Abstract
Bat carcasses from two wind farms in Tasmania (2002–2010) were assessed to determine the species, sex, age, reproductive state, morphometrics, presence of food in the gastrointestinal tract, and evidence of spatial and seasonal patterns. Thirty-eight of the 54 carcasses were Gould's wattled bats, with another 14 likely to be, and two Vespadelus sp. All but two were adults, with an equal ratio of females and males. None were actively breeding when found, and five of the six bats tested, had not been recently feeding. Mortalities predominantly occurred in autumn, with a small difference between sites. There was no pattern in the location of carcasses. There appear to be particular ecological, morphological and behavioural characteristics associated with bat collision risk—tree roosting bats with high wing aspect ratios that forage in the open air at high altitude appear to be susceptible. Seasonal patterns may be associated with specific behaviours.
Introduction
The wind industry is relatively new to the Australasian region, particularly compared with that in Europe, the UK and the USA. Partly as a result of this, there are no data published in the scientific literature on the impact of wind farms (WFs) on bats in this region. This taxon has been identified as being at risk of collisions with wind turbines, with bat fatalities being recorded in North America (e.g. Arnett et al. Citation2008) and Europe (Rydell et al. Citation2010b).
Studies to date have indicated that bat fatalities occur as a result of traumatic injury following collisions with moving turbines and possibly from barotrauma (Baerwald et al. Citation2008), although the incidence of barotrauma has been recently queried, with collisions thought to be the primary cause of fatality (Grodskey et al. Citation2011; Rollins et al. Citation2012).
Tree roosting or migratory bats have been found to be more prone to fatalities at WFs overseas than other groups (Cryan & Barclay Citation2009). However, it is unknown if the same patterns occur in the Australasian region. Understanding which species are susceptible to wind turbine associated fatalities and from which sex and age class they belong is necessary (Kunz Citation2004), and this, along with any existing population data, will inform debates around whether WFs are having a significant impact (i.e. resulting in a decline in the status of populations) on important (threatened or significant for another reason) bat populations. If significant impacts are found, greater focus can be turned to understanding why bat fatalities occur at WFs and to developing strategies to reduce these impacts. Furthermore, understanding the impacts in this region is important to inform the assessment of new WF projects.
One of the first large-scale WFs in Australia was approved in 2000 (the Woolnorth WF, now operated as the Bluff Point WF and Studland Bay WF and now owned and operated by Woolnorth Holdings, ). The Bluff Point WF was developed in two stages, with stage 1 (six turbines) commissioned in August 2002, and stage 2 (an additional 31 turbines) commissioned in May 2004. The Studland Bay WF was developed in one stage and was fully commissioned in April 2007.
Figure 1 Location of the Bluff Point and Studland Bay Wind Farms in north-west Tasmania.
Monitoring of bird and bat collisions was conducted from the commencement of operation (2002 at Bluff Point WF and 2007 at Studland Bay WF) until September 2010, when the focus of monitoring changed to eagles at both sites (although bat fatalities were still documented). Bat carcasses collected during dead bird and bat surveys at the two WFs were assessed in this study and cover the period 2002–2010. As these bats are those found, and it was impossible to determine the level of undetected bats due to a lack of randomness in the survey design, estimates of WF-wide bat mortality rates cannot be made, and the focus is only on an assessment of those found (note that Driessen et al. Citation2011 cite a WF-wide figure for bat fatalities at the Bluff Point WF, but as this figure did not take these factors into account, the figures do not represent WF-wide mortality rates).
The purpose of the current study was to identify the species of bat colliding with turbines and to determine if there were specific characteristics that may make them susceptible to fatality. Morphometrics and other information were also collected to better understand the specific characteristics of the bats and if particular cohorts or parts of the population were more vulnerable than others. An assessment of any evidence of temporal (seasonal) and spatial (whether some circuits of turbines had higher counts of bat fatalities than others) patterns was also undertaken to attempt to shed light on the factors involved in bat collision risk.
Methods
Study sites
Both WFs are in north-west Tasmania, with the Bluff Point WF comprising 37 Vestas V66 turbines (33-m blades, 60-m tower). It is located on 1524 ha, on a flat-topped bluff with a steep coastal cliff to the west (coastal side) and moderately steep slopes to the east, north and south. Approximately 40% of the land is cleared for cattle grazing (with some fodder cropping), with the remainder being high quality remnant vegetation (including closed melaleuca forest and wet coastal shrubby Eucalypt forest). This vegetation has been fenced to exclude cattle.
