Abstract
Data obtained during carcass and bird utilisation surveys conducted over an approximately 10-year period at two wind farms in northwest Tasmania (Bluff Point Wind Farm [BPWF] and Studland Bay Wind Farm [SBWF]) were compared. Of the species present onsite only 21% at the BPWF and 18% at the SBWF were found to collide with turbines, indicating that presence onsite was a poor indicator of collision risk. Furthermore, there was a poor relationship between abundance onsite and collisions with turbines. A classification and regression tree classified species into the groups that collided or did not collide, based on two classifiers. Specific families/superfamilies and foraging strategies/zones were associated with collision risk and indicated that particular morphological, ecological and behavioural factors were associated with a species' vulnerability to colliding with wind turbines. Future studies should investigate whether the patterns found at these sites are consistent across other habitats and sites.
Introduction
The potential impacts of wind farms on birds can potentially occur during two functional stages of the development of a wind farm—construction and operation. The effect of constructing a wind farm is similar to that of other developments, and is generally brief (approximately one year). Environmental impacts during this phase can be described as ‘pulse’ events (Bender et al. Citation1984) and can usually be minimised through careful environmental management. In contrast, the longer-term operational stage of a wind farm (in the order of 20–25 years) places a different persistence pressure, more in line with ‘press’ events (Bender et al. Citation1984), on populations present.
Press impacts from wind farms can be categorised into direct (collisions with wind turbines) and indirect (‘barrier’ and ‘alienation’, with the potential to reduce access to habitat on or near the wind farm; Langston & Pullan Citation2003). Obviously, collision effects cannot occur if alienation is operating, as the latter results in birds no longer utilising the area around turbines (Madders & Whitfield Citation2006).
Avian collisions with wind turbines have been documented at various sites in North America, the UK and Europe (e.g. Erickson et al. Citation2001; Langston & Pullan Citation2003; Bay Citation2010), but no data from Australia have yet been published in the scientific literature. Given the unique species in this region, and differences such as the lack of large-scale migrations of songbirds and raptors, it is important to understand the effects on Australian species to inform the assessment of new wind farm proposals and the management of existing ones.
Assessments for new wind farm proposals in Australia are currently based on a range of studies, including bird utilisation surveys, to determine the species and numbers present onsite, and then collision risk is estimated using tools such as collision risk modelling (see Smales Citation2006). These risk assessments calculate the extent of collision risk and can be used to inform strategies to reduce this risk (such as the placement of turbines). Often the focus of regulators is threatened species, largely because the consequences of mortality can be high. However, all risk assessments will be improved if there is a better understanding of the species at risk of collision. Furthermore, understanding the factors involved in collision risk is critical to any attempts to mitigate this risk (Barrios & Rodríguez Citation2004).
There has been a range of avian studies conducted at the Bluff Point Wind Farm (BPWF) and Studland Bay Wind Farm (SBWF) in northwest Tasmania (previously called the Woolnorth WF, now owned and operated by Woolnorth Holdings, a joint venture between Shenhua Clean Energy Holdings Pty Ltd. and Hydro Tasmania Wind Operations Pty Ltd.; ) including monitoring bird collisions and bird utilisation surveys. These data have been used in the current study.
Figure 1 Locations of the Bluff Point (top) and Studland Bay (bottom) wind farms.
Using monitoring data from these wind farms, the purpose of the current study was to document the species that collided with turbines and to determine the relationship between those colliding with turbines and those present onsite by comparing the species lists and proportions of taxa in each list. If a mismatch was found between species colliding and species present onsite then the characteristics of those species that collided were evaluated to better understand the factors involved in turbine collision risk. Finally, the study compared the broad characteristics of collision victims at these sites to overseas wind farms.
Other than the study on raptors by de Lucas et al. (Citation2008), we are not aware of any other studies that have assessed collision data in relation to utilisation data, and de Lucas et al. (Citation2008) used the comparison for different purposes.
The current study focused on detected mortalities and did not attempt to quantify undetected mortalities, due to the lack of randomness in the carcass-monitoring survey design and the lack of precision in undetected mortality estimates, which result in large ranges (Smallwood & Thelander Citation2008).
