Abstract
Dietary analyses were carried out on four species of native birds on Tiritiri Matangi and Motuora Islands, New Zealand, between February 2007 and February 2008. We examined regurgitated pellets of kingfishers, moreporks and swamp harriers, and the faeces of pukeko. Remains of lizards were found in 88% and 43% of regurgitated kingfisher pellets on Tiritiri Matangi and Motuora, respectively. All remains belonged to one species, Oligosoma moco. No evidence of lizard predation was found for the other three bird species. Translocation of lizards to areas supporting a high abundance of kingfishers should take avian predation risk into account.
Introduction
In New Zealand, studies addressing the impacts of predation on lizards have focused almost entirely on introduced mammalian predators (Whitaker Citation1973; McCallum Citation1986; Miskelly Citation1997; Jones et al. Citation2005; Hoare et al. Citation2007). This has been in response to significant population declines and extinctions of endemic herpetofauna caused by mammalian predation (Whitaker Citation1973, Citation1978; Worthy Citation1987). The effects of predation by native animals on lizards are poorly understood, although there are records of lizard predation by invertebrates (Lettink & Patrick Citation2006), fish (Anonymous Citation2003), reptiles (Whitaker Citation1994) and birds (Marples Citation1942; Fitzgerald et al. Citation1986).
Internationally, birds have been regarded the most important predators of reptiles (Martin & López 1996). In New Zealand, at least two species of introduced birds, Eurasian blackbird (Turdus merula merula; Bell Citation1996) and common starling (Sturnus vulgaris vulgaris; Thompson Citation2000), and 12 species of native birds have been recorded preying upon lizards. These include banded rail (Rallus philippensis; Whitaker Citation1968), black-fronted tern (Chlidonias albostriatus; O'Donnell & Hoare Citation2009), North Island fernbird (Bowdleria punctata vealeae; Ball & Parrish Citation2005), kakapo (Strigops habroptilus; Falla et al. Citation1979), long-tailed cuckoo (Eudynamys taitensis; Oliver Citation1955), morepork (Ninox novaeseelandiae; Ramsay & Watt Citation1971; Imboden Citation1975; Haw & Clout Citation1999), New Zealand falcon (Falco novaeseelandiae; Fox Citation1977; Worthy Citation1997), New Zealand kingfishers (Todiramphus sanctus vagans; Mead Citation1947; O'Donnell Citation1981), pukeko (Porphyrio melanotus melanotus; Muggeridge & Cottier Citation1931; Carroll Citation1966), swamp harrier (Circus approximans; Pierce & Maloney Citation1989), takahe (Porphyrio hochstetteri; Whitaker Citation1991) and western weka (Gallirallus australis australis; Coleman et al. Citation1983). Reports of avian predation on lizards are largely based on anecdotal accounts and the few studies that have explored the predator–prey relationships between birds and lizards in New Zealand have done so with limited success (Schneyer Citation2001; Jordan Citation2005).
On offshore islands, native bird populations are known to reach considerable densities and resident lizard populations may provide an especially important food resource for avian predators (Schoener & Schoener Citation1978). Such predator–prey relationships have typically evolved within natural communities where established prey populations are generally robust enough to withstand the pressures exerted by natural predators (Bodini Citation1991). However, populations translocated for conservation reasons may not be able to withstand such predation, as even if a small proportion of a bird's diet consists of lizards, this may be sufficient to impact negatively on a founder lizard population (Martín & López Citation1996). A lack of understanding of predator–prey dynamics between native species could have significant impacts on the outcome of threatened species translocations. For example, unforeseen predation by pukeko and swamp harriers appears to have limited the population expansion of brown teal (pateke, Anas chlorotis) translocated to Tiritiri Matangi Island (Armstrong Citation2006). Similarly, the failure of a translocated population of threatened shore plover (Thinornis novaeseelandiae) to establish on Motuora Island was attributed to unanticipated predation by morepork (Aikmen Citation1999). The introduction of threatened bird species to islands outside of their natural range (e.g. takahe on Tiritiri Matangi Island) may result in unanticipated predation of lizards and could lead to their extinction from the island if the lizard population were small (Atkinson Citation1990). In New Zealand, recognition of the significance of lizards in functioning ecosystems has been an important impetus for their translocation to islands undergoing restoration (Whitaker Citation1978; Payton et al. Citation2002). Clearly there is a need for research and insight into lizard predation by native birds in order to understand the impacts of such predations. Such knowledge is important for the success of further translocations of threatened species (Armstrong & McLean Citation1995; Towns Citation1999).
