
ABSTRACT
How seedling mortality and browsing affects species composition of regenerating forests has been mostly studied on a small scale. Yet, large-scale analyses based on extensive data are essential for robust prediction of species composition in young forests. In this study, we used a dataset from a national inventory of young forests (1–4 metres in height) to investigate the species composition of young forests across Sweden. We found that most of the regenerated forest area (almost 90%) was planted with Norway spruce (southern Sweden) and Scots pine (northern Sweden). Regeneration of Norway spruce was generally relatively successful but as a consequence of seedling mortality and browsing, almost 40% of the area regenerated with Scots pine will probably not develop into pine-dominated stands. Thus, low survival of Scots pine seedlings and trees can profoundly change the trajectory of species composition of the young forest from what was originally intended, and a large proportion of the young stands may develop into mixtures of conifers and broadleaves. While such mixtures may benefit certain biodiversity and ecosystem services, a loss of Scots pine dominated stands may also have adverse impacts on the economic returns as well as pine-dependent biodiversity and recreational values.
Introduction
Norway spruce (Picea abies L.) and Scots pine (Pinus sylvestris L.) are the two dominant tree species in Sweden, and crucial for economically profitable production of pulpwood and timber (Nilsson et al. Citation2012; Nilsson et al. Citation2011). According to the Swedish National Forest Survey (Citation2010), Norway spruce (spruce) contributes 40.5% of the total volume of Swedish forests, followed by Scots pine (pine) with 38.9%. Because of tradition, availability of seedlings, and for economic reasons, more than 70% of the forest land in Sweden was regenerated with these two conifer species after the rotation period in the 2000s (Bergquist et al. Citation2011). The density of planted seedlings varies from 2000 to 3000 trees ha−1, depending on site conditions and geographical location (Bergquist et al. Citation2011). In Sweden, the aim is to have at least 1800–2500 vigorous spruce or pine seedlings per hectare at the end of the regeneration stage (Nilsson et al. Citation2010). Successful regeneration is one of the most important components of sustainable forest ecosystems and a crucial management goal, although reaching the target regeneration outcome can be difficult, unpredictable, and in some cases also expensive (Kabrick et al. Citation2006; Loftis and McGee Citation1993).
In Sweden, the regeneration success of spruce and pine is threatened by several abiotic and biotic factors that can cause damage and mortality. In many regeneration areas and young forests, browsing by ungulates is a major concern (Bergquist et al. Citation2003; Cederlund Citation1983; Mysterud Citation2000) as it may cause mortality up to 30–35% of seedlings (Edenius et al., Citation1994; Wallgren et al., Citation2002). Ungulates cause damage to trees by browsing apical shoots, lateral twigs, breaking the main stem, or stripping the stem bark (Bergquist et al. Citation2001). This damage causes growth and quality losses. Mortality due to browsing may profoundly change the species composition of forests (Gill Citation1992). Moreover, ungulates are very selective in their browsing and prefer some species over others (Månsson et al. Citation2007). The favoured tree species in ungulate diets are rowan (Sorbus aucuparia L.), willow (Salix caprea L.), oak (Quercus spp L.), and aspen (Populus tremula L.) in Sweden, but their diet is dominated by Scots pine because of its availability, abundance and preference (Berström & Hjeljord, Citation1987; Cederlund &Berström, Citation1996; Månsson et al. Citation2007). This browsing pressure on Scots pine could change the species composition in forests.
So far, most studies on the effect of seedling mortality due to different agents have focused on a local scale. While it has been shown that the effect of browsing on forest composition and structure depends on the spatial and temporal scale (Wallgren et al. Citation2002), little is known about how seedling mortality during the regeneration stage influences the tree species composition of the developing forests at a larger scale. This requires a large number of observations over large areas to obtain a robust analysis of species’ vulnerability to browsing damage and the eventual consequences for forest species composition. In this study, we used a unique national-level dataset of damage in young forests (1–4 metres tall) to assess the effect of browsing on species composition in Swedish forests. In particular, we were interested in finding out how well the species composition of young forests corresponds to planted used in the regeneration stage. Our hypothesis was that browsing damage can alter the trajectory of species composition of developing forests, profoundly changing it from what was intended by the forest owners.
