
Abstract
Aim: To assess practical implications of genotype-based recall (GBR) studies, an increasingly popular approach for in-depth characterization of genotype–phenotype relationships.
Methods: We genotyped 2500 participants from the Swedish EpiHealth cohort and considered loss-of-function and missense variants in genes with relation to cardiometabolic traits as the basis for our GBR study. Therefore, we focused on carriers and non-carriers of the PPARG Pro12Ala (rs1801282) variant, as it is a relatively common variant with a minor allele frequency (MAF) of 0.14. It has also been shown to affect ligand binding and transcription, and carriage of the minor allele (Ala12) is associated with a reduced risk of type 2 diabetes. We re-invited 39 Pro12Pro, 34 Pro12Ala, and 30 Ala12Ala carriers and performed detailed anthropometric and serological assessments.
Results: The participation rates in the GBR study were 31%, 44%, and 40%, and accordingly we included 12, 15, and 13 individuals with Pro12Pro, Pro12Ala, and Ala12Ala variants, respectively. There were no differences in anthropometric or metabolic variables among the different genotype groups.
Conclusions: Our report highlights that from a practical perspective, GBR can be used to study genotype–phenotype relationships. This approach can prove to be a valuable tool for follow-up findings from large-scale genetic discovery studies by undertaking detailed phenotyping procedures that might not be feasible in large studies. However, our study also illustrates the need for a larger pool of genotyped or sequenced individuals to allow for selection of rare variants with larger effects that can be examined in a GBR study of the present size.
Introduction
The rapid increase in the prevalence of obesity and associated attributes, such as insulin resistance, is one of the major causes of the global epidemic of type 2 diabetes and related complications such as cardiovascular diseases (Citation1). Genome-wide association studies (GWAS) have identified around 150 common gene variants related to obesity (Citation2) and over 100 gene variants associated with type 2 diabetes (Citation3), but their function remains largely elusive. Both genetic and environmental factors play an important role in the pathogenesis of metabolic diseases (Citation4,Citation5). Thus, it is imperative to understand whether the effect of a given variant of a genotype is conditional on context. The meticulous characterization of individuals carrying specific gene variants that have been implicated in the progression of insulin resistance and dyslipidemia can deliver knowledge of possible mechanisms which can further help in the diagnosis and management of metabolic diseases. In this perspective, genotype-based recall (GBR) studies can prove to be a powerful tool for comprehensive characterization of metabolic diseases (Citation6).
The GBR method offers a participant recruitment approach for studies of genotype–phenotype relationships. The term refers to the prospective recruitment and phenotyping of subgroups of research participants with specific genotypes from a larger background cohort where the genotypes of all participants are known. The main advantage of the GBR approach is that a detailed phenotypic characterization can be done efficiently in a subgroup of participants, which may help maximize statistical power. The GBR design may be used both for assessment of genetic effects and gene–treatment interactions (Citation6,Citation7). For example, when treatment efficiency in randomized control trials is heterogeneous, or the treatment is expensive or could have deleterious side effects in a subgroup of patients, identification of underlying genetic components sometimes may not be possible in such trials as they are not always designed specifically for this purpose. In such cases, GBR could be a useful tool, as it is designed to test specific hypotheses about gene–treatment interaction. It can also provide information about underlying mechanisms and treatment efficacy between different genotype groups, which further can be used to optimize therapies.
This approach has been suggested by many investigators, but relatively few dedicated studies have been performed to date prospectively collecting data based on known genotypes. We are aware of a few studies in the cardiovascular and metabolic field, including a study of the relationship between the negative regulation of atrial natriuretic peptides by microRNA-425 (Citation8), a study of functional effects of ADIPOQ gene variants (Citation9), and a study by Tang and co-workers providing the proof of concept for the feasibility of individualized treatment using a GBR approach (Citation10). A few previous studies of peroxisome proliferator-activated receptor gamma (PPARG) Pro12Ala (Citation6,Citation7,Citation11–13), which is also a gene of interest in the present study, have employed a GBR-like approach.
In the present study, we aimed to test the feasibility and practical implications of applying a GBR approach to increase the understanding of a genetic variant associated with metabolic disease. We pursued this aim by re-inviting participants from an existing cohort study based on their carrier status of the PPARG Pro12Ala (rs1801282) variant to a smaller in-depth clinical investigation.
Methods
The EpiHealth study
EpiHealth is a population-based multicenter longitudinal cohort study, which has been conducted in the Uppsala and Malmö regions of Sweden. The primary goal of the EpiHealth cohort is to provide a resource to study interactions between several genotypes and lifestyle factors in a large cohort derived from the Swedish population within the age range of 45–75 years regarding development of common degenerative disorders, such as cardiovascular diseases, cancer, dementia, joint pain, obstructive lung disease, depression, and osteoporotic fractures. The EpiHealth cohort aims to collect data on 300,000 individuals.
