Abstract
This paper reviews regeneration research during the past 20 years. The focus is mainly on planting of Norway spruce and Scots pine. Research on root morphology and nursery practices has played an important role in the introduction of containerized seedlings in northern Europe. In recent years, mini-seedlings have been tested. Conventionally, seedlings are planted in spring and early autumn but recent research indicates that Norway spruce seedlings can be planted from spring until the end of September. Soil temperature strongly influences seedling establishment in the harsh northern Scandinavian climate and a good way to increase soil temperature is to plant in elevated planting spots, which can be achieved by mounding. Soil scarification also reduces competition from field vegetation and damage by pine weevils. In southern Finland, Sweden and Norway, pine weevils are by far the most serious causes of damage to both Norway spruce and Scots pine. Therefore, designing regeneration treatments for decreasing pine weevil damage without using insecticides is one of the more important current research challenges. Examples of possible future trends in forest regeneration research are an emphasis on more basic research, use of fast growing tree species, multidisciplinary approaches and site-specific regeneration regimes.
Introduction
In commercial forestry, regeneration sets the arena for the rest of the rotation. Hence, the most important silvicultural decisions are probably those related to regeneration treatments. Therefore, for a long time both forest companies and private forest owners in northern Europe have been attempting to increase the cost-effectiveness of regeneration treatments, and the degree to which they improve the survival and growth of seedlings. Research has played an important role in attempts to meet these goals.
In this paper, the regeneration stage is defined as the stage in a rotation between clear-felling and the time when the new generation of the main tree species has reached an average height of 1.3 m. The new generation may arise from naturally seeded material from the previous generation, or planted seeds or seedlings. For example, in planting of Norway spruce (Picea abies) and Scots pine (Pinus sylvestris) in Finland and Sweden, a frequent aim is to have at least 1800–2500 vigorous, planted seedlings per hectare at the end of the regeneration stage. However, this target is frequently not met. In Sweden, less than 80% of the regeneration areas met the requirement of the Forest Act during the years 1999–2009 if broadleaved species were included as crop-trees, and this proportion varied between 65 and 70% if only conifer species were included as crop-trees (). In southern Finland, a survey study showed that less than 60% of the regeneration areas had more than 1600 crop-trees per hectare (). Therefore, although substantial information about regeneration has been gathered, especially during the past 20 years, considerable further research efforts (and initiatives to ensure that acquired knowledge is rationally used) are still required to ensure that regeneration practices deliver consistently good results.
Figure 1. Proportion of regeneration area meeting the requirements in the Swedish law during the years 1999–2008. The total bars show the proportion of regeneration areas with sufficient seedling numbers including broadleaved species and the lower part shows the same proportion when only conifer species are included. Each bar represents the 3-year average. Data from a regeneration survey (Polytax) carried out by the Swedish Board of Forestry in Bergquist et al. (Citation2010). In the survey, 500 sites were inventoried during the years 1999–2007 and about 1800 sites year–1 during the years 2007–2008. In southern Sweden, the inventories were done 5 years after the landowner had submitted a notification of a planned clear-cut. In northern Sweden, the survey was done 7 years after submission of the clear-cut form. The survey is further described in Bergquist et al. (Citation2010).
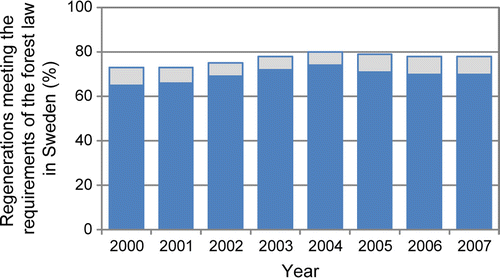
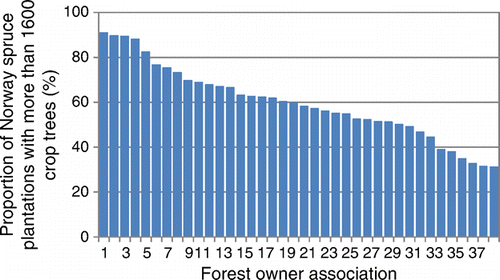
Figure 2. Proportion of Norway spruce plantations with more than 1600 crop trees ha–1. In total 9249 ha was inventoried during the years 2000–2006 in 40 forest owner associations in southern Finland. (Redrawn from Kankaanhuhta et al., Citation2009.)
The effectiveness of planting procedures has been increased by various means (outlined below), but in contrast to practices applied in other rotation stages they have remained largely unmechanized. Consequently, the costs of regeneration have not decreased to the same extent as costs of harvesting pulpwood and timber in thinnings and final felling. In the future, regeneration treatments will probably have to be more mechanized to reduce costs and to complete the tasks with fewer workers. Increased mechanization will probably place new demands on the regeneration treatments. Therefore, they may need to be more robust and applicable for longer planting seasons.
This paper reviews regeneration research and implementation of research results in practical forestry in northern Europe during the past 20 years (). Furthermore, the likely trends in research related to forest regeneration during the next 20 years are forecast. The focus is mainly on planting Norway spruce and Scots pine, but it is acknowledged that issues regarding direct seeding and natural regeneration, as well as the regeneration of broadleaves and other conifer species, have also been intensively addressed. In addition, although the review concentrates on research in Finland, Sweden and Norway, studies in other boreal regions are also mentioned.
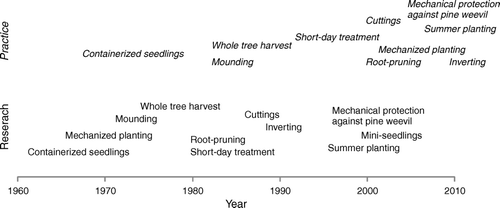
Figure 3. Period of the main research in various fields of regeneration research and when they were implemented in practical forestry in Scandinavia (italics).
Regeneration research in northern Europe
Nursery regimes and seedling types
The only seedlings that were available for planting programmes before the late 1960s were barerooted seedlings, but containerized seedlings were subsequently developed in attempts to make regeneration more economical and to avoid certain technical problems. The introduction of containerized seedlings offered considerable potential reductions in planting costs and extensions of the length of the planting season. However, their introduction was not problem free; notably, serious root deformation occurred among some containerized Scots pine and lodgepole pine (Pinus contorta) seedlings (Parviainen, Citation1976; Lindström, Citation1998; Rosvall, Citation1998) with profound implications for the growth and stability of plantations (Parvianen & Antola, Citation1986; Rune, Citation2003). Therefore, extensive efforts were made to improve the design of containers to avoid root deformation (e.g. Parviainen & Tervo, Citation1989; Almqvist, Citation1998). Tested design-based solutions have included the use of pots with slopes to direct roots downwards and containers that are either open to the air or treated with copper to kill root-tips when they reach the edge of the container (Rosvall, Citation1998).
