens are assigned to two new species, ?Paraortygoides argillae, sp. nov. and Waltonortyx bumbanipodiides, gen. et sp. nov. The latter is classified in a new family-level taxon, Waltonortygidae, fam. nov., which differs from other higher-level clades of stem group Galliformes in a proximo-distally narrow caput humeri and a short tuberculum dorsale of the humerus. With regard to these plesiomorphic features and a characteristic morphology of the coracoid, Waltonortyx is similar to early Eocene stem group Galliformes from Mongolia. Wing and pectoral girdle bones of a third, undetermined galliform from Walton-on-the-Naze resemble the Quercymegapodiidae; this fossil also shows a similarity to early Eocene galliforms from Mongolia. The affinities of “Paraortygoides” radagasti Dyke and Gulas, 2002, the only previously described putative galliform from Walton-on-the-Naze, are uncertain and neither its classification in the Galliformes nor its assignment to the taxon Paraortygoides are unequivocally established. For the first time we subjected a larger taxonomic sample of early Paleogene Galliformes to a formal analysis, and even though the resulting phylogenies were poorly resolved, they supported paraphyly of the Quercymegapodiidae, with the Early Miocene taxon Ameripodius being more closely related to the crown group than the late Eocene species of Quercymegapodius. Waltonortyx bumbanipodiides was recovered as the sister taxon of the Mongolian Bumbanipodius transitoria, which indicates early Paleogene dispersal events across the Turgai Strait.
INTRODUCTION
Until quite recently, galliform birds had a very limited fossil record from the Eocene epoch (Mayr, Citation2009, Citation2022). Gallinuloides wyomingensis from the lower Eocene North American Green River Formation, of which two articulated skeletons on slabs were described, was for a long time the only substantial galliform from the early Eocene (Eastman, Citation1900; Lucas, Citation1900; Mayr & Weidig, Citation2004; Shufeldt, Citation1915). The species belongs to the taxon Gallinuloididae, which also includes the latest early or earliest middle Eocene Paraortygoides messelensis from Messel in Germany (Mayr, Citation2000, Citation2006).
Other stem group representatives of the Galliformes from the Eocene of Europe belong to the Quercymegapodiidae and Paraortygidae, which are particularly well represented in the Quercy fissure fillings in France (Mourer-Chauviré, Citation1992). The quercymegapodiid taxon Quercymegapodius occurs in middle and late Eocene Quercy localities (Mourer-Chauviré, Citation1992). Other presumed quercymegapodiids were reported from the Oligocene of Brazil (Alvarenga, Citation1995) and the Early Miocene of France (Mourer-Chauviré, Citation2000). The Paraortygidae are known from the late Eocene and Oligocene of the Quercy fissure fillings and other localities in Belgium and Germany, and their European representatives are classified in the taxa Paraortyx and Pirortyx (Fischer, Citation1990; Mayr, Citation2009, Citation2022; Mayr & Smith, Citation2019; Mourer-Chauviré, Citation1988, Citation1992, Citation2006). A fragmentary coracoid from the middle Eocene of Utah (U.S.A.) was assigned to the Paraortygidae by Stidham et al. (Citation2020).
Even though some of the above taxa have a comprehensive fossil record, their interrelationships are not well understood. Mourer-Chauviré (Citation1992) hypothesized that the Paraortygidae is the sister taxon of a clade including the Quercymegapodiidae and crown group Galliformes, with a transverse ridge in the ventral portion of the incisura capitis of the humerus being a derived feature shared by quercymegapodiids and crown group galliforms (the clade including the Megapodiidae, Cracidae, and Phasianidae). Mayr (Citation2000) and Mayr and Weidig (Citation2004) placed the Gallinuloididae and Paraortygidae in a polytomy at the base of a clade including the Quercymegapodiidae and crown group Galliformes.
In addition to uncertainties surrounding the composition and affinities of the higher-level clades, the study of early Paleogene Galliformes is complicated by the fact that various species were based on very fragmentary material, which is particularly true for fossils from the lower Eocene British London Clay. Three species of putative galliforms from the Isle of Sheppey (Essex) were described as Argillipes aurorum, A. paralectoris, and Percolinus venablesi by Harrison and Walker (Citation1977). All of these were based on fragmentary tarsometatarsi (of A. aurorum a distal humerus was also identified). The hypotarsi of Argillipes aurorum and Percolinus venablesi exhibit a similar, galliform-like morphology, with a large canal for the tendon of musculus flexor digitorum longus (see Mayr, Citation2016). By contrast, the hypotarsus of “Argillipes” paralectoris lacks this canal and the species is unlikely to belong to the genus it was originally assigned to and probably does not even belong to the Galliformes. Based on the distal end of a tarsometatarsus, another species from the London Clay of Burnham-on-Crouch (Essex) was named Coturnipes cooperi by Harrison and Walker (Citation1977); this species was likened to the gruiform Messelornithidae by Mayr et al. (Citation2022), but its affinities are elusive owing to the fragmentary condition of the holotype.
Dyke and Gulas (Citation2002) described a fragmentary partial skeleton of a putative galliform from the London Clay of Walton-on-the-Naze (Essex) as Paraortygoides radagasti. Most recently, Houde et al. (Citation2023) assigned partial skeletons of a putative anseriform bird from Walton-on-the-Naze to the new species Danielsavis nazensis, which was classified in the monotypic taxon Danielsavidae by Mayr et al. (Citation2023), who hypothesized that D. nazensis may actually represent an archaic, anseriform-like stem group representative of the Galliformes.
Mayr and Smith (Citation2019) reported isolated bones of Argillipes-like stem group galliforms from the lower Eocene of Egem in Belgium. The identification of other specimens and species of putative Galliformes from the Eocene of Europe needs to be corroborated with more fossil material. This is true for an isolated foot of an unnamed species from the lower Eocene Fur Formation in Denmark (Lindow & Dyke, Citation2007) as well as two species from the middle Eocene of Hampshire (U.K.), which were described by Harrison and Walker (Citation1979) as Percolinus proudlocki (the holotype and only known specimen is the proximal end of a tarsometatarsus) and Litoripes medius (the holotype is a tarsometatarsus). Tegulavis corbalani is a putative galliform species from the lower Eocene of France, which is only known from a coracoid (Mourer-Chauviré et al., Citation2024).
In the past years, various Eocene stem group Galliformes were identified in African fossil sites. These include Namaortyx sperrgebietensis and Scopelortyx klinghardtensis from the middle Eocene of Namibia (Mourer-Chauviré et al., Citation2011, Citation2015, Citation2017) and the late early or early middle Eocene Chambiortyx cristata from Tunisia (Mourer-Chauviré et al., Citation2013). Whereas N. sperrgebietensis and C. cristata are only known from partial tarsometatarsi (and a referred fragmentary carpometacarpus in the case of C. cristata), S. klinghardtensis is well represented by most postcranial elements, even though it is the tarsometatarsus that is unknown from this taxon. Scopelortyx was assigned to the Paraortygidae (Mourer-Chauviré et al., Citation2015), but the exact affinities of Namaortyx and Chambiortyx are elusive.
The fossil record of Eocene stem group Galliformes from Asia has also improved. Specimens from the lower Eocene Naranbulag Formation of Tsagaan Khushuu in Mongolia have been identified by Hwang et al. (Citation2010) and Hood et al. (Citation2019). Zelenkov (Citation2021) described two species from this site as Bumbanortyx transitoria and Bumbanipodius magnus; owing to the fragmentary nature of the material, the affinities of both species could not be unambiguously determined, but Zelenkov (Citation2021) noted similarities to the Gallinuloididae and Quercymegapodiidae. Xorazmortyx turkestanensis from the middle Eocene of Uzbekistan was assigned to the Paraortygidae by Zelenkov and Panteleyev (Citation2019).
Here we report specimens of galliforms from Walton-on-the-Naze, which stem from the collection of the late Michael Daniels. With an age of 54.6–55 million years (Ma), these fossils are among the earliest unequivocal galliforms known so far, being coeval with the specimens from the Mongolian Naranbulag Formation, which date to about 55 Ma (Hood et al., Citation2019; Hwang et al., Citation2010). Most previous fossils of early Eocene Galliformes were skeletons on slabs or fragmentary isolated bones, so the fossils from Walton-on-the-Naze are of significance, because they include associated and three-dimensionally preserved bones of single individuals.
MATERIAL AND METHODS
The phylogenetic analysis was run with the heuristic search modus of PAUP*4.0a169 (Swofford, Citation2002) with the “simple” stepwise addition option, random-addition sequences (10,000 replicates), tree-bisection-reconnection (TBR) with reconnection limit = 8 for the branch swapping algorithm, and collapsing branches if maximum branch length is 0. The matrix contained 17 ingroup taxa and 32 characters; outgroup comparisons were performed with Chauna torquata (Anseriformes, Anhimidae). Bootstrap support values were calculated with 100 replicates. Tree length (L), consistency index (CI), and retention index (RI) were calculated. Four characters (3, 19, 21, and 22) were coded as ordered. Character descriptions and the matrix are given in the Supplementary Files 1–3. The data matrix is made available as a NEXUS file in the Supplementary Material.