To date, 14,000 individuals have been enrolled at the Uppsala test center. The design has been described in detail elsewhere (Citation14); see also the study website (https://www.epihealth.se/) (Citation15). Briefly, EpiHealth includes self-assessment of lifestyle factors using an internet-based questionnaire; a visit to a test center where blood samples are collected and physiological parameters are recorded; and follow-up of disease incidence via nationwide medical registers. The present study focused on participants enrolled at the EpiHealth test center in Uppsala before 7 March 2013, when the data extraction was performed for selection of DNA samples for genotyping.
The ULSAM study
All men born between 1920 and 1924 in Uppsala, Sweden were invited to participate at age 50 in this longitudinal cohort study that was started in 1970. Participants were reinvestigated at the ages of 60, 71, 77, 82, and 88 years (Citation16). Further details about ULSAM can be obtained online (http://www.pubcare.uu.se/ulsam) (Citation17). Participants have undergone extensive phenotyping at repeated time points, including euglycemic clamps, oral glucose tolerance test (OGTT), DXA, echocardiography, 24 h ambulatory blood pressure measurement, and a range of biomarkers. For the present study, we used data on 922 non-diabetic participants who had been genotyped using genome-wide microarrays, and who underwent a euglycemic clamp at the age 71 examination. The total amount of glucose infused during a euglycemic clamp was taken as an index of sensitivity of subjects to an increasing concentration of plasma insulin. The glucose disposal rate (M) was then determined as the amount of glucose taken up during the last 60 min of the clamp. These data were used exclusively in the process of selecting a variant for the GBR study.
Ethical approval
All participants in the EpiHealth and ULSAM studies provided written informed consents. A renewed consent was obtained from the EpiHealth participants for the present substudy, and all study protocols were approved by the Regional Ethics Review Board in Uppsala. A copy of informed consent is available on request.
Genotyping
The 2500 most recently enrolled participants at the EpiHealth test center in Uppsala (before 7 March 2013) who had DNA available, with minimal adjustments of the sampling scheme to achieve an age and sex distribution reflecting the underlying distribution of EpiHealth, were selected for genotyping (Supplemental Table 1, available online) with the Illumina HumanCoreExome-12 v1.0 BeadChip including 522,731 autosomal markers. The genotype data were called using Illumina GenomeStudio 2011.1 GenCall followed by zCall version 3.3 (Citation18). Sample exclusion filters applied prior to the genotype calling with zCall were: (1) discordant sex information when comparing reported sex and sex determined by the X-chromosome; (2) outlying, non-European ancestry based on the first two components in a multidimensional scaling analysis (>3 standard deviations [SD] from the mean); (3) outlying heterozygosity rate (>5 SD from the mean based on markers with a minor allele frequency [MAF] < 1% or markers with MAF ≥1%); and (4) low sample call rate (<98%). Markers with a call rate <97%, a Fisher’s exact test p value for Hardy–Weinberg equilibrium <10−4, a cluster separation score <0.4, or a GenTrain score <0.6 were also excluded. After genotype calling with zCall, markers with a call rate <99% or a Fisher’s exact test p value for Hardy–Weinberg equilibrium <10−4 were also excluded. In total, 2432 samples passed the quality control, and 2378 samples remained after further exclusion of related individuals. All quality filters were applied using PLINK v.1.0.7 (Citation19). All participants in the ULSAM study provided blood samples for DNA analysis. DNA from the blood samples was extracted using a standard procedure, and genotyping was performed in the same fashion as described above.
The genotype-based recall study of PPARG Pro12Ala
Participant recruitment for a GBR substudy
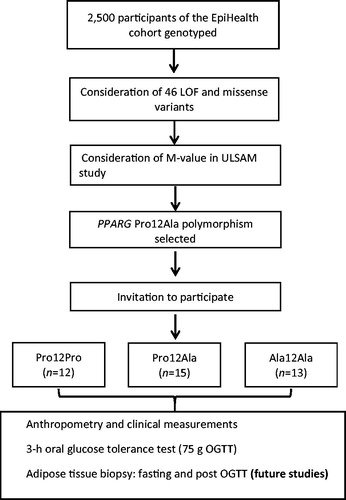
We considered loss of function (LOF) and missense variants that were directly genotyped on the Illumina HumanCoreExome microarray as the possible basis for our GBR study and decided to study carriers and non-carriers of the PPARG Ala12 allele at rs1801282 (see Results for rationale for choosing this variant). Potential participants who reported being on treatment with an anticoagulant or lipid-lowering agent, or having diabetes, were excluded from further consideration as this might have convoluted the assessment of genotype–phenotype associations. For each participant with the Ala12Ala genotypes, age- and sex-matched carriers of the Pro12 allele (heterozygotes and homozygotes) were selected. Age matching was based on a 2-year interval around the birth date. Two batches of invitations were sent out. Participant recruitment was stopped after a second round of invitation because of the limited resource allocation to perform the in-depth clinical investigation for a larger number of participants in this feasibility study. Participants who agreed to participate were contacted by a nurse before they arrived at the diabetes outpatient clinic at the Uppsala University Hospital, Uppsala, Sweden, where a detailed clinical examination was done, and study-specific exclusion criteria were critically examined (Supplemental data, available online). All study participants, physicians, nurses, and researchers, except the database manager, involved in the study remained blinded to participant genotypes until all 40 individuals had completed the investigation. A schematic overview of the study design is shown in .
