Abstract
Artificial retreats are increasingly being used as a tool for monitoring lizard populations in New Zealand. However, relationships between placement of retreats and skink sightings are not well understood. We investigated how microhabitat around and under artificial retreats influences sightings of common skinks (Oligosoma polychroma) in a grassland habitat in Fiordland. Our dataset comprised 2356 skink sightings from 4100 checks of artificial retreats. Mean vegetation height 0.5 m from retreats was positively correlated with skink sightings, but we did not detect relationships between other microhabitat variables and skink counts. Greater vegetation height may provide enhanced habitat complexity, thus supporting a richer faunal community and higher skink density. Alternatively, skink basking opportunities may be more limited under taller vegetation, making nearby artificial retreats more attractive for their thermal properties. We highlight the importance of considering microhabitat when placing artificial retreats to maximise detections of lizards.
Introduction
Reliable inventory and monitoring methods are essential for evaluating animal population trends. Catch (or sightings) per unit effort (CPUE) measures generated from timed searches or checking monitoring devices are often used to compare population trends over time or relative abundance among sites. To generate robust comparisons from CPUE measures, detection of animals must be constant over time or among sites. A range of factors, including detection method (Lettink & Cree Citation2007), sex or age of animals (Senar et al. Citation1999), resource availability (Alterio et al. Citation1999) and placement of monitoring devices relative to microhabitat features (Hyde & Simons Citation2001; Cameron et al. Citation2005; Lettink & Seddon Citation2007), can affect detection probabilities. Microhabitat may influence use of retreats by lizards because it can affect both the thermal advantage provided by retreats relative to the environment (Thierry et al. Citation2009) and resource availability (Lettink & Seddon Citation2007; Norbury et al. Citation2009). Evaluating effects of microhabitat variables on detection probabilities of animals is an important component of designing rigorous protocols for new monitoring techniques (Lettink & Seddon Citation2007).
Artificial retreats are increasingly being used to detect and evaluate trends in lizard abundance in New Zealand (Lettink & Cree Citation2007; Wilson et al. Citation2007; Bell Citation2009; Hoare et al. Citation2009; Thierry et al. Citation2009). Using artificial retreats is simple, cost-effective and low-impact relative to other monitoring techniques (Sutton et al. Citation1999; Lettink & Seddon Citation2007). Recent research has addressed optimal weather conditions during which retreats should be checked for consistency (Hoare et al. Citation2009) and accuracy and precision of index counts from retreats relative to known density (Lettink et al. Citation2011). The present study focussed on the effect of microhabitat of retreats on detecting skinks.
We investigated the influence of microhabitat beneath and around retreats in grassland habitat on the sightings of common skinks (Oligosoma polychroma). Previous work has demonstrated that substrate type (grass or moss) beneath retreats alone does not influence skink sightings, but interactions with substrate height have not been tested (Hoare et al. Citation2009). We evaluated the effects of habitat composition and vegetation height beneath and 0.5 m away from retreats on skink sightings, using data collected over the summer of 2010–11. Based on field observations, we predicted that skinks would be seen more frequently where deep moss occurs beneath retreats (J.M. Hoare, pers. obs.).
Materials and methods
The study was conducted in a 1.5 km2 mixed exotic–indigenous grassland on the floor of the Eglinton Valley, Fiordland. The grassland has been recovering from sheep (Ovis aries) grazing since their removal 13 years ago. Exotic grasses (especially red fescue [Festuca rubra] and sweet vernal [Anthoxanthum odoratum]) dominate the emergent layer, but the grassland now contains a large proportion of indigenous mosses and scattered, prostrate shrubs and herbs. Common skinks are small diurnal, heliothermic skinks that reach densities of 3600–9200 individuals per hectare and readily use artificial retreats in the grassland (Lettink et al. Citation2011).
Artificial retreats used comprised single 670×420 mm sheets of Onduline, a light-weight corrugated roofing material that provides lizards with an opportunity for indirect basking (Lettink Citation2007; Thierry et al. Citation2009). Retreats were set along eight transect lines located randomly, at least 100 m apart, in January 2009. Each line comprised 25 retreats at 10 m intervals, and each sheet was placed on a patch of grass cut to c. 5 cm prior to placement in order to aid skink detection.
Each line of retreats was checked during optimal weather conditions (Hoare et al. Citation2009) between 20 and 22 times between November 2010 and February 2011. A retreat check consisted of lifting the Onduline sheet and recording the number of skinks observed.
