Abstract
Reliable survey methods for detection are critically important for the monitoring and management of exotic species. The eastern rosella (Platycercus eximius), a broad-tailed parakeet endemic to southeastern Australia, was introduced to New Zealand a century ago and is now geographically widespread. We studied the necessary timeframe for surveying the eastern rosella within its introduced range, testing the hypothesis that there are seasonal differences in the likelihood of detection. Although our comparisons were limited to surveys conducted during a single year, they are suggestive of an important impact of season on the survey duration required to detect eastern rosella confidently. Median latency until first detection was less during summer months (2.55 min) in comparison with winter months (11.2 min). Furthermore, 90% of first detections occurred within the first 13 min in summer surveys, compared with 22 min in winter. These results have implications for the design of surveys aiming to monitor rosella populations in New Zealand, and reiterate the importance of tailoring survey methods to the species of interest.
Introduction
Surveying animal populations is a fundamental component of ecology, both at a theoretical and practical level. Information on occupancy and abundance is vital for describing species distributions and community assemblage patterns (Noon Citation1981; Taylor Citation1984), identifying and analysing the factors influencing these patterns (Fernández-Juricic Citation2000; Perault & Lomolino Citation2000), assessing and predicting the impact of anthropogenic activities and stressors (Griffiths & van Schaik Citation1993; Bibby Citation1999), and planning the conservation management of populations (McLennan & McCann Citation2002; Witmer Citation2005; Bart Citation2009). Diverse methods have been developed to survey avian populations specifically (Bibby et al. Citation2000). These techniques can be broadly divided into three categories: counts of singing/displaying individuals or breeding pairs within a defined area; transect counts of all individuals along a specified distance; and counts undertaken at fixed points, known as point counts or circular-plot surveys, which record all individuals within a stated distance (Dawson & Bull Citation1975; Reynolds et al. Citation1980; Bibby & Buckland Citation1987). Each method requires a different amount of effort, has advantages and disadvantages, and varies in its accuracy and costs (Reynolds et al. Citation1980).
In New Zealand, 5-min point counts are widely used in avian surveys (Dawson & Bull Citation1975; Spurr Citation2005); however, few studies have evaluated the suitability of this method for particular New Zealand species (Gill Citation1980; Cassey et al. Citation2007). In most cases, allowances are not made for diversity in the ecology and behaviour of different target taxa. Most survey methods, including standard 5-min point counts, assume that all birds have the same probability of being detected (Reynolds et al. Citation1980), an assumption that is generally false (Boulinier et al. Citation1998). Even from brief observations, it is evident that conspicuousness varies considerably among different bird species and even between the sexes of the same species. Accordingly, the timeframe required to detect an individual of a particular species, if it is present, will also vary. Another key problem associated with accurately surveying animal populations in general is that most methods do not detect all individuals that are actually present in an area (Wobeser Citation2007). The probability of detecting an animal, or its detectability, is therefore usually <1 (Wobeser Citation2007).
The problems associated with low detection probability may be reduced in a number of ways. Additional observers can be used to count animals simultaneously, enabling a probability of detection to be estimated (Nichols et al. Citation2000; Thompson Citation2002), and/or replicate counts may be carried out (Verner & Milne Citation1989; Wobeser Citation2007). Regarding survey design, longer survey duration plays a crucial role in increasing the probability of detection, particularly for species that are inconspicuous and/or have large home ranges (Johnson Citation1995). By establishing the length of time necessary to detect a species if it is present, the accuracy and efficiency of surveys can be increased (Petit et al. Citation1995; Lee & Marsden Citation2008). Additionally, distance sampling methods can be added to point-count protocols, generating an estimate of density, and can reduce detectability problems in many cases (Buckland et al. Citation2001; Thompson Citation2002), although these are not evaluated here.
The eastern rosella (Platycercus eximius), an introduced broad-tailed parakeet, has been established in the wild in New Zealand for a century (Higgins Citation1999). While their spread has been documented through analysis of Ornithogical Society of New Zealand (OSNZ) observer records (Wright Citation2001), no published studies have monitored changes in eastern rosella abundance at specific New Zealand locations. There are a number of species-typical behavioural barriers to reliably detecting eastern rosella (hereafter rosella) across the New Zealand landscape. Superficially, it may seem that rosella have a high probability of detection, being both visually and vocally conspicuous to the human observer (Berg and Bennett Citation2010). When silent and stationary in foliage, however, these birds can be extremely difficult to detect (Galbraith Citation2010); other studies have also noted the difficulty of detecting seemingly conspicuous parrots in dense vegetation (Heinsohn et al. Citation2005; Rivera-Milan et al. Citation2005). Furthermore, rosella are particularly wary in the presence of humans (Higgins Citation1999; Blumstein Citation2003), and could potentially avoid observers during surveys. Finally, rosella home ranges can be much larger than a survey area, with singular flights between perch sites spanning distances of more than 0.5 km recorded for the species in New Zealand (Galbraith Citation2010). These behavioural factors suggest rosella do not, in fact, have a high probability of detection during standard surveys.