In recognition of the lack of quantitative data, this study aimed to estimate the rate of avian predation on native lizards, on two offshore island sanctuaries, and provide a basis on which informed management decisions can be made during threatened lizard species translocations. This study was conducted on two offshore islands; Tiritiri Matangi (36°36′ S, 174°53′ E, 220 ha) and Motuora (36°30′ S, 174°47′ E, 80 ha) Islands (Hauraki Gulf, Auckland, New Zealand) as part of a wider lizard translocation and monitoring project. Introduced mammals have never established on Motuora (Gardner-Gee et al. Citation2007); however, Tiritiri Matangi once supported populations of kiore (Rattus exulans), rabbits (Oryctolagus cuniculus), cats (Felis catus) and goats (Carpus hircus). By 1993, all introduced mammals had been eradicated from Tiritiri Matangi (Mitchell Citation1985; Galbraith & Hayson Citation1994). These islands are currently free of introduced mammals and are subject to ecological restoration by way of re-vegetation and faunal re-introduction programmes. These two islands share four species of lizards, while Tiritiri Matangi supports one further lizard species and tuatara (Sphenodon punctatus) ().
Table 1 Status and density of lizard species present on Motuora and Tiritiri Matangi Islands.
Materials and methods
Dietary analyses were conducted on four resident species of native birds known to prey upon lizards. These included pukeko, kingfisher, morepork and swamp harriers. Diets were assessed using several techniques, including analyses of regurgitated pellets, faecal material, nest material and stomach contents (where available).
The two islands were systematically searched for kingfisher and morepork nest and roost sites between February 2007 and February 2008. Faecal and regurgitated pellet samples were collected from across the islands and frozen to prevent decay and damage by invertebrates. Nest samples were collected from morepork and kingfishers after each successive fledging to avoid disturbing the nesting birds. Faecal samples from pukeko were collected from across the islands every month. Dead birds of these four species were collected opportunistically as they became available, and stored in a −20 °C freezer until examination. The entire alimentary tract (i.e. oesophagus, gizzard, stomach and intestines) of deceased birds was examined for the presence of lizard remains.
All regurgitated pellets were examined under a dissecting microscope. The prey composition from each pellet was expressed as the frequency of pellets containing a prey item. Prey items included the following categories: insects, decapods (e.g. crabs), fish and lizards. The number of individual lizards per pellet was estimated by counting the number of brain cases, jaw bones (i.e. two dentaries and two maxilla per lizard), frontal bones and pelvic girdles. Lizard remains were identified to species by morphological characteristics of the dentaries and maxilla (Worthy Citation1987), and by comparisons with museum reference skeletons. Prey remains in nest samples were not quantified since some remains may have been deposited during previous nesting seasons. Seasonal dietary analyses were not carried out because of the lack of comparable data across seasons.
Results
One hundred regurgitated pellets were collected from nine individual kingfishers on Tiritiri Matangi and seven pellets collected from three individual kingfishers on Motuora. Lizard remains were present in 88 (88%) of the Tiritiri Matangi kingfisher pellets and in three (43%) of the Motuora kingfisher pellets. Details of pellet contents are shown in . All lizard remains were identified as one species, Oligosoma moco, based on the size and shape of the jaw bones. The average number of lizards per pellet varied depending on the type of skeletal remains present in the pellets (i.e. braincase, jaw bones, frontal bone and pelvic girdle) (). Conservative estimates of the number of lizards per pellet were two per pellet on Tiritiri Matangi, and one per pellet on Motuora.
Figure 1 Percentage of regurgitated kingfisher pellets containing prey items from Tiritiri Matangi (n=100) and Motuora (n=7) Islands.
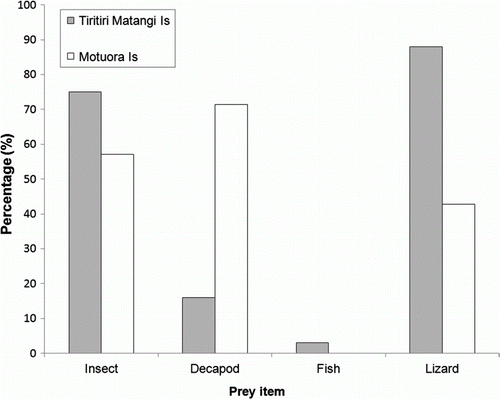
Table 2 Average number (±SEM) of skink bones recorded in regurgitated kingfisher pellets containing lizard remains, on Tiritiri Matangi and Motuora Islands in 2007.