Institutional Abbreviations—IRSNB, Royal Belgian Institute of Natural Sciences, Brussels, Belgium; NHMUK, Natural History Museum, London, U.K.; NMS, National Museums Scotland, Edinburgh, U.K.; PIN, Borissiak Paleontological Institute, Russian Academy of Sciences, Moscow, Russia; SMF, Senckenberg Research Institute Frankfurt, Frankfurt, Germany; UM, Institut des Sciences de l’Evolution, Université de Montpellier, Montpellier, France; WDC, Wyoming Dinosaur Center, Thermopolis, Wyoming, U.S.A.
SYSTEMATIC PALEONTOLOGY
Remarks—Our assignment of the new species described below to the taxon Paraortygoides is based on a similar size and overall morphology of the tarsometatarsus and has to be regarded as tentative until more material is identified.
FIGURE 1. Fossils of newly identified galliform birds from Walton-on-the-Naze (Essex, U.K.) and the holotype of “Paraortygoides” radagasti. A, ?Paraortygoides argillae, sp. nov. (holotype, NMS.Z.2021.40.174). B, Waltonortyx bumbanipodiides, gen. et sp. nov. (holotype, NMS.Z.2021.40.175). C, undetermined galliform from Walton-on-the-Naze (NMS.Z.2021.40.176). D, “Paraortygoides” radagasti (holotype, NHMUK A 6217); not shown are pedal phalanges associated with the specimen (see Dyke & Gulas, Citation2002:fig. 3A). The scale bars equal 5 mm.

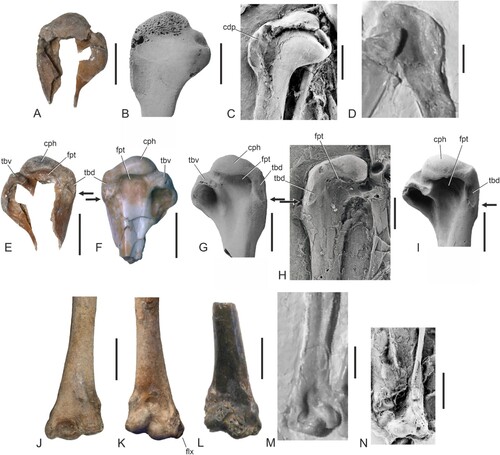
←FIGURE 2. Tarsometatarsi and feet of Eocene galliform and galliform-like birds. A–D, ?Paraortygoides argillae, sp. nov. from the London Clay of Walton-on-the-Naze (holotype, NMS.Z.2021.40.174), right (A, B) and left (C, D) tarsometatarsus in dorsal (A, C) and plantar (B, D) view. E, F, Argillipes aurorum, from the London Clay of the Isle of Sheppey (holotype, NHMUK A 3130), proximal portion of right tarsometatarsus in dorsal (E) and plantar (F) view. G, H, A. aurorum, proximal portion of a referred right tarsometatarsus from the Isle of Sheppey (NHMUK A 4283) in dorsal (G) and plantar (H) view. I, J, “A.” paralectoris from the London Clay of the Isle of Sheppey (holotype, NHMUK A 3604), proximal portion of left tarsometatarsus in dorsal (I) and plantar (J) view. K, L, Percolinus venablesi from the London Clay of the Isle of Sheppey (holotype, NHMUK A 3680), proximal portion of left tarsometatarsus in dorsal (K) and plantar (L) view. M, N, cf. A. aurorum from the lower Eocene of Egem in Belgium (IRSNB Av 166), proximal portion of right tarsometatarsus in dorsal (M) and plantar (N) view; the specimen was coated with ammonium chloride. O, P, cf. A. aurorum from the lower Eocene of Egem in Belgium (IRSNB Av 165), distal end of right tarsometatarsus in dorsal (O) and plantar (P) view; the specimen was coated with ammonium chloride. Q–T, ?P. argillae, sp. nov. (holotype, NMS.Z.2021.40.174), distal end of left tarsometatarsus in dorsal (Q), plantar (R), and distal (S) view; distal end of right tarsometatarsus in distal view (T). U–W, Coturnipes cooperi from the London Clay of Burnham-on-Crouch (holotype, NHMUK A 3706), distal end of left tarsometatarsus in dorsal (U), plantar (V), and distal (W) view. X–Z, cf. A. aurorum from Egem (IRSNB Av 165), distal end of right tarsometatarsus in dorsal (X), plantar (Y), and distal (Z) view; the specimen was coated with ammonium chloride. AA, ?P. argillae, sp. nov. (holotype, NMS.Z.2021.40.174), distal end of left tarsometatarsus (mirrored) in dorsal view. BB, Gallinuloides wyomingensis from the lower Eocene Green River Formation (WDC CGR−012), distal end of right tarsometatarsus in dorsal view. CC, ?P. argillae, sp. nov. (holotype, NMS.Z.2021.40.174), distal end of left tarsometatarsus (mirrored) in plantar view. DD, Paraortygoides messelensis from the latest early or earliest middle Eocene of Messel (holotype, SMF-ME 1303a), distal end of right tarsometatarsus in plantar view. EE, “Paraortygoides” radagasti (holotype, NHMUK A 6217), distal end of right tarsometatarsus in plantar view. FF–KK, proximal end of the tarsometatarsus (proximal view) of FF, ?P. argillae, sp. nov. (holotype, NMS.Z.2021.40.174, left side); GG, A. aurorum (holotype, NHMUK A 3130, right side); HH, A. aurorum (NHMUK A 4283, right side); II, cf. A. aurorum (IRSNB Av 166, right side); JJ, “A.” paralectoris (holotype, NHMUK A 3604, left side); KK, P. venablesi (holotype, NHMUK A 3680, left side). LL, ?P. argillae, sp. nov. (holotype, NMS.Z.2021.40.174), distal end of left tarsometatarsus with pedal phalanges (in different orientations); the asterisks denote tentatively identified phalanges. MM, G. wyomingensis (WDC CGR−012), right foot in dorsal view. NN, P. messelensis (holotype, SMF-ME 1303a), right foot in plantar view; the specimen was coated with ammonium chloride. In AA–EE the trochleae are numbered; the arrows in AA and BB denote the proximal terminus of the articular surface of the trochlea metatarsi III. In LL–NN the first phalanges of the second and third toes are labeled. Abbreviations: fdl, hypotarsal canal/sulcus for tendon of musculus flexor digitorum longus; fhl, hypotarsal sulcus for tendon of musculus flexor hallucis longus; fp2/fpp2, sulcus for the tendons of musculus flexor perforatus digiti 2 and m. perforans et perforatus digiti 2; fvd, foramen vasculare distale; ttc, tuberositas musculi tibialis cranialis. The scale bars equal 5 mm.

Holotype—NMS.Z.2021.40.174 (; proximal end of left femur, distal ends of both femora, distal portion of right tibiotarsus, proximal and distal portions of both tarsometatarsi, pedal phalanges); collected in 1982 by M. Daniels (original collector’s number WN 82383).
Differential Diagnosis—Smaller than Argillipes aurorum, “A.” paralectoris, Percolinus venablesi, and “Paraortygoides” radagasti. Distinguished from Paraortygoides messelensis Mayr, Citation2000 (Gallinuloididae) in that first phalanx of second toe proportionally shorter (slightly shorter than the first phalanx of third toe in P. messelensis, but somewhat longer in ?Paraortygoides argillae, sp. nov.; ) and in that ungual phalanges straighter and proportionally longer. Apart from a smaller size the new species differs from “Paraortygoides” radagasti Dyke and Gulas, Citation2002 in that trochlea metatarsi II proportionally shorter and hypotarsus with closed canal. Distinguished from Gallinuloides wyomingensis Eastman, Citation1900 (Gallinuloididae) in that tarsometatarsus with proportionally shorter dorsal portion of articular surface of trochlea metatarsi III (proximal terminus at level of distal end of trochlea metatarsi II, whereas it reaches to middle of trochlea metatarsi II in Gallinuloides; ), in that first phalanx of second toe proportionally shorter (slightly shorter than first phalanx of third toe in G. wyomingensis but slightly longer in ?P. argillae; ), and in that distal end of trochlea metatarsi IV more laterally slanted. Differs from the somewhat larger Argillipes aurorum Harrison and Walker, Citation1977 (fam. inc. sed.) in that hypotarsus with proportionally smaller canal for tendon of musculus flexor digitorum longus (); the distal end of the tarsometatarsus differs from a distal tarsometatarsus that was considered by Mayr and Smith (Citation2019) to possibly be from A. aurorum in that the trochleae are more splayed. Distinguished from Percolinus venablesi Harrison and Walker, Citation1977 (fam. inc. sed.) in that tuberositas musculi tibialis cranialis less pronounced and in that hypotarsus proximodistally shorter (). Differs from “Argillipes” paralectoris and “Percolinus” proudlocki in that hypotarsus with canal for tendon of musculus flexor digitorum longus (this canal is absent in “A.” paralectoris and “P.” proudlocki; ). Differentiated from Coturnipes cooperi Harrison and Walker, Citation1977 (fam. inc. sed.) in that trochlea metatarsi IV more laterally splayed, incisura intertrochlearis lateralis wider, and plantar articular surface of trochlea metatarsi III less asymmetric (). Distinguished from Litoripes medius Harrison and Walker, Citation1979 (fam. inc. sed.) in that tarsometatarsus with proportionally wider distal end and trochlea metatarsi IV reaching farther distally. The proximal tarsometatarsus referred to B. transitoria by Zelenkov (Citation2021) differs from that of the new species in being smaller and in that crista medialis hypotarsi not as strongly plantarly protruding. ?Paraortygoides argillae, sp. nov. is distinguished from Bumbanipodius magnus Zelenkov, Citation2021 (fam. inc. sed.) in that tarsometatarsus without laterally prominent tubercle for ligamentum collaterale mediale. It can be differentiated from the species of Quercymegapodius Mourer-Chauviré, Citation1992 (Quercymegapodiidae) in that foramen vasculare distale situated farther proximally, plantar articular surface of trochlea metatarsi proximodistally longer than mediolaterally wide (about the same length and width in Quercymegapodius), and in that trochleae metatarsorum II and IV proportionally longer. Differs from Ameripodius alexis Mourer-Chauviré, Citation2000 (Quercymegapodiidae) in that hypotarsus with intermediate crest less developed (see Zelenkov, Citation2021: fig. 2l), and in that foramen vasculare distale situated farther proximally (comparisons with Ameripodus silvasantosi Alvarenga, Citation1995 are not possible owing to a lack of overlap in the known skeletal elements). Distinguished from the species of the Paraortygidae in that trochleae metatarsorum of tarsometatarsus more splayed. Differs from Chambiortyx cristata Mourer-Chauviré et al., Citation2013, in that trochlea metatarsi III dorsoplantarly deeper. Distinguished from Namaortyx sperrgebietensis Mourer-Chauviréet al., 2011, in that tarsometatarsus proportionally more elongated. Differentiated from Scopelortyx klinghardtensis Mourer-Chauviré et al., Citation2015, in that distal end of femur less excavated by sulcus intercondylaris, and proximal portion of condylus medialis more gradually merging with shaft.
