
Abstract
Background: Glucose-6-phosphate dehydrogenase (G6PD) deficiency is the most common inherited enzyme defect worldwide. There is a growing scientific evidence for a protective role of G6PD deficiency against carcinogenesis. In this retrospective analysis, we tested the hypothesis that G6PD deficiency may reduce the risk of developing cancer in a tissue-specific manner.
Material and methods: The study was conducted using data from 11,708 subjects undergoing gastrointestinal endoscopic procedures between 2002 and 2018 and tested for G6PD status in a teaching hospital of Northern Sardinia, Italy.
Results: A 40% reduction of risk for cancer of endodermal origin was observed among G6PD-deficient patients compared with subjects with normal enzyme activity (relative risk (RR) 0.61, 95% confidence interval (CI) 0.47–0.80) in both genders, confirmed by multivariable generalized linear regression after adjusting for age, sex, smoking habits, body mass index, diabetes and socio-economic status. The ‘protective’ effect of G6PD deficiency was larger for gastric cancer (RR 0.41, 95% CI 0.18–0.99), hepatocellular carcinoma (RR 0.48, 95% CI 0.26–0.92) and colorectal cancer (RR 0.72, 95% CI 0.53–0.98), while a non-significant risk was observed for breast, prostate, lung, hematopoietic and metastases (primary site unknown).
Conclusions: Our results suggest a reduced susceptibility to develop cancers, mostly of endodermal origin (stomach, colon and liver), but not of ectodermal/mesodermal origin, in carriers of G6PD deficiency. The effects of G6PD deficiency on carcinogenesis need further studies to better understand how cancer cells originating from different germ layers use pentose phosphate pathway to proliferate.
Introduction
The hypothesis that deficiency of glucose-6-phosphate dehydrogenase (G6PD), a cytoplasmic enzyme that catalyzes the first and rate-limiting step of the pentose phosphate pathway (PPP), may be associated with reduced tumorigenesis was object of debate for the last half century, and recently has spurred a renewed interest in medical literature [Citation1–6]. G6PD deficiency was claimed to counteract the development of certain cancers through downregulation of PPP and consequent shortage of the extra-mitochondrial nicotinamide adenine dinucleotide phosphate (NADPH) as well as the pentose sugars needed by rapidly proliferating cancer cells to synthesize DNA [Citation7,Citation8]. The putative anti-carcinogenic effect of G6PD deficiency has been investigated in animal models, where a reduced NADPH production resulting from PPP downregulation has often been documented, but in general these studies have yielded mixed results, likely due to interspecies diversity [Citation9,Citation10]. While in bacteria and yeasts complete G6PD deletion does not affect viability [Citation11] in the nematode Caenorhabditis elegans, whose G6PD protein shows 56% amino-acid sequence homology with the corresponding human counterpart [Citation12], microRNA-induced G6PD gene disruption dramatically impairs oogenesis and embryogenesis [Citation13]. Similarly, in mice, severe G6PD deficiency leads to embryonic lethality [Citation14], whereas milder G6PD deficiency exacerbates brain oxidative damage [Citation15,Citation16]. On the contrary, overexpression of the enzyme in Drosophila melanogaster is associated with 30% lifespan extension [Citation17], and in mice even a modest increase in G6PD activity is associated with increased levels of NADPH, and a general strengthening of antioxidant defense [Citation16]. These findings suggest an important role of G6PD activity during embryogenesis that may be related mainly to protection against oxidative stress and DNA oxidative damage. Despite the rising interest in altered PPP metabolism in cancer [Citation18], a recent G6PD knockout murine model failed to provide evidence of any effect on tumor development [Citation9], while confirming a modest increase in health span probably through raised NADPH levels and protection from the deleterious effects of free radicals. In humans, evidence of overexpression of G6PD activity was found in several site-specific tumors, including gastric and colorectal cancers [Citation19,Citation20].