Figure 1. Schematic flow chart describing the study design.
Clinical examination
Participants arrived in the morning after overnight fasting and were examined by a study nurse and physician. Anthropometric measurements were done. A 3-h OGTT with 75 g of glucose was performed, and blood samples were collected at baseline, 15, 30, 60, 90, 120, and 180 min. Subcutaneous adipose tissue was obtained by needle biopsies at baseline and after the OGTT, to be used for future studies.
Anthropometric and biochemical measurements
Participants underwent a detailed investigation of their medical history and a physical examination, including height (cm), weight (kg), waist (cm) and hip (cm) circumference, which was carried out according to the World Health Organization guidelines. Body composition was determined by bioelectrical impedance (Kroppsanalysator BC-418MA, Tania). Body mass index (BMI) was calculated as weight (kg) divided by height (m2). A 3-h OGTT (75 g oral glucose load) was performed, and blood samples for analysis of glucose, insulin, non-esterified fatty acids (NEFA), and glycerol were drawn at baseline, 15, 30, 60, 90, 120, and 180 min. NEFA was measured using NEFA fluorometric assay kit (Cayman Chemicals, Anna Arbor, MI, USA), and glycerol was quantified using free glycerol reagent (Sigma Chemicals Co., St Louis, MO, USA). In addition, the fasting baseline blood sample was used for the analysis of lipid profile (triglyceride, total cholesterol, high-density cholesterol [HDL], and low-density lipoprotein [LDL]). Blood plasma and serum samples were analyzed at the Department of Clinical Chemistry, Uppsala University Hospital. Fasting glucose and insulin values were used to calculate a homeostatic model assessment of insulin resistance (HOMA-IR), which was used as an estimate of insulin resistance. In addition, glucose, insulin, and NEFA levels obtained during OGTT were used to calculate insulin sensitivity indices (ISI), which were determined by the Matsuda Index, and a revised quantitative insulin sensitivity check (QUICKI) (Citation20–22). Plasma glucose at 2 h post-glucose load was used as an additional glucose assessment. The following formulas were used to calculate different insulin sensitivity indices. Revised QUICKI = 1/(log fasting glucose + log fasting insulin + log fasting NEFA); Matsuda Index = 10,000/√fasting glucose × fasting insulin × mean glucose × mean insulin.
Confirmatory genotyping
The fasting venous blood sample was collected in an EDTA tube and stored at −20 °C until DNA extraction using NucleoSpin Blood extraction kit (Macherey-Nagel GmbH & Co. KG, Düren, Germany). De novo genotyping of PPARG Pro12Ala polymorphism (rs1801282) was done to confirm the previous genotyping using single base primer extension with detection of the incorporated allele by fluorescent polarization template dye incorporation (Citation23). Signal intensities were read using a Tecan Genios Pro fluorescence absorbance reader, and raw data from the fluorescence polarization were converted to genotype data using AlleleCaller 4.0.0.1.
Statistical analysis
GBR variant selection
We used the ULSAM and EpiHealth cohorts for variant selection. Fasting plasma glucose and triglycerides were natural log transformed to promote normality. All metabolic phenotypes were z-score transformed (mean = 0 and SD =1) prior to the association tests to allow for comparisons of effect sizes across measures. Analyses were restricted to non-diabetic individuals (based on fasting plasma glucose and diabetes medication), and the association tests for all phenotypes measured in plasma were further restricted to individuals fasting for at least 6 h. We excluded first- or second-degree relatives and individuals of non-European ancestry from further analyses. The associations of each variant (independent variable) with each phenotype (dependent variable) were tested in linear regression models adjusting for age, sex (EpiHealth), and the first three (EpiHealth) or two (ULSAM) ancestry components (from multidimensional scaling analyses) assuming additive effects.