Containerized seedlings have captured a large share of the market in the Nordic countries, but barerooted seedlings have some advantages; most importantly, they are larger and more tolerant of pine weevil (Hylobius abietis) damage (Örlander & Nilsson, Citation1999). Therefore, barerooted seedlings are still often used, especially in the southern parts of the Nordic countries on fertile, vegetation-rich sites with high risks of pine weevil damage. Since they establish more slowly and are older, barerooted seedlings also flush later than containerized seedlings in the spring, which may decrease the risk of frost damage (Langvall et al., Citation2000). However, compared to containerized seedlings, it takes longer for barerooted seedlings to establish good contact between roots and soil, and thus commence water uptake (Nilsson & Örlander, Citation1995). Therefore, first and second year height growth is often greater for containerized than for barerooted seedlings, and height differences between them have been found to become insignificant 3–5 years after planting (Nilsson & Örlander, Citation1999).
Another possible option to reduce seedling and planting costs that has been evaluated in Finland, Sweden and Canada is to use very small, 7–10 week-old, mini-seedlings (Kumpare, Citation1998; Lindström et al., Citation2005; Murphy & Wiebe, Citation2006). Scots pine seedlings were used initially in these tests, but Norway spruce seedlings have also been included in subsequent field trials (Lindström et al, 2005; Johansson et al., Citation2007). Johansson et al. (Citation2007) concluded that mini-seedlings grow as well as or even better than larger seedlings if they are successfully established, but they are more sensitive to their planting environment and handling. Danielsson et al. (Citation2008) found that mini-seedlings were damaged less by pine weevils than conventional 1-year-old seedlings, possibly because mini-seedlings emit limonene, which is known to repel the pine weevil. Furthermore, when wounded, green leaf volatiles were found to be released by mini-seedlings while the pine weevil attractant α-pinene was released by conventional seedlings.
Growth media for containerized seedlings should ideally be well aerated, retain water and nutrients, be easy to handle and cheap, have low pH and be mechanically cohesive, but naturally biodegradable after outplanting (Bernier & Gonzalez, Citation1995). Sphagnum peat material meets many of these requirements for growth medium for nursery production (Hellum, Citation1975; Heiskanen, Citation1993). However, Sphagnum peat is a variable material so factors that influence its suitability (and possible ways to improve its properties) for both nursery and postplanting have been investigated in several studies. Bernier and Gonzalez (Citation1995) found that including <1.3 mm particles in the peat influenced seedling morphology whereas humification did not, indicating that variation in water retention characteristics is more important than chemical properties of the peat in this respect. It should also be noted that physical properties that are desirable for growth media during nursery production may not always coincide with optimal properties for postplanting performance. For example, in the nursery it is important for growing media to drain quickly, to allow adequate aeration, but the use of coarse growth media may result in low water availability in the peat plug under field conditions (Bernier et al., Citation1995). It is also essential to handle the seedlings appropriately before planting; for instance, preplanting watering of container-grown Scots pine, Norway spruce and silver birch (Betula pendula) increases survival rates after planting, even if the most easily retained water is released to the soil within hours of planting (Heiskanen & Rikala, Citation2000).
A treatment that has been applied to seedlings in the autumn, for either autumn planting or winter storage, is exposure to short-day (SD) treatments to induce growth cessation (Rosvall-Åhnebrink, Citation1985, Citation1990). In Scots pine, SD treatments have also been used to induce two flushes and hence produce 2-year-old like seedlings in a year (Rosvall-Åhnebrink, Citation1982). The effects of varying the timing and duration of SD treatments in many tree species have been extensively studied in both the Nordic countries and North America. For Norway spruce seedlings, it has been shown that SD treatment has to last for 2–3 weeks and it cannot start before early July to cease height growth and harden seedlings effectively (Konttinen et al., Citation2003). If SD treatment is started earlier, the seedlings may have a second flush (Kohmann & Johnsen, Citation2007; Luoranen et al., Citation2009). Budburst may also occur earlier in the following spring among SD-treated seedlings than among untreated seedlings (Konttinen et al., Citation2003).
The thawing procedure applied after winter storage can also have substantial effects on subsequent physiological processes. For instance, Fløistad and Kohmann (Citation2001) found that Norway spruce seedlings that were thawed rapidly in water had greater frost tolerance, higher carbohydrate contents and later budbreak than slowly thawed seedlings. In North America, Camm et al. (Citation1995) and Kooistra and Bakker (Citation2002) showed that when Engelmann spruce (Picea engelmannii) seedlings are planted in warm soil (>18°C), thawing may be unnecessary. However, in boreal conditions, when Norway spruce seedlings are planted in cool soil in the spring or early autumn, thawing is necessary for successful planting (Helenius, Citation2005), especially during dry conditions. A 4-day period in cardboard boxes was recommended to ensure complete thawing of the root plug.
Various fertilization regimes have been developed, tested and applied over the years in attempts to produce high-quality seedlings for planting. Nutrient loading, in which fertilization is applied in autumn to increase the nutrient content of seedlings, has been successfully tested in several Nordic countries (Fløistad, Citation2002; Rytter et al., Citation2003; Fløistad & Kohmann, Citation2004; Luoranen et al., Citation2008; Heiskanen et al., Citation2009).
In the Nordic countries, the interest in clonal forestry has been focused on Norway spruce. An increased growth of about 30% compared to seedlings has been reported (Rosvall et al., Citation2001). The total gain of clones includes effects of the cutting propagation method as well as genetic selection effects. Gemmel et al. (Citation1991) found that growth of cuttings was superior to that of seedlings during the first 8 years, even if the genetic selection was negligible, and that this could be explained by differences in morphology and physiology. Cuttings are also reported to have lower mortality due to pine weevils than conventional seedlings with the same initial stem base diameter, possibly because they have thicker bark and needles at the base of their stems (Hannerz et al., Citation2002).
Planting date
Conventionally, seedlings are planted in spring and early autumn. These short planting windows are very busy times, in both the nurseries and forests. In addition, in the future, the number of forest workers will probably decrease, causing labour shortages and thus further problems for plantings. A solution may be to mechanize planting. For this to be economic the mechanized planting season should be as long as possible. Results of field studies in Finland indicate that Norway spruce seedlings can be planted from spring until the end of September without any negative effects on growth or survival (Luoranen et al., Citation2005, Citation2006b). Actively growing Norway spruce, silver birch and hybrid aspen (Populus tremula×P. tremuloides) containerized seedlings planted in summer may even grow better than spring- or autumn-planted seedlings owing to the warm soil and rapid root egress just after planting (Luoranen et al., Citation2003, Citation2006a, Citationb). For successful summer planting seedlings must be well watered at planting, and they must not dry out during transport and field storage before planting (Helenius et al., Citation2002, Citation2005). Furthermore, actively growing conifer seedlings used in plantings may be damaged by some insecticides that have been used against pine weevils (Luoranen & Viiri, Citation2005).
If the planting period is extended the nursery growing procedures must also be adjusted, since seedlings have to be prepared for each planting date. From spring until early summer, the planting season can be extended simply by planting freezer-stored seedlings. In Finland, freezer-stored seedlings can be used until mid-June. Computer simulations with long-term air temperature data (Hänninen et al., Citation2009) and experimental studies over a few years (Luoranen et al., Citation2005) have shown that if seedlings are planted before mid-June the remaining growing season is long enough for proper growth and frost hardening before the first autumn frosts. Before mid-June seedlings stored outdoors can also be planted, even if they are actively growing at planting (Luoranen et al., Citation2005). After mid-June, actively growing seedlings targeted for summer planting can be planted until late July, when SD-treated seedlings are ready for planting (Luoranen et al., Citation2006b).