To evaluate and customise survey design to an invasive New Zealand bird species, we analysed the necessary timeframe for surveying the eastern rosella. The study focused on the effect of survey duration on detection frequency (the percentage of counts where the species was detected; Verner & Milne Citation1989); we also briefly looked at its role in generating relative abundance estimates. Additionally, we provide a preliminary test of the hypothesis that the probability of detecting rosella, and therefore the optimal survey time, varies between seasons. Other factors, suggested in the avian survey technique literature that may also affect species detectability, including time of day, location and relative abundance, were evaluated simultaneously. The reliability of detection estimates was also examined through a series of parallel data collection sessions with additional observers.
Methods
Study sites
Bird surveys were conducted at nine sites in the Auckland region, North Island, New Zealand (): Motuihe (36°48′S, 174°56′E), Tiritiri Matangi Island (36°36′S, 174°53′E), Shakespear Regional Park (36°36′S, 174°49′E), Tawharanui Regional Park (36°22′S, 174°50′E), Wenderholm Regional Park (36°32′S, 174° 42′E), Waitakere (Ark in the Park) (36°53′S, 174°31′E), Blockhouse Bay (36°55′S, 174°42′E), Chelsea Estate Heritage Park (36°49′S, 174°43′E) and the Auckland Domain (36° 51′S, 174°46′E). Rosella were known, from personal records and encounters, to be present at all sites prior to commencing the surveys. The sites included a variety of habitat types with differing mammalian predator control regimes (). The suburban site in Blockhouse Bay was surveyed in winter only, and was substituted for Chelsea Estate Heritage Park in the summer surveys for ease of access and to reduce travel time; both are suburban areas with semi-native coastal forest, bordering on harbours, without pest control. Shakespear Regional Park was not one of the initial winter survey sites, but was included in the summer surveys as it was of interest for a separate component of our broader research, given the recent sightings of an endemic parakeet, the red-crowned kakariki (Cyanoramphus novaezelandiae) in this park (JAG, pers. obs.). Shakespear is one of the only known mainland New Zealand sites where the self-reintroduction of kakariki, from the nearby mammal-free Tiritiri Matangi Island (Ortiz-Catedral & Brunton Citation2009), in the presence of the exotic rosella, may be occurring.
Figure 1 Map of the Auckland region, New Zealand, showing point-count survey sites.
Table 1 Description of study sites in the Auckland region, New Zealand, where 30-min point-count surveys were conducted.
Sampling regime
A series of point-count surveys were conducted during the austral winter months, between April and August 2008 (by EAF), and over the following summer months, between December 2008 and March 2009 (by JAG). Surveys were undertaken in the morning, from 07:00 h, and completed no later than 14:00 h local time. Timing was limited by the ability of observers to access the two island sites. Surveying during particularly wet or windy weather was avoided. A single survey consisted of a 30-min point count, divided into 5-min periods. All birds (of any species) seen or heard within a 150-m radius were recorded for each 5-min period (adapted from Dawson & Bull Citation1975). Birds that remained within the survey area, and were either sighted continuously or called consistently from the same location, were taken to be the same individual. For our analyses of rosella specific surveys, we recorded the latency of the species’ initial detection.
Within each site, surveys were carried out at six different sub-locations. These were randomly selected using a bi-coordinate grid. Points close to streams, where noise levels were sufficient to interfere with auditory detection of birds, were excluded (Simons et al. Citation2009). To enable the surveys to be relatively independent, but completed within the designated time frame, locations were chosen that were greater than 300 m apart and within 30 min walking distance of each other. Counts at each study site were repeated three times on different days, both in summer and winter. Where possible, the order of bird counts at the six locations was varied on each repeat. In total, 143 counts of 30 min duration were conducted over the summer months (one count was not done for the first Tiritiri Matangi Island survey day because of time restrictions), and 126 counts over the preceding winter.