Methods
Site characteristics
The BPWF (40°43′9″S, 144°41′49″) comprises 37 Vestas V66 turbines (tower height 60 m, blade length 33 m), located on 1524 ha and developed in two stages, with stage 1 (six turbines) completed in August 2002 and stage 2 (an additional 31 turbines) commissioned (all testing completed and the wind farm fully operational) in May 2004. It is located on a flat-topped bluff with a steep coastal cliff to the west (coastal side) and moderately steep slopes to the east, north and south. Approximately 40% of the land is cleared for cattle grazing (with some fodder cropping), with the remainder being high quality remnant vegetation (including closed melaleuca forest and wet coastal shrubby eucalypt forest). This vegetation has been fenced to exclude cattle.
The SBWF (40° 47′8″S, 144° 44′19″) is approximately 3 km south of Bluff Point and comprises 25 V90 wind turbines (45 m long blades, 80 m tower) located on 1410 ha. The SBWF was developed in one stage and fully commissioned in April 2007. It is on less elevated terrain and consists of flat undulating land with isolated rocky outcrops and large consolidated dunes with some small ephemeral fresh water lagoons close to the coast (and to the north of the site). Approximately 60% of the land is cleared for cattle grazing (with some fodder cropping) and the remaining land (comprising coastal swamp forest, coastal heath and pockets of wet coastal shrubby eucalypt forest) is fenced to exclude stock.
Data from two surveys (carcass monitoring and bird utilisation) were used in this study, and covered approximately a 10-year period.
Carcass monitoring surveys
Carcass monitoring surveys were conducted from the commencement of operations (BPWF: August 2002; SBWF: April 2007) until the end of December 2010 when the survey strategy changed. Surveys were conducted throughout the year, twice weekly in spring and autumn, and fortnightly outside these periods (based on regulatory requirements and scavenger trials, Hydro Tasmania unpublished data). From 2006, survey frequency was increased to twice weekly during seasons with what were thought to be heightened Tasmanian wedge-tailed eagle activity (eagle display, 1 June to 31 August and eagle fledging, 15 December to 1 March; see and Appendix 1 for scientific names of birds).
All turbines were surveyed after the completion of stage 1 at the BPWF, but a subsample (27%) of turbines was surveyed during the formal surveys after stage 2 was completed. A subsample of turbines was surveyed at the SBWF (25%) following its completion. However, in 2007 the survey regime was changed, with additional surveys focused on eagles undertaken at all turbines at both sites. The formal surveys were complemented by ad hoc surveys, with all site personnel and visitors required to report bird (and bat) injuries or carcasses found onsite.
Observers searched beneath turbines in concentric circles on foot and using slow-moving 4-wd (quad) bikes from the base of the turbine tower usually out to 100 m (consistent with Hull & Muir Citation2010). The subset of turbines surveyed for all species were surrounded with predator-proof fences to minimise removal by mammalian scavengers and pasture was maintained at a low height to increase visibility. All birds and featherspots (a collection of 10 feathers and/or three flight feathers) were documented and then removed to avoid double-counting.
Bird utilisation surveys
Bird utilisation surveys were conducted during autumn and spring (based on regulatory requirements) at both sites for approximately three years prior to the construction of the wind farms and for approximately three years after the wind farms commenced operation, spanning approximately 10 years ().
Table 1 Seasons and years when bird utilisation surveys were conducted at the Bluff Point and Studland Bay Wind Farms. Shading indicates construction (no monitoring).
Five-minute observations documenting species and numbers of bird movements observed were conducted by experienced observers at nine fixed points on two transects (spaced 250 m apart), which crossed the site on an approximate east–west axis, and nine reference points at each wind farm. As birds were not marked, it was not possible to determine if the same individual was observed more than once during an observation period, hence the metric ‘bird movement’ was used. Approximately 10 replicates of each survey point were conducted during each surveyed season, each year and site. Observations were conducted during daylight hours.
Statistical analyses
The species counts from both survey types were compared between sites to determine if they could be combined, using a Spearman rank test. The species lists from the carcass surveys were then compared to those from the bird utilisation surveys, using a Spearman rank test, to test the null hypothesis that the two were the same.
To ensure that the datasets adequately represented the species onsite, a species accumulation curve was produced to determine if the duration of carcass surveys and bird utilisation surveys was adequate to detect all species. This combined data from both sites.