In Canada, seedlings are also planted in summer, and the following procedure has been developed. The growth of selected seedlings is initiated in January or February, in a heated and lighted greenhouse, in which they are grown until SD treatment in late May or early June, then planted in July (Grossnickle & Folk, Citation2003). This method has not yet been tested or applied in northern Europe owing to the high energy costs. For plantings in late summer and early autumn, SD-treated seedlings are better since they are tolerant of drought and early autumn frosts after planting (Luoranen et al., Citation2006b, Citation2007). In Norway, Norway spruce seedlings exposed to early summer SD treatments for summer plantings have also been examined (Kohmann & Sønsteby, Citation2007).
Site preparation
Prescribed burning was a major soil treatment method from the 1930s until the early 1960s. A need for developing soil treatment equipment arose because prescribed burning became expensive when the cost of labour increased and the size of clear-cuts decreased. During the 1960s and 1970s, soil treatment was mainly done by hand tools or with small mechanical equipment, resulting in scarified patches of about 30×30 cm. Commonly used patch scarification, however, did not always produce satisfying regeneration results, especially when treating stony sites or sites with a high amount of slash. Therefore, disc-trenching was developed to achieve a large number of planting spots.
Various studies from the 1970s onwards found that soil temperature strongly influences seedling establishment in the harsh climate in northern Finland, Sweden and Norway (Örlander, Citation1984; Örlander et al., Citation1990). Accordingly, root growth is linearly correlated with soil temperature in the normal temperature interval during the vegetation period in this area (Vaapavuori et al., Citation1992), and since root growth is essential for water and nutrient uptake seedling establishment is limited by the growth of new roots. However, Johansson et al. (Citation2005) found poor correlation between the amount of new roots and gas exchange, which indicates that root growth and seedling establishment is not linearly correlated. A good way to increase soil temperature is to plant in elevated planting spots and this can be achieved by mounding (Örlander et al., Citation1990).
Another important aspect of soil scarification is that it reduces competition from field vegetation. Nilsson and Örlander (Citation1999) found that planted Norway spruce seedlings were only affected by competing vegetation during the first 2 years after planting, so a mounding treatment was as effective in reducing the negative effect of competing vegetation on their growth as an intensive herbicide treatment. However, these effects were observed during a period with high browsing pressure from roe deer and moose, resulting in sites where the field vegetation mainly consisted of one grass species (Deschampsia flexuosa, hairy grass) (Bergquist et al., Citation1999). Long-term studies have shown that in other circumstances more radical treatments than soil scarification may be needed to release planted seedlings from competition from unwanted vegetation (Boateng et al., Citation2009). However, since forest laws and certification rules restrict the use of herbicide treatments in northern Europe, forest managers have to find other ways to reduce competing vegetation. Soil scarification by disc-trenching or mounding, combined with precommercial thinning, is usually sufficient to release planted seedlings, but on fertile sites with abundant field vegetation or in afforestation on former farmland, more radical scarification methods may be needed. Nordborg and Nilsson (Citation2003) showed that deep soil cultivation enabled good seedling establishment on very fertile sites with abundant competing vegetation.
Soil scarification has been shown to be one way to protect seedlings from damage by pine weevils (H. abietis) (Petersson, Citation2004). Insecticide has to be combined with soil scarification to achieve good survival in areas with high populations of pine weevils (Petersson, Citation2004). It has been shown that pine weevils react very differently when walking on bare mineral soil compared to undisturbed humus (Kindvall et al., Citation2000; Björklund et al., Citation2003). It has also been found that if planted seedlings are surrounded by at least 10 cm bare mineral soil the effect of scarification on pine weevil damage is maximized (Lindström et al., Citation1986). However, with the scarification methods that are used today, it is difficult to obtain enough planting spots with bare mineral soil and a large number of seedlings are planted in humus and mineral soil mixtures, which have much weaker effects on pine weevil damage.
The inverse scarification treatment, in which the humus layer is buried by a 5–10 cm deep mineral soil layer in patches, has been shown to give good results in terms of both survival and early growth of planted seedlings (Örlander et al., Citation1998; Nordborg et al., Citation2003). Until recently inversion has been a costly technique, requiring the use of excavators, but good inverted planting spots can be made at reasonable cost using recently developed equipment mounted on ordinary excavators. This may enable new scarification methods to be applied that can be used in conjunction with mechanical planting.
Mechanized planting
In the 1970s, automated machines for continuous planting were developed in both Sweden (Silva Nova) and Finland (Serlachius), but they were too expensive to replace manual planting. In the early 1990s the Swedish company Bracke introduced a machine that both prepared soil and planted single seedlings. Two later machines, the Ecoplanter from Sweden and M-Planter from Finland, also performed both of these operations, but planted two seedlings simultaneously. These devices offer productivity and costs that are similar to or better than those of manual planting (Saarinen, Citation2006; Rantala et al., Citation2009). Of course, regeneration results after planting are important as well as costs. The Ecoplanter makes planting spots in which humus and mineral soil are mixed and, thus, risks for pine weevil damage are high (Saarinen, Citation2006). However, the Bracke and M-Planter machines make mounds with double humus layers covered by bare mineral soil, and regeneration results are comparable to those of manual planting (Nieuwenhuis & Egan, Citation2002; Luoranen & Saarinen, Citation2004). In Finland, approximately 30 planting devices are now commercially available, and almost 5% of all potential planting areas are mechanically planted, although in company-owned forest in some areas up to 20–30% of planting areas are planted by machines.
Pine weevil damage
In southern Finland, Sweden and Norway, pine weevils are by far the most serious cause of damage to both Norway spruce and Scots pine (Örlander & Nilsson, Citation1999; Petersson, Citation2004; Wallertz, Citation2009). Insecticides such as DDT and lindane were introduced in the late 1950s, but they were prohibited in the Nordic countries in the mid-1970s for environmental reasons. After that, the main treatment used to protect seedlings against damage by pine weevils was to leave sites fallow for several years (i.e. to plant seedlings on 3- or 4-year-old clear-cuts). However, planting on old clear-cuts has obvious disadvantages, e.g. reduced production because of competition from ground vegetation and leaving clear-cuts unplanted. Thus, the introduction of a synthetic pyrethroid (permethrin) in the late 1970s improved regeneration results in southern Finland, Sweden and Norway significantly. However, permethrin has been prohibited in all countries in the European Union (EU) since 2004. Currently, other insecticides can be used within the EU, but their use has been questioned because of the environmental and health hazards they pose.
When concerns about the use of permethrin were first raised, research into alternative methods to protect seedlings was initiated, and several mechanical protection systems have been tested (Lindström et al., Citation1986; Petersson et al., Citation2004). As mentioned above, scarification provides protection against pine weevil damage if seedlings can be planted in bare mineral soil. In addition, planting under shelterwoods has been shown to reduce pine weevil damage (Von Sydow & Örlander, Citation1994; Wallertz, Citation2009), as well as using seedlings with large root collar diameters (Örlander & Nilsson, Citation1999; Thorsén et al., Citation2001). However, none of these treatments provides sufficient protection alone, if the aim is to plant on fresh or 1-year-old clear-cuts in areas with high pine weevil populations. Instead, several treatments need to be combined and it has been shown that the treatment effects are generally additive (Petersson & Örlander, Citation2003). A decision support system has been constructed to help forest managers to find alternatives to insecticide treatments in southern Sweden (http://www2.ess.slu.se/snytbagge/default.htm).