Analyses of clinical measures in the GBR study
In order to promote normal distribution, fasting glucose, fasting insulin, HOMA-IR, area under the curve (AUC) glucose, AUC insulin, triglycerides, AUC NEFA, AUC glycerol (0–120 min), and NEFA and glycerol at baseline were natural log transformed. Associations between a number of minor alleles and phenotypes were tested under an additive model using linear regressions adjusted for age, sex, and ancestry components. All statistical analyses were performed in STATA 13.1 (StataCorp LP, College Station, TX, USA) or R 3.0.0 (Citation24).
Results
Selection of a gene variant as basis for the GBR study
In order to select the gene variant for our GBR study, we considered LOF and missense variants that were directly genotyped on the Illumina HumanCoreExome microarray. For this pilot study, we considered variants across the whole genome for which at least 10 participants were homozygous for the minor allele. According to these criteria, we identified 46 gene variants to be further considered (). We assessed associations of these 46 variants with clinical parameters in the EpiHealth cohort (n = 2500) and with M-values from clamp studies in ULSAM (n = 922) (Supplemental Table 2, available online). Among these variants, we opted to invite carriers and non-carriers of the PPARG Pro12Ala (rs1801282) to this pilot study, as it is a relatively common variant (MAF = 0.14) in the EpiHealth cohort, allowing inclusion of our projected number of 15 participants in each genotype group. Also, functionally, the Ala12 variant has a decreased transactivation capacity, and its association with cardiometabolic traits is well known. In addition, between the two significantly associated variants, Ala12 was strongly associated with the glucose disposal rate (M-value) as recorded in the euglycemic clamps of the ULSAM study (β = 0.19, p = 0.0032).
Table 1. Protein-altering variants in the EpiHealth cohort with ≥10 participants homozygous for minor allelesTable Footnotea.
Participation rate in the GBR study
In total, we invited 39, 34, and 30 people with 0, 1, and 2 copies of the minor Ala12 allele, respectively. After sending two rounds of invitations, we included 12 Pro12Pro, 15 Pro12Ala, and 13 Ala12Ala participants with a response rate of 31%, 44%, and 40%, respectively (total n = 40), in this GBR study.
Clinical characteristics of carriers of PPARG Pro12Ala
As should be expected from the modest sample size, we did not observe any significant differences in the anthropometric and clinical characteristics across the three different genotype groups (). Participants with the PPARG Pro12Ala variant were studied with an OGTT. The area under the curve for glucose, insulin, NEFA, and glycerol did not differ between the variant groups (). Likewise, the insulin sensitivity determined by calculating Matsuda Index, and revised QUICKI did not differ between the three groups of carriers ().
Table 2. Baseline characteristics of participants in the GBR study as genotyped by their carrier status of PPARG Pro12Alaa.
Table 3. Metabolic variables of participants in the GBR study as genotyped by their carrier status of PPARG Pro12AlaTable Footnotea.
Discussion
GWAS have made significant progress over the past decade in the discovery of genetic variants associated with disease risk (Citation25). In the field of cardiovascular disease and type 2 diabetes research, hundreds of loci associated with these traits have been discovered. However, the functional impact of these gene variants on the phenotype needs more detailed follow-up studies using measurements that may not be feasible to undertake in large study populations where variants have been discovered. Here, we have performed a pilot study addressing the feasibility and practical implications of using a genotype-based recall approach to perform functional characterization of genetic variants established in GWAS. We genotyped 2500 individuals from the EpiHealth cohort, Uppsala, and identified 46 potential candidate variants based on a priori criteria. We decided to base our GBR study on the PPARG Pro12Ala polymorphism due to its relatively high minor allele frequency, its strong association with influenced glucose disposal rates in the ULSAM study, and its well-characterized role in the context of obesity, insulin resistance, and type 2 diabetes (Citation26–29). As the genotypes of EpiHealth participants were already known, it was possible to invite a smaller number of individuals with the desired genotype distribution. This is an advantage of the GBR approach over conventional recruitment paradigms, where genotypes are randomly assigned, as GBR can be more powerful, particularly when rarer variants are of interest and when oversampling homozygotes of the rare variant. Another advantage of the GBR design is that you can match invitees on their baseline characteristics (in this case, age and sex) before inviting them to the substudy. This can be important to avoid confounding results, especially if factors other than the polymorphism itself are known to influence the phenotype. Similarly, in another GBR study of PPARG Pro12Ala by Stefan et al. (Citation6), the participants were matched for age, sex, BMI, and waist–hip ratio, since the authors were evaluating the effect of free fatty acids (FFA) on insulin secretion and sensitivity and the above factors are known to influence these variables.