As two different observers conducted the surveys independently for each season, there is a potential inter-observer bias. To examine this effect, a second independent observer was present for 16 of the summer surveys, simultaneously recording rosella detection time and relative abundance, and richness of other avian species. Three different people assisted as the secondary observer; all were trained biologists familiar with New Zealand avifauna, and had a good level of skill in identification prior to commencement of the surveys. These data were then used to estimate observer bias through the measure of repeatability; with the explicit assumption that should the surveys show a high degree of consistency any differences found between summer and winter surveys would not be solely attributable to the effect of inter-observer bias.
Statistical analyses
Repeatability (r), a measure of intraclass correlation, was calculated using ANOVAs of primary and secondary observer data and formulae given in Lessels & Boag (1987) (see below), where s 2 A is the variance component among groups and s 2 is the variance component within groups, calculated from the mean squares in the analysis of variance (MS W=mean squares within groups and MS A=mean squares among groups). Repeatability essentially describes the percentage of variance that is attributable to intergroup variation rather than within-group variation (e.g. Igic et al. Citation2010); ‘groups’ in this case are the different observers.
We used a Pearson's chi-square test to determine whether the proportion of surveys detecting rosella differed between seasons, and subsequently tested whether detection latency differed between summer and winter surveys using a Wilcoxon rank-sum test. We then investigated what factors best predicted detection latency by performing a generalised linear mixed model (GLMM) analysis, selecting important explanatory variables based on a stepwise backward regression. The variables of date, time, site type, location, season, relative rosella abundance, and season×relative rosella abundance interaction, were entered in the full model. Other interactions were not included as there were insufficient degrees of freedom. Factors were tested as whole effects to preserve the degrees of freedom. The least significant terms were removed sequentially from the full model, with the ‘probability to leave’ criterion set at 10%, until only significant variables remained. A final GLMM was fitted using the three most significant predictors of detection latency: season, relative rosella abundance, and their interaction term, with site included as a random effect.
Pearson's chi-square tests were used to determine whether the overall probabilities of identifying rosella at the time of first detection by auditory and by visual means were equal, and also whether the observed ratio of auditory to visual detection differed between sites.
Finally, the asymptote of detection, or the point beyond which an increase in survey time will not result in an increase in the proportion of surveys that detect rosella, was visually estimated for the summer surveys from the plotted data, which clearly levelled off in an asymptotic manner. Mathematical methods were not used to model an asymptote for these data as they did level off distinctly. An asymptote could therefore be quickly and easily estimated by visual means (visual examination of cumulative curves is common in species richness-sampling effort studies, e.g. Newmark Citation1991). An asymptote was not fitted to the winter data, as the data showed a strong linear tendency without appearing to level off (Thompson et al. Citation2003). To calculate an accurate asymptote for winter rosella detection, surveys of longer duration would be necessary (see Results and Discussion).
Parametric tests were used wherever possible; however, relative rosella abundance and time to first rosella detection were not normally distributed (Shapiro-Wilks test) and could not be fitted to the normal distribution even after transformation, hence the use of non-parametric tests. Averages are given as medians (Mdn) where non-parametric tests were used. Means are shown with their standard errors (±SEM). The alpha level was set at 0.05 for all two-tailed statistical tests. Statistical analyses were performed using JMP version 7.0.1 for Mac (SAS Institute Inc., Cary, NC, 2007).
Results
Repeatability
The three survey variables, which were specifically compared between data collected simultaneously by two independent observers during the summer sessions, showed high levels of agreement and repeatability (A–C). The highest value of repeatability was associated with estimating relative rosella abundance between the observers, followed by avian species richness (). A small but significant difference was found between observers in the number of avian species recorded per summer surveys (t 15=2.24, P=0.04). Specifically, the secondary observer counted significantly fewer avian species on average (11.0±0.68 spp./survey) compared with the primary observer (11.75±0.69 spp./survey) (x obs2-obs1= −0.75±0.34 spp.). No significant differences were found between observers either in counts of relative abundance (Mdn obs1=2.5 rosella/survey; Mdn obs2=2.5 rosella/survey; W=1.50, n=16, P=0.63), or in the average time to first detection (Mdn obs1=0.15 min; Mdn obs2=0.02 min; W=2.50, n=16, P=0.69).