Meaningful comparisons with Tegulavis corbalani Mourer-Chauviré et al., 2023 (in Mourer-Chauviré et al., Citation2024) are not possible owing to a lack of overlap in the known bones; however, with a length of 41.7 mm (Mourer-Chauviré et al., Citation2024), the coracoid of T. corbalani is much too large to belong to the new species.
Etymology—The species epithet is derived from argilla (Lat.), clay.
Type Locality and Horizon—Walton-on-the-Naze, Essex, U.K.; Walton Member of the London Clay Formation (previously Division A2; Aldiss, Citation2012; Jolley, Citation1996; Rayner et al., Citation2009), early Eocene (early Ypresian, 54.6–55 Ma; Collinson et al., Citation2016).
Measurements (Maximum Length, in mm)—Left tarsometatarsus, estimated length, ∼31.6.
Remarks—Derived features of galliforms shown by the holotype of ?Paraortygoides argillae, sp. nov. include an asymmetric plantar articular surface of the trochlea metatarsi III and a characteristic hypotarsus morphology (Mayr, Citation2016).
Description and Comparisons—Of the femur, only the proximal and distal ends are preserved in the holotype. These show morphologies that agree with other stem group Galliformes. The crista trochanteris, on the proximal end of the bone, is well developed. The distal end exhibits a deeply incised sulcus patellaris and a mediolaterally wide condylus medialis.
The holotype includes the distal end of the tibiotarsus. The condyles are of similar size, and as in some extant Galliformes (e.g., Alectura lathami, Megapodiidae) the condylus medialis is inclined relative to the longitudinal axis of the bone, so that the medial margin of the distal tibiotarsus forms a marked kink. The area of the pons supratendineus is covered by sediment encrustation.
The tarsometatarsus resembles that of the species of the Gallinuloididae and Quercymegapodiidae in that the trochleae are more splayed than in crown group Galliformes. The impressiones retinaculi extensorii, on the proximal end of the bone, are poorly marked. The hypotarsus exhibits a canal for the tendon of musculus flexor digitorum longus, which is proportionally smaller than the hypotarsal canal of Argillipes aurorum (). The plantar hypotarsal surface bears two sulci for the flexor tendons of the second toe and for the tendon of musculus hallucis longus (Mayr, Citation2016). The hypotarsus corresponds with that of Bumbanortyx and Bumbanipodius, but in contrast to B. magnus the proximal end of the tarsometatarsus lacks a laterally prominent tubercle for the ligamentum collaterale mediale. On the distal end of the bone the dorsal opening of the foramen vasculare distale is situated in a deep fossa. The trochlea metatarsi IV reaches farther distally than the trochlea metatarsi II. As in other Galliformes (Mayr, Citation2000), the plantar articular surface of the trochlea metatarsi III is asymmetric, with the lateral portion reaching farther proximally than the medial one. A canalis interosseus distalis is present. The distal end of the bone differs distinctly from a distal tarsometatarsus from the lower Eocene of Egem in Belgium (), which was tentatively referred to Argillipes by Mayr and Smith (Citation2019), in that the trochleae are more splayed, the trochlea metatarsi II less plantarly deflected, the trochlea metatarsi III proportionally smaller, and the trochlea metatarsi IV more strongly dorsally raised. The distal tarsometatarsus of NMS.Z.2021.40.174 resembles that of Gallinuloides wyomingensis in its overall shape, but differs in that the dorsal articular surface of the trochlea metatarsi III does not reach as far proximally (). Compared with the larger-sized “Paraortygoides” radagasti (), the trochlea metatarsi II does not reach as far distally.
The first phalanx of the second toe slightly exceeds the first phalanx of the third toe in length. The relative lengths of the ungual phalanges are between those of Gallinuloides wyomingensis and Paraortygoides messelensis ().
WALTONORTYGIDAE, fam. nov.
Type Genus—Waltonortyx, gen. nov.
Differential Diagnosis—Small stem group Galliformes with a deeply excavated cotyla scapularis (coracoid) of circular shape. This feature distinguishes the new taxon from the Gallinuloididae as exemplified by the type genus Gallinuloides, in which the cotyla scapularis has an ellipsoid outline. The proximal humerus has a distinctive morphology and is distinguished from that of the Gallinuloididae (as exemplified by Paraortygoides messelensis; the feature is not observable in Gallinuloides), Quercymegapodiidae, Paraortygidae, and crown group Galliformes in a proximodistally narrower caput humeri and a shorter and wider tuberculum dorsale, which reaches only slightly farther distally than the tuberculum ventrale.
WALTONORTYX, gen. nov.
Type Species—Waltonortyx bumbanipodiides, sp. nov.
Differential Diagnosis—In addition to the above-listed features that distinguish the Waltonortygidae, fam. nov. from the Gallinuloididae, Quercymegapodiidae, Paraortygidae, and crown group Galliformes, the new taxon Waltonortyx, gen. nov. is distinguished from Gallinuloides Eastman, Citation1900, and Paraortygoides Mayr, Citation2000, in that the crista deltopectoralis is less dorsally extensive. Differs from a coracoid referred to Bumbanipodius by Zelenkov (Citation2021) in that medial surface of processus acrocoracoideus (that is, the facies articularis clavicularis) forming a medial protrusion (flatter in B. magnus), facies articularis humeralis (coracoid) more laterally projected and with more convex outline (in dorsoventral view), and in that cotyla scapularis proportionally smaller. Distinguished from Bumbanortyx Zelenkov, Citation2021, in that cotyla scapularis (coracoid) with circular outline (ellipsoid in Bumbanortyx) and processus procoracoideus less developed. Differs from Tegulavis Mourer-Chauviré, Bourdon, Duffaud, Le Roux, and Laurent, 2023, in that facies articularis humeralis of coracoid less laterally projected and sternal margin of crista articularis sternalis markedly concave (straight in Tegulavis). Distinguished from Scopelortyx Mourer-Chauviré, Pickford, and Senut, Citation2015, in that coracoid with more deeply excavated cotyla scapularis, humerus with proximodistally narrower caput humeri, and second (dorsal) fossa pneumotricipitalis shallower.
Direct comparisons with the taxon Argillipes are impeded by the lack of overlap in the known elements (only proximal tarsometatarsi and a referred distal humerus are known from A. aurorum, the type species of the taxon), but compared with a specimen that was likened to A. aurorum by Mayr and Smith (Citation2019), the tuberculum dorsale of the humerus is proximodistally shorter and mediolaterally wider, and the caput humeri is proximodistally somewhat longer.
Comparisons with Namaortyx Mourer-Chauviré et al., Citation2011 and Chambiortyx Mourer-Chauviré et al., Citation2013 are not possible owing to a lack of overlap in the known skeletal elements.
Etymology—The name is derived from the type locality and ortyx (Gr.), which denotes a quail. The name is feminine in gender.
WALTONORTYX BUMBANIPODIIDES, gen. et sp. nov.
(, , and )
Holotype—NMS.Z.2021.40.175 (; thoracic vertebra, right coracoid, fragmentary proximal end of right humerus, left phalanx proximalis digiti majoris); collected in 1981 by M. Daniels (original collector’s number WN 81315A).
Diagnosis—As for genus. The species is distinctly smaller than ?Paraortygoides argillae, sp. nov.
Etymology—The species epithet refers to the resemblance of the coracoid of the new species to that of the taxon Bumbanipodius from the lower Eocene of Mongolia.
Type Locality and Horizon—Walton-on-the-Naze, Essex, U.K.; Walton Member of the London Clay Formation (previously Division A2), early Eocene (early Ypresian).
Measurements (Maximum Length, in mm)—Right coracoid, 22.5.
Description and Comparisons—The thoracic vertebra exhibits distinct fossae on the lateral surfaces of the corpus. This is a plesiomorphic trait of neornithine birds and likely to be due to a lesser degree of skeletal pneumatization (Mayr, Citation2021a).