Materials and methods
Data source and analysis
We used data from the Äbin (Moose browsing data) national inventory for Sweden collected during 2015–2018. Äbin is an inventory method developed by the Swedish Forest Agency and the Swedish University of Agricultural Sciences (SLU) to estimate amounts of browsing damage on young trees in Sweden. The inventory is carried out in young forests with mean stand heights between 1 and 4 metres. A detailed description of the Äbin measurements and data is given in Appendix 1. In total, 147,736 plots in 32,894 stands evenly distributed and located in all forested regions of Sweden were measured and used in this study (). In our analysis, we divided the country into three domains (northern, central, and southern) to investigate the species composition in different regions of Sweden ().
Figure 1. An example plot (left) and a schematic illustration (right) of the half-height calculation based on the two tallest trees (right) in an Äbin inventory.
Figure 2. Äbin inventory stands located in northern, southern and central Sweden. Each dot represents the location of each stand and boundaries show the division of different regions.
The Äbin dataset includes an estimate of tree species used in regeneration by field crew during inventory. We used this information to investigate which species were used in the regeneration phase in different regions of Sweden.
The proportion of density classes of Scots pine in Scots pine regenerations and Norway spruce in Norway spruce regeneration were estimated by averaging trees per ha for all plots within each stands with a specific regeneration species. Thereafter, the total area that these plots represented were summed up in the various density classes and divided by the total area of respective regeneration species. This was done for each of the four years of inventory and standard error of the mean for the mean value of these four years is indicated in the figures. In stands where more than one regeneration species had been used, the stand was divided into smaller stands with an area proportional to the number of plots with the different regeneration species.
Stands with more than 500 trees ha−1 (TPH) were classified into five different forest types based on the species composition: (1) Scots pine monoculture, (2) Norway spruce monoculture, (3) conifer mixture, (4) conifer-dominated mixture and (5) broadleaf-dominated mixture. In the first three classes, pine, spruce or both in combination had more than 1500 (TPH), respectively. In the fourth class, pine and spruce together were between 1000 and 1500 TPH and additional broadleaves resulted in a total stem density exceeding 1500 TPH. The fifth class had more than 1500 TPH but conifers were below 1000 TPH.
In the analysis, we assumed that 50% of the damaged trees will develop into a future crop tree. In order to investigate the importance of this assumption, we performed a sensitivity analysis where all, every second, or none of the damaged trees were included as future crop trees.
The spatial distribution of species composition (occurrence of species) was analysed using the Inverse Distance Weighted (IDW) interpolation method. This method uses the measured values to predict the unmeasured value. IDW assumes that each measured point has a local influence that diminishes with distance. It gives greater weights to points close to the prediction location, and the weights diminish as a function of distance. In this study, we used species composition in 1 × 1km grids to obtain a continuous map for whole Sweden by using Arcmap 10.6 (ESRI, Citation2010)
Results
Tree species used in the regeneration stage
Norway spruce and Scots pine were the dominant tree species for planted regeneration in different regions of Sweden. More than 90% of the forest area inventoried in this study was regenerated with one of these conifer species in all the three regions of Sweden ( and ). A small proportion of the area was regenerated with other species (2–5%) and around 3–6% did not receive any regeneration treatments. Moreover, the regeneration species could not be determined in plots representing about 2–3% of the total area ().
Figure 3. Species used in regeneration and area coverage by each species in different regions of Sweden.
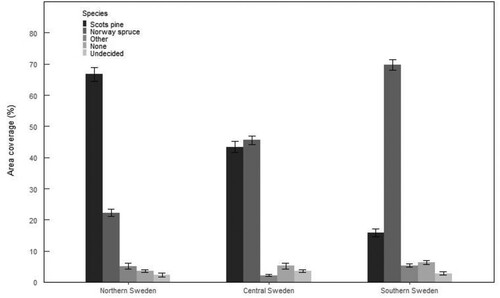
Figure 4. Spatial distribution of species used in the regeneration of Swedish forests. The map shows the proportion of total conifer and both Norway spruce and Scots pine used in different regions of Sweden in regeneration. The different colour of the scale shows different proportion of species, for example dark green represents 81-100% of species used in regeneration.