Figure 2 Agreement between primary and secondary observer avian point-count surveys (n=16) conducted in the Auckland region during the austral summer. A, Agreement between observers in detection latency. B, Agreement between observers in eastern rosella relative abundance (number of individuals/30 min). C, Agreement between observers in avian richness (number of species/30 min). Observer 1 was the primary researcher (JAG). The regression line is shown as solid grey and the line of x=y is dashed in grey. Units are the same for both axes.
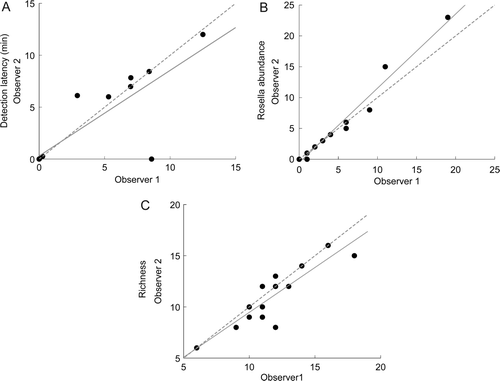
Table 2 Repeatability of eastern rosella (Platycercus eximius) point-count surveys for summer months.
Detectability
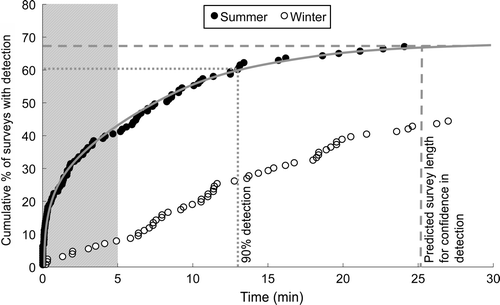
Rosella were detected in 96 of the 143 surveys undertaken during the summer. The detection frequency (percentage of counts where rosella were detected) was significantly higher than that from the winter surveys (67.1% vs 44.4%) (χ2=14.03, P<0.0002). In those surveys with rosella detection, rosella were detected much earlier on average in summer months (Mdn=2.55 min) in comparison with winter months (Mdn=11.2 min) (Z=6.01, n summer=96, n winter =56, P<0.0001). During the summer, 90% of first detections were within the first 13 min of a survey, compared with the winter surveys where 90% of first detections occurred within 22 min. From the estimated asymptote of detection (), a survey length of 25 min or longer is necessary in summer months to be confident rosella have been detected if they are present. In contrast, no such estimation was possible for the continuously rising winter survey data (), implying that a duration of greater than 30 min is required to effectively detect rosella in the winter.
Figure 3 Time to first detection of eastern rosella against the cumulative percentage of surveys with detection at this time. The shaded area denotes where data were truncated for 5-min comparisons. The estimated asymptote of detection (solid line), the point at which 90% of first detections occurred (dotted line), and the predicted necessary survey time for confidence in detecting the presence of rosella (dashed line), are shown for summer data only.
A significantly greater proportion of the first rosella detections in summer surveys were identified using auditory (79.2%) rather than visual means (20.8%) (χ2=32.67, P<0.0001). The observed ratio of auditory to visual detection did not differ significantly between sites (χ2=8.33, P=0.30).
Site type, replicate location, date and time of day were found to have no significant effect on detection latency in the stepwise regression model (F≤2.01, P≥0.11 in all cases). These were removed from the GLMM model (with the exception of site, retained as a random effect). Season (F=36.41, df = 1, P<0.0001), relative rosella abundance (F=45.28, df = 1, P<0.0001) and their interaction term (F=21.53, df = 1, P<0.0001) were found to be significant predictors in the final GLMM that explained variation in detection latency (whole model test: F=144.57, df = 49, P<0.0001).
Survey duration
Detection frequency was reduced dramatically when survey data were truncated at the 5-min mark compared with data collected over the entire 30-min survey duration (). For summer surveys, where rosella were detected in 67.1% of surveys after 30-min, the overall detection frequency would have decreased to 40.6% if the survey timeframe was limited to 5-min periods; a 39.6% reduction in detection success. This decrease was more substantial for winter surveys, where a 44.4% detection frequency at 30 min was reduced to just 7.9% at 5 min; a decrease of 82.1%. In contrast, overall relative rosella abundances at 5 vs 30 min increased roughly proportionally to the increase in survey duration, for both summer and winter data ().
Table 3 Mean eastern rosella relative abundance measures for surveys conducted in the Auckland region.