The coracoid () agrees with that of Bumbanipodius magnus from the lower Eocene of Mongolia (), in that the short processus acrocoracoideus is only weakly medially projected. Unlike in most other stem and crown group Galliformes, the extremitas omalis therefore does not have a hook-like shape (an exception is, for example, the extant phasianid species Bambusicola thoracicus; see Zelenkov, Citation2016:fig. 1c). As in other galliforms, the impressio ligamenti acrocoracohumeralis is marked. The cotyla scapularis is deeply excavated and has a circular outline. There is an incipient incisura nervi supracoracoidei on the medial margin of the shaft, which is not known from other unequivocal stem group Galliformes (however, Danielsavis nazensis, which was considered to be a possible stem group galliform by Mayr et al., [Citation2023] exhibits a foramen nervi supracoracoidei). The extremitas sternalis forms a medial projection and its dorsal surface bears muscle striae. The processus lateralis is better developed than in extant Galliformes and tapers into a pointed projection; in Scopelortyx from the middle Eocene of Namibia this process is likewise shorter and less pointed (Mourer-Chauviré et al., Citation2017). The bone agrees in size with a coracoid from the lower Eocene of Belgium (), which was likened to Argillipes aurorum by Mayr and Smith (Citation2019), but, owing to the poor preservation of the latter specimen, detailed comparisons are not possible.
FIGURE 3. Coracoids and scapulae of stem group Galliformes from the Paleogene of Europe, Asia, and North America. A, B, Waltonortyx bumbanipodiides, gen. et sp. nov. from Walton-on-the-Naze, right coracoid (holotype, NMS.Z.2021.40.175) in ventral (A) and dorsal (B) view; the arrow denotes a detail of the extremitas omalis. C, D, undetermined galliform from Walton-on-the-Naze (NMS.Z.2021.40.176), right coracoid in ventral (C) and dorsal (D) view; the arrow denotes a detail of the extremitas omalis. E, Bumbanipodius magnus from the lower Eocene of Mongolia (PIN 3104/195), omal extremity of left coracoid in dorsal view. F, Bumbanortyx transitoria from the lower Eocene of Mongolia (holotype, PIN 3104/265), omal extremity of left coracoid in dorsal view. G, Quercymegapodius brodkorbi (Quercymegapodiidae) from the lower Eocene of France (UM BFI 1849), omal extremity of left coracoid in dorsal view. H, Gallinuloides wyomingensis (Gallinuloididae) from the lower Eocene Green River Formation (WDC CGR−012), left coracoid in dorsal view. I, Paraortygoides messelensis (Gallinuloididae) from the latest early or earliest middle Eocene of Messel (holotype, SMF-ME 1303a), left coracoid in dorsal view; the specimen was coated with ammonium chloride. J, P. messelensis (SMF-ME 11112a), right coracoid in ventral view; the specimen was coated with ammonium chloride. K, cf. Argillipes aurorum from the lower Eocene of Egem in Belgium (IRSNB Av 167), right coracoid in dorsal view; the specimen was coated with ammonium chloride. L, Paraortyx brancoi (Paraortygidae) from the lower Oligocene of Belgium (IRSNB Av 116a), left coracoid in dorsal view; the specimen was coated with ammonium chloride. M, undetermined galliform from Walton-on-the-Naze (NMS.Z.2021.40.176), right scapula in lateral view. N, P. messelensis (holotype, SMF-ME 1303a), cranial extremity of left scapula in lateral view; the specimen was coated with ammonium chloride. Abbreviations: acr, acromion; agm, angulus medialis; cas, crista articularis sternalis; csc, cotyla scapularis; fah, facies articularis humeralis; inc, incisura nervi supracoracoidei; mpr, medial projection; pac, processus acrocoracoideus; pla, processus lateralis; ppc, processus procoracoideus; str, muscle striae; tbc, tuberculum coracoideum. The scale bars equal 5 mm.
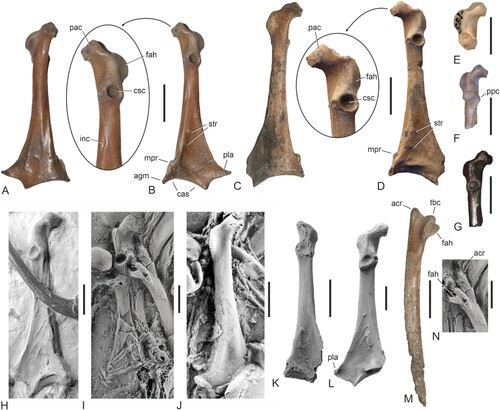
The proximal end of the humerus () resembles a similar-sized proximal humerus from the lower Eocene of Mongolia () that was referred to Bumbanortyx transitoria by Zelenkov (Citation2021), with which it shares a proximodistally narrow caput humeri and a short tuberculum dorsale that reaches only slightly farther distally than the tuberculum ventrale (much farther distally in most other galliforms). The humerus of NMS.Z.2021.40.175 is also similar to an early Eocene proximal humerus from Egem in Belgium (), which was tentatively assigned to Argillipes aurorum by Mayr and Smith (Citation2019). However, in the latter specimen the tuberculum dorsale is somewhat longer and mediolaterally narrower, and the caput humeri is wider. The humeri of all other galliforms also have a wider caput humeri and a longer and wider tuberculum dorsale. There is a second (dorsal) fossa pneumotricipitalis, which is shallower than in the humerus from Egem, in which it undercuts the caput humeri and is dorsally more excavated. The primary fossa pneumotricipitalis is larger than in the Egem humerus. Unlike in Gallinuloides wyomingensis () and Paraortygoides messelensis (), the crista deltopectoralis has little dorsal extent.
FIGURE 4. Humeri of Eocene stem group Galliformes. A, Waltonortyx bumbanipodiides, gen. et sp. nov. from Walton-on-the-Naze, proximal end of right humerus (holotype, NMS.Z.2021.40.175) in cranial view. B, cf. Argillipes aurorum from the lower Eocene of Egem in Belgium (IRSNB Av 163), proximal end of right humerus in cranial view; the specimen was coated with ammonium chloride. C, Paraortygoides messelensis from the latest early or earliest middle Eocene of Messel (SMF-ME 11112a), proximal end of right humerus is cranial view; the specimen was coated with ammonium chloride. D, Gallinuloides wyomingensis from the lower Eocene Green River Formation (WDC CGR−012), proximal end of left humerus is cranial view. E, W. bumbanipodiides, gen. et sp. nov., proximal end of right humerus in caudal view. F, proximal left humerus from the early Eocene of Mongolia that was referred to Bumbanortyx transitoria by Zelenkov (Citation2021) (PIN 3104/128), caudal view. G, cf. Argillipes aurorum from Egem (IRSNB Av 163), proximal end of right humerus in caudal view. H, P. messelensis (holotype, SMF-ME 1303a), proximal end of left humerus in caudal view; the specimen was coated with ammonium chloride. I, Paraortyx lorteti from the lower Eocene of Belgium (IRSNB Av 115), proximal end of right humerus in caudal view. J, K, undetermined galliform from Walton-on-the-Naze, from the London Clay of Walton-on-the-Naze (NMS.Z.2021.40.176), distal portion of a right humerus in caudal (J) and cranial (K) view. L, Argillipes aurorum from the London Clay of the Isle of Sheppey (NHMUK A 4282), distal portion of left humerus (mirrored to ease comparisons) in cranial view. M, G. wyomingensis (WDC CGR−012), distal end of left humerus in cranial view. N, P. messelensis (SMF-ME 11112a), distal end of right humerus is cranial view; the specimen was coated with ammonium chloride. The arrows in E–I denote the distal terminus of the tuberculum dorsale. In A and B the humeri are shown to scale, in E–I they are brought to the same size. Abbreviations: cdp, crista deltopectoralis; cph, caput humeri; flx, processus flexorius; fpt, second (dorsal) fossa pneumotricipitalis; tbd, tuberculum dorsale; tbv, tuberculum ventrale. The scale bars equal 5 mm.
The phalanx proximalis digiti majoris is short and craniocaudally wide, which indicates a broad and rounded wing.
Referred Specimen—NMS.Z.2021.40.176 (; right coracoid, right scapula, right humerus lacking proximal end); collected in 1992 by M. Daniels (original collector’s number WN 92715).
Type Locality and Horizon—Walton-on-the-Naze, Essex, U.K.; Walton Member of the London Clay Formation (previously Division A2), early Eocene (early Ypresian).
Measurements (Maximum Length, in mm)—Right coracoid, maximum length, 25.0; length from angulus medialis to tip of processus acrocoracoideus, 23.8.