The Studland Bay WF is approximately 3 km south of Bluff Point and comprises 25 Vestas V90 wind turbines (45-m blades, 80-m tower). It is located on 1410 ha, and is on less elevated terrain consisting of flat undulating land with isolated rocky outcrops and large consolidated dunes with some small ephemeral fresh water lagoons close to the coast (and to the north of the site). Approximately 60% of the land is cleared for cattle grazing (with some fodder cropping) and the remaining land (comprising coastal swamp forest, coastal heath and pockets of wet coastal shrubby Eucalypt forest) is fenced to exclude stock.
Carcass monitoring surveys
The strategy used in the carcass monitoring surveys was broadly consistent with that described in US Fish and Wildlife Service Guidelines (Citation2012) and Rodrigues et al. (Citation2008). It was consistent in terms of early liaison with Regulators, the duration of surveys, surveys during relevant seasons, scavenger and detectability trials, survey interval, attempts to survey different parts of the WFs and assessment of migrants vs residents. Some of the requirements in the guidelines, including estimating WF-wide fatality rates, comparisons with other WFs (in Australia) and comparisons with estimated collision rates could not be achieved because of a lack of the necessary data or information.
Surveys for bats beneath the turbines were conducted throughout the year with survey frequency determined from a series of scavenger trials (Hydro Tasmania, unpubl. data) and regulatory requirements. The initial survey frequency was twice weekly in spring and autumn, and fortnightly outside these periods. However, from 2006, survey frequency was increased during seasons with what were thought to be heightened Tasmanian wedge-tailed eagle (Aquila audax fleay) activity.
Following completion of stage 1 at the Bluff Point WF, all turbines were surveyed, but after stage 2, a sub-sample (27%) of turbines were surveyed during the formal surveys. A sub-sample (25%) of turbines was surveyed at the Studland Bay WF following its completion. In 2007 the survey regime was changed further, with all turbines surveyed at both sites. The formal surveys were complimented by ad hoc surveys, where all site personnel and visitors were required to report bat (or bird) injuries or carcasses found on site.
Observers searched beneath turbines in concentric circles on foot and using slow-moving 4WD motor bikes from the base of the turbine tower out to 100 m (consistent with Hull & Muir Citation2010). The concentric circles were 2–10 m apart, with the wider spacing used during eagle searches (as eagle carcasses were more conspicuous and readily detected; Hydro Tasmania, unpubl. data). Sub-sampled turbines were surrounded with predator-proof fences (chicken mesh, buried beneath the ground, with additional electrified wires at the top of the fence to prevent climbing by scavengers) to minimise removal by mammalian scavengers, and pasture was maintained at a low height to increase visibility.
A total of 54 bats (37 from the Bluff Point WF, 10 from the Studland Bay WF and seven where the site could not be determined) were available for this analysis, from a total of 75 (62 from the Bluff Point WF and 13 from the Studland Bay WF) found at these sites. All had been stored in a freezer following collection until the time of analysis.
Morphometric and other measurements taken
The following measurements and investigations were made on the carcasses, wherever possible:
| • | Species—identified by head characteristics, and head, forearm and head–body measurements (based on Taylor et al. Citation1987, Parnaby Citation1992 and Churchill Citation2009). Carcasses with missing heads or with broken forearms or skulls could not be identified to species. Carcasses with missing heads and intact forearms were classified to the nearest species matching that forearm length; | ||||
| • | Age—determined through examination of wing joints (based on Churchill Citation2009 and categorised as adult, sub-adult or juvenile); | ||||
| • | Sex (based on Churchill Citation2009); | ||||
| • | Reproductive state (adapted from Phillips & Inwards Citation1985 and Churchill Citation2009. Females were categorised as: non-reproductive (nipples regressed or absent), pregnant (palatable foetus in abdomen), lactating (nipple enlarged, area surrounding the nipple hairless will milk production evident) and post-lactating (nipples prominent, but fur regrowing and no milk production evident). Males were categorised as: non-reproductive (testis not enlarged), stage 1–3 (testis enlarged, testis increasing in size from 1 to 4 mm: sperm production), stage 4 (peak testicular development, testis increasing in size from 4 to 7 mm, epididymis distended: sperm production and storage) and stage 5 (testis regressed with distended epididymis: sperm storage); | ||||
| • | Morphometrics: forearm, head, head to body length, tail length (based on Churchill Citation2009); | ||||
| • | Presence of food in the gastrointestinal tract through dissection. | ||||
As some individuals were partially decomposed, not all the above measurements could be made. Only six of the samples were in a sufficiently fresh condition to determine if there was food in the gastrointestinal tract. Morphometrics were not taken on individuals that showed evidence of decomposition or damage to the relevant body part. Measurements were made with calipers and are reported to 0.1 mm. Mean±standard deviations are provided. Morphometrics between the sexes were compared using t-tests and a probability of 0.05 was used to indicate a significant effect.