Epidemiological studies exploring the susceptibility to cancer in subjects carrying an inherited defect that reduces G6PD enzyme activity have been conducted in populations where the frequency of the genetic defect is relatively higher, mostly because of malaria resistance: sub-Saharan Africa, South-East Asia and the Mediterranean island of Sardinia. In the latter, the overall prevalence of G6PD deficiency ranges between 12% and 24%, and in up to 95% of cases is due to the G6PD Mediterranean (Med) variant (a C → T transition at nucleotide 563) [Citation21]. The first hint that G6PD deficiency can offer protection against carcinogenesis emerged more than half a century ago when Sulis reported a lower cancer prevalence in Sardinia compared to the rest of Europe where G6PD deficiency is rare [Citation1]. This claim was further supported by large-scale epidemiological studies conducted in Sardinia in the 1980s and 1990s by Cocco et al. addressing cancer mortality among G6PD-deficient men [Citation3,Citation4]. These early investigations suggested that cancer rate was lower in this group, but subsequent epidemiological studies failed to confirm the initial findings, especially when the analysis focused on site-specific cancers [Citation2,Citation22]. Therefore, after more than five decades of investigations, the hypothesis of an inverse correlation between G6PD deficiency and cancer susceptibility is still insufficiently supported by empirical evidence. The past few years, however, have witnessed a renovated interest in the G6PD/cancer hypothesis, especially because PPP has been shown to exert an unexpected crucial role in cancer cell growth by providing both nucleotide precursors, needed for proliferation, and the high levels of NADPH necessary for the synthesis of metabolites such as endogenous fatty acids and cholesterol [Citation18,Citation23,Citation24]. Recently, we have reported that G6PD deficiency was associated with a reduced risk of colorectal cancer and hepatocellular carcinoma [Citation25,Citation26].
Based on our previous findings, we tested the G6PD/cancer hypothesis in a wider range of tumors originating from various germ layers.
Methods
Study design
This study was a retrospective case control one-site study.
Data sources
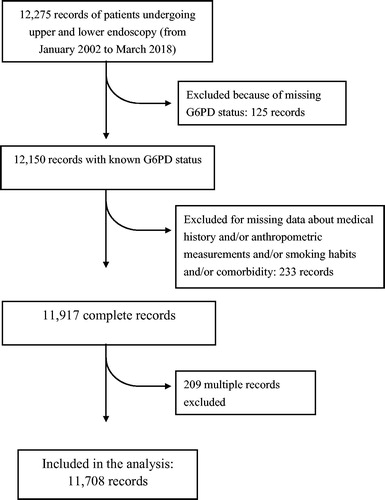
We used data from a computerized database of patients from Northern Sardinia undergoing upper and lower endoscopy for any reason, from January 2002 to March 2018. The database is currently updated, and smaller subsets of it have previously been used for epidemiological and observational studies [Citation25,Citation26]. The patients included in the database are broadly representative of the overall population of Northern Sardinia. The inclusion and exclusion criteria of the study are shown in . One hundred and twenty-five records from the original database were excluded due to the lack of G6PD status, and 233 additional records were excluded due to missing data about medical history (including information concerning major risk factors for cancer in general, such as smoking habits, low socioeconomic status and diabetes mellitus), anthropometric measurements and comorbidity. Finally, in 209 patients with multiple examinations within the study period, only the last record corresponding to a diagnosis of any cancer was retained for the analysis. Socio-economic status (SES) was defined according to current or past occupation and divided into four groups as previously described although, in the final analysis, these groups were stratified into two categories, i.e., high-SES and low-SES [Citation25,Citation26].
Figure 1. Flowchart of study inclusions and exclusions.
G6PD assay
G6PD activity had been determined in all patients by a quantitative assay (Nurex Diagnostici) based on the ratio between G6PD/6GPD in erythrocytes, as reported and validated in other studies [Citation25,Citation26]. According to the manufacturer’s instructions, G6PD enzyme deficiency was defined as a ratio ≤0.1. Results were expressed qualitatively as a binary variable (normal/deficient). Molecular analysis for G6PD deficient patients was not available.
Ethical considerations
The study protocol was approved by the local ethics committee Comitato di Bioetica, Azienda Ospedaliero Universitaria di Sassari, Italy (Prot No. 3004/CE, 2016), and the 1975 Declaration of Helsinki guidelines were followed. From each participant was obtained a written informed consent.