Two regurgitated pellets were collected from swamp harriers on Tiritiri Matangi, and neither of these pellets contained lizard remains. No swamp harrier pellets were collected on Motuora.
Lizard remains were found in six (60%; n=10) kingfisher nests on Tiritiri Matangi, but were recorded less often in nests on Motuora (23.8%, n=21). Remains of all other prey categories were detected in nest holes on both islands.
No lizard remains were identified in regurgitated morepork pellets from Tiritiri Matangi (n=67, 12 birds) or Motuora (n=107, nine birds). Prey consumption consisted almost entirely of invertebrates, with wētā (Orthoptera: Anostostmatidae) occurring in 85.1% and 89.5% of owl pellets on Tiritiri Matangi and Motuora, respectively. Bird remains were occasionally apparent in pellets from owls on both islands. No lizard remains were found in morepork nests on either island.
Pukeko faecal samples from Tiritiri Matangi (n=50) and Motuora (n=50) were comprised entirely of vegetative matter and did not contain lizard remains. Similarly, no lizard remains were observed within the guts of a swamp harrier and partially decomposed pukeko.
Discussion
Evidence of lizard predation was found only in the diet of kingfishers. The high proportion of kingfisher pellets and nests containing lizard remains, relative to the other identified prey categories, suggests that lizards form an important component of the kingfishers’ diet on these islands. Previous studies have also noted the importance of lizard prey items in the diet of both adult and nestling kingfishers (Burton Citation1998; Blomberg & Shine Citation2000; Asokan et al. Citation2009; Soud et al. Citation2010). For example, Asokan et al. (Citation2009) reported that 24% of white-breasted kingfisher (Halcyon smyrnensis) regurgitated pellets contained the remains of lizard prey.
All lizards consumed by kingfishers in our study consisted exclusively of O. moco. This lizard is a diurnal heliotherm and is common on the islands, with an estimated density of up to 4617 per ha in grassland habitat on Tiritiri Matangi (Habgood Citation2003). The diurnal, sun-basking activity exhibited by this species is likely to have contributed to the high rate of predation by visually oriented predators such as kingfishers (Anholt & Werner Citation1975). The vulnerability of diurnal heliotherms to predation is emphasised by the lack of Oligosoma aeneum (a crepuscular species) in the diet, despite this species occurring at densities of up to 6173 per ha in grassland habitat on Tiritiri Matangi (Habgood Citation2003).
Predation is a function of prey availability (Anholt & Werner Citation1975), and the higher proportion of lizard remains in kingfisher pellets on Tiritiri Matangi, compared with that of invertebrate remains, may be related to high lizard prey abundance, or perhaps reflects selection of more protein-rich prey items, especially during periods of high energy demand such as breeding seasons (O'Donnell & Hoare Citation2009). However, the overall impact of kingfishers on the island lizard populations could not be determined because of the lack of information pertaining to demographics and structure of the lizard populations.
Despite the high density estimates of O. moco and O. aeneum on Tiritiri Matangi, lizard remains were not detected in the diet of morepork, pukeko and swamp harrier in this study, even though these birds have been recorded preying on lizards. This may indicate that lizard predation by these species is low and occurs opportunistically. However, larger sample sizes would be required to verify this.
Translocations
In New Zealand, lizard conservation efforts have focused on translocating threatened species to offshore islands free of introduced mammalian predators (Armstrong & McLean Citation1995; Towns & Ferreira Citation2001). Founder populations are generally small because of the limitations of harvesting source populations, and opportunistic predation of founders may be great enough to exacerbate the effects associated with K-strategist lizard species. For example, following the translocation of highly threatened Cyclodina whitakeri to Korapuki Island, population models suggested that the population became highly vulnerable to collapse if predation of adults exceeded 5% per annum (Towns Citation1994).