Remarks—Even if a sexual dimorphism in size is accounted for, the bones of NMS.Z.2021.40.176 are much too small to belong to Paraortygoides radagasti. The coracoid () is larger than that of Waltonortyx bumbanipodiides, gen. et sp. nov. and differs in the shape of the processus acrocoracoideus. Based on comparisons with the paraortygid Paraortyx lorteti—the coracoid and tarsometatarsus of which measure 24.7 and 29.3–36.8 mm, respectively (Mourer-Chauviré, Citation1992) —the length of the coracoid of NMS.Z.2021.40.176 would correspond to that of the tarsometatarsus of ?Paraortygoides argillae, sp. nov. (NMS.Z.2021.40.174). By contrast, in Gallinuloides wyomingensis, the ratio of tarsometatarsus length to coracoid length is 1.15, whereas it is ∼1.32 for the tarsometatarsus of ?P. argillae (NMS.Z.2021.40.174) and the coracoid of NMS.Z.2021.40.176. Compared with the tarsometatarsus of the ?P. argillae holotype, the coracoid of NMS.Z.2021.40.176 is therefore proportionally shorter than that of G. wyomingensis. However, crown group Galliformes exhibit pronounced sexual dimorphism in size, and stem group galliforms also show considerable intraspecific variation in the lengths of the major skeletal elements (Mourer-Chauviré, Citation1992). Therefore, NMS.Z.2021.40.176 possibly represents a female of ?P. argillae. On the other hand, the coracoid of the specimen is distinguished from that of gallinuloidids in the circular shape of the cotyla scapularis and, as noted below, the distal humerus of NMS.Z.2021.40.176 shows a close resemblance to a humerus referred to Argillipes aurorum by Harrison and Walker (Citation1977).
Description and Comparisons—The coracoid has a deeply excavated, concave cotyla scapularis with a circular outline. It differs from the somewhat shorter coracoid of Waltonortyx bumbanipodiides, gen. et sp. nov., in that the processus acrocoracoideus is more medially protruding and in that the facies articularis humeralis is proportionally longer and less strongly laterally projected. In its overall morphology the omal extremity resembles that of Bumbanortyx transitoria from the lower Eocene of Mongolia (Zelenkov, Citation2021), but the processus procoracoideus of NMS.Z.2021.40.176 is less developed than in B. transitoria () and the cotyla scapularis has a circular outline (ellipsoid in Bumbanortyx). The omal extremity of NMS.Z.2021.40.176 also resembles that of another, unnamed, stem group galliform from the lower Eocene of Mongolia (Hood et al., Citation2019), in which the medial surface of the processus acrocoracoideus is somewhat straighter. This latter specimen was tentatively assigned to the Quercymegapodiidae and we also regard the coracoid of NMS.Z.2021.40.176 to be similar to that of Quercymegapodius (). The bone differs from the coracoid of Gallinuloides (), in that the cotyla scapularis is more deeply excavated and, unlike in Gallinuloides and Paraortygoides (), the cotyla scapularis has a circular (rather than ellipsoid) outline. In Bumbanipodius () the processus acrocoracoideus is less strongly medially protruding. The coracoid of Tegulavis has a more laterally projected facies articularis humeralis and the sternal margin of the crista articularis sternalis is straight. In the Paraortygidae () and other stem group Galliformes, the cotyla scapularis also has an ellipsoid outline.
The scapula () exhibits a prominent tuberculum coracoideum, which is better developed than in, e.g., Scopelortyx (in crown group Galliformes it is obsolete, which is functionally related to the flat facies articularis scapularis of the coracoid). The acromion is proportionally shorter and wider than in the paraortygid taxa Paraortyx (Mourer-Chauviré, Citation1992:fig. 2i) and Scopelortyx (Mourer-Chauviré et al., Citation2015:fig. 5). In crown group Galliformes the acromion is likewise more elongated, a tuberculum coracoideum is virtually absent, and the facies articularis humeralis is craniocaudally much narrower. As in other stem group Galliformes (e.g., Mayr, Citation2000), the shaft of the bone is narrow and straight (in crown group Galliformes other than the Odontophorinae and Phasianinae it is wider and more angled); its caudal portion is broken.
The size and morphology of the distal humerus () closely resembles a distal humerus from the Isle of Sheppey (), which was referred to Argillipes aurorum by Harrison and Walker (Citation1977). Unfortunately, this resemblance is difficult to contextualize, because there exists no basis for an unequivocal identification of the specimen from the Isle of Sheppey. Compared with the distal humerus of Gallinuloides wyomingensis (), the processus flexorius of NMS.Z.2021.40.176 is more protruding.
RESULTS OF THE PHYLOGENETIC ANALYSIS
Analysis of the character matrix in the appendix with PAUP*4.0a169 yielded 75 most parsimonious trees (L = 55, CI = 0.65, RI = 0.80), the strict consensus tree of which was poorly resolved (). However, the analysis did not support monophyly of the Quercymegapodiidae and recovered Quercymegapodius and Ameripodius as successive sister taxa of crown group Galliformes.
FIGURE 5. Results of the phylogenetic analysis. A, strict consensus tree of 75 most parsimonious trees (L = 55, CI = 0.65, RI = 0.80); bootstrap support is given next to the internodes. B, majority rule consensus tree; the numbers next to the internodes indicate the percentage of replications in which the node was retained.
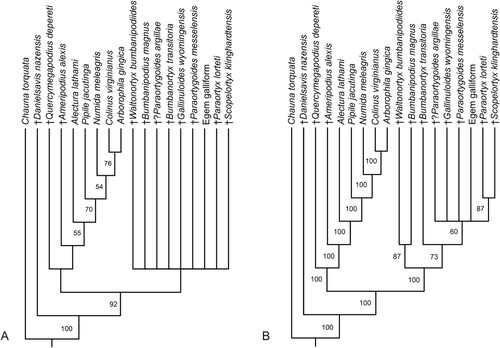
This topology was also retained in the majority rule consensus tree (), which furthermore supported a sister group relationship between Bumbanipodius magnus and Waltonortyx bumbanipodiides. The majority rule consensus tree also recovered a clade including Paraortygoides messelensis, Gallinuloides wyomingensis, Bumbanipodius magnus, Waltonortyx bumbanipodiides, Bumbanortyx transitoria, the Egem galliform and the paraortygid species Paraortyx lorteti and Scopelortyx klinghardtensis.
DISCUSSION
Phylogenetic Affinities and Evolutionary Significance of the New Fossils from Walton-on-the-Naze—In its composition, the avifauna from the London Clay closely corresponds to that of the slightly younger locality Messel in Germany and that of the North American Green River Formation. Shared taxa include, among others, the Rhynchaeitinae (Mayr & Kitchener, Citation2023a), Masillaraptoridae (Mayr & Kitchener, Citation2021), stem group Leptosomiformes (Mayr & Kitchener, Citation2023b), as well as various taxa that belong to the Psittacopasseres (Mayr & Kitchener, Citation2023c, Citationd, Citation2024). Therefore, it would be expected that representatives of the Gallinuloididae, which are well documented in Messel and the Green River Formation, also occurred in the London Clay. As detailed below, the affinities of “Paraortygoides” radagasti are uncertain, and the assignment of this species to the taxon Paraortygoides was not well supported. We identified another fossil that may fill the role of the “London Clay gallinuloidid,” but even though the leg bones of the holotype of ?Paraortygoides argillae, sp. nov. appear to be very similar to those of Paraortygoides messelensis, our assignment to the taxon Paraortygoides is tentative and only based on overall similarity. ?P. argillae was not recovered together with P. messelensis in the phylogenetic analysis, but the characters that support a clade to the exclusion of ?P. argillae are actually not preserved in the holotype of the new species.
The second new galliform from Walton-on-the-Naze, Waltonortyx bumbanipodiides, gen. et sp. nov. is distinguished from the Gallinuloididae (as exemplified by Paraortygoides messelensis; ), Quercymegapodiidae, Paraortygidae () and crown group Galliformes in the proximodistally narrow caput humeri and short tuberculum dorsale (which reaches only slightly farther distally than the tuberculum ventrale) of the humerus. Outgroup comparisons with the Anseriformes and other neornithine birds suggest that at least the short tuberculum dorsale of Waltonortyx is plesiomorphic, which may indicate that the taxon falls outside a clade formed by gallinuloidids, quercymegapodiids, paraortygids, and crown group galliforms. However, this position conflicts with the plesiomorphic shape of the dorsally prominent crista deltopectoralis of gallinuloidids (), which resembles that of anseriforms (Mayr, Citation2006) and is absent in Waltonortyx.
Another characteristic feature of Waltonortyx bumbanipodiides is the circular shape of the cotyla scapularis of the coracoid, which also occurs in Quercymegapodius (), Ameripodius, and Bumbanipodius (). In Bumbanortyx () and other stem group galliforms the cotyla scapularis has an ellipsoid outline, and in Gallinuloides () and the Paraortygidae () it is less deeply excavated. Again, outgroup comparisons with the Anseriformes suggest that a deeply concave cotyla scapularis with a circular (rather than ellipsoid) outline is plesiomorphic for the Galliformes. Our analysis did not resolve the position of Waltonortyx relative to gallinuloidids and other stem group galliforms, and more data on the skeletal morphology of the Waltonortygidae are required for a robust placement of the new taxon.
The humerus of W. bumbanipodiides shows a close resemblance to humeri from the lower Eocene of Mongolia that were described by Hwang et al. (Citation2010) and Zelenkov (Citation2021). These humeri () were referred to Bumbanortyx transitoria by Zelenkov (Citation2021) and derive from the Tsagaan-Khushuu locality, which also yielded the holotype of Bumbanipodius magnus. As noted above, the coracoid of this latter species is more similar to that of W. bumbanipodiides than is the coracoid of B. transitoria, in which the cotyla scapularis is ellipsoid, the processus acrocoracoideus more medially projected, and the processus procoracoideus better developed. This raises the question of whether the humeri from Tsagaan-Khushuu may actually belong to B. magnus rather than B. transitoria, and although the holotype coracoid of B. magnus appears to be somewhat too large for the humeri referred to B. transitoria (see Zelenkov, Citation2021), the species may have shown sexual dimorphism in size. For the phylogenetic analysis, we followed Zelenkov (Citation2021) and scored the humeri for B. transitoria, which was placed in a clade together with gallinuloidids and paraortygids. This position was supported by the ellipsoid shape of the cotyla scapularis, which constitutes very weak evidence, because the distribution of this feature shows homoplasy in Galliformes and also characterizes the crown group representatives. If the Mongolian humeri indeed belong to Bumbanortyx (rather than Bumbanipodius), we consider it likely that the former is also a representative of the Waltonortygidae, fam. nov. In any case, the close similarity between the galliform taxa from Walton-on-the-Naze and those from the Bumbanian locality Tsagaan Khushuu in Mongolia suggests dispersal events across the Turgai Strait, which separated Europe and Asia in the early Paleogene (see, e.g., Hooker and Dashzeveg [Citation2003] for biogeographic connections in the mammalian faunas).