Discussion
The levelling off of the summer detection curve for eastern rosella in its introduced range in New Zealand well below 100% is indicative of the general difficulties associated with avian surveys for detecting species presence. When we consider the wide-ranging behaviour of this species, particularly combined with low-density sites, we predict that there will be some instances where rosella will not be present within a survey area, even though they are known to occupy a study site. This variation in distribution across time and space is one of many barriers to detection, which cannot be avoided by standardising sampling protocols; thus any analyses must account for some level of error in any survey method (Bibby et al. Citation2000, Thompson Citation2002). More concerning is that the results we would have seen, had we employed the standard 5-min point-count method, would have fallen severely short of the detection frequencies reported overall, and disproportionately so between the different seasons of our surveys. For instance, 5-min counts in summer that recorded the presence of rosella would have been reduced by more than a third, and strikingly, winter surveys with detection would be reduced by more than three quarters. Where detection probability is low (as observed for the 5-min point counts), it is predicted abundance estimates will be underestimated (Thompson Citation2002; Wobeser Citation2007). Contrary to expectations, observed estimates of relative rosella abundance for 5 and 30 min periods were roughly proportional to the increase in survey timeframe. It is possible that the tendency of rosella to move through the landscape in small flocks (Cannon Citation1984; Higgins Citation1999; Fraser Citation2008; JAG unpubl. data) offsets the expected underestimation of abundance, as this flocking behaviour means that, where rosella are detected within 5 min, multiple individuals are counted. Five-minute point counts do have advantages in being fast, easily implemented and widely known (Thompson Citation2002); nevertheless, our results highlight the quantitative reduction and disproportional shifts in detection frequencies when detectability is not accounted for. These reductions, in a formal study of rosella populations, would subsequently influence conclusions about site occupancy, and potentially estimates of rosella abundance.
Our study used extensive ecological replication of site types, including habitat, locality, level of urbanisation and mammal control management, whereas the comparison of seasonal effects was limited to a single year study. Therefore, the distinctive seasonal effects reported here in the latencies required to detect eastern rosella confidently in point-count surveys in New Zealand can only be taken as suggestive evidence for a potentially prevalent seasonal effect. Nonetheless, three lines of evidence indicated that rosella detectability was higher over summer months in our surveys. First, detection of rosella in summer surveys occurred much sooner on average in comparison with winter months. Second, a significantly greater proportion of summer point-count surveys detected rosella overall. Finally, 90% of first detections were within the first 13 min of a survey, compared with the winter surveys where 90% of first detections occurred within 22 min. The survey length necessary for near certain detection of rosella was estimated to be 25 min or greater in the summer. Although also expected to follow an exponential curve and reach an asymptote, the winter data did not cease to track a positive linear relationship, indicating that the survey length required for maximum detection likelihood in winter was longer than the 30-min duration we employed in our own study. Accurately calculating the true asymptote would require a further set of winter surveys to be carried out, with the duration of point-count sessions extended.
The variation seen in detection latency was attributable to relative abundance and season, as well as their interaction. We would expect to see a link between detection time and relative abundance, as a greater number of individuals in a given area will result in a greater likelihood of observing at least one of them, therefore decrease the time to first detection. Population sizes of avian species typically fluctuate with season, because of breeding and juvenile recruitment contributing in the spring/summer and increased mortality over the winter (Ekman Citation1984; Galetti Citation1997; Drechsler et al. Citation1998). The differences seen in rosella detection between summer and winter surveys may be related in part to these actual oscillations in population size via juvenile recruitment and mortality.
However, population dynamics are almost certainly not the sole factor driving the observed patterns in rosella detections. For example, there are also changes in rosella behaviour throughout the year. From studies conducted in its native range, eastern rosella individuals are described as ‘vocal and active’ during the breeding season, and ‘quiet and less conspicuous’ during the non-breeding season (Higgins Citation1999, p. 347). Agonistic behaviours increase between adult pairs and within juvenile groups with the arrival of spring (Brereton Citation1971a, Citationb). We noted similar behavioural changes in introduced rosella (Galbraith Citation2010). Within a 2-week period in mid-August, birds became perceptibly more vocal, with increased occurrence of the piping call, and the noisy chatter associated with the agonistic tail-wagging behaviour, indicating an increase in intraspecific interactions and communication. These observations support the scenario that there are demonstrable differences in the conspicuousness of individuals between seasons, which are responsible, at least in part, for the results generated in these surveys, as conspicuousness is invariably linked to detectability.