Statistical analysis
Categorical variables, such as frequency of cancers, patients’s age grouped in 10-year intervals, and G6PD status, were described by using percentage. The association between G6PD deficiency and cancer risk was tested by calculating unadjusted and adjusted relative risk (RR) and their 95% confidence intervals (95% CIs) through a multivariable generalized linear model regression with a Poisson distribution and a log link. Models were adjusted for the following covariates: (i) patient’s age and sex, (ii) smoking habits (never smokers, current or former smokers), (iii) body mass index calculated as weight/height2 (kg/m2), (iv) diabetes (type 1 and type 2 pooled together in a dichotomic variable) and (v) SES. The control group for each site-specific cancer included subject’s with a negative clinical history of cancer at the time of the study. Relative risks and their 95% CIs were calculated by exponentiating the regression coefficients. All statistical analyses were carried out using SPSS statistical software (version 16·0, Chicago, IL, USA) and two-sided p-values lower than .05 were considered statistically significant.
Results
A total of 11,708 clinical records of patients (4468 men and 7240 women) who underwent upper endoscopy and colonoscopy were available for the analysis. One thousand four hundred and twenty-two patients (12.1%) had G6PD deficiency and 10,286 patients were normal for G6PD enzyme activity in line with previously reported data [Citation27]. G6PD enzyme deficiency was detected in 8.6% of men and in 14.3% of women, as expected for a X-linked trait. The frequency of all cancers according to age decades and sex is reported in . No significant differences were found across age groups. The frequency of cancers according to the specific tissue origin revealed a predominance of colorectal cancer (3.9%), followed by breast cancer (3.7%), prostate cancer (1.5%) and hepatocellular carcinoma (1.4%). The unadjusted RR and 95% CI for cancers of specific tissue origin in patients with G6PD deficiency and those with normal enzyme activity are reported in .
Table 1. Demographic characteristics, cancer frequency and glucose-6-phosphate dehydrogenase frequency in 11,708 patients undergoing endoscopy included in the study.
Table 2. Unadjusted relative risk and 95% confidence interval for all-cancer among 11,708 patients with and without glucose-6-phosphate dehydrogenase (G6PD) deficiency, according to the embryological origin of cancer.
Overall, cancers of endodermal origin were less frequent among G6PD deficient patients (unadjusted RR: 0.61; 95% CI 0.47–0.80) compared with controls, whereas for cancers of ectodermal/mesodermal origin the RR was close to 1 (unadjusted RR: 1.01; 95% CI 0.80–1.27). shows the absolute risk and the RRs calculated by the generalized linear regression model. After adjusting for age, sex, smoking habits, diabetes and SES, G6PD deficiency remained associated with a decreased risk for cancers although the effect size was larger, in descending order, for gastric cancer (RR = 0.41, 95% CI 0.18–0.99), hepatocellular carcinoma (RR = 0.48, 95% CI 0.26–0.92) and colorectal cancer (RR = 0.72, 95% CI 0.53–0.98). The other site-specific cancers, of ectodermal/mesodermal origin, did not display significantly increased RR.
Table 3. Absolute and relative risk for site-specific cancers among 11,708 patients with and without glucose-6-phosphate dehydrogenase (G6PD) deficiency.
Discussion
The hypothesis that inherited G6PD deficiency may contrast the development of cancer emerged nearly 50 years ago and several epidemiological studies have tried to evaluate its plausibility. Much of the research was conducted in the Sardinia population where the frequency of this hereditary condition, due to the G6PD Med variant, is one of the highest in the world. In previous studies, using a relatively large computerized database, we provided evidence in favor of the existence of an inverse relationship between G6PD deficiency and the risk of colorectal and hepatocellular carcinoma [Citation25,Citation26]. Studies targeted to other site-specific cancers have been less frequent. The present investigation aimed at widening the spectrum of malignancies tested for the association with G6PD status, by controlling for a number of risk factors common to several types of cancers such as age, sex, smoking habits, BMI, diabetes and socioeconomic status. Based upon the findings of this study, we argue that carriers of G6PD deficiency may benefit of a reduced cancer susceptibility, although the effect size varies with the type of cancer. Specifically, a considerable risk reduction was observed for gastric and colorectal cancer and hepatocellular carcinoma, while the risk of developing breast, prostate, lung, hematopoietic cancers as well as metastases does not appear to be significantly influenced by G6PD status. The effect of the chronic enzyme deficiency seems to be essentially restricted to malignancies of endodermal origin, while those of ectodermal/mesodermal origin are apparently unaffected.