Besides its potential, the GBR approach also has considerable ethical challenges (Citation30), mainly the potential disclosing of the genotype to the participants. For the present study, we considered ethical recommendations specific to GBR studies (Citation31), in addition to the viewpoint of the local ethics committee at the Uppsala University. In brief, participants were informed about the reason and nature of this follow-up substudy in the invitation letter. However, although the participants were informed that they were being recruited to a genetic study, their Pro12Ala genotype was not disclosed to them. It would not have been easy to provide this information at the test center given that the staff was blinded to the genotype of the participant; but following the ethical principle of autonomy, if the participant had wanted to know their genotype, this would have been disclosed (by breaking the code). However, this did not occur in any case. We acknowledge that knowledge about the Pro12Ala variant being investigated in the present study may not be the most informative for a research participant, given the very small variance explained. Hence, the ethical considerations with regard to this specific study were very different to those of a study of more predictive variants, such as for example rare BRCA variants, where this information is of much higher importance. Thus the non-disclosure of carriership would be harder to justify, and the consequences of this knowledge for the participant and relatives would be much greater.
The participation rate is another important aspect of a GBR study, especially when studying rarer variants than Pro12Ala, and when the number of genotyped or sequenced individuals to recruit from is limited. As shown in our study, among the total number of participants genotyped (n = 2500), we had 561 Pro12Ala and 31 Ala12Ala carriers (). Based on our previous experience of a moderate participation rate (about 50%), which we almost reached, we realistically expected to recruit about 15 participants being homozygous for the Ala allele. This is also a reason for why we chose to recruit heterozygote carriers, even if the statistical power would be higher by contrasting only homozygotes. This highlights one of the challenges with GBR studies. Even for a relatively common variant as Pro12Ala (MAF = 14%) and a relatively large sampling frame (n = 2500), we did not have enough homozygotes to perform a study of only homozygotes. Indeed, among the uncommon and rare variants (MAF < 0.05) where the effects could be expected to be larger, we had five or fewer individuals homozygous for the risk allele (data not shown), making them unfeasible as basis for recruitment to a GBR. Hence, for future GBR efforts, a considerably larger cohort, at least 10-fold larger, and preferably sequenced to pick up all rare variants in the target gene, would be ideal.
PPARG is a nuclear transcription factor known to regulate adipogenesis and expression of genes involved in adipose tissue glucose and lipid metabolism (Citation32). Two PPARG isoforms, PPARG 1 and PPARG 2, exist due to mRNA splicing. The difference between these two isoforms is that the latter isoform has an additional stretch of 28 amino acids towards its N-terminal (Citation33). The expression of PPARG 1 is ubiquitous, whereas PPARG 2 is selectively expressed in adipose tissue (Citation34). As a result of the amino acid substitution of proline to alanine, a common Pro12Ala polymorphism was identified at the 12th codon in exon B of PPARG (Citation35). The frequency of the Ala variant varies from 4% to 28%, with higher prevalence among people of northern European ancestry (Citation36). Previous studies have shown an association of the PPARG Pro12Ala polymorphism with improved insulin sensitivity and reduced risk of type 2 diabetes (Citation26–28), but, in contrast, higher BMI (Citation37,Citation38). In addition to glucose metabolism, some studies have also indicated an association of the Pro12Ala polymorphism with lipid metabolism. Even if such associations have been inconsistent across different studies (Citation39–44), there is evidence in our work in the Global Lipids Genetics Consortium (Citation45) that there are associations of variation in PPARG with lipid fractions, presumably due to high correlations between insulin resistance and dyslipidemia. In line with our findings, a few prior studies characterizing the role of the PPARG Pro12Ala polymorphism using a GBR approach also failed to report any significant differences between the Pro12Pro and Pro12Ala participants for the phenotypes they studied, presumably due to a similarly low statistical power in these previous studies, as in ours. Specifically, a study by Stefan et al. (Citation6) could not show any differences in insulin sensitivity in response to the FFA between the Pro12Pro and Pro12Ala groups, which they suggested could be due to the small number of participants included (10 Pro12Pro and 10 Pro12Ala). Another study by Pihlajamäki et al. (Citation13) investigating the relation between the PPARG Pro12Ala and polyunsaturated fatty acids (PUFA) and its influence on serum lipid profile also employed the GBR approach. They were unable to find any effect of PUFA on the PPARG gene, which also remained unaffected by the diet–genotype interaction. Moreover, they could not find any differences in serum lipid profile and glucose between Pro12Pro and Ala12Ala individuals, and suggested that this was due to a small sample size. In line with these observations and as expected based on known effect sizes, we were unable to report any association of Pro12Ala polymorphism with measures of adiposity or insulin sensitivity.