Temporal and spatial analyses
The small sample sizes precluded some statistical analyses, but tests were conducted wherever possible.
To determine if there were patterns in the seasonal distribution of bat carcass finds, a Spearman's Rank Correlation Test (with correction for tied ranks) was used to test the null hypothesis that there was no difference in the distribution between months, sites, and whether the months males and females were found were similar (rejection of the null hypothesis therefore indicating a difference). Tests were conducted after scaling for survey effort.
A spatial analysis was undertaken to assess whether a higher number of bats were found at different parts on the WFs. Given the small sample sizes at each turbine, this comparison was made by comparing the number of bats found at turbine circuits. At the Bluff Point WF, A circuit has six turbines in open pasture, B circuit has seven turbines in cleared pasture, C circuit has six turbines adjacent to remnant vegetation, D circuit has 10 turbines adjacent to remnant vegetation and E circuit has eight turbines predominantly in cleared pasture. At the Studland Bay WF, F circuit has 10 turbines predominantly in cleared pasture, G circuit has five turbines predominantly adjacent to remnant vegetation and H circuit has 10 turbines in a mixture of pasture and adjacent to vegetation. Survey effort was taken into account and χ2 tests for independence were used to test for patterns. A probability of 0.05 was used.
Results
Of the 54 bats analysed, 38 were Gould's wattled bats (Chalinolobus gouldii). Based on forearm length and head appearance (long vs short ears and muzzle ridge shape), 14 were determined to most likely be Gould's wattled bat, although 100% confirmation was not possible. Two of the samples were Vespadelus sp. (species could not be determined with confidence), but based on forearm length, one of these was most likely V. darlingtoni.
The vast majority (45) of bats were adults. One was a sub-adult and the age of eight individuals could not be ascertained.
There were 14 females and 19 males (indeterminable in 21) amongst the samples. Using the approaches of Clopper & Pearson, as described in Agresti (Citation2002), the proportion of females was 42.4%, and was not significantly different to a 50:50 ratio (P=0.48; the 95% confidence levels were 25.5% and 60.8%).
There was a significant correlation between the months that males and females were found (R s=0.57, P=0.09, n=10) at the 90% confidence level, but not at the 95% confidence level, suggesting weak evidence for a difference in the months males and females collided with turbines. Given the weakness in this effect, it was not examined further.
Reproductive state was determined in 23 individuals (indeterminable in 31), with 12 being classified as non-reproductive, four post-lactating, four in reproductive stage 4 and three were in reproductive stage 5.
The morphometrics for each sex (sites combined) are provided in , with only forearm significantly different between the sexes. There were insufficient samples from the Studland Bay WF to test for a difference between sites.
Table 1 The mean±standard deviation of morphometrics of Gould's wattled bats found at the Bluff Point and Studland Bay Wind Farms in north-west Tasmania; both sites combined.
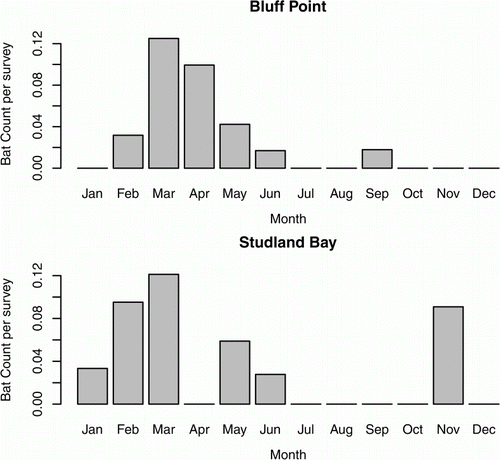
Both sites showed a strong seasonal pattern in bat detections, with the majority of finds occurring in March and April (therefore autumn, ). The months in which bats were detected were not correlated between the sites (R s=0.43, P=0.2, n=12), indicating a difference in months that mortalities occurred. However, given the small sample sizes at the Studland Bay WF, some caution is necessary, as there is a risk of a Type I error.