The observation that G6PD deficiency, although implying a lower defense from oxidative stress, can paradoxically entail a lower risk of carcinogenesis dates back a long time ago and raised a debate over decades, with pro and con arguments. Recently, the interest in this controversy seems to be revived also as a result of the availability of new in vitro and in vivo models of G6PD deficiency [Citation6]. These models suggested that the damage caused by PPP disruption is essentially genotoxic, namely DNA is the main target of oxidative damage [Citation13]. For example, G6PD-knockdown cells in C. elegans display DNA oxidative damage but not lipid peroxidation, indicating a predominant nucleic acid insult [Citation13].
G6PD is not expressed equally by all types of cancers but appears to be overexpressed in more aggressive tumors where cells grow very rapidly [Citation28]. It can be hypothesized that downregulation of the non-oxidative branch of PPP induced by G6PD deficiency, and the consequent reduction of ribose synthesis, predominantly hinders fast-growing tumors such as those of the digestive system [Citation29] where extra ribose is required to sustain replication. The situation could be similar to G6PD deficiency in embryonic tissues where the result is often non-viability [Citation13]. Moreover, it has recently been shown that endodermal differentiation from embryonic stem cells (ESCs) requires the activation of PPP [Citation30]. It is reasonable to speculate that cancers derived from endoderm maintain some features of the original tissue. Based on that, endodermal tumors, depending strictly from PP pathway, may experience a more difficult growth in G6PD deficient patients compared to ectodermal/mesodermal cancers. According to this conjecture, endodermal cancers would be less capable to activate alternative pathways (e.g., mitochondria) to produce redox equivalents necessary for the synthesis of membrane components such as fatty acids and cholesterol [Citation31]. Furthermore, since it has been reported that G6PD is implicated in the apoptotic machinery, as it downregulates the expression of apoptosis inhibitory factors Bcl-2 and Bcl-xL [Citation10,Citation32], while increasing the expression of Fas protein compared with expression levels observed in controls [Citation33], we may speculate that tissues where the enzyme is deficient may counteract the apoptotic inhibition that characterizes the molecular events of carcinogenesis. The mammalian target of rapamycin (mTOR) and Myc transcription factors regulate PPP and controls cell proliferation and growth and normally inhibit the differentiation of ESCs into endoderm [Citation34]. However, mouse ESC carrying a deletion in G6PD gene are able to express endodermal-specific markers such as Sox 17 [Citation30]. These findings suggest that PPP is crucial for the differentiation of ESC into definitive endoderm cells, and cell of endodermal origin with deficient G6PD may have a lower carcinogenetic potential than cells with normal G6PD activity. This may be due to the reduced production of pentose sugars, DNA precursors, rather than to the reduced antioxidant defense resulting from NADPH shortage [Citation30].
The finding that lung cancer, in our study, seems to be an exception as its development is not affected by G6PD deficiency, as already reported [Citation35], may be due to several causes: this cancer may originate from distinct types of cells but mostly from ciliated airway epithelial cells that are probably a terminally differentiated lineage [Citation36]. As a consequence, their proliferative rate was estimated as a half-life of 6 months in the trachea and 17 months in the lungs, much longer than the three days estimated for mature enterocytes in the villi tips [Citation37]. This suggests that the proliferative stimuli capable to amplify the hypothetical genotoxic damage induced by G6PD deficiency may be greater for enteric cells than for cell derived from the respiratory tract. Moreover, lung cancer cells as well as cancer cells of ectodermal origin may be less dependent from NADPH compared to gastric, colonic and liver tumor cells, thus overcoming the energy restriction induced by the G6PD enzyme deficiency. In any case, the protective effect of G6PD against cancers of endodermal origin would hardly influence significantly the survivorship curve of the population [Citation38] since deaths from these tumors represent a near-negligible proportion of the overall mortality in the general population.