We measured vegetation height and composition underneath and 0.5 m from each retreat. Vegetation height was measured beneath an Onduline sheet placed on top of the vegetation. We recorded:
| 1. | Mean vegetation height beneath each retreat: calculated from measurements at the central high and low corrugations of each sheet (four measures/sheet). | ||||
| 2. | Vegetation composition beneath each retreat: percentage of bare ground, moss, live shrubs/herbs and grass, and dry material beneath each retreat. | ||||
| 3. | Mean vegetation height 0.5 m from each retreat: calculated from measurements at the central low corrugation of an Onduline sheet temporarily placed 0.5 m from each side of the retreat (eight measurements). | ||||
| 4. | Primary vegetation type 0.5 m from each retreat: categorised as either live shrubs/herbs and grass, dry material, moss, mixture of grass material (both live and dry grass and shrubs/herbs) or a mixture of all vegetation types. | ||||
Results
The 4100 retreat checks conducted resulted in 2356 skink sightings (). Sightings per retreat ranged from zero to four skinks.
Table 1 Summary of common skink (Oligosoma polychroma) sightings beneath artificial retreats in the Eglinton Valley, Fiordland, in the summer of 2010–11.
Mean height of vegetation beneath retreats was 36 mm (range 20–62 mm) compared with 84 mm (range 29–120 mm) 0.5 m from retreats. Dry grass material was the dominant vegetation type under the retreats (mean = 73%) while moss was dominant under 14% of retreats. Live vascular plants were dominant around 60% of retreats, but an equal mixture of moss and vascular plants were found around 20% of retreats.
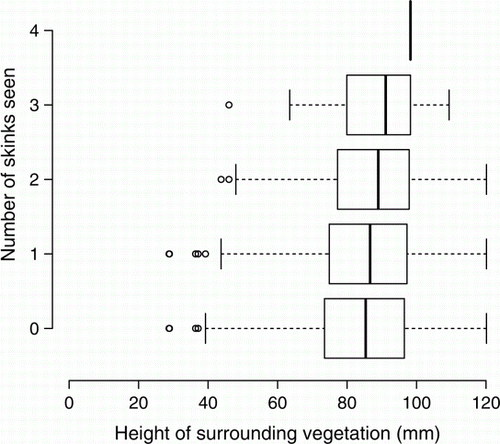
Mean vegetation height 0.5 m from retreats was the only microhabitat factor (or interaction) for which we detected a relationship with skinks (at α=0.05); it was positively correlated with skink sightings (t 191=3.214, P=0.002, r 2 =0.141; ). Based on GLMM results, we predict an extra 0.06 skink sightings per retreat for every extra centimetre of vegetation surrounding retreats; this equates to an extra 12 skinks per 200 retreat checks.
Figure 1 Sightings of common skinks (Oligosoma polychroma) beneath Onduline artificial retreats in grassland habitat in relation to mean height of vegetation 0.5 m from the retreats. Dark vertical lines represent the median data point in each category; boxes represent quartiles; dotted lines extend to 1.5 times the interquartile range; ○, outliers beyond this range.
Discussion
Our finding, that mean vegetation height surrounding artificial retreats is positively correlated with sightings of common skinks in grassland, may be because of either habitat complexity supporting more skinks or lack of basking opportunities in taller vegetation. Norbury et al. (2009) found that faunal species richness was positively correlated with vegetation cover and reported a positive (though non-significant) relationship between skink numbers and vegetation ground cover. Vegetation height may act as a proxy for habitat complexity, invertebrate richness and potential refugia, which are all likely to be positively correlated with carrying capacity for skinks. Alternatively, taller vegetation may limit basking opportunities within the home range of skinks (which is likely to be small in the Eglinton grassland given the high density estimates [Lettink et al. Citation2011]). Placing Onduline artificial retreats in tall grass may represent a thermal opportunity for skinks disproportionate to that offered in short grassland habitat.
Other factors that may influence skink sightings beneath artificial retreats include distance to the nearest divaricating shrubs (i.e. fruit resources) and abiotic microsite effects (e.g. substrate type [Lettink et al. Citation2008]). Lettink and Seddon (Citation2007) found that presence of divaricating shrubs and vines within 5 m of pitfall traps influenced sightings of common and McCann's skinks (O. maccanni) in coastal habitat; a similar relationship is possible in the Eglinton grassland. However, we consider abiotic microsite effects unlikely to influence skink sightings in homogeneous terrain such as that found in the Eglinton grassland.