Figure 2 Monthly patterns when bats were detected at the Bluff Point and Studland Bay Wind Farms. Data have been scaled for survey effort. Due to the scaling for survey effort for each month, the results for November at Studland Bay should be interpreted with caution as it represents one bat find.
Five of the six bats that could be assessed had no food or remains in the gastrointestinal tract. One bat had a full stomach, comprising insect remains.
There was no significant difference in the number of bats detected in the turbine circuits at either site (Bluff Point WF χ2=3.22, DF=4, P>0.5; Studland Bay χ2=4.14, DF=2, P>0.1).
Discussion
There are eight species of bat recorded in Tasmania and two vagrant species (Driessen et al. Citation2011; L Cawthen, unpubl. data). The vast majority of bat fatalities at these WFs were Gould's wattled bat. They are a widespread species endemic to Australia and occur throughout Tasmania in a variety of habitats (Churchill Citation2009; Driessen et al. Citation2011; L Cawthen, unpubl. data). The other species likely to be in the samples, but could not be definitively identified, the large forest bat (V. darlingtoni) is also widely distributed and occurs throughout Tasmania in a variety of habitats (Driessen et al. Citation2011; L Cawthen, unpubl. data). Both species are considered dependent on trees as roost sites year-round (Koch et al. Citation2008; Lumsden et al. Citation2002), but their patterns of roost site selection and behaviour in Tasmania are unknown.
The bat surveys conducted for the Development Proposal and Environmental Management Plan for these two WFs (Hydro Tasmania Citation2000) using harp trapping and call identification, identified lesser long-eared bat (Nyctophilus geoffroyi) and Tasmanian long-eared bat (Nyctophilus sherrini) at both sites, but not Gould's wattled bats or Vespadelus spp. The duration of surveys was limited (4 days), but they were conducted during the season bat fatalities have been recorded in the current study (mid-late autumn).
There are three possible explanations why the species detected in our study were not recorded during the pre-construction surveys: biological (activity of some species, such as Gould's wattled bats and large forest bats, might be affected by poor weather as they forage in open areas, and these surveys may have been in poor weather); technical [surveys were conducted at ground level and not at rotor height—previous studies overseas have found echolocation detectors need to be placed at least 52 m above the ground to adequately detect high flying bats in the region of the rotor swept area (Weller & Baldwin Citation2012; see also Kunz Citation2004; Johnson Citation2010), information that was not available in 1999, when surveys were conducted]; or the species composition of bats has changed since construction of the WFs. There is insufficient information available to definitively determine which of these factors might explain this inconsistency, but given there is currently no evidence to indicate that WFs cause a change in species composition, more likely explanations are technical or biological.
The two species of bats observed on site, but not found in the carcass surveys, are low flyers, foraging in the understory (O'Neil & Taylor Citation1986) and therefore they are not expected in the region of the rotor swept area (at the Bluff Point WF the rotor swept area is 27–93 m and at the Studland Bay WF it is 35–125 m above the ground). In contrast, Gould's wattled bats are high-flying, open-air foragers and V. darlingtoni flies predominantly in the mid-story away from vegetation (O'Neill & Taylor Citation1986), with likely occasional higher flights, potentially placing them in the risk zone. The only other species that is a high flyer in Tasmania is the Eastern falsistrelle (Falsistrellus tasmaniensis; O'Neil & Taylor Citation1986), but they have not been recorded on these sites.
Although there are no data published in the scientific literature on bat fatalities at large, modern wind turbines from Australia, Hall & Richards (Citation1972) reported white-striped freetail bat (Tadarida australis) fatalities at small wind turbines, with 127-cm-long blades, mounted at 12 m above the ground, in NSW, as did Bennett (Citation2012) at two large turbines, with 41-m-long blades and a hub height of 68 m, at the Hepburn WF in Victoria. Pre-construction surveys at the Hepburn WF identified Gould's wattled bats, Southern forest bat (Vespadelus regulus), the eastern broadnosed Bat (Scotorepens orion) and the white-striped freetail bat on site (Richards Citation2011), but not Gould's wattled bats in carcass surveys. Given that the only report available at the time of writing (Bennett Citation2012) did not report on carcass surveys during the autumn period, they may appear in surveys during that season. Like Gould's wattled bats, white-striped freetail bats are high flying, open-air foragers (O'Neill & Taylor Citation1986; Rhodes & Richards Citation2008).