A number of caveats may concern the validity of our study. First, a detailed distribution of potential confounders between normal and G6PD deficient patients was not entirely checked.
Second, it might be objected to the use of a database of patients undergoing gastrointestinal endoscopic examination instead of a random sample. However, in our opinion, there is no a priori reason to think this may have introduced any serious bias. Due to its elevated frequency in the general Sardinian population, the enzyme defect is routinely assessed since childhood. Therefore, all patients recruited in the study undergoing endoscopic procedure, were tested for the enzymatic activity at least once in their lifetime. More importantly, the frequency of G6PD deficiency, in our cohort, was very close to the average frequency found in previous studies in the Sardinian population [Citation21,Citation39].
Other limitations of the study are the lack of detailed data regarding other risk factors or comorbidities usually associated with gastric cancer, including H. pylori infection, dietary habits, medications, etc. However, we previously observed that H. pylori infection is not influenced by G6PD status [Citation40]. On the other hand, for hepatocellular carcinoma, usually known to be strongly related to cirrhosis and virus infections, the reduced risk produced by G6PD deficiency remains significant also after adjusting for these covariates [Citation26]. Finally, some of the cancer-specific analyses (e.g., lung) were based on small numbers, which makes it difficult to show statistical significance even though the RR is high.
In conclusion, the results of our study indicate that an inherited mutation of the G6PD enzyme can delay or reduce the risk to develop a variety of cancers especially those of endodermal origin. The protective effect of G6PD deficiency against the development of various cancers observed in several epidemiological studies over the past years, is consistent with preclinical studies. Moreover, the demonstration of a major role of G6PD in cancer cell metabolism stimulated investigators to search for strategies to selectively inhibit G6PD activity in patients with malignancy [Citation41,Citation42]. In recent years, the synthesis of competitive inhibitors capable to directly disrupt the catalytic activity of G6PD [Citation43,Citation44], and the use of antisense oligonucleotides to downregulate the expression of G6PD by targeting its mRNA [Citation45], have attracted considerable interest. Although a number of compounds with inhibitory effect on G6PD have been identified, their use is limited in clinical setting by their severe adverse effects [Citation46]. Currently, new potential inhibitors with less undesired side-effects are being evaluated by screening chemical libraries [Citation46]. This approach may prove promising since G6PD inhibition was reported to be 100- to 1000-fold more powerful than for well-known G6PD inhibitors such as dehydroepiandrosterone and 6-aminonicotinamide [Citation46]. In addition, G6PD deficiency decreases cancer cells migration and proliferation [Citation47], and even enhances sensitivity of cancer cell to chemotherapy [Citation48]. Our findings provide an additional source of information on the potential risks and benefits of targeting G6PD in the treatment of a variety of cancers, although further research is needed to fully understand the relationship between G6PD deficiency and cancer.
Disclosure statement
No potential conflict of interest was reported by the authors.
References
- Sulis E. G6PD deficiency and cancer. The Lancet. 1972;1:1185.
- Ferraris AM, Broccia G, Meloni T, et al. Glucose-6-phosphate dehydrogenase deficiency and incidence of hematologic malignancy. Am J Hum Genet. 1988;42:516–520.
- Cocco PL. Does G6PD deficiency protect against cancer? A critical review. J Epidemiol Community Health. 1987;41:89–93.
- Cocco PL, Dessí S, Avataneo G, et al. Glucose-6-phosphate dehydrogenase deficiency and cancer in a Sardinian male population: a case-control study. Carcinogenesis. 1989;10:813–816.
- Kowalik MA, Columbano A, Perra A. Emerging role of the pentose phosphate pathway in hepatocellular carcinoma. Front Oncol. 2017;7. doi:10.3389/fonc.2017.00087
- De Santis MC, Porporato PE, Martini M, et al. Signaling pathways regulating redox balance in cancer metabolism. Front Oncol. 2018;8. doi:10.3389/fonc.2018.00126
- Luzzatto L, Metha A. Glucose-6-phosphate dehydrogenase deficiency. In: Scriver CR, Beaudet AL, Sly WS, et al., editors. The metabolic basis of inherited diseases. New York: McGraw-Hill, Inc.; 1989.