Damage by frost
After planting, spring frosts may severely damage planted Norway spruce seedlings if they occur while new shoots are extending (Langvall et al., Citation2000). Planting under dense shelterwoods will reduce damage by frost (Langvall, Citation2000), and studies in the 1980s and 1990s identified several reasons for the protective effects of shelterwoods. First, the near-ground temperature during calm, clear nights is higher under shelterwoods than on clear-cuts because of higher turbulence in the air (Ottosson-Lövenius, Citation1993; Langvall, Citation2000). Secondly, the shade that shelter-trees provide during the day after a frost reduces damage to the photosynthetic apparatus (Lundmark & Hällgren, Citation1987; Lundmark et al., Citation1988). Thirdly, seedlings under shelterwoods flush later than seedlings on clear-cuts (Langvall, Citation2000). However, in order to be effective, the shelterwood has to be relatively dense (Langvall, Citation2000), and the density must be increased with reduced height of the shelter-trees. Frost damage reduces the growth of planted Norway spruce seedlings, but only extreme, successive frosts generally cause mortality among planted Norway spruce seedlings (Langvall et al., Citation2000; Bigras et al., Citation2004). In addition to reduced growth, frost damage can induce flaws in the wood that may reduce the quality (and value) of the end-product (Klang, Citation2000).
Late budburst reduces the risk of frost injuries by shifting the flushing period to a period with more favourable nocturnal air temperatures. Hence, selection of a late-flushing provenance is regarded as one of the best strategies for avoiding frost damage (Hannerz, Citation1999). A model, accompanied by a decision support system, for assessing the risk of frost damage for various provenances is available in Sweden (http://www.skogforsk.se/sv/KunskapDirekt/Alla-Verktyg/Frostrisk/).
In Scots pine plantations in northern Finland, Sweden and Norway, extreme weather conditions during the end of the winter are important causes of mortality (Eiche, Citation1966; Rosvall et al., Citation1998). Weather-related damage can be reduced by seed-source transfer from north to south (Eriksson et al., Citation1980; Rosvall et al., Citation1998). However, this seed-source transfer negatively influences growth and the trade-off between hardiness and growth potential is of major importance in cold climates, such as in most upland areas of northern Finland, Sweden and Norway (Rosvall et al., Citation1998).
Damage by browsing
Populations of browsing animals (moose, deer, roe deer) have increased considerably in the Nordic countries during the past 20–30 years (Bergquist, Citation1998). Browsing on Scots pine in many areas is so severe that forest managers avoid this tree species in plantations. Indeed, the area planted with Scots pine in southern Sweden is currently close to zero, whereas it was a major tree species 30 years ago. In many regions, the populations of browsing animals need to be reduced considerably before planting of Scots pine (or broadleaves) can be considered.
As long as browsing is done on small seedlings and only occurs during a few years, protective treatments can be applied to individual seedlings (Bergquist & Örlander, Citation1996). However, if significant levels of browsing may persist for a longer time, the animals need to be excluded from the regeneration area by fencing. Many of the protective treatments are relatively expensive and therefore little used (Bergquist & Örlander, Citation1996).
It has been shown that vigorous seedlings are more heavily browsed than less vital seedlings and that browsing on individual seedlings is correlated with the colour of the needles (Bergquist et al., Citation2003b). However, reducing the vitality of seedlings to avoid browsing will prolong the period when browsing occurs, and is also incompatible with the general aim of high production. Furthermore, Bergquist et al. (Citation2003a) found that roe deer browsing on spruce is less harmful than many foresters have feared, since the negative effects on height growth they observed were minor, and the multiple trunks it induced developed into spike-knots located near the ground.
Competing vegetation
Competing vegetation is commonly regarded as a severe regeneration problem (e.g. Davies, Citation1985; Boateng et al., Citation2009) and many studies have shown the importance of keeping planting spots free from competing vegetation (e.g. Bärring, Citation1967; Sands & Nambiar, Citation1984; Sutton, Citation1995). As described above, for most sites in northern Europe it is possible to achieve good vegetation control with the use of scarification, even if radical methods may have to be applied to vegetation-rich sites, and the use of herbicides is more or less forbidden by law or certification rules. Competing woody vegetation is currently removed using brush-saws. However, equipment that can be mounted on excavators or harvesters is being developed in both Finland (Strandström et al., Citation2009) and Sweden (Bergkvist et al., Citation2006).
Long-term effects of regeneration treatments
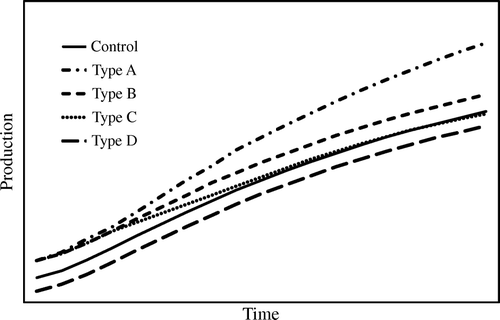
Four types of long-term growth response to early cultural treatments have been described (Hughes et al., Citation1979; Morris & Lowery, Citation1988). A type A response occurs when growth gains on treated areas continue to increase throughout the rotation (). A type B response occurs when growth gains achieved early in the rotation are maintained but do not continue to increase after the initial response period. A type C response occurs when early growth gains are subsequently lost, and a type D response occurs when early growth on treated areas falls below levels observed on non-treated areas (). The effect of various regeneration methods may be any of the above described types depending on site characteristics and how the regeneration methods are applied.
Figure 4. Hypothetical long-term growth response to early cultural treatments. See text for description of the type A–D responses.
Site preparation will result in a type B response if the only effect is a reduction in the abundance of competing herbaceous vegetation or a short-term increase in the rooting volume (Sutton, Citation1995; Nilsson & Örlander, Citation1999). In northern Europe, type B responses are probably the most common responses to conventional regeneration treatments. However, type A responses may occur after radical site preparation treatments that make previously unavailable nutrient pools available (Nordborg et al., Citation2006), if the site is drained (Hånell, Citation1988) or if site preparation also reduces competition from hardwoods by enabling planted crop trees to occupy the site before competing hardwoods establish (Nilsson & Allen, Citation2003; Boateng et al., Citation2009). Site preparation can also result in a type C or D response if the treatment involves removal of nutrients, e.g. harvest of slash for biofuel (Egnell & Valinger, Citation2003).
Since rates of various soil processes may increase after scarification, scientists have warned that nutrient losses might lead to reduced long-term site productivity (Lundmark, Citation1977, Citation1988). However, this conclusion is not supported by observed declines in productivity in long-term experiments (Örlander et al., Citation1990). Örlander et al. (Citation1996) studied the long-term development of planted Scots pine stands on poor sandy sediments after five different scarification treatments, and concluded that the risks of decreased long-term productivity on poor Scots pine sites caused by mechanical scarification methods used today are negligible since the top height on scarified plots was higher than on control plots.