In summary, we have performed a pilot study to test the feasibility and practical implications of applying a GBR approach to increase the understanding of cardiometabolic diseases. We did this by performing a comprehensive clinical study of individuals recruited based on their carriership for the PPARG Pro12Ala polymorphism. Due to the modest sample size, we observed no differences in anthropometric and metabolic parameters between the three groups of carriers. Our study suggests that the GBR approach is straightforward, and can be used as a tool to disentangle the nature of genotype–phenotype relationships. This approach can prove to be a valuable means to carry out a follow-up of studies where detailed, precise phenotyping in a large population is costly and time-consuming. Our study highlights several practical aspects of a GBR study, and that the approach is feasible. Specifically, no ethical concerns were raised by either the ethics committee or any participants. Further, the participation rate was unexpectedly high even though the study protocol was extensive in comparison to the less extensive prescreening in EpiHealth. Also, in spite of such an extensive study protocol, an in-depth examination was possible at a quite affordable cost in relation to examining all study individuals. Finally, the genotypes of all participants were confirmed in the recall phase. A different outcome of any of these four practical aspects could have falsified our hypothesis that GBR would be a workable strategy.
Our study also illustrates that a considerably larger number of genotyped—or preferably sequenced—individuals would be optimal as a basis for a more efficient GBR study. With a larger pool of genotyped samples, it would be possible to select more rare variants with larger effects, which would make the GBR more efficient and increase statistical power. Ultimately, it would be even better if the blood samples were subjected to DNA sequencing—either using whole-genome, exome, or targeted resequencing of the gene of interest—as that would also allow identification of carriers of rare or private mutations, which could be assumed to have even larger effects. Such rare or private mutations could then be grouped based on their inferred effects, for example, loss-of-function variant carriers could be studied in comparison with non-carriers. Given the limited number of samples (n = 2500) we had the resources to genotype in the present study, it was not feasible to consider rarer variants, and based on power calculations it was not surprising that we were not able to observe any significant effects of the chosen common variant with modest, or even minute, effects on the clinical phenotype.
Notes on contributors
Prasad G. Kamble, MSc, is a PhD student at Uppsala University, Uppsala, Sweden.
Stefan Gustafsson, PhD, is a post-doctoral researcher at Uppsala University, Uppsala, Sweden.
Maria J. Pereira, PhD, is a post-doctoral researcher at Uppsala University, Uppsala, Sweden.
Per Lundkvist, MD, is a PhD student at Uppsala University, Uppsala, Sweden.
Naomi Cook, PhD, is a post-doctoral researcher at Uppsala University, Uppsala, Sweden.
Lars Lind, MD, PhD, is a Professor at Uppsala University, Uppsala, Sweden.
Paul W. Franks, PhD, is a Professor at Lund University, Malmö, Sweden.
Tove Fall, VMD, PhD, is an Associate Professor at Uppsala University, Uppsala, Sweden.
Jan W. Eriksson, MD, PhD, is a Professor at Uppsala University, Uppsala, Sweden.
Erik Ingelsson, MD, PhD, FAHA, is a Professor at Stanford University, Stanford, CA, USA.
Supplemental data
Download MS Word (81.7 KB)Acknowledgements
We gratefully thank the participants of the EpiHealth study for their important contributions and acknowledge the valuable technical, administrative, and analytical contributions and expert advice from co-workers at Uppsala University Hospital.
Disclosure statement
E.I. is a scientific advisor for Precision Wellness and Olink Proteomics for work unrelated to this article.
Additional information
Funding
Related Research Data
References
- Hu FB. Globalization of diabetes: the role of diet, lifestyle, and genes. Diabetes Care. 2011;34:1249–57.
- Shungin D, Winkler TW, Croteau-Chonka DC, Ferreira T, Locke AE, Mägi R, et al. New genetic loci link adipose and insulin biology to body fat distribution. Nature. 2015;518:187–96.
- Prasad RB, Groop L. Genetics of type 2 diabetes-pitfalls and possibilities. Genes (Basel). 2015;6:87–123.
- Hinton RB. Genetic and environmental factors contributing to cardiovascular malformation: a unified approach to risk. J Am Heart Assoc. 2013;2:e000292.
- Speakman JR. Obesity: the integrated roles of environment and genetics. J Nutr. 2004;134:2090S–105S.
- Stefan N, Fritsche A, Haring H, Stumvoll M. Effect of experimental elevation of free fatty acids on insulin secretion and insulin sensitivity in healthy carriers of the Pro12Ala polymorphism of the peroxisome proliferator–activated receptor-gamma2 gene. Diabetes. 2001;50:1143–8.
- Tan GD, Neville MJ, Liverani E, Humphreys SM, Currie JM, Dennis L, et al. The in vivo effects of the Pro12Ala PPARgamma2 polymorphism on adipose tissue NEFA metabolism: the first use of the Oxford Biobank. Diabetologia. 2006;49:158–68.
- Arora P, Wu C, Khan AM, Bloch DB, Davis-Dusenbery BN, Ghorbani A, et al. Atrial natriuretic peptide is negatively regulated by microRNA-425. J Clin Invest. 2013;123:3378–82.