The humerus of W. bumbanipodiides also exhibits a similar morphology to that of a galliform from Egem, which was tentatively assigned to Argillipes aurorum by Mayr and Smith (Citation2019). However, in the fossil from Egem () the caput humeri is proximodistally somewhat wider, the tuberculum dorsale is proportionally somewhat longer and mediolaterally narrower, and the dorsal fossa pneumotricipitalis is deeper in its dorsal portion. These differences also distinguish the Egem galliform from the Mongolian humerus that was referred to Bumbanortyx by Zelenkov (Citation2021). Overall, the Egem galliform is very similar to Scopelortyx klinghardtensis from the late middle Eocene of Namibia and our analysis also suggests that the Belgian species is more closely related to Paraortyx than to Waltonortyx.
Our analysis furthermore recovered a clade including Scopelortyx, Paraortyx, and the Egem galliform, as well as the Gallinuloididae, Waltonortyx and the Mongolian taxa (). However, this clade was only retained in the majority rule consensus tree and the single apomorphy supporting it is the presence of a second (dorsal) fossa pneumotricipitalis. This fossa is poorly developed in Waltonortyx bumbanipodiides and the Mongolian humeri assigned to Bumbanortyx transitoria, and its presence cannot be verified for Gallinuloides wyomingensis, because the two known specimens of this taxon do not expose the caudal surface of the proximal humerus. Moreover, the occurrence of a dorsal fossa pneumotricipitalis shows homoplasy within Galliformes and also evolved in some crown group taxa (Odontophorinae and early diverging Phasianinae). Thus, the evidence for a clade including Bumbanortyx, Bumbanipodius, Waltonortyx, as well as the Gallinuloididae and Paraortygidae, is weak. The wider caput humeri and longer tuberculum dorsale may indicate that Scopelortyx, Paraortyx, and the Egem galliform are more closely related to crown group Galliformes than is Waltonortyx, but the currently known material does not allow an unequivocal assessment of the affinities of these birds. Therefore, we consider the affinities of the Waltonortygidae to be unresolved relative to the Gallinuloididae and Paraortygidae (). A second fossa pneumotricipitalis is absent in the poorly known taxon Pirortyx, which was assigned to the Paraortygidae by Mourer-Chauviré (Citation1992), but is here regarded to be of uncertain affinities.
FIGURE 6. Temporal occurrences of early Paleogene stem group galliforms (gray bars) and our preferred tentative hypothesis on their interrelationships.
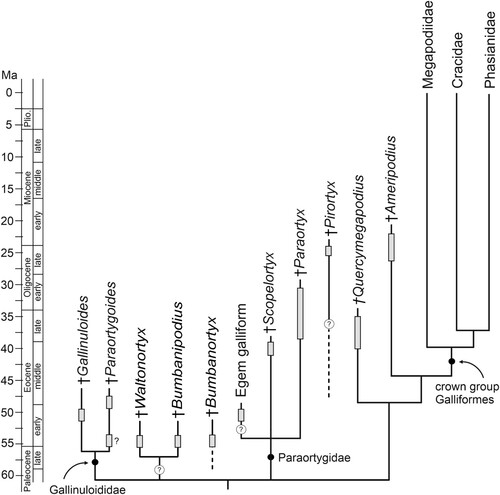
The position of the late Oligocene/Early Miocene Ameripodius, which was shown to be the sister taxon of crown group Galliformes in our analyses, is of particular interest. Ameripodius differs from late Eocene species of Quercymegapodius in that the ridge in the incisura capitis of the humerus is much more developed and in that the sternal extremity of the coracoid exhibits a large pneumatic opening, which also occurs in the Megapodiidae and Cracidae (although only the Early Miocene A. alexis was included in our analysis, the late Oligocene/Early Miocene Ameripodius silvasantosi, the type species of the taxon Ameripodius [see Alvarenga, Citation1995], exhibits the same characteristics of the humerus and coracoid). We hypothesize that the pneumatic opening of the coracoid is an apomorphy of crown group Galliformes and was secondarily lost in the Phasianidae. An assignment of Ameripodius to the Quercymegapodiidae was largely based on overall similarity of the bones (Alvarenga, Citation1995; Mourer-Chauviré, Citation2000), but the above-mentioned derived features shared with extant galliforms suggest that Ameripodius is more closely related to crown group Galliformes than are the species of Quercymegapodius.
Judging from the similar morphology of the coracoid (which resembles that of Quercymegapodius), it is possible that the wing bones of the undetermined specimen NMS.Z.2021.40.176 belong to the Quercymegapodiidae. However, such an assignment cannot be established based on shared derived characteristics, and the affinities between NMS.Z.2021.40.176 and the Waltonortygidae likewise remain elusive. There are no definitive records of quercymegapodiids from early Eocene sites, but Hood et al. (Citation2019) tentatively assigned a fragmentary coracoid from the lower Eocene of Mongolia to this taxon, and Zelenkov (Citation2021) also noted some similarities between the galliform fossils from the lower Eocene of Mongolia and quercymegapodiids. Crown group Galliformes have a flat facies articularis scapularis, and in galliforms and other avian lineages flattening of the scapular cotyla appears to have been related to the reduction of the furcular shafts (Mayr, Citation2021b). In the Gallinuloididae the furcular shafts are particularly robust, and it was hypothesized that these birds lacked the large crop that characterizes crown group Galliformes (Mayr, Citation2006).
Finally, we note that all currently known early Eocene Galliformes are relatively small (Coturnix- to Perdix-sized) and that no larger-sized galliforms are known from this epoch. This sharply contrasts with the size spectrum of extant Galliformes, which include very large species in all three major clades (Megapodiidae, Cracidae, and Phasianidae). Therefore, it is probable that the stem species of Galliformes was also a rather small bird, and ecological factors that favored size increase within the crown group—such as niche segregation, increased predation by larger carnivorans, or intraspecific competition—were less significant by the early Eocene.
The Affinities of “Paraortygoides” radagasti—“Paraortygoides” radagasti is only known from the holotype—a fragmentary partial skeleton ()—and a referred distal tarsometatarsus from Walton-on-the-Naze (the latter was not figured by Dyke and Gulas [Citation2002] and has not been examined by us). Even if sexual dimorphism in size is considered, the species is distinctly larger than the new galliforms described herein.
“Paraortygoides” radagasti differs from unequivocal Eocene stem group Galliformes in the morphology of the cranial extremity of the scapula, with the tuberculum coracoideum not being sharply delimited from the facies articularis humeralis and the latter having a broadly convex ventral margin (). “P.” radagasti is also distinguished from ?P. argillae in that the crista trochanteris on the proximal end of the femur is proportionally shorter and more strongly laterally protruding (). Unlike in ?P. argillae and other unequivocal Galliformes, the hypotarsus of “P.” radagasti lacks a canal for the tendon of musculus flexor digitorum longus (; however, a similar hypotarsus morphology to that in “P.” radagasti occurs in Danielsavis nazensis; see Mayr et al. [Citation2023:fig. 6]). The trochlea metatarsi II of “P.” radagasti reaches farther distally than in ?P. argillae, P. messelensis, and Gallinuloides wyomingensis.
Mayr (Citation2009) hypothesized that “P.” radagasti may be a synonym of one of the galliform species described from the Isle of Sheppey, but close examination does not support this hypothesis. Even though the hypotarsus of the species shows a resemblance to that of “Argillipes” paralectoris (), differences in the shape of the crista medialis, which is mediolaterally narrower in “A.” paralectoris, indicate a distinction at species level.
There are no unambiguously referable specimens of “P.” radagasti in the Daniels collection, but we identified four larger-sized coracoids that show a resemblance to the coracoid of galliforms and are cataloged as NMS.Z.2021.40.177 (original collectors number WN 86519), NMS.Z.2021.40.178 (WN 97963), NMS.Z.2021.40.179 (WN 92716), and NMS.Z.2021.40.180 (WN 95880; ). The coracoid is not preserved in the holotype of “P.” radagasti, but specimen NMS.Z.2021.40.177 includes the cranial extremities of both scapulae (), which—except for a somewhat larger size—resemble the partial scapula of the holotype of “P.” radagasti (). However, this similarity is not diagnostic enough for an unambiguous referral of the coracoids to “P.” radagasti.