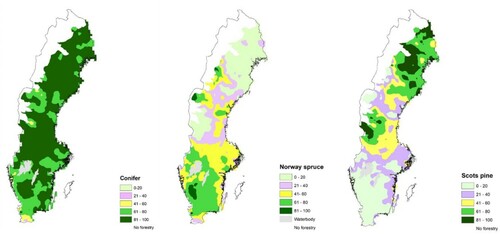
Norway spruce was the dominant regeneration species in southern Sweden and Scots pine in northern Sweden. Almost 70% of the area was regenerated with Norway spruce in southern Sweden and Scots pine in northern Sweden ( and ). The proportion of spruce decreased from central to northern Sweden and was very low in the interior northern Sweden (). The proportion of Scots pine decreased towards the south. In southern Sweden, pine was more common on the east coast but there was no area in southern Sweden with a dominance of pine regeneration ()
Tree species composition in young forests
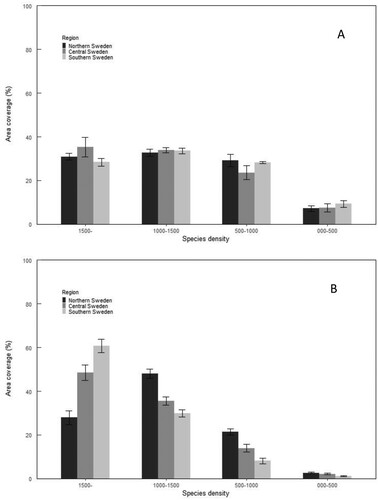
A little more than 60% of the stands between 1 and 4 m mean height regenerated with Norway spruce had more than 1500 Norway spruce TPH and around 90% had more than 1000 Norway spruce TPH in southern Sweden. The density of Norway spruce in Norway spruce regenerations was lower in the two northern regions but around 80% of the area regenerated with Norway spruce had more than 1000 Norway spruce TPH in central and northern Sweden. In Scots pine regenerations, about 30–35% of the total area had more than 1500 Scots pine trees and about 62–68% of the area had more than 1000 Scots pine trees in pine regenerations. ().
Figure 5. Area coverage of different stem number classes by regeneration species: (a) Scots pine in stands regenerated with Scots pine and (b) Norway spruce in Norway spruce regenerated stands.
Spatial distribution of species
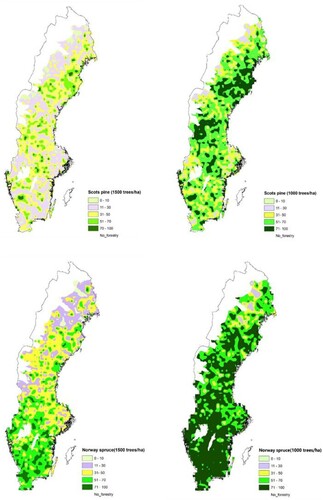
Neither Norway spruce or Scots pine had high dominance in any of the regions if 1500 TPH is considered as a threshold for dominance. However, a large part of southern Sweden is dominated by Norway spruce if 1000 TPH is set as the criterion of dominance. With the same standard for Scots pine, areas of high dominance could be found in the east coast in northern Sweden and in some part the interior of central Sweden and in small pockets in southern Sweden ().
Figure 6. Spatial distribution of Scots pine (upper figures) and Norway spruce (lower figures) dominated stands when 1000 and 1500 TPH of the respective tree species was considered as the definition of dominance.
Comparison of species composition forecasts with different weighting for damaged trees
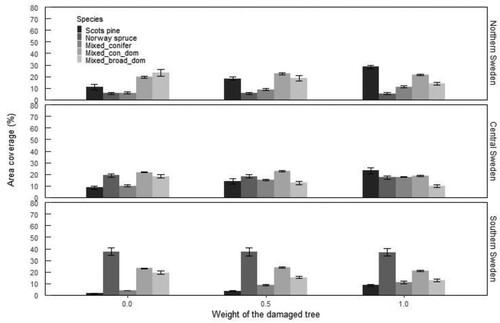
Damage weighting had almost no effect on the proportion of Norway spruce across all regions, but it did have a significant impact on the proportion of Scots pine. There are almost no Scots pine-dominated stands in southern and central Sweden if only undamaged trees are considered as future crop trees. In northern Sweden, the proportion of Scots pine-dominated stands increased from 11% if no damaged trees were included to almost 30% when all damaged trees were included (). In the northern and central regions, mixed forests dominated regardless of whether the damaged trees were included or not. However, the change in the proportion of Scots pine when damaged trees were included or not also changed the proportion of mixtures. If damaged Scots pine were excluded it was replaced by broadleaves which increased the proportion of broadleaf-dominated mixtures ().