In addition to their effects on growth, regeneration methods may affect stand development in terms of variability in size, which arises because of variations in environmental conditions and the seedlings’ physiological state, genotype and damage (Weiner & Thomas, Citation1986). By reducing the variability in seedling stands, variability can be reduced in mature stands (Lieffers & Titus, Citation1989), and since production is higher in a homogeneous stand than in a heterogeneous stand of the same dominant height, a type A response may occur as a result of improved homogeneity. In addition, there are indications that self-thinning may be delayed in stands with less variability before crown closure than in stands originating from seedling stands with large variation in sizes (Nilsson & Allen, Citation2003).
Forest regeneration research in 2030
Although the prediction of future research is difficult, strong current trends may give some indications of what may be needed and should be done in the future of forest regeneration research. General trends in research and development include strong progress in basic sciences, the need for multidisciplinary research, rapid technological developments, advances in information technology and management, global climate and energy policy, and the requirement for a holistic view. All these will also influence forest regeneration research.
Since its rise in the 1950s, forest regeneration research could be said to have been predominantly descriptive up to the 1980s, after which research has become both more quantitative and more basic. This trend of quantitative research will continue to be driven to a large extent by methodological and experimental development in other, more basic natural sciences and by new technology and equipment development in physical sciences (e.g. nanoscale applications). Forest regeneration as an applied science will benefit from these developments and allow researchers to measure variables that cannot be directly measured today, e.g. measures of in situ soil nutrient availability and nutrient mineralization, frost hardiness and root development.
The Nordic forest tree species will continue to be investigated using gene technology but it is unlikely that genetically modified (GM) seedlings will be commonly available in 2030, partly owing to the long required field testing times for risk assessment. Elsewhere, e.g. in Asia and South America, GM seedlings will be used in biomass production. Fast growing tree species may be used to a greater extent, and this will raise new regeneration questions, especially regarding the establishment of fast growing broadleaved tree species.
The EU's Climate and Energy Package will target a 20% renewable energy share (RES-D, Directive 2009/28/EC), which will require increased production of wood biomass to meet the total biomass demand. This will probably require an expansion of the areas to grow biomass and increasing efficiency in the production of wood biomass. Therefore, forest regeneration research will be needed but it will have to be better integrated with a strong economic and production process-orientated approach. New outcomes will be delivered by multidisciplinary approaches.
The forecast future climate change may raise important new regeneration research questions. For instance, the earlier start of the growing season may affect the risk of frost damage, damage by pine weevils may be more common in the northern parts of Finland, Sweden and Norway, competing vegetation may be increased and damage that currently is minor or non-existent may be important in the future if the climate is changing. The choice of tree species and genetic material, which greatly affect future production, should probably already today be adapted to possible future climate change.
In regeneration practices, mechanized planting with associated site preparation will be the dominant planting method in 20 years’ time. Mechanized planting both requires and makes it possible to develop new site-specific regeneration methods. In 20 years’ time and as a result of multidisciplinary research, appropriate regeneration methods will be better targeted to growing sites with abundant information on seedling progeny, from the nursery growing phase, and on site characteristics, achieved, for example, by different ground-based or airborne sensing systems and knowledge of past regeneration success at the site. Hence, the use of Geographical Information System (GIS) and other advanced information synthesizing systems (e.g. Sutinen et al., Citation2002) will be commonplace. Most unforeseen advances will probably be in the use of knowledge and data in various information technology and management applications for reforestation management, e.g. in risk assessments and other simulations.
Realization of this still requires that research has to test further and develop site-specific regeneration regimes, e.g. combinations of regeneration methods, choice of tree species and genetics, and protection against various damaging agents. Interactions occur between actions taken during different stages of the regeneration chain, e.g. site preparation may have a very different effect depending on nursery treatments. Research into possible interactions will be invaluable in designing site-specific regeneration methods.
Regeneration success depends on a series of events that start in seed procurement, continue at the nursery and end in precommercial thinning. Even if forest regeneration research will use more fundamental approaches in the future, regeneration research continues to combine and synthesize knowledge from various disciplines for applied forest management.
Related Research Data
References
- Almqvist C. 1998 Root development and stability Conference held in Garpenberg, Sweden, 30 September–1October 1997 Forestry Research Institute of Sweden Rep. 7 1 99
- Bärring , U. 1967 . Studier av metoder för plantering av gran och tall på åkermark I södra och mellersta Sverige [Studies of methods employed in the planting of Picea abies (L.) H. Karst and Pinus sylvestris (L.) on farmland in southern and central Sweden] . Studia Forestalia Suecica , 50 : 1 – 332 .
- Bergkvist , I. , Lundmark , T. , Rytter , L. and Thor , M. 2006 . Uttag av biobränslen i ungskog Arbetsrapport 611 , 1 – 17 . Uppsala : Skogforsk . (In Swedish.)
- Bergquist , J. 1998 . Influence by ungulates on early plant succession and forest regeneration in south Swedish spruce forests. Swedish University of Agricultural Sciences. Acta Universitatis Agriculturae Sueciae . Silvestria , 55 : 1 – 29 .
- Bergquist , J. , Bergström , R. and Zakharenka , A. 2003a . Responses of young Norway spruce (Picea abies) to winter browsing by roe deer (Capreolus capreolus): Effects on height growth and stem morphology . Scandinavian Journal of Forest Research , 18 : 368 – 376 .
- Bergquist J. Claesson S. Eriksson A. Fries C. 2010 Polytax 5/7 återväxttaxering: Resultat från 1999–2008 Rep. 6 1 44 Swedish Board of Forestry
- Bergquist , J. and Örlander , G. 1996 . Browsing deterrent and phytotoxic effects of roe-deer repellents on Pinus sylvestris and Picea abies seedlings . Scandinavian Journal of Forest Research , 11 : 145 – 152 .
- Bergquist , J. , Örlander , G. and Nilsson , U. 1999 . Deer browsing and slash removal affects field vegetation on south Swedish clearcuts . Forest Ecology and Management , 115 : 171 – 182 .
- Bergquist , J. , Örlander , G. and Nilsson , U. 2003b . Interactions among forestry regeneration treatments, plant vigour and browsing damage by deer . New Forests , 25 : 25 – 40 .
- Bernier , P. Y. and Gonzales , A. 1995 . Effects of the physical properties of Sphagnum peat on the nursery growth of containerized Picea mariana and Picea glauca seedlings . Scandinavian Journal of Forest Research , 10 : 176 – 183 .
- Bernier , P. Y. , Stewart , J. D. and Gonzales , A. 1995 . Effects of the physical properties of Sphagnum peat on water stress in containerized Picea mariana seedlings under simulated field conditions . Scandinavian Journal of Forest Research , 10 : 184 – 189 .
- Bigras , F. J. , Coursolle , C. and Margolis , H. A. 2004 . Survival and growth of Picea glauca seedlings as a function of freezing temperatures and exposure times during budbreak and shoot elongation . Scandinavian Journal of Forest Research , 19 : 206 – 216 .
- Björklund , N. , Nordlander , G. and Bylund , H. 2003 . Host-plant acceptance on mineral soil and humus by the pine weevil Hylobius abietis (L) . Agricultural and Forest Entymology , 5 : 61 – 65 .