Moreover, these coracoids show some distinct differences to those of unequivocal galliforms. In particular, the processus procoracoideus is proportionally longer than in any unambiguously identified galliform. The crista articularis sternalis is also proportionally longer than in Gallinuloides wyomingensis and most other stem group Galliformes, but an equally long albeit straighter crista occurs in Tegulavis corbalani from the lower Eocene of France. These differences challenge an assignment of the four coracoids to “P.” radagasti, and a further caveat on an assignment of these coracoids to “P.” radagasti arises from the fact that the bones also show a strong resemblance to the coracoid of the stem group ibis Rhynchaeites litoralis from Walton-on-the-Naze (), which was described by Mayr and Kitchener (Citation2023a). However, the cranial extremity of the scapula of NMS.Z.2021.40.177 is distinguished from the scapula of the holotype of R. litoralis () by a shorter acromion and in that the tuberculum coracoideum is less well delimited from the facies articularis humeralis.
Mayr and Kitchener (Citation2023a:427) already commented on the similarities between “P.” radagasti and R. litoralis. In addition to the features mentioned by these authors, we note that the hypotarsus of “P.” radagasti closely resembles that of the holotype of R. litoralis (). A derived galliform feature of “P.” radagasti is an asymmetric plantar articular facet of the trochlea metatarsi III, in which the lateral portion is proximodistally longer than the medial one. This morphology is also present in R. litoralis (Mayr & Kitchener, Citation2023a), but it is much more pronounced in “P.” radagasti. However, and as also detailed by Mayr and Kitchener (Citation2023a), other bones of “P.” radagasti and R. litoralis are clearly distinguishable from each other, with the processus extensorius of the carpometacarpus of “P.” radagasti being narrower, the crista trochanteris of the femur less protruding (), and the thoracic vertebrae having pleurocoels (deep fossae) rather than pneumatic openings (Mayr, Citation2021a).
FIGURE 7. Selected elements of the holotype of “Paraortygoides” radagasti (NHMUK A 6217) in comparison to the holotype of ?Paraortygoides argillae, sp. nov. (NMS.Z.2021.40.174) and Rhynchaeites litoralis, a stem group representative of the Threskiornithidae from Walton-on-the-Naze; the dashed frame highlights partial scapulae and coracoids from Walton-on-the-Naze that may belong to “P.” radagasti or a closely related bird but also show a close resemblance to the ibis Rhynchaeites. A, “Paraortygoides” radagasti (holotype), cranial extremity of right scapula in lateral view. B, undetermined galliform from Walton-on-the-Naze (NMS.Z.2021.40.176), cranial extremity of right scapula in lateral view. C, “P.” radagasti (holotype), proximal end of left femur in cranial view. D, ?P. argillae (holotype), proximal end of left femur in cranial view. E, “P.” radagasti (holotype), distal end of right tarsometatarsus in plantar view. F, ?P. argillae (holotype), distal end of left tarsometatarsus (mirrored) in plantar view. G, “P.” radagasti (holotype), proximal end of right tarsometatarsus in proximal view. H, ?P. argillae (holotype), proximal end of left tarsometatarsus in proximal view. I, “P.” radagasti (holotype), cranial extremity of right scapula in lateral view. J, K, NMS.Z.2021.40.177, cranial extremities of both scapulae (J, lateral view) and left coracoid (K, dorsal view), which may belong to “P.” radagasti. L, NMS.Z.2021.40.180, left coracoid (dorsal view), which may belong to “P.” radagasti. M, N, NMS.Z.2021.40.179, partial right coracoid, which may belong to “P.” radagasti, in ventral (M) and dorsal (N) view. O, NMS.Z.2021.40.178, extremitas omalis of right coracoid (dorsal view), which may belong to “P.” radagasti. P, Q, Rhynchaeites litoralis (holotype, NMS.Z.2021.40.28), right coracoid (P, dorsal view) and cranial portion of left scapula (Q, medial view). R, S, fourth cervical vertebra (dorsal view) of “P.” radagasti (R: holotype) and R. litoralis (S: holotype, NMS.Z.2021.40.28). T, U, proximal portion of right carpometacarpus (ventral view) of “P.” radagasti (T: holotype) and R. litoralis (U: NMS.Z.2021.40.29). V, R. litoralis (holotype, NMS.Z.2021.40.28), proximal end of left femur in caudal view. W, R. litoralis (holotype, NMS.Z.2021.40.28), proximal end of left tarsometatarsus (mirrored) in proximal view. In E and F the trochleae are numbered. Abbreviations: acr, acromion; cas, crista articularis sternalis; csc, cotyla scapularis; ctr, crista trochanteris; fah, facies articularis humeralis; fdl, hypotarsal sulcus for tendon of musculus flexor digitorum longus; fhl, hypotarsal sulcus for tendon of musculus flexor hallucis longus; fp2/fpp2, sulcus for the tendons of musculus flexor perforatus digiti 2 and m. perforans et perforatus digiti 2; pex, processus extensorius; ppc, processus procoracoideus; tbc, tuberculum coracoideum. The scale bars equal 5 mm.

While we do not advocate close relationships between “P.” radagasti and R. litoralis, the similarities between both species challenge the presumed galliform affinities of the former, which, as detailed above, have not yet been convincingly established. Until more material can be unambiguously referred to “P.” radagasti, caution is advised in using this species to draw inferences about the evolution of galliforms.
AUTHOR CONTRIBUTIONS
GM analyzed the data and wrote the first draft of the manuscript. ACK contributed to the writing of the final version.
SUPPLEMENTARY FILES
Supplementary File 1. Character descriptions.
Supplementary File 2. Character matrix of 32 morphological characters for the 17 ingroup taxa included in this study.
Supplementary File 3. Nexus file used in the analysis with PAUP*4.0a169.
Supplemental Material
Download Zip (56.6 KB)DISCLOSURE STATEMENT
No potential conflict of interest was reported by the author(s).
DATA AVAILABILITY STATEMENT
The character descriptions and character matrix are available in the Supplementary Files 1–3. The nexus file used in the phylogenetic analysis was uploaded to the Science Data Bank (www.scidb.cn; doi: 10.57760/sciencedb.16556
).ACKNOWLEDGMENTS
We are indebted to Nikita Zelenkov for providing images of Bumbanortyx, Bumbanipodius, and Quercymegapodius and for permitting their use. We furthermore thank Sven Tränkner for taking some of the other photographs (additional images are by GM). We thank Trevor Worthy and an anonymous reviewer for comments that improved the paper.
LITERATURE CITED
- Aldiss, D. T. (2012). The stratigraphical framework for the Palaeogene successions of the London Basin, UK. British Geological Survey Open Report, OR/12/004, 1–87.
- Alvarenga, H. (1995). Um primitivo membro da ordem Galliformes (Aves) do Terciário Médio da Bacia de Taubaté, Estado de São Paulo, Brasil. Anais da Academia Brasileira de Ciências, 67(1), 33–44.
- Collinson, M. E., Adams, N. F., Manchester, S. R., Stull, G. W., Herrera, F., Smith, S. Y., Andrew, M. J., Kenrick, P., & Sykes, D. (2016). X-ray micro-computed tomography (micro-CT) of pyrite-permineralized fruits and seeds from the London Clay Formation (Ypresian) conserved in silicone oil: a critical evaluation. Botany, 94(9), 697–711. doi:10.1139/cjb-2016-0078
- Dyke, G. J., & Gulas, B. E. (2002). The fossil galliform bird Paraortygoides from the Lower Eocene of the United Kingdom. American Museum Novitates, 3360, 1–14. doi:10.1206/0003-0082(2002)360<0001:TFGBPF>2.0.CO;2
- Eastman, C. R. 1900. New fossil bird and fish remains from the Middle Eocene of Wyoming. Geological Magazine, decade 4, 7, 54–58. doi:10.1017/S0016756800181701
- Fischer, K. (1990). Der Hühnervogel Pirortyx major (Gaillard, 1939) aus dem marinen Mitteloligozän bei Leipzig (DDR). Mitteilungen aus dem Zoologischen Museum Berlin, 66, Supplement Annalen für Ornithologie, 14, 133–136.
- Harrison, C. J. O., & Walker, C. A. (1977). Birds of the British Lower Eocene. Tertiary Research Special Papers, 3, 1–52.
- Harrison, C. J. O., & Walker, C. A. (1979). Birds of the British Middle Eocene. In C. J. O. Harrison, & C. A. Walker (Eds.), Studies in Tertiary avian paleontology. Tertiary Research Special Papers, 5 (pp. 19–27.
- Hood, S. C., Torres, C. R., Norell, M. A., & Clarke, J. A. (2019). New fossil birds from the earliest Eocene of Mongolia. American Museum Novitates, 3934(3934), 1–24. doi:10.1206/3934.1
- Hooker, J. J., & Dashzeveg, D. (2003). Evidence for direct mammalian faunal interchange between Europe and Asia near the Paleocene-Eocene boundary. In S. L. Wing, P. D. Gingerich, B. Schmitz, & E. Thomas (Eds.), Causes and Consequences of Globally warm climates in the early Paleogene. Geological Society of America Special Paper, 369 (pp. 479–500.
- Houde, P., Dickson, M., & Camarena, D. 2023. Basal Anseriformes from the early Paleogene of North America and Europe. Diversity, 15(2), 233. doi:10.3390/d15020233
- Hwang, S. H., Mayr, G., & Minjin, B. (2010). The earliest record of a galliform bird in Asia, from the late Paleocene/early Eocene of the Gobi Desert, Mongolia. Journal of Vertebrate Paleontology, 30(5), 1642–1644. doi:10.1080/02724634.2010.501454
- Jolley, D. W. (1996). The earliest Eocene sediments of eastern England: an ultra-high resolution palynological correlation. Geological Society, London, Special Publications, 101(1), 219–254. doi:10.1144/GSL.SP.1996.101.01.14
- Lindow, B. E. K., & Dyke, G. J. (2007). A small galliform bird from the Lower Eocene Fur Formation. Bulletin of the Geological Society of Denmark, 55, 59–63. doi:10.37570/bgsd-2007-55-04
- Linnaeus, C. (1758). Systema naturae per regna tria naturae, 10th edition, 2 vols. L. Salmii.