- Lee BP, Lloyd-Laney HO, Locke JM, McCulloch LJ, Knight B, Yaghootkar H, et al. Functional characterisation of ADIPOQ variants using individuals recruited by genotype. Mol Cell Endocrinol. 2016;428:49–57.
- Tang Y, Axelsson AS, Spegel P, Andersson LE, Mulder H, Groop LC, et al. Genotype-based treatment of type 2 diabetes with an alpha2A-adrenergic receptor antagonist. Sci Transl Med. 2014;6:257ra139.
- Herrmann J, Rubin D, Hasler R, Helwig U, Pfeuffer M, Auinger A, et al. Isomer-specific effects of CLA on gene expression in human adipose tissue depending on PPARgamma2 P12A polymorphism: a double blind, randomized, controlled cross-over study. Lipids Health Dis. 2009;8:35.
- Rubin D, Herrmann J, Much D, Pfeuffer M, Laue C, Winkler P, et al. Influence of different CLA isomers on insulin resistance and adipocytokines in pre-diabetic, middle-aged men with PPARgamma2 Pro12Ala polymorphism. Genes Nutr. 2012;7:499–509.
- Pihlajamaki J, Schwab U, Kaminska D, Ågren J, Kuusisto J, Kolehmainen M, et al. Dietary polyunsaturated fatty acids and the Pro12Ala polymorphisms of PPARG regulate serum lipids through divergent pathways: a randomized crossover clinical trial. Genes Nutr. 2015;10:43.
- Lind L, Elmstahl S, Bergman E, Englund M, Lindberg E, Michaelsson K, et al. EpiHealth: a large population-based cohort study for investigation of gene-lifestyle interactions in the pathogenesis of common diseases. Eur J Epidemiol. 2013;28:189–97.
- EpiHealth. Epihealth 2011. Available from: https://www.epihealth.se
- Berglund L, Berne C, Svardsudd K, Garmo H, Melhus H, Zethelius B. Seasonal variations of insulin sensitivity from a euglycemic insulin clamp in elderly men. Ups J Med Sci. 2012;117:35–40.
- ULSAM. Uppsala Longitudinal Study of Adult Men 2017. Available from: http://www.pubcare.uu.se/ulsam
- Goldstein JI, Crenshaw A, Carey J, Grant GB, Maguire J, Fromer M, et al. zCall: a rare variant caller for array-based genotyping: genetics and population analysis. Bioinformatics. 2012;28:2543–5.
- Purcell S, Neale B, Todd-Brown K, Thomas L, Ferreira MA, Bender D, et al. PLINK: a tool set for whole-genome association and population-based linkage analyses. Am J Hum Genet. 2007;81:559–75.
- Perseghin G, Caumo A, Caloni M, Testolin G, Luzi L. Incorporation of the fasting plasma FFA concentration into QUICKI improves its association with insulin sensitivity in nonobese individuals. J Clin Endocrinol Metab. 2001;86:4776–81.
- Katz A, Nambi SS, Mather K, Baron AD, Follmann DA, Sullivan G, et al. Quantitative insulin sensitivity check index: a simple, accurate method for assessing insulin sensitivity in humans. J Clin Endocrinol Metab. 2000;85:2402–10.
- Matsuda M, DeFronzo RA. Insulin sensitivity indices obtained from oral glucose tolerance testing: comparison with the euglycemic insulin clamp. Diabetes Care. 1999;22:1462–70.
- Chen X, Levine L, Kwok PY. Fluorescence polarization in homogeneous nucleic acid analysis. Genome Res. 1999;9:492–8.
- Team RC. R: A language and environment for statistical computing. 2017. Available from: https://www.r-project.org/
- Stranger BE, Stahl EA, Raj T. Progress and promise of genome-wide association studies for human complex trait genetics. Genetics. 2011;187:367–83.
- Stumvoll M, Wahl HG, Loblein K, Becker R, Machicao F, Jacob S, et al. Pro12Ala polymorphism in the peroxisome proliferator-activated receptor-gamma2 gene is associated with increased antilipolytic insulin sensitivity. Diabetes. 2001;50:876–81.
- Jacob S, Stumvoll M, Becker R, Koch M, Nielsen M, Löblein K, et al. The PPARgamma2 polymorphism pro12Ala is associated with better insulin sensitivity in the offspring of type 2 diabetic patients. Horm Metab Res. 2000;32:413–16.
- Altshuler D, Hirschhorn JN, Klannemark M, Lindgren CM, Vohl MC, Nemesh J, et al. The common PPARgamma Pro12Ala polymorphism is associated with decreased risk of type 2 diabetes. Nat Genet. 2000;26:76–80.