Observer bias may have contributed to some of the variation in rosella detection between seasons; however, our results demonstrate that there is a good degree of repeatability (0.67) in detection latency measures between different observers. Interestingly, the measure of repeatability calculated for detection latency was strongly influenced by one extreme data point. With the removal of the point from calculations, repeatability increased from 0.67 to 0.98. This particular survey was conducted on Motuihe, where rosella abundance is relati vely low (Galbraith Citation2010). The primary observer (JAG) heard a rosella, but the secondary independent observer failed to detect this or other calls. This was the sole opportunity to detect rosella during that particular survey session; hence the large discrepancy between detection latency recorded by observer 1 and observer 2.
Recently, Simons et al. (Citation2009) suggested that the probability of detection depends on both the probability that a bird is available for detection (i.e. calls during the count period) and the probability that the observer will detect it, given that it is available. We predict that in situations where there is low rosella abundance, consequently low availability, the failure of an observer to perceive a single call shall affect survey reliability to a much greater extent than should the same occur during a survey in an area of high abundance, as there will be fewer opportunities to detect an individual in the areas where the former occurs. This scenario highlights the necessity of having observers with extensive experience in avian surveying, and in particular a strong ability in call recognition and discrimination where vocalisations occur infrequently, as ultimately what a surveyor records depends on their ability to distinguish and identify avian species (Sauer et al. Citation1994; Robertson et al. Citation2007). The need for a strong ability in auditory discrimination of bird calls is further supported by the finding that the majority of first detected rosella were heard rather than seen. This requirement is also supported by results from other studies, which show a strong reliance in avian surveys on auditory detection (DeJong & Emlen Citation1985; Simons et al. Citation2009).
In conclusion, our study demonstrates that the standard 5-min point-count method is not sufficient for accurately surveying eastern rosella populations within its introduced range in New Zealand. Differences in detectability between seasons are arguably a consequence of behavioural changes in conspicuousness. These differences are also sizeable enough to warrant the design of rosella-specific survey techniques that account for seasonal fluctuations in detectability. To detect the presence of rosella confidently at a site during the summer, a survey length of 25 min is necessary. This timeframe is predicted to be much longer for the winter months, and dedicated further studies of surveying over longer durations are needed to determine the winter asymptote of detection accurately for the species. A problem arises though with increasing survey length, as the probability of detecting the same bird on more than one occasion also increases (Reynolds et al. Citation1980), leading to an overestimation of abundance. Thus, there is a trade-off between confidence in detecting rosella when they are present in a survey area and accurately estimating abundance or density. Where winter surveying is essential, results should be interpreted with caution. We advocate that surveys undertaken in the summer months should be longer than the standard 5-min period, with a duration of 13–15 min being favourable. This will ensure a high level of confidence in detecting any rosella present, and simultaneously maintain a minimal risk of counting the same individual more than once. Ultimately, survey efficiency for detection will be increased, while maintaining reliability and accuracy of other population parameter estimates.
Acknowledgements
We thank the Auckland Regional Council, the University of Auckland and the Ornithological Society of New Zealand for funding this study. Many thanks also to M. Galbraith (Unitec), S. Galbraith and K. Stevens for their assistance in the field, and T. Lovegrove (Auckland Regional Council) for stimulating discussions. Finally we are grateful to M. Stanley and two anonymous reviewers, whose helpful comments greatly improved the manuscript. This study was conducted with permission given by the Auckland Regional and North Shore City Councils, to conduct research in regional and local parks, and was approved by the University of Auckland Animal Ethics Committee (Permit R653).
References
- Bart , J . 2009 . Monitoring the abundance of bird populations . The Auk , 122 : 15 – 25 .
- Berg , ML and Bennett , ATD . 2010 . The evolution of plumage colouration in parrots: a review . Emu , 110 : 10 – 20 .
- Bibby , CJ . 1999 . Making the most of birds as environmental indicators . Ostrich , 70 : 81 – 88 .
- Bibby , CJ and Buckland , ST . 1987 . Bias of bird census results due to detectability varying with habitat . Acta Oecologica/Oecologia Generalis , 8 : 103 – 112 .
- Bibby , CJ , Burgess , ND , Hill , DA and Mustoe , S . 2000 . Bird census techniques , 2nd edition , New York : Academic Press .
- Blumstein , DT . 2003 . Flight-initiation distance in birds is dependent on intruder starting distance . The Journal of Wildlife Management , 67 : 852 – 857 .