- Patra KC, Hay N. The pentose phosphate pathway and cancer. Trends Biochem Sci. 2014;39:347–354.
- Nóbrega-Pereira S, Fernandez-Marcos PJ, Brioche T, et al. G6PD protects from oxidative damage and improves healthspan in mice. Nat Commun. 2016;7:10894.
- Hu T, Zhang C, Tang Q, et al. Variant G6PD levels promote tumor cell proliferation or apoptosis via the STAT3/5 pathway in the human melanoma xenograft mouse model. BMC Cancer. 2013;13:251.
- Fraenkel DG, Vinopal RT. Carbohydrate metabolism in bacteria. Annu Rev Microbiol. 1973;27:69–100.
- C. elegans Sequencing Consortium. Genome sequence of the nematode C. elegans: a platform for investigating biology. Science. 1998;282:2012–2018.
- Yang HC, Chen TL, Wu YH, et al. Glucose 6-phosphate dehydrogenase deficiency enhances germ cell apoptosis and causes defective embryogenesis in Caenorhabditis elegans. Cell Death Dis. 2013;4:e616.
- Longo L, Vanegas OC, Patel M, et al. Maternally transmitted severe glucose-6-phosphate dehydrogenase deficiency is an embryonic lethal. EMBO J. 2002;21:4229–4239.
- Mejías R, Villadiego J, Pintado CO, et al. Neuroprotection by transgenic expression of glucose-6-phosphate dehydrogenase in dopaminergic nigrostriatal neurons of mice. J Neurosci. 2006;26:4500–4508.
- Cao L, Zhang D, Chen J, et al. G6PD plays a neuroprotective role in brain ischemia through promoting pentose phosphate pathway. Free Radic Biol Med. 2017;112:433–444.
- Legan SK, Rebrin I, Mockett RJ, et al. Overexpression of glucose-6-phosphate dehydrogenase extends the life span of Drosophila melanogaster. J Biol Chem. 2008;283:32492–32499.
- Jiang P, Du W, Wu M. Regulation of the pentose phosphate pathway in cancer. Protein Cell. 2014;5:592–602.
- Yu J, Liang Q, Wang J, et al. REC8 functions as a tumor suppressor and is epigenetically downregulated in gastric cancer, especially in EBV-positive subtype. Oncogene. 2017;36:182–193.
- Van Driel BE, Valet GK, Lyon H, et al. Prognostic estimation of survival of colorectal cancer patients with the quantitative histochemical assay of G6PDH activity and the multiparameter classification program CLASSIF1. Cytometry. 1999;38:176–183.
- Fiorelli G, Meloni T, Palomba V, et al. Gene frequency of glucose-6-phosphate dehydrogenase (G6PD) polymorphic variants in Sardinia. Gene Geogr. 1990;4:139–142.
- Cocco PL, Todde P, Fornera S, et al. Mortality in a cohort of men expressing the glucose-6-phosphate dehydrogenase deficiency. Blood. 1998;91:706–709.
- Riganti C, Gazzano E, Polimeni M, et al. The pentose phosphate pathway: an antioxidant defense and a crossroad in tumor cell fate. Free Radic Biol Med. 2012;53:421–436.
- Ai G, Dachineni R, Kumar DR, et al. Aspirin inhibits glucose-6-phosphate dehydrogenase activity in HCT 116 cells through acetylation: identification of aspirin-acetylated sites. Mol Med Rep. 2016;14:1726–1732.
- Dore MP, Davoli A, Longo N, et al. Glucose-6-phosphate dehydrogenase deficiency and risk of colorectal cancer in Northern Sardinia: a retrospective observational study. Medicine. 2016;95:e5254.
- Dore MP, Vidili G, Marras G, et al. Inverse association between glucose-6-phosphate dehydrogenase deficiency and hepatocellular carcinoma. Asian Pac J Cancer Prev. 2018;19:1069–1073.
- De Vita G, Alcalay M, Sampietro M, et al. Two point mutations are responsible for G6PD polymorphism in Sardinia. Am J Hum Genet. 1989;44:233–240.
- Buj R, Aird KM. Deoxyribonucleotide triphosphate metabolism in cancer and metabolic disease. Front Endocrinol. 2018;9:177.