Figure 7. Species composition of young forest in Sweden as a function of damaged tree weighting. The bars show the proportion of area with more than 1500 TPH in each species composition class. In the species composition classes, the respective species alone or together made up more than 1500 TPH (see Materials and Methods for further description of species composition classes).
Discussion
Species composition in young forests differs from the planting intention
We found only about one third of the area that had been regenerated with pine that contained more than 1500 TPH. The proportion of Norway spruce regeneration that had more than 1500 TPH of Norway spruce was higher (48% in the central and 60% in the southern regions). A high discrepancy of species composition between regeneration phase and young forest was also found by Ackzell et al. (Citation1993) in northern Sweden. However, Bergquist et al. (Citation2017) found regeneration success above 70% in most regions of Sweden in five growing seasons after clear-cut. In their inventory, both conifer and broadleaf saplings were considered as crop trees, although less than 20% of the stems were of broadleaved species. These results indicate that even if there are relatively high survival in five years after clear-cut, the state of young forests (1–4 m mean height) can be different from what was intended in the regeneration. Considering 1000 TPH as a criterion to be dominant, we found high regeneration success for Norway spruce in the southern and central regions but only 62–68% of the stands regenerated with pine had more than 1000 Scots pine trees. Therefore, this study indicates that a relatively large proportion of the stands regenerated with Scots pine may not develop into pine-dominated stands.
The discrepancy between species used in regeneration and species dominating in the young stands may either be caused by low planting density or high mortality. Pine weevil is considered the biggest source of mortality for young conifer seedlings in southern and central Sweden (Day et al. Citation2004; Örlander and Nilsson Citation1999) and it could cause up to 90% of mortality if no treatment is applied (Örlander and Nilsson Citation1999; von Sydow Citation1997). Moreover, browsing by ungulates could also causes mortality of seedlings (Bergquist et al. Citation2003; Bergquist and Örlander Citation1998; Bergström and Bergqvist Citation1997; Palmer and Truscott Citation2003; Miller et al. Citation1982). Severe damage has been reported on newly-planted pine seedlings by roe deer (Bergström & Bergqvist, Citation1997; Bergquist and Örlander Citation1996) and moose (Bergquist & Örlander, Citation1998) which increases mortality (Edenius et al. Citation1994; Wallgren et al. Citation2002). Experimental evidence has showed that roe deer can uproot half of the regenerated pine within a few weeks after planting (Miller et al. Citation1982) and for other species (Harmer Citation2001; Shimoda et al. Citation1994). Also, abiotic stress, such as frost and drought may make seedlings susceptible to other damage agents which may cause mortality (Burdet Citation1990; Nilsson et al. Citation2006).
The discrepancy between intention in the regeneration phase and tree species composition in young forest will not only result in a loss of invested money in planted seedlings but can also negatively influence future stand productivity and sustainable production of raw material and long-term economy. Species selection for regeneration should be done based on site suitability and experimental evidence rather than by tradition to improve regeneration success. For example, mixed species stands can be an insurance if one of the tree-species is heavily damaged. Holmström et al. (Citation2018) showed that mixtures of Scots pine and Norway spruce had almost the same production as the best monoculture but there are other mixtures that also could be tested in the future.
We found pine as a dominant regeneration species in northern Sweden whereas it was less used in southern Sweden. The high risk for browsing by ungulates probably contributes to the choice of spruce over pine in southern Sweden (Lodin Citation2020). However, Scots pine is heavily browsed also in northern Sweden where it was extensively used. A possible explanation for this may be the documented poor growth of spruce in the interior of northern Sweden (Nilsson et al. Citation2011). Recent unpublished studies from SLU have shown that in contrast to current ideas, production of Scots pine is better than Norway spruce on poor and medium fertile sites in southern Sweden and similar on relatively fertile sites unpublished). Therefore, it is not only for biodiversity and spread of risks that it is important to increase the use of Scots pine in southern Sweden but also because of sustainable wood production.