- Meirhaeghe A, Fajas L, Helbecque N, Cottel D, Auwerx J, Deeb SS, et al. Impact of the Peroxisome Proliferator Activated Receptor gamma2 Pro12Ala polymorphism on adiposity, lipids and non-insulin-dependent diabetes mellitus. Int J Obes Relat Metab Disord. 2000;24:195–9.
- Beskow LM, Linney KN, Radtke RA, Heinzen EL, Goldstein DB. Ethical challenges in genotype-driven research recruitment. Genome Res. 2010;20:705–9.
- Beskow LM, Fullerton SM, Namey EE, Nelson DK, Davis AM, Wilfond BS. Recommendations for ethical approaches to genotype-driven research recruitment. Hum Genet. 2012;131:1423–31.
- Ahmadian M, Suh JM, Hah N, Liddle C, Atkins AR, Downes M, et al. PPARgamma signaling and metabolism: the good, the bad and the future. Nat Med. 2013;19:557–66.
- Yanase T, Yashiro T, Takitani K, Kato S, Taniguchi S, Takayanagi R, et al. Differential expression of PPAR gamma1 and gamma2 isoforms in human adipose tissue. Biochem Biophys Res Commun. 1997;233:320–4.
- Fajas L, Auboeuf D, Raspe E, Schoonjans K, Lefebvre AM, Saladin R, et al. The organization, promoter analysis, and expression of the human PPARgamma gene. J Biol Chem. 1997;272:18779–89.
- Yen CJ, Beamer BA, Negri C, Silver K, Brown KA, Yarnall DP, et al. Molecular scanning of the human peroxisome proliferator activated receptor gamma (hPPAR gamma) gene in diabetic Caucasians: identification of a Pro12Ala PPAR gamma 2 missense mutation. Biochem Biophys Res Commun. 1997;241:270–4.
- Tonjes A, Scholz M, Loeffler M, Stumvoll M. Association of Pro12Ala polymorphism in peroxisome proliferator-activated receptor gamma with pre-diabetic phenotypes: meta-analysis of 57 studies on nondiabetic individuals. Diabetes Care. 2006;29:2489–97.
- Mansoori A, Amini M, Kolahdooz F, Seyedrezazadeh E. Obesity and Pro12Ala polymorphism of peroxisome proliferator-activated receptor-gamma gene in healthy adults: a systematic review and meta-analysis. Ann Nutr Metab. 2015;67:104–18.
- Galbete C, Toledo E, Martinez-Gonzalez MA, Martínez JA, Guillén-Grima F, Marti A. Pro12Ala variant of the PPARG2 gene increases body mass index: An updated meta-analysis encompassing 49,092 subjects. Obesity (Silver Spring). 2013;21:1486–95.
- Barbieri M, Rizzo MR, Papa M, Acampora R, De Angelis L, Olivieri F, et al. Role of interaction between variants in the PPARG and interleukin-6 genes on obesity related metabolic risk factors. Exp Gerontol. 2005;40:599–604.
- Swarbrick MM, Chapman CM, McQuillan BM, Hung J, Thompson PL, Beilby JP. A Pro12Ala polymorphism in the human peroxisome proliferator-activated receptor-gamma 2 is associated with combined hyperlipidaemia in obesity. Eur J Endocrinol. 2001;144:277–82.
- Iwata E, Matsuda H, Fukuda T, Fukuen S, Motomura T, Igarashi T, et al. Mutations of the peroxisome proliferator-activated receptor gamma (PPAR gamma) gene in a Japanese population: the Pro12Ala mutation in PPAR gamma 2 is associated with lower concentrations of serum total and non-HDL cholesterol. Diabetologia. 2001;44:1354–5.
- Hamada T, Kotani K, Tsuzaki K, Sano Y, Murata T, Tabata M, et al. Association of Pro12Ala polymorphism in the peroxisome proliferator-activated receptor gamma2 gene with small dense low-density lipoprotein in the general population. Metabolism. 2007;56:1345–9.
- Tai ES, Corella D, Deurenberg-Yap M, Adiconis X, Chew SK, Tan CE, et al. Differential effects of the C1431T and Pro12Ala PPARgamma gene variants on plasma lipids and diabetes risk in an Asian population. J Lipid Res. 2004;45:674–85.
- Robitaille J, Despres JP, Perusse L, Vohl MC. The PPAR-gamma P12A polymorphism modulates the relationship between dietary fat intake and components of the metabolic syndrome: results from the Quebec Family Study. Clin Genet. 2003;63:109–16.
- Willer CJ, Schmidt EM, Sengupta S, Peloso GM, Gustafsson S, et al.; Global Lipids Genetics Consortium. Discovery and refinement of loci associated with lipid levels. Nat Genet. 2013;45:1274–83.