- Boulinier , T , Nichols , JD , Sauer , JR , Hines , JE and Pollock , KH . 1998 . Estimating species richness: the importance of heterogeneity in species detectability . Ecology , 79 : 1018 – 1028 .
- Brereton , JL . 1971a . “ A self-regulation to density-independent continuum in Australian parrots, and its implication for ecological management ” . In The scientific management of animal and plant communities for conservation , Edited by: Duffy , E and Watts , AS . 207 – 221 . Oxford : Blackwell .
- Brereton , JL . 1971b . “ Inter-animal control of space ” . In Behaviour and environment: the use of space by animals and men , Edited by: Esser , AH . 69 – 91 . New York : Plenum Press .
- Buckland , ST , Anderson , DR , Burnham , KP , Laake , JL , Borchers , DL and Thomas , L . 2001 . Introduction to distance sampling: estimating abundance of biological populations , Oxford : Oxford University Press .
- Cannon , CE . 1984 . Flock size of feeding eastern and pale-headed rosellas (Aves: Psittaciformes) . Australian Wildlife Research , 11 : 349 – 355 .
- Cassey , P , Craig , JL , McArdle , BH and Barraclough , RK . 2007 . Distance sampling techniques compared for a New Zealand endemic passerine (Philesturnus carunculatus rufusater) . New Zealand Journal of Ecology , 31 : 223 – 231 .
- Dawson , DG and Bull , PC . 1975 . Counting birds in New Zealand forests . Notornis , 22 : 101 – 109 .
- DeJong , MJ and Emlen , JT . 1985 . The shape of the auditory detection function and its implications for songbird censusing . Journal of Field Ornithology , 56 : 213 – 223 .
- Drechsler , M , Burgman , MA and Menkhorst , PW . 1998 . Uncertainty in population dynamics and its consequences for the management of the orange-bellied parrot Neophema chrysogaster . Biological Conservation , 84 : 269 – 281 .
- Ekman , J . 1984 . Density-dependent seasonal mortality and population fluctuations of the temperate-zone willow tit (Parus montanus) . Journal of Animal Ecology , 53 : 119 – 134 .
- Fernández-Juricic , E . 2000 . Bird community composition patterns in urban parks of Madrid: the role of age, size and isolation . Ecological Research , 15 : 373 – 383 .
- Fraser EA 2008 . The winter ecology of the eastern rosella (Platycercus eximius) in New Zealand . Unpublished BSc (Hons) dissertation , University of Auckland , Auckland . P. 73 .
- Galbraith JA 2010 . The ecology and impact of the introduced eastern rosella (Platycercus eximius) in New Zealand . Unpublished MSc thesis , University of Auckland , Auckland .
- Galetti , M . 1997 . Seasonal abundance and feeding ecology of parrots and parakeets in a lowland Atlantic forest of Brazil . Ararajuba , 5 : 115 – 126 .
- Gill , BJ . 1980 . Abundance, feeding and morphology of passerine birds at Kowhai Bush, Kaikoura, New Zealand . New Zealand Journal of Zoology , 7 : 235 – 246 .
- Griffiths , M and van Schaik , CP . 1993 . The impact of human traffic on the abundance and activity periods of Sumatran rainforest wildlife . Conservation Biology , 7 : 623 – 626 .
- Heinsohn , R , Legge , S and Endler , JA . 2005 . Extreme reversed sexual dichromatism in a bird without sex role reversal . Science , 309 : 617 – 619 .
- Higgins , PJ . 1999 . Handbook of Australian, New Zealand and Antarctic birds , Vol. 4 , Melbourne : Oxford University Press .
- Igic , B , Hauber , ME , Galbraith , JA , Grim , T , Dearborn , DC , Brennan , PLR , Moskat , C , Choudary , PK and Cassey , P . 2010 . Comparison of micrometer-and scanning electron microscope-based measurements of avian eggshell thickness . Journal of Field Ornithology , 81 : 402 – 410 .
- Johnson DH 1995 . Point counts of birds: what are we estimating? In : Ralph CJ , Sauer JR , Droege S Monitoring bird populations by point counts . Forest Service General Technical Report PSW-149 . Albany, NY , US Department of Agriculture . Pp 117 – 123 .
- Lee , DC and Marsden , SJ . 2008 . Adjusting count period strategies to improve the accuracy of forest bird abundance estimates from point transect distance sampling surveys . Ibis , 150 : 315 – 325 .