Translocating reptiles into unfamiliar environments has potential to disrupt their behaviours, especially as reptiles may move more frequently to explore, or as a result of disorientation (Plummer & Mills Citation2000; Sullivan Citation2004). Increased activity and frequency of movement through unfamiliar habitat may ultimately affect the survival and establishment of a lizard population because of increased risk of predation (Plummer & Mills Citation2000). Post-release monitoring of 30 translocated Oligosoma smithi on Motuora has revealed a decrease in detectability through time (M. Baling pers. comm. 2011) following their release at a beach site known to support kingfishers. Although the ultimate cause of this decline in delectability cannot be ascertained, a combination of storm damage to habitat and predation by kingfishers could be proximate causes. In comparison, the O. smithi population translocated to Tiritiri Matangi has shown a marked increase in numbers. The success of this population establishment has been attributed to the provision of adequate natural refuges throughout the release habitat, which are likely to have offered the skinks protection from storm events and avian predators. Future studies involving lizard population monitoring and kingfisher dietary analyses following the second release of O. smithi onto Motuora in February 2011 may provide a more rigorous insight into the effects of avian predation on small translocated lizard populations.
Assessing the risk of predation on translocated lizard populations may incorporate approaches such as modelling possible outcomes based on known life history traits of the lizard species involved. An understanding of predator–prey dynamics between birds and lizards, and the associated risks, can potentially improve the outcome of threatened species translocations. Conservation managers should be aware of the potential impacts of resident birds, notably kingfishers, on small native lizards and should consider these factors when planning translocations of threatened endemic lizards; especially diurnally active species. Strategies to mitigate predation risk include: avoiding the release of lizards at locations or microhabitats where densities of avian predators are known to be high, releasing lizards outside of the avian predator breeding season, selecting release sites with adequate natural refuges and food resources, and enhancing habitat with additional refuges (artificial or otherwise) (Webb & Shine Citation2000; Lettink Citation2007). In addition, removing potential hunting posts (e.g. disused fence posts, dead tree stumps) at the release sites might aid in reducing the risk of kingfisher predation of released lizards.
Acknowledgements
We are grateful to Richard Sharp and Helen Lindsay for their abseiling skills, to Dave Jenkins for his assistance on Motuora Island, and to Trent Bell, Sarah Wells and Marleen Baling for their constructive comments. Funding was provided by the Society for Research on Amphibians and Reptiles in New Zealand Herpetological Research Award, the Supporters of Tiritiri Matangi Research Fund, the Motuora Restoration Society and Massey University. This study was endorsed by the Department of Conservation (permit AK-19734-FAU) and Massey University Animal Ethics Committee (Protocol 06/98).
References
- Aikmen , H . 1999 . Attempts to establish shore plover (Thinornis novaeseelandiae) on Motuora Island, Hauraki Gulf . Notornis , 46 : 195 – 205 .
- Anholt , BR and Werner , EE . 1975 . Interaction between food availability and predation mortality mediated by adaptive behaviour . Ecology , 76 : 2230 – 2234 .
- Anonymous 2003 . Lizards as snapper bait? In : New Zealand Fishing News . Auckland , Rugby Press . 8 .
- Armstrong DP 2006 . Reintroduction projects in New Zealand . Reintroduction Specialist Group, Oceania Section . Retrieved 16 April 2008, from http://www.massey.ac.nz/~darmstro/nz_projects.htm .
- Armstrong , DP and McLean , IG . 1995 . New Zealand translocations: theory and practice . Pacific Conservation Biology , 2 : 39 – 54 .
- Asokan , S , Ali , AMS and Manikannan , R . 2009 . Diet of three insectivorous birds in Nagapattinam District, Tamil Nadu, India—a preliminary study . Journal of Threatened Taxa , 1 : 327 – 330 .
- Atkinson IAE 1990 . Ecological restoration on islands: prerequisites for success . In : Towns DR , Daugherty CH , Atkinson IAE . Ecological restoration of New Zealand islands . Conservation Sciences Publication No. 2, Department of Conservation , Wellington . 73 – 90 .
- Ball , O and Parrish , R . 2005 . Lizard predation by North Island fernbird (Bowdleria punctata vealeae) . Notornis , 52 : 250 – 251 .
- Bell , BD . 1996 . Blackbird (Turdus merula) predation on the endemic copper skink (Cyclodina aenea) . Notornis , 43 : 213 – 217 .
- Blomberg , SP and Shine , R . 2000 . Size-based predation by kookaburras (Dacelo novaeguineae) on lizards (Eulamprus tympanum: Scincidae): what determines prey vulnerability? . Behavioral Ecology and Sociobiology , 48 : 484 – 489 .
- Bodini , A . 1991 . What is the role of predation on stability of natural communities? A theoretical investigation . BioSystems , 26 : 21 – 30 .