- Boateng , J. O. , Heineman , J. L. , Bedford , L. , Harper , G. J. and Linnell Nemec , F. L. 2009 . Long-term effects of site preparation and postplanting vegetation control on Picea glauca survival, growth and predicted yield in boreal British Columbia . Scandinavian Journal of Forest Research , 24 : 111 – 129 .
- Camm , E. L. , Gay , R. D. , Kubien , D. S. , Goetze , D. C. , Silim , S. N. and Burton , P. J. 1995 . Physiological recovery of freezer-stored white and Engelmann spruce seedlings planted following different thawing regimes . New Forests , 10 : 55 – 77 .
- Danielsson , M. , Kännaste , A. , Lindström , A. , Hellqvist , C. , Stattin , E. , Långström , B. and Borg-Karlsson , A.-K. 2008 . Mini-seedlings of Picea abies are less attacked by Hylobius abietis than conventional ones: Is plant chemistry the explanation? . Scandinavian Journal of Forestry Research , 23 : 299 – 306 .
- Davies , R. J. 1985 . The importance of weed control and the use of tree shelters for establishing broadleaved trees on grass-dominated sites in England . Forestry , 58 : 167 – 180 .
- Egnell , G. and Valinger , E. 2003 . Survival, growth and growth allocation of planted Scots pine trees after different levels of biomass removal in clear-felling . Forest Ecology and Management , 177 : 65 – 74 .
- Eiche , V. 1966 . Cold damage and plant mortality in experimental plantations with Scots pine in northern Sweden . Studia Forestalia Suecica , 36 : 1 – 218 .
- Eriksson , G. , Andersson , S. , Eiche , V. , Ifver , J. and Persson , A. 1980 . Severity index and transfer effects on survival and volume production of Pinus sylvestris in northern Sweden . Studia Forestalis Suecica , 156 : 1 – 32 .
- Fløistad , I. S. 2002 . Effects of excessive nutrient supply and short day treatment on autumn frost hardiness and time of budbreak in Picea abies seedlings . Scandinavian Journal of Forestry Research , 17 : 295 – 303 .
- Fløistad , I. S. and Kohmann , K. 2001 . Effects of thawing procedure on frost hardiness, carbohydrate content and timing of budbreak in Picea abies . Scandinavian Journal of Forest Research , 16 : 30 – 36 .
- Fløistad , I. S. and Kohmann , K. 2004 . Influence of nutrient supply on spring frost hardiness and time of bud break in Norway spruce (Picea abies (L.) Karst.) seedlings . New Forests , 27 : 1 – 11 .
- Gemmel , P. , Örlander , G. and Högberg , K. A. 1991 . Norway spruce cuttings perform better than seedlings of the same genetic origin . Silvae Genetica , 40 : 198 – 202 .
- Grossnickle , S. C. and Folk , R. S. 2003 . Spring versus summer spruce stock of western Canada: Nursery development and field performance . Western Journal of Applied Forestry , 18 : 267 – 275 .
- Hånell , B. 1988 . Postdrainage forest productivity of peatlands in Sweden . Canadian Journal of Forest Research , 18 : 1443 – 1456 .
- Hannerz , M. 1999 . Early testing of growth rhythm in Picea abies for prediction of frost damage and growth in the field. Swedish University of Agricultural Sciences. Acta Universitatis Agriculturae Sueciae . Silvestria , 85 : 1 – 45 .
- Hannerz , M. , Thorsén , Å. , Mattsson , S. and Weslien , J. 2002 . Pine weevil (Hylobius abietis) damage to cuttings and seedlings of Norway spruce . Forest Ecology and Management , 160 : 11 – 17 .
- Hänninen , H. , Luoranen , J. , Rikala , R. and Smolander , H. 2009 . Frozen-stored seedling of Norway Spruce: Effect of planting date on the risk of autumn frost damage . Silva Fennica , 43 : 817 – 830 .
- Heiskanen , J. 1993 . Favourable water and aeration conditions for growth media used in containerized tree seedling production: A review . Scandinavian Journal of Forest Research , 8 : 337 – 358 .
- Heiskanen , J. , Lahti , M. , Luoranen , J. and Rikala , R. 2009 . Nutrient loading has a transitory effect on the nitrogen status and growth of outplanted Norway spruce seedlings . Silva Fennica , 43 : 249 – 260 .
- Heiskanen , J. and Rikala , R. 2000 . Effect of peat-based container media on establishment of Scots pine, Norway spruce and silver birch seedlings after transplanting in contrasting water conditions . Scandinavian Journal of Forest Research , 15 : 49 – 57 .
- Helenius , P. 2005 . Effect of thawing regime on growth and mortality of frozen-stored Norway spruce container seedlings planted in cold and warm soil . New Forests , 29 : 33 – 41 .
- Helenius , P. , Luoranen , J. and Rikala , R. 2005 . Physiological and morphological responses of dormant and growing Norway spruce container seedlings to drought after planting . Annals of Forest Science , 62 : 201 – 207 .
- Helenius , P. , Luoranen , J. , Rikala , R. and Leinonen , K. 2002 . Effect of drought on growth and mortality of actively growing Norway spruce container seedlings planted in summer . Scandinavian Journal of Forest Research , 17 : 218 – 224 .
- Hellum , A. K. 1975 . Selecting peat for rearing coniferous container seedlings . Forest Chronicle , 51 : 200 – 202 .
- Hughes , J. H. , Campbell , R. G. , Duzan , H. W. Sr and Dudley , C. S. 1979 . Site index adjustment for intensive forest management treatments in North Carolina. Forest Research Technical Report , New Bern, NC : Weyerhauser .
- Johansson , K. , Nilsson , U. and Allen , H. L. 2007 . Interactions between soil scarification and Norway spruce seedling types . New Forests , 33 : 13 – 27 .
- Johansson , K. , Söderbergh , I. , Nilsson , U. and Allen , H. L. 2005 . Effects of scarification and mulch on establishment and growth of six different clones of Picea abies . Scandinavian Journal of Forest Research , 20 : 421 – 430 .
- Kankkaanhuhta , V. , Saksa , T. and Miina , J. 2009 . “ Quality management of forest regeneration services processes ” . In Forest regeneration: Ecology, management and economics , Edited by: Fournier , M. V. New York : Nova Science Publishers .
- Kindvall , O. , Nordlander , G. and Nordenhem , H. 2000 . Movement behaviour of pine weevil Hylobius abietis in relation to soil type: An arena experiment . Entomologia Experimentalis et Applicata , 95 : 53 – 61 .
- Klang , F. 2000 . The influence of silvicultural practices on tree properties in Norway spruce. Swedish University of Agricultural Sciences. Acta Universitatis Agriculturae Sueciae . Silvestria , 128 : 1 – 33 .
- Kohmann , K. and Johnsen , Ø. 2007 . Effects of early long-night treatment on diameter and height growth, second flush and frost tolerance in two-year-old Picea abies container seedlings . Scandinavian Journal of Forest Research , 22 : 375 – 383 .