Some studies in North America and parts of Europe have documented migratory bats as being most at risk of collision with wind turbines (Dürr & Bach Citation2004; Kunz Citation2004; Pasqualetti et al. Citation2004; Baerwald et al. Citation2009), but other studies from Europe did not find this pattern (Rydell et al. Citation2010b). Rydell et al. (Citation2010a,Citationb) found that the bat genera at most risk of colliding with wind turbines in north-west Europe were Nyctalus, Pipistrellus and Vespertilio and to some extent also Eptesicus, all of which have long and narrow wings, forage in the open air and tend to be migratory. The North American bats most often killed at wind turbines are morphologically and ecologically similar to those in Europe, but belong to other genera (Lasiurus, Lasionycerteris and Perimyotis; Johnson et al. Citation2004; Kunz et al. Citation2007), features that are consistent with our study.
Bat population sizes in Tasmania are poorly understood, so it is impossible to determine to what extent WF fatalities are impacting on local bat populations. Gould's wattled bats are widespread in Tasmania (Driessen et al. Citation2011; L Cawthen, unpubl. data) and mainland Australia (Churchill Citation2009) and are not currently listed as a threatened species in Tasmania or at the national level. A review by Lumsden & McKenzie (Citation2008) considered the population large and stable. As we were unable to determine the WF-wide bat fatality rate, it is not possible to determine definitively the impact to local populations, but we assume that it is unlikely impacts to the species would be occurring. Given the current level of knowledge about bat moralities at WFs in this region and the recent findings of Voigt et al. (Citation2012) that bats from far afield can collide with WFs in Europe, further studies are required to understand the source of the bats that collide at Australasian WFs and the effect on populations.
The vast majority of bats in the current study were adults. Few studies in the USA reported the age of the bats that collided with wind turbines, often because decomposition or scavenging prevented this, but the few studies that did found mainly adults (Arnett et al. Citation2008). This is consistent with our studies and those in north-western Europe (Rydell et al. Citation2010b). The predominance of adults indicates a lack of support for the hypothesis that young bats may be at risk of collision due to unskilled flight or as a result of dispersal activity (Altringham Citation1996; Arnett et al. Citation2008; Baerwald et al. Citation2008; Cryan & Barclay Citation2009).
An even ratio of males and females was found at these sites. Studies in the USA (Arnett et al. Citation2008) and north-western Europe (Rydell et al. Citation2010a,Citationb) found a predominance of adult males amongst bat fatalities at WFs. However, Rydell et al. (Citation2010b) found that the composition of sex and age differed across north-west Europe, indicating site variability. As habitat use and behaviour can vary with the sex of the bat, although not in Gould's wattled bats (Lumsden et al. Citation2002), it is plausible that sex biases in bat fatalities may occur at some sites in Australasia, particularly if they involve species other than Gould's wattled bats.
The morphometrics of these individuals, including sexual dimorphism, were within the same range as that documented from field and museum specimens from Tasmania (), suggesting that the WF victims did not represent a morphologically distinct subset of the Tasmanian population. Of the morphometrics measured, only forearm had the potential to discriminate the sex of Gould's wattle bat.
Table 2 The mean±standard deviation of morphometrics of Gould's wattled bats from throughout Tasmania obtained from field and museum specimens (L Cawthen, unpubl. data).
There was a strong seasonal pattern (autumn) in bat fatalities at these sites. As surveys were conducted all year round and survey effort taken into account in the analysis, this represents a real pattern and not a sampling artefact. Autumn is predominantly the period of post-reproduction, as females become pregnant in spring and give birth in late spring and throughout summer, while males begin producing sperm in December in preparation for mating in early autumn (Churchill Citation2009). Therefore, the timing of bat fatalities would not result in the direct loss of young.