Our result that vegetation composition beneath and around retreats has no detectable influence on sightings of grassland skinks extends a conclusion previously drawn by Hoare et al. (2009). Our hypothesis that skinks were seen more frequently under retreats with deep moss beneath them was not supported by these analyses, possibly because moss provides more opportunities for concealment and thus lower detectability than other substrate types. Furthermore, variability in height of vegetation beneath retreats (which we attempted to minimise, but could not eliminate, prior to deployment of retreats) did not influence detections. Instead, the thermal advantages offered by retreats appear to exert more influence on sightings than microhabitat characteristics (Thierry et al. Citation2009; this study).
Our findings support previous work by Lettink & Seddon (2007) and have implications for designing monitoring protocols for sampling skinks using artificial retreats. We recommend that practitioners using artificial retreats to detect lizards consider microhabitat preferences of focal species and use a stratified design, or deliberately place retreats at microsites within a fixed sampling configuration, to maximise detections.
Acknowledgements
We thank Maddie Charlton, Aaron Fenner, Nicola Fullerton, Iris Jacobs, Dan Palmer and Jason van der Wetering for field assistance and Marieke Lettink and Grant Norbury for valuable advice on the manuscript. The Department of Conservation (Science Investigation No. 4231) and Victoria University of Wellington provided funding.
References
- Alterio , N , Moller , H and Brown , K . 1999 . Trappability and densities of stoats (Mustela erminea) and ship rats (Rattus rattus) in a South Island Nothofagus forest, New Zealand . New Zealand Journal of Ecology , 23 : 95 – 100 .
- Bell , TP . 2009 . A novel technique for monitoring highly cryptic lizard species in forests . Herpetological Conservation and Biology , 4 : 415 – 425 .
- Cameron , BG , Van Heezik , Y , Maloney , RF , Seddon , PJ and Harraway , JA . 2005 . Improving predator capture rates: analysis of river margin trap site data in the Waitaki Basin, New Zealand . New Zealand Journal of Ecology , 29 : 117 – 128 .
- Hoare , JM , O'Donnell , CFJ , Westbrooke , I , Hodapp , D and Lettink , M . 2009 . Optimising the sampling of skinks using artificial retreats based on weather conditions and time of day . Applied Herpetology , 6 : 379 – 390 .
- Hyde , EJ and Simons , TR . 2001 . Sampling plethodontid salamanders: sources of variability . Journal of Wildlife Management , 65 : 624 – 632 .
- Lettink M 2007 . Adding to nature: can artificial retreats be used to monitor and restore lizard populations? Unpublished PhD thesis: University of Otago , Dunedin . 190 p.
- Lettink , M and Cree , A . 2007 . Relative use of three types of artificial retreats by terrestrial lizards in grazed coastal shrubland, New Zealand . Applied Herpetology , 4 : 227 – 243 .
- Lettink M , Cree A , Norbury G , Seddon PJ 2008 . Monitoring and restoration options for lizards on Kaitorete Spit, Canterbury . DOC Research & Development Series , 301 . Wellington, Department of Conservation . 44 p.
- Lettink , M , O'Donnell , CFJ and Hoare , JM . 2011 . Accuracy and precision of skink counts from artificial retreats . New Zealand Journal of Ecology , 35 : 236 – 243 .
- Lettink , M and Seddon , PJ . 2007 . Influence of microhabitat factors on capture rates of lizards in a coastal New Zealand environment . Journal of Herpetology , 41 : 187 – 196 .
- Norbury , G , Heyward , R and Parkes , J . 2009 . Skink and invertebrate abundance in relation to vegetation, rabbits and predators in a New Zealand dryland ecosystem . New Zealand Journal of Ecology , 33 : 24 – 31 .
- Senar , JC , Conroy , MJ , Carrascal , LM , Domènech , J , Mozetich , I and Uribe , F . 1999 . Identifying sources of heterogeneity in capture probabilities: an example using the great tit, Parus major . Bird Study , 46 : S248 – S252 .
- Sutton , PE , Musinsky , HR and McCoy , ED . 1999 . Comparing the use of pitfall drift fences and cover boards for sampling the threatened sand skink (Neoseps reynoldsi) . Herpetological Review , 30 : 149 – 151 .
- Thierry , A , Lettink , M , Besson , AA and Cree , A . 2009 . Thermal properties of artificial refuges and their implications for retreat-site selection in lizards . Applied Herpetology , 6 : 307 – 326 .
- Wilson , DJ , Mulvey , RL and Clark , RD . 2007 . Sampling skinks and geckos in artificial cover objects in a dry mixed grassland-shrubland with mammalian predator control . New Zealand Journal of Ecology , 31 : 169 – 185 .