Future forest species composition considering the different weighting of damaged trees
To our knowledge, there are no studies regarding the contribution of damaged saplings to future crop-trees. Because many Scots pine trees were damaged, the proportion of broadleaved crop trees increased when damaged Scots pine trees were not considered as potential crop-trees. The reason not to include damaged pine saplings as crop trees is reduced growth due to ungulates browsing (Bergquist et al. Citation2003; Gill Citation1992). Growth reduction is likely to provide a competitive advantage to neighbouring broadleaves. Broadleaves, and especially birch which was the most common broadleaved tree species in this study, are also preferred by ungulates but birch has a high capacity to tolerate high browsing pressure without significant reduction in its biomass production (Danell et al. Citation1985; Persson et al. Citation2005). Browsed shoots of birches can continue to produce biomass within browsing height for a longer time compared to Scots pine (Danell et al. Citation1985). Because of their compensatory growth capacity, flexible growth pattern, and high absolute number, broadleaves may overtop damaged Scots pine trees. Our results indicate a higher amount of mixed forest in all regions of Sweden irrespective of whether damaged trees become crop trees or not. Therefore, this study indicates that future forest stands in Sweden will be dominated by Norway spruce and mixed-species stands but a reduction in the proportion of Scots pine-dominated stands.
Uncertainty in future tree species composition
The Äbin inventory was designed to assess moose browsing damage. In this study, we evaluated tree species composition which is a by-product of this data. Clearly, it is not possible to confidently predict future stand structure based on this data alone. However, our results show that future Swedish forests may be dominated by spruce and mixed-species forests and that pine-dominated forests may decrease in abundance. An increased broadleaved component in future stands may increase biodiversity as well as other ecosystem services such as recreation, resilience to disturbances, and water quality (Ammer et al. Citation2006; Chauvat et al. Citation2011; Felton et al. Citation2010; Felton et al. Citation2019; Foley et.al., Citation2005). However, a significant decrease in pine-dominated stands in Sweden may also reduce biodiversity since many species are dependent on this forest type (Petersson Citation2019). It might also impact recreational activities such as hunting and berry picking which depend on pine-dominated stands. However, it is difficult to predict the future of young mixed forest stands. Future stand structure depends on future forest management as well as ecological and environmental interactions. Broadleaved trees may be removed in pre-commercial and commercial thinning, resulting in conifer dominance in the mature stands. Low density spruce and pine plantations can also result in low density monocultures or conifer mixtures that are managed without thinning.
Conclusion
Our study provides evidence that the species composition of young forests deviates from the regeneration-phase intention where monocultures of Scots pine and Norway spruce probably to a large extent was the goal. Instead, young forests are dominated by Norway spruce with and mixed conifer and broadleaved forests. Failures in Scots pine regenerations, may change the species composition of future forests. The observed low abundance of Scots pine dominated stands may reduce production and profits as well as biodiversity and recreational opportunities. As spruce and birch benefit from dense ungulate populations, the species diversity of Swedish forests may be at stake (Felton et al. Citation2019). However, it is difficult to predict future stand structure because it is dependent on future management and other events taking place during the rotation.
Acknowledgments
We thank Skogsstyrelsen for providing access to the Äbin data. Thanks to all the field crews and other people associated with the data collection process. We also want to thank Emma Holmström for her suggestions during the data analysis. The FRAS (Future Silviculture of Southern Sweden) research program funded Mostarin Ara to conduct this study.
Disclosure statement
No potential conflict of interest was reported by the author(s).
References
- Ackzell L, Elfving B, Lindgren D. 1993. Occurrence of naturally regenerated and planted main crop plants in plantations in boreal Sweden. For Ecol Manag. 65(2–3):105–113.
- Ammer S, Weber K, Abs C, Ammer C, Prietzel J. 2006. Factors influencing the distribution and abundance of earthworm communities in pure and converted Scots pine stands. Appl Soil Ecol. 33(1):10–21. doi:https://doi.org/10.1016/j.apsoil.2005.09.005.
- Bergquist J, Bergström R, Zakharenka A. 2003. Responses of young Norway spruce (Picea abies) to winter browsing by roe deer (Capreolus capreolus): Effects on height growth and stem morphology. Scand J For Res. 18(4):368–376. doi:https://doi.org/10.1080/0282758031005431.
- Bergquist J, Fries C, Svensson L. 2017. Skogsstyrelsens återväxtuppföljning Resultat från 1999–2016.
- Bergquist J, Kalén C, Berglund H. 2011. Hjortdjurens inverkan på tillväxt av produktionsträd och rekrytering av betesbegärliga trädslag. Jönköping: Skogsstyrelsen.
- Bergquist J, Kullberg Y, Örlander G. 2001. Effects of shelterwood and soil scarification on deer browsing on planted Norway spruce Picea abies L. (Karst) seedlings. Forestry. 74(4):359–367. doi:https://doi.org/10.1093/forestry/74.4.359.