This seasonality in fatalities is consistent with studies in both the USA (August and September, the northern hemisphere autumn, Kunz et al. Citation2007; Arnett et al. Citation2008) and north-western Europe (Rydell et al. Citation2010b), and is thought to be linked with the bat migratory period (some studies reported in Arnett et al. Citation2008), dispersal (Rydell et al. Citation2010b) or flocking and mating behaviour (Cryan Citation2008). As bats in Tasmania do not migrate, and no juveniles were recorded in the samples, flocking and mating behaviour or a change in foraging behaviour are more parsimonious explanations for this seasonal pattern. Bats may be attracted to the wind turbines as they would tall trees in the forest for attracting a mate (Cryan Citation2008) or as a potential roost site (Cryan & Barclay Citation2009), placing them in the high risk zone. There have been no studies on roost site selection of Gould's wattled bats in Tasmania and no studies on this species mating behaviour to support this theory. However, a study from mainland Australia found that they roosted at the tops of tall trees (Lumsden et al. Citation2002), which may add some support for the theory that bat mortality is, in part, a result of bats investing turbine towers as potential roost sites or gathering sites for mating.
An interesting finding in the current study was the subtle difference in the months that collisions occurred at the two WFs. Although these sites are located in close proximity (approximately 3 km apart), there are some topographical and vegetation differences between them. These differences may result in a slightly different use of the sites by Gould's wattled bats, but further studies are required to confirm this. This result does caution against extrapolating findings from one site to another, even when they are geographically close.
No pattern was found in the location of the bat fatalities at circuits within these WFs, indicating no particular relationship with proximity to the coast, or vegetation (which may have roost sites). Topography and its effect on vegetation have been related to bat habitat choice in the previous studies (e.g. Sedgley & O'Donnell Citation1999; Lloyd et al. Citation2006), but not all (Johnson et al. Citation2004). Given that Gould's wattled bats can travel up to 11 km from their roosts to forage (Lumsden et al. Citation2002), it suggests the small-scale habitat variability on site may not be an important factor in influencing collision risk, or that the bats are not using particular routes through the WF (Piorkowski & O'Connell Citation2010). However, the analysis that was possible on the data in our study was relatively coarse.
The limited evidence indicates that bats had not been recently feeding around the turbines when they died. Johnson (Citation2004) found that collision victims did not involve foraging populations, but other authors have contended that bats might be foraging on insects attracted to the turbines (Kunz Citation2004; Rydell et al. Citation2010a,Citationb).
This study found that high-flying, open-air foraging bats are more at risk of fatality at wind turbines than other species. Males and females were impacted similarly, but there was a predominance of adults suggesting that the WFs were not resulting in mortality of dispersing juveniles and sub-adults. The seasonality in fatalities may be linked with specific behaviours at this time of year, perhaps mating. Further studies are required to determine if the patterns found at these sites are consistent with other sites and habitats in Australasia.
Acknowledgements
We would like to thank Elizabeth Stark of Symbolix for assistance with the statistical analyses, and Mark Venosta, Chris Sims, Stephen Ross and two anonymous referees for comments on the draft. Finally, we would like to thank Woolnorth Holdings Pty Ltd, a Joint Venture between Shenhua Clean Energy Holdings Pty Ltd and Hydro Tasmania, for use of the data.
References
- Agresti , A . 2002 . Categorical data analysis , 2nd edition , New York : John Wiley & Sons .
- Altringham , JD . 1996 . Bats: biology and behaviour , New York : Oxford University Press .
- Arnett , EB , Brown , WK , Erickson , WP , Fielder , JK , Hamilton , BL , Henry , TH , Jain , A , Johnson , GD , Kerns , J , Koford , RR , Nicholson , CP , O'Connell , TJ , Piorkowski , MD and Tankersley , RD Jr . 2008 . Patterns of bat fatalities at wind energy facilities in North America . Journal of Wildlife Management , 72 : 61 – 78 . doi: 10.2193/2007-221
- Baerwald , EF , D'Amours , GH , Klug , BJ and Barclay , RMR . 2008 . Barotrauma is a significant cause of bat fatalities at wind turbines . Current Biology , 18 : R695 – 696 . doi: 10.1016/j.cub.2008.06.029
- Baerwald , EF , Edworthy , J , Holder , M and Barclay , RM . 2009 . A large-scale mitigation experiment to reduce bat fatalities at wind energy facilities . Journal of Wildlife Management , 73 : 1077 – 1081 . doi: 10.2193/2008-233
- Bennett E 2012 . Hepburn Wind Farm bird and bat mortality survey interim report 11th July 2011–9th January 2012 . Elmoby Ecology. http://hepburnwind.com.au/downloads/Hepburn%20Wind%20interim%20report%20Bird%20&%20Bat%20Monitoring.pdf (accessed 25 May 2012) .
- Churchill , S . 2009 . Australian bats , Crows Nest , NSW : Allen and Unwin .