- Kohmann K. Sønsteby F. 2007 Summer planting with early long night treated seedlings Forskning fra Skog og landskap 2/07. (In Norwegian with English summary
- Konttinen , K. , Rikala , R. and Luoranen , J. 2003 . Timing and duration of short-day treatment of Picea abies seedlings . Baltic Forestry , 9 ( 2 ) : 2 – 9 .
- Kooistra , C. M. and Bakker , J. D. 2002 . Planting frozen conifer seedlings: Warming trends and effects on seedling performance . New Forests , 23 : 225 – 237 .
- Kumpare T. 1998 Minitaimien hyvä kehitys jatkuu [The good development of mini-seedlings continues] Mätsähallitus, kehittämisyksikön tiedote 4 2 (In Finnish.)
- Langvall , O. 2000 . Interactions between near-ground temperature and radiation, silvicultural treatments and frost damage to Norway spruce seedlings. Swedish University of Agricultural Sciences. Acta Universitatis Agriculturae Sueciae . Silvestria , 330 : 1 – 34 .
- Langvall , O. , Nilsson , U. and Örlander , G. 2000 . Frost damage to planted Norway spruce seedlings—Influence of site preparation and seedling type . Forest Ecology and Management , 141 : 225 – 237 .
- Lieffers , V. J. and Titus , S. J. 1989 . The effects of stem density and nutrient status on size inequality and resource allocation in lodgepole pine and white spruce seedlings . Canadian Journal of Botany , 67 : 2900 – 2903 .
- Lindström , A. 1998 . Rotdeformationer och deras konsekvenser för täckrotsplantornas etablering och framtida kvalitetsutveckling. Rotutveckling och Stabilitet. Konferens i Garpenberg, 30 sep–1 okt 1997 . SkogForsk Redogörelse , 7 : 53 – 62 .
- Lindström , A. , Hellqvist , C. , Gyldberg , B. , Långström , Å. and Mattsson , A. 1986 . Field performance of a protective collar against damage by Hylobius abietis . Scandinavian Journal of Forest Research , 1 : 3 – 15 .
- Lindström A. Hellqvist C. Stattin E. 2005 Mini seedlings—A new forest regeneration system J. S. Colombo The Thin Green Line—A symposium on the state-of-the-art in reforestation—Proceedings Forest Research Information Paper No. 160 59 61 Sault Ste Marie, ON, Canada
- Lundmark J.-E. 1977 Marken som en del I det skogliga ekosystemet Sveriges Skogsvårdsförbunds Tidskrift 2 109 122 (In Swedish.)
- Lundmark J.-E. 1988 Skogsmarkens ekologi- ståndortsanpassat skogsbruk. Del 2—Tillämpning . Jönköping Skogsstyrelsen (In Swedish.)
- Lundmark , T. and Hällgren , J.-E. 1987 . Effects of frost on shaded and exposed spruce and pine seedlings planted in the field . Canadian Journal of Forest Research , 17 : 1197 – 1201 .
- Lundmark , T. , Hällgren , J.-E. and Degermark , C. 1988 . Effects of summer frosts on the gas exchange of field-grown Pinus sylvestris L. Seedlings . Scandinavian Journal of Forest Research , 3 : 441 – 448 .
- Luoranen , J. , Helenius , P. , Huttunen , L. and Rikala , R. 2007 . Short-day treatment enhances root egress of summer planted Picea abies seedlings under dry conditions . Scandinavian Journal of Forest Research , 22 : 384 – 389 .
- Luoranen , J. , Konttinen , K. and Rikala , R. 2009 . Frost hardening and risk of a second flush in Norway spruce seedlings after an early-season short-day treatment . Silva Fennica , 43 : 235 – 247 .
- Luoranen , J. , Lahti , M. and Rikala , R. 2008 . Frost hardiness of nutrient-loaded two-year-old Picea abies seedlings in autumn and at the end of freezer storage . New Forests , 35 : 207 – 220 .
- Luoranen , J. , Lappi , J. , Zhang , G. and Smolander , H. 2006a . Field performance of hybrid aspen clones planted in summer . Silva Fennica , 40 : 257 – 269 .
- Luoranen , J. , Rikala , R. , Konttinen , K. and Smolander , H. 2005 . Extending the planting period of dormant and growing Norway spruce container seedlings to early summer . Silva Fennica , 39 : 481 – 496 .
- Luoranen , J. , Rikala , R. , Konttinen , K. and Smolander , H. 2006b . Summer planting of Picea abies container-grown seedlings: Effects of planting date on survival, height growth and root egress . Forest Ecology and Management , 237 : 534 – 544 .
- Luoranen , J. , Rikala , R. and Smolander , H. 2003 . Root egress and field performance of actively growing Betula pendula container seedlings . Scandinavian Journal of Forest Research , 18 : 133 – 144 .
- Luoranen J. Saarinen V.-M. 2004 Koneellinen istutus ja sen onnistuminen [Success of mechanized planting] TaimiUutiset 2/2004 17 21 (In Finnish.)
- Luoranen , J. and Viiri , H. 2005 . Insecticides sprayed on seedlings of Picea abies during active growth: Damage to plants and effect on pine weevils in bioassay . Scandinavian Journal of Forest Research , 20 : 47 – 53 .
- Morris , L. A. and Lowery , R. F. 1988 . Influences of site preparation on soil conditions affecting stand establishment and tree growth . Southern Journal of Applied Forestry , 12 : 170 – 178 .
- Murphy W. F. Wiebe R. P. F. S. 2006 Shallow site jack pine outplanting success using Jiffy mini-plug 1842 mm pellets with two planting tools (Res. Rep. X-11-06) Thunder Bay, ON Forest Renewal Cooperative, Faculty of Forestry and the Forest Environment
- Nieuwenhuis , M. and Egan , D. 2002 . An evaluation and comparison of mechanized and manual planting on afforestation and reforestation sites in Ireland . International Journal of Forest Engineering , 13 ( 2 ) : 11 – 23 .
- Nilsson , U. and Allen , H. L. 2003 . Short- and long-term effects of site preparation, fertilization and vegetation control on growth and stand development of planted loblolly pine . Forest Ecology and Management , 175 : 367 – 377 .
- Nilsson , U. and Örlander , G. 1995 . Effects of some regeneration methods on drought damage of newly planted Norway spruce seedlings . Canadian Journal of Forest Research , 25 : 790 – 802 .
- Nilsson , U. and Örlander , G. 1999 . Vegetation management on grass-dominated clearcuts planted with Norway spruce in southern Sweden . Canadian Journal of Forest Research , 29 : 1015 – 1026 .
- Nordborg , F. and Nilsson , U. 2003 . Growth and net nitrogen uptake in Picea abies (L.) Karst. seedlings, effects of fertilization and site preparation . Annals of Forest Science , 60 : 657 – 666 .
- Nordborg , F. , Nilsson , U. , Gemmel , P. and Örlander , G. 2006 . Carbon and nitrogen stocks in soil, trees and field vegetation in conifer plantations 10 years after deep soil cultivation and patch scarification . Scandinavian Journal of Forest Research , 21 : 356 – 363 .
- Nordborg , F. , Nilsson , U. and Örlander , G. 2003 . Effects of different soil treatments on growth and net nitrogen uptake of newly planted Picea abies (L.) Karst. seedlings . Forest Ecology and Management , 180 : 571 – 582 .