- Burton , NHK. 1998 . Notes on the diet of nestling white-throated kingfishers Halcyon smyrnensis in Malaysia . Forktail , 14 : 79 – 80 .
- Carroll , ALK . 1966 . Food habits of pukeko (Porphyrio melanotus Temmink) . Notornis , 13 : 133 – 142 .
- Coleman , JD , Warburton , B and Green , WQ . 1983 . Some population statistics and movements of the western weka . Notornis , 30 : 93 – 109 .
- Falla , RA , Sibson , RB and Turbott , EG . 1979 . The new guide to the birds of New Zealand and outlying islands , Auckland : Collins .
- Fitzgerald , BM , Meads , MJ and Whitaker , AH . 1986 . Food of the kingfisher (Halcyon sancta) during nesting . Notornis , 31 : 23 – 33 .
- Fox N 1977 . The biology of the New Zealand falcon (Falco novaeseelandiae Gmelin 1788). Unpublished PhD thesis , University of Canterbury , Christchurch .
- Galbraith MP , Hayson CR 1994 . Tiritiri Matangi Island, New Zealand: public participation in species translocation to an open sanctuary . In : Serena M . Reintroduction biology of Australian and New Zealand fauna . Chipping Norton , Surrey Beatty & Sons . 149 – 154 .
- Gardner-Gee R , Graham S , Griffiths R , Habgood M , Dunlop SH , Lindsay H 2007 . Motuora native species restoration plan . Auckland , Motuora Restoration Society Inc .
- Habgood MJ 2003 . Behavioural interactions between copper (Cyclodina aenea) and moko (Oligosoma moco) skinks: implications for translocations . Unpublished MSc thesis , University of Auckland , Auckland .
- Haw , JM and Clout , MN . 1999 . Diet of morepork (Ninox novaeseelandiae) throughout New Zealand by analysis of stomach contents . Notornis , 46 : 333 – 345 .
- Hoare , JM , Pledger , S , Nelson , NJ and Daugherty , CH . 2007 . Avoiding aliens: behavioural plasticity in habitat use enables large, nocturnal geckos to survive Pacific rat invasions . Biological Conservation , 136 : 510 – 519 .
- Imboden , C . 1975 . A brief radio-telemetry study on moreporks . Notornis , 22 : 221 – 231 .
- Jones , C , Moss , K and Sanders , M . 2005 . Diet of hedgehogs (Erinaceus europaeus) in the upper Waitaki Basin, New Zealand: implications for conservation . New Zealand Journal of Ecology , 29 : 29 – 35 .
- Jordan M 2005 . An impact assessment report on the pukeko (Porphyrio porphyrio melanotus) population at Shakespear Regional Park . Auckland Regional Council Report .
- Lettink , M and Patrick , BH . 2006 . Use of artificial cover objects for detecting red katipo, Latrodectus katipo Powell (Araneae: Theridiidae) . New Zealand Entomologist , 29 : 99 – 102 .
- Lettink M 2007 . Adding to nature: can artificial retreats be used to monitor and restore lizard populations . Unpublished PhD thesis , University of Otago , Dunedin .
- Marples , BJ . 1942 . A study of the little owl, Athene noctua, in New Zealand . Transactions and Proceedings of the Royal Society of New Zealand , 72 : 237 – 252 .
- Martin J , López P 1990 . Amphibians and reptiles as prey of birds in Southwestern Europe . Smithsonian Herpetological Information Service No. 82 . Washington , Smithsonian Institute . 1 – 43 .
- Martín , J and López , P . 1996 . Avian predation on a large lizard (Lacerta lepida) found at low population densities in Mediterranean habitats: an analysis of bird diets . Copeia , 1996 : 722 – 726 .
- McCallum , J . 1986 . Evidence of predation by kiore upon lizards from the Mokohinau Islands . New Zealand Journal of Ecology , 9 : 83 – 87 .
- Mead , WP . 1947 . Classified summarised notes . Notornis , 2 : 50
- Miskelly CM 1997 . Whitaker's skink Cyclodina whitakeri eaten by a weasel Mustela nivalis . Conservation Advisory Science Notes No. 146 , Department of Conservation , Wellington .
- Mitchell , ND . 1985 . The revegetaion of Tiritiri Matangi Island: the creation of an open sanctuary . Royal New Zealand Horticultural Society Annual Journal , 13 : 36 – 41 .
- Muggeridge J , Cottier W 1931 . Food habits of the pukeko . New Zealand Journal of Science and Technology XIII : 36 .