- Lessells , CM and Boag , PT . 1987 . Unrepeatable repeatabilities: a common mistake . The Auk , 104 : 116 – 121 .
- McLennan J , McCann T 2002 . Genetic variability, distribution and abundance of great spotted kiwi (Apteryx haastii) . In : Overmars F Some early 1990s studies in kiwi (Apteryx spp.) genetics and management . Science and Research Internal Report 191 . Wellington , Department of Conservation . Pp. 35 – 55 .
- Newmark , WD . 1991 . Tropical forest fragmentation and the local extinction of understory birds in the eastern Usambara Mountains, Tanzania . Conservation Biology , 5 : 67 – 78 .
- Nichols , JD , Hines , JE , Sauer , JR , Fallon , FW , Fallon , JE and Heglund , PJ . 2000 . A double-observer approach for estimating detection probability and abundance from point counts . The Auk , 117 : 393 – 408 .
- Noon , BR . 1981 . The distribution of an avian guild along a temperate elevational gradient: The importance and expression of competition . Ecological Monographs , 51 : 105 – 124 .
- Ortiz-Catedral , L and Brunton , DH . 2009 . Nesting sites and nesting success of reintroduced red-crowned parakeets (Cyanoramphus novaezelandiae) on Tiritiri Matangi Island, New Zealand . New Zealand Journal of Zoology , 36 : 1 – 10 .
- Perault , DR and Lomolino , MV . 2000 . Corridors and mammal community structure across a fragmented, old-growth forest landscape . Ecological Monographs , 70 : 401 – 422 .
- Petit DR , Petit LJ , Saab VA , Martin TE 1995 . Fixed-radius point counts in forests: factors influencing effectiveness and efficiency . In : Ralph CJ , Sauer JR , Droege S monitoring bird populations by point counts . Forest Service General Technical Report PSW-149 . Albany, NY , US Department of Agriculture . Pp 51 – 59 .
- Reynolds , RT , Scott , JM and Nussbaum , RA . 1980 . A variable circular-plot method for estimating bird numbers . The Condor , 82 : 309 – 313 .
- Rivera-Milan , FF , Collazo , JA , Stahala , C , Moore , WJ , Davis , A , Herring , G , Steinkamp , M , Ron , P , Thompson , JL and Bracey , W . 2005 . Estimation of density and population size and recommendations for monitoring trends of Bahama Parrots on Great Abaco and Great Inagua . Wildlife Society Bulletin , 33 : 823 – 834 .
- Robertson , CJR , Hyvonen , P , Fraser , MJ and Prichard , CR . 2007 . Atlas of bird distribution in New Zealand , Wellington : Ornithological Society of New Zealand .
- Sauer , JR , Peterjohn , BG and Link , WA . 1994 . Observer differences in the North American Breeding Bird Survey . The Auk , 111 : 50 – 62 .
- Simons , TR , Alldredge , MW , Pollock , KH , Wettroth , JM and Dufty , AM . 2009 . Experimental analysis of the auditory detection process on avian point counts . The Auk , 124 : 986 – 999 .
- Spurr , EB . 2005 . Monitoring bird populations in New Zealand: a workshop to assess the feasibility of a national bird population monitoring scheme , Lincoln : Manaaki Whenua Pr ess .
- Taylor , LR . 1984 . Assessing and interpreting the spatial distributions of insect populations . Annual Review of Entomology , 29 : 321 – 357 .
- Thompson , GG , Withers , PC , Pianka , ER and Thompson , SA . 2003 . Assessing biodiversity with species accumulation curves; inventories of small reptiles by pit-trapping in Western Australia . Austral Ecology , 28 : 361 – 383 .
- Thompson , WL . 2002 . Towards reliable bird surveys: accounting for individuals present but not detected . The Auk , 119 : 18 – 25 .
- Verner , J and Milne , KA . 1989 . Coping with sources of variability when monitoring population trends . Annales Zoologici Fennici , 26 : 191 – 199 .
- Witmer , GW . 2005 . Wildlife population monitoring: some practical considerations . Wildlife Research , 32 : 259 – 263 .
- Wobeser , GA . 2007 . Disease in wild animals: investigation and management , 2nd edition , Springer : Berlin .
- Wright D 2001 . Population dynamics and nesting and feeding ecology of the eastern rosella (Platycercus eximius) in northern New Zealand . Unpublished MSc thesis , University of Auckland , Auckland .