- Örlander G. 1984 Some aspects of water relations in planted seedlings of Pinus sylvestris PhD thesis, Department of Silviculture, Swedish University of Agricultural Sciences, Umeå
- Örlander , G. , Egnell , G. and Albrektson , A. 1996 . Long-term effects of site preparation on growth of Scots pine . Forest Ecology and Management , 86 : 27 – 37 .
- Örlander , G. , Gemmel , P. and Hunt , J. 1990 . Site preparation. A Swedish overview . FRDA Report , 105 : 1 – 62 .
- Örlander , G. , Hallsby , G. , Gemmel , P. and Wilhelmsson , C. 1998 . Inverting improves establishment of Pinus contorta and Picea abies: 10 year results from a site preparation trial in northern Sweden . Scandinavian Journal of Forest Research , 13 : 160 – 168 .
- Örlander , G. and Nilsson , U. 1999 . Effects of reforestation methods on pine weevil (Hylobius abietis) damage and seedling survival . Scandinavian Journal of Forest Research , 14 : 341 – 354 .
- Ottosson-Löfvenius M. 1993 Temperature and radiation regimes in pine shelterwoods and clearcut areas 1 109 Doctoral thesis, Department of Forest Ecology, Swedish University of Agricultural Sciences.
- Parviainen J. 1976 Männyn eri taimilajien juuriston alkukehitys [Initial development of root systems of various types of nursery stock for Scots pine] Folia Forestalia 268
- Parviainen J. Antola J. 1986 Taimien kehitys ja juuriston morfologia eri taimilajeilla perustetuissa mäntyistutuksissa [Summary: The root system morphology and stand development of different types of pine nursery stock] Folia Forestalia 671
- Parviainen , J. and Tervo , L. 1989 . “ A new approach for production of containerised coniferous seedlings using peat sheets coupled with root pruning ” . In Conifer planting stock. Forestry Supplement Edited by: Mason , W. L. , Deans , J. L. and Thompson , S. 87 – 94 .
- Petersson , M. 2004 . Regeneration methods to reduce pine weevil damage to conifer seedlings. Swedish University of Agricultural Sciences. Acta Universitatis Agriculturae Sueciae . Silvestria , 330 : 1 – 34 .
- Petersson , M. and Örlander , G. 2003 . Effectiveness of combinations of shelterwood, scarification, and feeding barriers to reduce pine weevil damage . Canadian Journal of Forest Research , 33 : 64 – 73 .
- Petersson , M. , Örlander , G. and Nilsson , U. 2004 . Feeding barriers to reduce damage by pine weevil (Hylobius abietis) . Scandinavian Journal of Forest Research , 19 : 48 – 59 .
- Rantala , J. , Harstela , P. , Saarinen , V.-M. and Tervo , L. 2009 . A techno-economic evaluation of Bracke and M-Planter tree planting devices . Silva Fennica , 43 : 659 – 667 .
- Rosvall O. 1998 Rotsystemets stabilitetsmekanik ställer krav på plantodlingsbehållares utformning. Rotutveckling och Stabilitet. Konferens i Garpenberg, 30 sep–1 okt 1997 Skogforsk Redogörelse 7 32 36 (In Swedish.)
- Rosvall O. Andersson B. Ericsson T. 1998 Species-specific guidelines for choosing forest regeneration material for northern Sweden Skogforsk Redogörelse 1 1 66 (In Swedish with English summary)
- Rosvall O. Jansson G. Andersson B. Ericsson T. Karlsson B. Sonesson J. Stener L.-G. 2001 Genetiska vinster i nuvarande och framtida fröplantager och klonblandningar Redogörelse nr 01, 2001. (In Swedish with English summary.)
- Rosvall-Åhnebrink G. 1982 Practical application of dormancy induction techniques to greenhouse-grown conifers in Sweden Proceedings of the Canadian containerized tree seedling symposium Toronto Ontario September 1981 14–16 163 170
- Rosvall-Åhnebrink G 1985 Invintring av plantor för höstplantering eller vinterlagring. I: Skogsskötsel i södra Sverige. Swedish University of Agricultural Sciences, Faculty of Forestry Skogsfakta, Konferens 7 33 37 Uppsala. (In Swedish.)
- Rosvall-Åhnebrink G 1990 Bättre plantkvalitet genom styrning av fotoperiod och temperatur. I: Fytotronen 25 års forskning i plantodlingens och skogsträdsförädlingens tjänst. Swedish University of Agricultural Sciences, Faculty of Forestry , Skogsfakta, Konferens 14 27 34 Uppsala. (In Swedish.)
- Rune , G. 2003 . Instability in plantations of container grown Scots pine and consequences on stem form and wood properties. Swedish University of Agricultural Sciences. Acta Universitatis Agriculturae Sueciae . Silvestria , 281 : 1 – 35 .
- Rytter , L. , Ericsson , T. and Rytter , R.-M. 2003 . Effects of demand-driven fertilization on nutrient use, root:plant ratio and field performance of Betula pendula and Picea abies . Scandinavian Journal of Forest Research , 18 : 401 – 415 .
- Saarinen , V.-M. 2006 . The effects of slash and stump removal on productivity and quality of forest regeneration operations—Preliminary results . Biomass and Bioenergy , 30 : 349 – 356 .
- Sands , R. and Nambiar , E. K. S. 1984 . Water relations of Pinus radiata in competition with weeds . Canadian Journal of Forest Research , 14 : 233 – 237 .
- Strandström M. Hämäläinen J. Pajuoja H. 2009 Mechanizing of silvicultural work Metsätehon raportti 206 (In Finnish)
- Sutinen , R. , Teirilä , A. , Pänttäjä , M. and Sutinen , M.-L. 2002 . Distribution and diversity of tree species with respect to soil electrical characteristics in Finnish Lapland . Canadian Journal of Forest Research , 32 : 1158 – 1170 .
- Sutton , R. F. 1995 . White spruce establishment: initial fertilisation, weed control and irrigation evaluated after three decades . New Forests , 9 : 123 – 133 .
- Thorsén , Å. , Mattsson , S. and Weslien , J. 2001 . Influence of stem diameter on the survival and growth of containerized Norway spruce seedlings attacked by pine weevils (Hylobius abietis) . Scandinavian Journal of Forest Research , 16 : 54 – 66 .
- Vapaavuori , E. M. , Rikala , R. and Ryyppö , A. 1992 . Effect of root temperature on growth and photosynthesis in conifer seedlings during shoot elongation . Tree Physiology , 10 : 217 – 230 .
- Von Sydow , F. and Örlander , G. 1994 . The influence of shelterwood density on Hylobius abietis (L.) occurrence and feeding on planted conifers . Scandinavian Journal of Forest Research , 9 : 367 – 375 .
- Wallertz , K. 2009 . Pine weevil feeding in Scots pine and Norway spruce regenerations. Doctoral thesis, Swedish University of Agricultural Sciences . Acta Universitatis Agriculturae Sueciae , 60 : 1 – 46 .
- Weiner , J. and Thomas , S. C. 1986 . Size variability and competition in plant monocultures . Oikos , 47 : 211 – 222 .