- Cryan , PM . 2008 . Mating behavior as a possible cause of bat fatalities at wind turbines . Journal of Wildlife Management , 72 : 845 – 849 . doi: 10.2193/2007-371
- Cryan , PM and Barclay , RMR . 2009 . Causes of bat fatalities at wind turbines: hypotheses and predictions . Journal of Mammalogy , 90 : 1330 – 1340 . doi: 10.1644/09-MAMM-S-076R1.1
- Driessen , M , Brereton , R and Pauza , M . 2011 . “ Status and conservation of bats in Tasmania ” . In The biology and conservation of Australasian Bats , Edited by: Law , B , Eby , P , Lunney , D and Lumsden , L . 324 – 336 . Mosman , NSW : Royal Zoological Society of NSW .
- Dürr , T and Bach , L . 2004 . Bat deaths and wind turbines—a review of current knowledge, and of the information available in the database for Germany . Bremer Beiträge für Naturkunde und Naturshutz , 7 : 253 – 264 .
- Grodsky , SM , Behr , MJ , Gendler , A , Drake , D , Dieterle , BD , Rudd , RJ and Walrath , NL . 2011 . Investigating the causes of death for wind turbine-associated bat fatalities . Journal of Mammalogy , 92 : 917 – 925 . doi: 10.1644/10-MAMM-A-404.1
- Hall , LS and Richards , GC . 1972 . Notes on Tadarida australis (Chiroptera:Molossidae) . Australian Mammalogy , 1 : 46 – 47 .
- Hull , CL and Muir , S . 2010 . Search areas for monitoring bird and bat carcasses at wind farms using a Monte-Carlo model . Australasian Journal of Environmental Management , 17 : 77 – 87 . doi: 10.1080/14486563.2010.9725253
- Hydro Tasmania 2000 . West Coast Wind Farm Woolnorth, North-West Tasmania . Development proposal and environmental management plan . Hobart , Hydro Tasmania .
- Johnson G 2004 . A review of bat impacts at Wind Farms in the US . In : Proceedings of the Wind Energy and Birds/Bats Workshop: understanding and resolving bird and bat impacts . Washington , DC , American Wind Energy Association and The American Bird Conservancy . Resolve . 46 – 50 .
- Johnson G 2010 . Relationships between bat fatality and weather, marine radar, AnaBat, and night vision data at a wind energy facility in the midwest . In : Wind Wildlife Research Meeting VIII Meeting Proceedings . Washington , DC , National Wind Coordinating Collaborative . Resolve . 21 – 23 .
- Johnson , GD , Perlik , MK , Erickson , WP and Strickland , MD . 2004 . Bat activity, composition and collision mortality at a large wind plant in Minnesota . Wildlife Society Bulletin , 32 : 1278 – 1288 . doi: 10.2193/0091-7648(2004)032[1278:BACACM]2.0.CO;2
- Koch , AJ , Munks , SA and Woehler , EJ . 2008 . Hollow-using vertebrate fauna of Tasmania: distribution, hollow requirements and conservation status . Australian Journal of Zoology , 56 : 323 – 349 . doi: 10.1071/ZO08003
- Kunz TH 2004 . Wind power: bats and wind turbines . In : Proceedings of the Wind Energy and Birds/Bats Workshop: understanding and resolving bird and bat impacts . Washington , DC , American Wind Energy Association and The American Bird Conservancy . Resolve . 50 – 55 .
- Kunz , TH , Arnett , EB , Erickson , WP , Hoar , AR , Johnson , GD , Larkin , RP , Strickland , MD , Thresher , RW and Turtle , MD . 2007 . Ecological impacts of wind energy development on bats: questions, research needs, and hypotheses . Frontiers in Ecology and the Environment , 5 : 315 – 324 . doi: 10.1890/1540-9295(2007)5[315:EIOWED]2.0.CO;2
- Lumsden , LF , Bennett , AF and Silins , JE . 2002 . Location of roosts of the lesser long-eared bat Nyctophilus geoffroyi and Gould's wattled bat Chalinolobus gouldii in a fragmented landscape in south-eastern Australia . Biological Conservation , 106 : 237 – 249 . doi: 10.1016/S0006-3207(01)00250-6
- Lumsden L , McKenzie N 2008 . Chalinolobus gouldii . In: IUCN 2011. IUCN Red List of Threatened Species. Version 2011.2. www.iucnredlist.org (accessed 3 June 2012) .