- Bergquist J, Örlander G. 1996. Browsing deterrent and phytotoxic effects of roe deer repellents on Scots pine and Norway spruce seedlings. Scand J For Res. 11:145–152.
- Bergquist J, Örlander G. 1998. Browsing damage by roe deer on Norway spruce seedlings planted on clearcuts of different ages: 2. Effect of seedling vigour. For Ecol Manag. 105(1–3):295–302. doi:https://doi.org/10.1016/S0378-1127(97)00296-X.
- Bergström R, Bergqvist G. 1997. Frequencies and patterns of browsing by large herbivores on conifer seedlings. Scand J For Res. 12(3):288–294. doi:https://doi.org/10.1080/02827589709355412.
- Bergström R, Hjeljord O. 1987. Moose and vegetation interactions in northwestern Europe and Poland. Swedish Wildlife Research. 1:213–228.
- Burdet AN. 1990. Physiological processes in plantation establishment and their development of specifications for forest planting stock. Can J For Res. 20:415–427.
- Cederlund G. 1983. Home range dynamics and habitat selection by roe deer in a boreal area in central Sweden. Acta Theriol. 28:443–460. doi:https://doi.org/10.4098/at.arch.83-39.
- Cederlund G, Bergström R. 1996. Trends in the moose—forest system in Fennoscandia, with special reference to Sweden. Conservation of Faunal Diversity in Forested Landscapes. 265–281. doi:https://doi.org/10.1007/978-94-009-1521-3_10.
- Chauvat M, Titsch D, Zaytsev AS, Wolters V. 2011. Changes in soil faunal assemblages during conversion from pure to mixed forest stands. For Ecol Manag. 262(3):317–324. doi:https://doi.org/10.1016/j.foreco.2011.03.037.
- Danell K, Danel HK, Bergstrom R. 1985. Interaction between browsing moose and two species of birch in Sweden. Ecology. 66(6):1867–1878. doi:https://doi.org/10.2307/2937382.
- Day KR, Nordlander G, Kenis M, Halldorson G. 2004. General Biology and Life Cycles of Bark Weevils. In Bark and Wood Boring Insects in Living Trees in Europe, a Synthesis. https://doi.org/https://doi.org/10.1007/978-1-4020-2241-8_14.
- Edenius L, Danell K, Nyquist H. 1994. Effects of simulated moose browsing on growth, mortality, and fecundity in Scots pine: relations to plant productivity. Can J For Res. 25(4):529–535. doi:https://doi.org/10.1139/x95-060.
- ESRI. 2010. ArcGIS Desktop release 10. Environment System Release Institute.
- Felton A, Lindbladh M, Brunet B, Fritz Ö. 2010. Replacing coniferous monocultures with mixed-species production stands: An assessment of the potential benefits for forest biodiversity in Northern Europe. Forest Ecology &Management. 260:939–947.
- Felton A, Petersson L, Nilsson O, et al. 2019. The tree species matters: biodiversity and ecosystem service implications of replacing Scots pine production stands with Norway spruce. Ambio. 49(5):1035–1049. doi:https://doi.org/10.1007/s13280-019-01259-x.
- Foley JA, DeFries R, Asner GP, Barford C, Bonan G, Carpenter SR, Chapin FS, Coe MT, Daily GC, Gibbs HK, et al. 2005. Global consequences of land use. Science. 309(5734):570–574. doi:https://doi.org/10.1126/science.1111772.
- Gill RMA. 1992. A review of damage by mammals in north temperate forests:3. impact on trees and forests. Forestry. 65:363–388.
- Harmer R. 2001. The effect of plant competition and simulated summer browsing by deer on tree regeneration. J Appl Ecol. 38(5):1094–1103. doi:https://doi.org/10.1046/j.1365-2664.2001.00664.x.
- Holmström E, Gålnander H, Petersson M. 2018. Within-site variation in seedling survival in Norway spruce plantations. Forests. 10(2):8–10.
- Kabrick J, Dey D, Gwaze D. 2006. Shortleaf Pine Restoration and Ecology in the Ozarks: Proceedings of a Symposium. The Missouri department of conservation, USA.
- Lodin I. 2020. Current versus alternative forest management practices in southern Sweden. PhD dissertation, Swedish University of Agricultural Sciences.