- O'Donnell , CFJ . 1981 . Foods of the New Zealand kingfisher (Halcyon sancta vagans) . Notornis , 28 : 140 – 141 .
- O'Donnell , CFJ and Hoare , JM . 2009 . Predation of lizards by black-fronted terns (Sterna albostriata) . Notornis , 56 : 167 – 168 .
- Oliver , WRB . 1955 . New Zealand birds , 2nd edition , Wellington : Reed .
- Payton IJ , Fenner M , Lee WG 2002 . Keystone species: the concept and its relavance for conservation management in New Zealand . Science for Conservation 203 . Department of Conservation , Wellington .
- Pierce , RJ and Maloney , RF . 1989 . Responses of harriers in the MacKenzie Basin to the abundance of rabbits . Notornis , 36 : 1 – 12 .
- Plummer , MV and Mills , NE . 2000 . Spatial ecology and survivorship of resident and translocated hognose snakes (Heterodon platirhinos) . Journal of Herpetology , 34 : 565 – 575 .
- Ramsay , GW and Watt , C . 1971 . Notes on the birds of Great Island, Three Kings Islands . Notornis , 18 : 287 – 290 .
- Schneyer N 2001 . Effects of avian predation and habitat degradation on the population dynamics of the jewelled gecko (Naultinus gemmeus) from the Every Scientific Reserve, Otago Peninsula, New Zealand . Unpublished MSc thesis , Otago University , Dunedin .
- Schoener , TW and Schoener , A . 1978 . Inverse relation of survival of lizards with island size and avifaunal richness . Nature , 274 : 685 – 687 .
- Soud , R , Mazumdar , K and Grupta , A . 2010 . Predation by white-throated kingfisher Halcyon smyrnensis on common wolf snake Lycodon aulicus (Linnaeus) . NeoBIO , 1 : 53 – 54 .
- Sullivan , BK , Kwiatkowski , MA and Schuett , GW . 2004 . Translocation of urban gila monsters: a problematic conservation tool . Biological Conservation , 117 : 235 – 242 .
- Thompson , MB . 2000 . Oligosoma spp. (New Zealand skinks) predation . Herpetological Review , 31 : 175
- Towns , DR . 1994 . The role of ecological restoration in the conservation of Whitaker's skink (Cyclodina whitakeri), a rare New Zealand lizard (Lacertilia: Scincidae) . New Zealand Journal of Zoology , 21 : 457 – 471 .
- Towns DR 1999 . Cyclodina spp. skink recovery plan 1999–2004 . DOC Threatened Species Recovery Plan 27 , Department of Conservation , Wellington .
- Towns , DR and Ferreira , SM . 2001 . Conservation of New Zealand lizards (Lacertilia: Scincidae) by translocation of small populations . Biological Conservation , 98 : 211 – 222 .
- Webb , JK and Shine , R . 2000 . Paving the way for habitat restoration: can artificial rocks restore degraded habitats of endangered reptiles? . Biological Conservation , 92 : 93 – 99 .
- Whitaker , AH . 1968 . Lizards of the Poor Knights Islands New Zealand . New Zealand Journal of Science , 11 : 653 – 651 .
- Whitaker , AH . 1973 . Lizard populations on islands with and without polynesian rats, Ratus exulans (Peale) . Proceedings of the New Zealand Ecological Society , 20 : 121 – 130 .
- Whitaker AH 1978 . The effects of rodents on reptiles and amphibians . The ecology and control of rodents in New Zealand Nature Reserves . Wellington , New Zealand Department of Lands and Survey Information Service 4 . 75 – 86 .
- Whitaker AH 1991 . Research on the striped gecko (Hoplodactylus stephensi) on Maud Island, Pelorus Sound, Marlborough: 6–12 March 1991 . Unpublished report , Department of Conservation, Marlborough-Nelson Conservancy , Nelson .
- Whitaker AH 1994 . New Zealand lizards ecology and conservation . Threatened Species Unit, Ecological Management, No. 2, Department of Conservation , Wellington .
- Worthy , TH . 1987 . Osteological observations on the larger species of the skink Cyclodina and the subfossil occurrence of these and the gecko Hoplodactylus duvaucelii in the North Island, New Zealand . New Zealand Journal of Zoology , 14 : 219 – 229 .
- Worthy , TH . 1997 . Diets of New Zealand falcons and laughing owls—a palaeontological perspective . Kokako , 4 : 10 – 12 .