- Lloyd , A , Law , B and Goldingay , R . 2006 . Bat activity on riparian zones and upper slopes in Australian timber production forests and the effectiveness of riparian buffers . Biological Conservation , 129 : 207 – 220 . doi: 10.1016/j.biocon.2005.10.035
- O'Neill , MG and Taylor , RJ . 1986 . Observations of the flight patterns and foraging behaviour of Tasmanian bats . Australian Wildlife Research , 13 : 427 – 432 . doi: 10.1071/WR9860427
- Parnaby , H . 1992 . An interim guide to identification of insectivorous bats of south-eastern Australia . Technical Reports of the Australasian Museum , 8 : 1 – 35 . doi: 10.3853/j.1031-8062.8.1999.1528
- Pasqualetti M , Richter R , Gip P 2004 . History of wind energy . In : Cleveland C . Encyclopaedia of energy , 6 . Oxford , Elsevier . 419 – 433 .
- Phillips , WR and Inwards , SJ . 1985 . The annual activity and breeding cycles of Gould's long-eared bat, Nyctophilus gouldi (Microchiroptera: Vespertilionidae) . Australian Journal of Zoology , 33 : 111 – 126 . doi: 10.1071/ZO9850111
- Piorkowski , MD and O'Connell , TJ . 2010 . Spatial pattern of summer bat mortality from collisions with wind turbines in mixed-grass prairie . American Midland Naturalist , 164 : 260 – 269 . doi: 10.1674/0003-0031-164.2.260
- Rhodes M , Richards GC 2008 . White-striped free-tailed bat . In : Van Dyck S , Strahan R. The mammals of Australia , 3rd edition . Sydney , NSW , Martin Ford, Reed New Holland . 501 – 502 .
- Richards GC 2011 . An assessment of the bat fauna at the Hepburn Community Wind Farm, Leonards Hill, Victoria . Prepared for Hepburn Wind Park Co-operative Ltd. http://hepburnwind.com.au/downloads/planning/20110304-bat-assessment-report.pdf (accessed 25 May 2012) .
- Rodrigues L , Bach L , Dubourg-Savage M-J , Goodwin J , Harbusch C 2008 . Guidelines for consideration of bats in wind farm projects . EUROBATS Publication Series No. 3 (English version). Bonn, Germany , UNEP/EUROBATS Secretariat .
- Rollins , KE , Meyerholz , DK , Johnson , GD , Capparella , AP and Loew , SS . 2012 . A forensic investigation into the etiology of bat mortality at a wind farm: barotrauma or traumatic injury? . Veterinary Pathology , 49 : 362 – 371 . doi: 10.1177/0300985812436745
- Rydell , J , Bach , L , Dubourg-Savage , MJ , Green , M , Rodrigues , L and Hedenstrom , A . 2010a . Mortality of bats at wind turbines links to nocturnal insect migration? . European Journal of Wildlife Research , 56 : 823 – 827 . doi: 10.1007/s10344-010-0444-3
- Rydell , J , Bach , L , Dubourg-Savage , M-J , Green , M , Rodrigues , L and Hedenström , A . 2010b . Bat mortality at wind turbines in northwestern Europe . Acta Chropterlogica , 12 : 261 – 274 . doi: 10.3161/150811010X537846
- Sedgeley , JA and O'Donnell , CFJ . 1999 . Roost selection by the long-tailed bat, Chalinolobus tuberculatus, in temperate New Zealand rainforest and its implications for the conservation of bats in managed forests . Biological Conservation , 88 : 261 – 276 . doi: 10.1016/S0006-3207(98)00069-X
- Taylor , R , O'Neill , MG and Reardon , T . 1987 . Tasmanian bats: identification, distribution and natural history . Papers and Proceedings—Royal Society of Tasmania , 121 : 109 – 117 .
- US Fish and Wildlife Service 2012 . Land-based Wind Energy Guidelines . US Fish and Wildlife Service , Arlington , VA .
- Weller , TJ and Baldwin , JA . 2012 . Using echolocation monitoring to model bat occupancy and inform mitigations at wind energy facilities . Journal of Wildlife Management , 76 : 619 – 631 . doi: 10.1002/jwmg.260
- Voigt , CC , Popa-Lisseanu , AG , Niermann , I and Kramer-Schadt , S . 2012 . The catchment area of wind farm for European bats: a plea for international regulations . Biological Conservation , 153 : 80 – 86 . doi: 10.1016/j.biocon.2012.04.027