- Loftis DL, McGee CE. 1993. Symposium Proceedings, 1992 September 8-10, Knoxville, Tennessee.
- Månsson J, Kalén C, Kjellander P, Andrén H, Smith H. 2007. Quantitative estimates of tree species selectivity by moose (Alces alces) in a forest landscape. Scandinavian Journal of Forest Research. 22(5):407–414. doi:https://doi.org/10.1080/02827580701515023.
- Miller GR, Kinnaird JW, Cummins RP. 1982. Liability of saplings to browsing on a red deer range in the Scottish highlands. J Appl Ecol. 19:941–951.
- Mysterud A. 2000. Diet overlap among Fennoscandia. Oecologia. 124(1):130–137.
- Nilsson U, Elfving B, Karlsson K. 2012. Productivity of Norway spruce compared to Scots pine in the interior of Northern Sweden. Silva Fenn. 46(2):197–209. doi:https://doi.org/10.14214/sf.54.
- Nilsson U, Fahlvik N, Johansson U, Lundström A, Rosvall O. 2011. Simulation of the effect of intensive forest management on forest production in Sweden. Forests. 2(1):373–393. doi:https://doi.org/10.3390/f2010373.
- Nilsson U, Luoranen J, Kolström T, Örlander G, Puttonen P. 2010. Reforestration with planting in Northern Europe. Scandinavian Journal of Forest Research. 25(4):283–294.
- Nilsson U, Örlander G, Karlsson M. 2006. Establishing mixed forests in Sweden by combining planting and natural regeneration-effects of shelterwoods and scarification. For Ecol Manag. 237(1–3):301–311.
- Örlander G, Nilsson U. 1999. Effect of reforestation methods on pine weevil (Hylobius abietis) damage and seedling survival. Scandinavian Journal of Forest Research. 14(4):341–354. doi:https://doi.org/10.1080/02827589950152665.
- Palmer SCF, Truscott AM. 2003. Browsing by deer on naturally regenerating Scots pine (Pinus sylvestris L.) and its effects on sapling growth. For Ecol Manag. 182(1–3):31–47. doi:https://doi.org/10.1016/S0378-1127(03)00026-4.
- Persson IL, Kjell D, Roger B. 2005. Different moose densities and accompanied changes in tree morphology and browse production. Ecol Appl. 15(4):1296–1305.
- Petersson L. 2019. Replacing Scots pine with Norway spruce. PhD dissertation. Swedish University of Agricultural Science.
- Shimoda K, Kimura K, Kanzaki M, Yoda K. 1994. The regeneration of pioneer tree species under browsing pressure of sika deer in an evergreen oak forest. Ecol Res. 9(1):85–92. doi:https://doi.org/10.1007/BF02347245.
- Swedish National Forest Survey. 2010. http://www.slu.se/en/collaborative-centres-andprojects/swedish-national-forest-inventory/.
- Sydow V. 1997. Abundance of pine weevil (Hylobius abietis) and damage to conifer seedlings in relation to the silvicultural practices. Scandinavian Journal of Forest Researches. 12:157–167.
- Wallgren M, Bergström R, Bergqvist G, Olsson M. 2002. Spatial distribution of browsing and tree damage by moose in young pine forests, with implications for the forest industry. For Ecol Manag. 305:229–238. doi:https://doi.org/10.1016/j.foreco.2013.05.057.
Appendix
Äbin data
Äbin is an inventory method developed by the Swedish Forest Agency and the Swedish University of Agricultural Sciences (SLU) to estimate browsing damage on young trees. One half of Sweden is systematically inventoried each year, thus covering the entire country every second year. Every year about 10,000 square sampling regions (1 × 1 km) are randomly selected. Within these squares, all stands with a potential mean height between 1 and 4 metre are identified using remote sensing methods. All identified stands are visited and if the stand qualifies as an Äbin stand, it is included in to the inventory. Depending on the stand size, 1–15 plots are placed within the stand in a square grid and each plot location is recorded using a GPS.
The plots have a radius of 3.5 m and trees within the plots are recorded if their height is above the “half height”, which is the mean height of the two tallest conifer trees in the plot (). If conifers are absent from the plot, the two tallest broadleaved trees are used to determine the half height. The number of trees higher than the half height is recorded by species. In this study, the numbers of pine, spruce and broadleaves (a class heavily dominated by birch) were analysed separately.