- Lucas, F. A. (1900). Characters and relations of Gallinuloides wyomingensis Eastman, a fossil gallinaceous bird from the Green River shales of Wyoming. Bulletin of the Museum of Comparative Zoology, 36(1), 79–84.
- Mayr, G. (2000). A new basal galliform bird from the Middle Eocene of Messel (Hessen, Germany). Senckenbergiana lethaea, 80(1), 45–57. doi:10.1007/BF03043663
- Mayr, G. (2006). New specimens of the early Eocene stem group galliform Paraortygoides (Gallinuloididae), with comments on the evolution of a crop in the stem lineage of Galliformes. Journal of Ornithology, 147(1), 31–37. doi:10.1007/s10336-005-0006-8
- Mayr, G. (2009). Paleogene fossil birds (1st edition). Springer.
- Mayr, G. (2016). Variations in the hypotarsus morphology of birds and their evolutionary significance. Acta Zoologica, 97(2), 196–210. doi:10.1111/azo.12117
- Mayr, G. (2021a). On the occurrence of lateral openings and fossae (pleurocoels) in the thoracic vertebrae of neornithine birds and their functional significance. Vertebrate Zoology, 71, 453–463. doi:10.3897/vz.71.e71268
- Mayr, G. (2021b). The coracoscapular joint of neornithine birds - extensive homoplasy in a widely neglected articular surface of the pectoral girdle and its functional correlates. Zoomorphology, 140(2), 217–228. doi:10.1007/s00435-021-00528-2
- Mayr, G. (2022). Paleogene fossil birds (2nd edition). Springer.
- Mayr, G., & Kitchener, A. C. (2021). New fossils from the London Clay show that the Eocene Masillaraptoridae are stem group representatives of falcons (Aves, Falconiformes). Journal of Vertebrate Paleontology, 41(6), e2083515. doi:10.1080/02724634.2021.2083515
- Mayr, G., & Kitchener, A. C. (2023a). Multiple skeletons of Rhynchaeites from the London Clay reveal the osteology of early Eocene ibises (Aves, Threskiornithidae). PalZ, 97(2), 425–442. doi:10.1007/s12542-022-00647-1
- Mayr, G., & Kitchener, A. C. (2023b). New species from the early Eocene London Clay suggest an undetected early Eocene diversity of the Leptosomiformes, an avian clade that includes a living fossil from Madagascar. Palaeobiodiversity and Palaeoenvironments, 103(3), 585–608. doi:10.1007/s12549-022-00560-0
- Mayr, G., & Kitchener, A. C. (2023c). Psittacopedids and zygodactylids: The diverse and species-rich psittacopasserine birds from the early Eocene London Clay of Walton-on-the-Naze (Essex, UK). Historical Biology, 35(12), 2372–2395. doi:10.1080/08912963.2022.2141629
- Mayr, G., & Kitchener, A. C. (2023d). The Vastanavidae and Messelasturidae (Aves) from the early Eocene London Clay of Walton-on-the-Naze (Essex, UK). Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen, 307(2), 113–139. doi:10.1127/njgpa/2023/1119
- Mayr, G., & Kitchener, A. C. (2024). The Halcyornithidae from the early Eocene London Clay of Walton-on-the-Naze (Essex, UK): a species complex of Paleogene arboreal birds. Geobios, 83, 45–60. doi:10.1016/j.geobios.2023.06.003
- Mayr, G., & Smith, T. (2019). A diverse bird assemblage from the Ypresian of Belgium furthers knowledge of early Eocene avifaunas of the North Sea Basin. Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen, 291(3), 253–281. doi:10.1127/njgpa/2019/0801
- Mayr, G., & Weidig, I. (2004). The Early Eocene bird Gallinuloides wyomingensis - a stem group representative of Galliformes. Acta Palaeontologica Polonica, 49(3), 211–217.
- Mayr, G., De Pietri, V. L., & Scofield, R. P. (2022). New bird remains from the early Eocene Nanjemoy Formation of Virginia (USA), including the first records of the Messelasturidae, Psittacopedidae, and Zygodactylidae from the Fisher/Sullivan site. Historical Biology, 34(2), 322–334. doi:10.1080/08912963.2021.1910820
- Mayr, G., Carrió, V., & Kitchener, A. C. 2023. On the “screamer-like” birds from the British London Clay: An archaic anseriform-galliform mosaic and a non-galloanserine “barb-necked” species of Perplexicervix. Palaeontologia Electronica, 26(2), a3328; doi: 10.26879/1301.
- Mourer-Chauviré, C. (1988). Le gisement du Bretou (Phosphorites du Quercy, Tarn-et-Garonne, France) et sa faune de vertébrés de l’Eocène supérieur. II oiseaux. Palaeontographica A, 205, 29–50.
- Mourer-Chauviré, C. 1992. The Galliformes (Aves) from the Phosphorites du Quercy (France): systematics and biostratigraphy. In: K. E. Campbell (Ed.), Papers in Avian Paleontology Honoring Pierce Brodkorb. Natural History Museum of Los Angeles County, Science Series, 36, 67–95.
- Mourer-Chauviré, C. (2000). A new species of Ameripodius (Aves: Galliformes: Quercymegapodiidae) from the Lower Miocene of France. Palaeontology, 43(3), 481–493. doi:10.1111/j.0031-0239.2000.00136.x
- Mourer-Chauviré, C. 2006. The avifauna of the Eocene and Oligocene Phosphorites du Quercy (France): an updated list. Strata, Série 1(13), 13, 135–149.
- Mourer-Chauviré, C., Pickford, M., & Senut, B. (2011). The first Palaeogene galliform from Africa. Journal of Ornithology, 152(3), 617–622. doi:10.1007/s10336-010-0630-9
- Mourer-Chauviré, C., Tabuce, R., El Mabrouk, E., Marivaux, L., Khayati, H., Vianey-Liaud, M., & Ben Haj Ali, M. 2013. A new taxon of stem group Galliformes and the earliest record for stem group Cuculidae from the Eocene of Djebel Chambi, Tunisia. In: U. B. Göhlich & A. Kroh (Eds.), Paleornithological Research 2013 - Proceedings of the 8th International Meeting of the Society of Avian Paleontology and Evolution. Natural History Museum Vienna, Vienna, pp. 1–15.
- Mourer-Chauviré, C., Pickford, M., & Senut, B. (2015). Stem group galliform and stem group psittaciform birds (Aves, Galliformes, Paraortygidae, and Psittaciformes, family incertae sedis) from the Middle Eocene of Namibia. Journal of Ornithology, 156(1), 275–286. doi:10.1007/s10336-014-1124-y
- Mourer-Chauviré, C., Pickford, M., & Senut, B. (2017). New data on stem group Galliformes, Charadriiformes, and Psittaciformes from the middle Eocene of Namibia. Contribuciones del MACN, 7, 99–131.
- Mourer-Chauviré, C., Bourdon, E., Duffaud, S., Le Roux, G., & Laurent, Y. (2024). New avian remains from the early Eocene of La Borie, southern France. Geobios, 83, 61–84. doi:10.1016/j.geobios.2022.10.004
- Rayner, D., Mitchell, T., Rayner, M., & Clouter, F. (2009). London Clay fossils of Kent and Essex. Medway Fossil and Mineral Society.
- Shufeldt, R. W. (1915). A critical study of the fossil bird Gallinuloides wyomingensis Eastman. The Journal of Geology, 23(7), 619–634. doi:10.1086/622280
- Stidham, T. A., Townsend, K. E., & Holroyd, P. A. 2020. Evidence for wide dispersal in a stem galliform clade from a new small-sized Middle Eocene pangalliform (Aves: Paraortygidae) from the Uinta Basin of Utah (USA). Diversity, 12 (3), 90. doi:10.3390/d12030090
- Swofford, D. L. (2002). PAUP* Phylogenetic analysis using parsimony (*and other methods), version 4.0b10 [computer software]. Sinauer Associates.
- Temminck, C. J. (1820–1840). Manuel d’ornithologie, ou, Tableau systématique des oiseaux qui se trouvent en Europe: précédé d'une analyse du système général d’ornithologie, et suivi d’une table alphabétique des espèces (2nd edition). Chez H. Cousin.
- Zelenkov, N. V. (2016). Revision of non-passeriform birds from Polgárdi (Hungary, Late Miocene): 2. Galliformes. Paleontological Journal, 50(6), 623–634. doi:10.1134/S0031030116060162
- Zelenkov, N. (2021). New bird taxa (Aves: Galliformes, Gruiformes) from the early Eocene of Mongolia. Paleontological Journal, 55(4), 438–446. doi:10.1134/S0031030121040158
- Zelenkov, N. V., & Panteleyev, A. V. (2019). A small stem-galliform bird (Aves: Paraortygidae) from the Eocene of Uzbekistan. Comptes Rendus Palevol, 18(5), 517–523. doi:10.1016/j.crpv.2019.04.005