- Zhou Z, Ibekwe E, Chornenkyy Y. Metabolic alterations in cancer cells and the emerging role of oncometabolites as drivers of neoplastic change. Antioxidants (Basel). 2018;7. pii: E16.
- Manganelli G, Fico A, Masullo U, et al. Modulation of the pentose phosphate pathway induces endodermal differentiation in embryonic stem cells. PLoS One. 2012;7:e29321.
- Currie E, Schulze A, Zechner R, et al. Cellular fatty acid metabolism and cancer. Cell Metab. 2013;18:153–161.
- Wang X, Liu H, Zhang X, et al. G6PD downregulation triggered growth inhibition and induced apoptosis by regulating STAT3 signaling pathway in esophageal squamous cell carcinoma. Tumor Biol. 2016;37:781–789.
- Adams JM, Cory S. The Bcl-2 apoptotic switch in cancer development and therapy. Oncogene. 2007;26:1324–1337.
- Zhou J, Su P, Wang L, et al. mTOR supports long-term self-renewal and suppresses mesoderm and endoderm activities of human embryonic stem cells. Proc Natl Acad Sci USA. 2009;106:7840–7845.
- Pisano M, Cocco P, Cherchi R, et al. Glucose-6-phosphate dehydrogenase deficiency and lung cancer: a hospital based case-control study. Tumori. 1991;77:12–15.
- Potten CS. A comprehensive study of the radiobiological response of the murine (BDF1) small intestine. Int J Radiat Biol. 1990;58:925–973.
- Mayhew TM, Myklebust R, Whybrow A, et al. Epithelial integrity, cell death and cell loss in mammalian small intestine. Histol Histopathol. 1999;14:257–267.
- Pes GM, Errigo A, Bitti A, et al. Effect of age, period and birth-cohort on the frequency of glucose-6-phosphate dehydrogenase deficiency in Sardinian adults. Ann Med. 2018;50:68–73.
- Pes GM, Bassotti G, Dore MP. Colorectal cancer mortality in relation to glucose-6-phosphate dehydrogenase deficiency and consanguinity in Sardinia: a spatial correlation analysis. Asian Pac J Cancer Prev. 2017;18:2403–2407.
- Dore MP, Marras G, Rocchi C, et al. G6PD deficiency does not enhance susceptibility for acquiring Helicobacter pylori infection in Sardinian patients. PLoS One. 2016;11:e0160032.
- Gupte SA. Glucose-6-phosphate dehydrogenase: a novel therapeutic target in cardiovascular diseases. Curr Opin Investig Drugs. 2008;9:993–1000.
- Zhang C, Zhang Z, Zhu Y, et al. Glucose-6-phosphate dehydrogenase: a biomarker and potential therapeutic target for cancer. ACAMC. 2014;14:280–289.
- Mele L, Paino F, Papaccio F, et al. A new inhibitor of glucose-6-phosphate dehydrogenase blocks pentose phosphate pathway and suppresses malignant proliferation and metastasis in vivo. Cell Death Dis. 2018;9:572.
- Jin K, Li L, Sun X, et al. Mycoepoxydiene suppresses HeLa cell growth by inhibiting glycolysis and the pentose phosphate pathway. Appl Microbiol Biotechnol. 2017;101:4201–4213.
- Cai T, Kuang Y, Zhang C, et al. Glucose-6-phosphate dehydrogenase and NADPH oxidase 4 control STAT3 activity in melanoma cells through a pathway involving reactive oxygen species, c-SRC and SHP2. Am J Cancer Res. 2015;5:1610–1620.
- Preuss J, Richardson AD, Pinkerton A, et al. Identification and characterization of novel human glucose-6-phosphate dehydrogenase inhibitors. J Biomol Screen. 2013;18:286–297.
- Fang Z, Jiang C, Feng Y, et al. Effects of G6PD activity inhibition on the viability, ROS generation and mechanical properties of cervical cancer cells. Biochim Biophys Acta. 2016;1863:2245–2254.
- Catanzaro D, Gaude E, Orso G, et al. Inhibition of glucose-6-phosphate dehydrogenase sensitizes cisplatin-resistant cells to death. Oncotarget. 2015;6:30102–30114.