A classification and regression tree (CART) (Brieman et al. Citation1984) was used to classify species that collided with turbines. The response variable was the binary collided/did not collide. Taking a binary approach, rather than using count data limited the confounding effects of species detectability and susceptibility to scavenging (e.g. larger species are detected more readily and not so readily removed from the observation area by scavengers, Hydro Tasmania unpublished data). Each species present at the sites was grouped using a simple combination of classifiers. The classifiers were family or superfamily (if there was one) and foraging strategy/zone. The latter were allocated to one of the following (based on information in the Handbook of Australian, New Zealand and Antarctic Birds Citation1990–2006):
| • | AP: aerial pursuit of prey (e.g. peregrine falcon or swift); | ||||
| • | AS: aerial searching/ground pursuit or scavenging (e.g. eagle); | ||||
| • | C: canopy searching and feeding (e.g. thornbills); | ||||
| • | G: ground searching and feeding (e.g. geese, some ducks); | ||||
| • | W: water searching and feeding (e.g. some species of duck); | ||||
| • | A: ambush predator, chase prey from a perched location (e.g. some raptors such as goshawk); | ||||
| • | S: shrub searching and feeding (e.g. honeyeaters); | ||||
| • | AW: aerial searching, pursuit in water (e.g. seabirds such as gannets). | ||||
The small size of the dataset limited the utility of CART as a predictive tool in this instance, but it provided useful insight by allowing collision risk at these sites to be described using taxonomical and ecological classifiers.
Spatial analysis
An assessment of broad spatial patterns (comparing open versus vegetated habitats) in the collision data was conducted to determine if there were site characteristics associated with collision risk. The null hypothesis was that no difference would be found in the number of collisions in open compared to vegetated habitats. This could not be conducted at the SBWF due to small sample sizes in the collision data. The number of collisions per turbine circuit was assessed to determine if there were any differences. Each circuit has between five and 10 turbines and are located on different parts of the wind farm, with C and D circuits located within remnant vegetation and the A, B and E circuits predominantly in open paddocks. The vegetation adjacent to E circuit is eucalypt forest, while C and D circuits are in closed melaleuca forest. Again due to small sample sizes, the analysis could not be conducted at the species level, so was assessed by comparing passerines and non-passerines, to allow a broad indication of any spatial variability in where collisions occurred amongst these groups. Chi square tests were used to determine if there were deviations from the expected mortality count after accounting for variation in survey effort.
Results
Carcass monitoring surveys
A total of 1228 surveys were conducted at both sites (901 at the BPWF and 327 at the SBWF), comprising 12,908 turbine visits (this includes all survey types).
A total of 245 bird mortalities were detected during these surveys. Eighteen species of birds were identified at the BPWF and 14 species at the SBWF (). However, there were 82 and 14 featherspots detected at the BPWF and SBWF, respectively, which were not identified to any level (laboratory assessments were not made). The only featherspots that could be identified to any level were those of eagles due to their size and distinctive plumage, and hence have been included in the identified species list.
Table 2 Bird and bat collision records at the Bluff Point and Studland Bay Wind Farms.
Of the carcasses that could be identified to the species level, 80 were non-passerines (of these 19 were raptors) and 33 were passerines at the BPWF, while 23 were non-passerines (of which 10 were raptors) and 13 passerines at the SBWF. Of the identified individuals, 68 were migrants and 45 residents at the BPWF, and 14 migrants and 22 residents at the SBWF.
The lists of species mortalities, ranked by counts, were moderately positively correlated between the two wind farms (r s =0.37, P<0.01), even though they involved different-sized turbines.
Bird utilisation surveys
A total of 545 surveys were conducted at the BPWF (315 during autumn and 230 during spring) and 854 surveys at the SBWF (471 during autumn and 383 spring).
Eighty-five species of birds were observed at the BPWF and 77 at the SBWF (Appendix 1). The majority of movements at both sites were by resident passerines (). At both sites the passerine movements were dominated by forest ravens (with more than 9000 movements), Eurasian skylarks (more than 4000 movements) and black currawongs (more than 2000 movements).
Table 3 Number of movements of birds (categorised by migration and broad taxonomic status) observed during bird utilisation surveys at the Bluff Point and Studland Bay Wind Farms.
Species utilisation counts were strongly correlated between the two wind farms (r s =0.69, P<0.01).
Species accumulation curve
Given that the data for both sites were positively correlated, they were combined for development of the risk profiles, and for species accumulation curves.
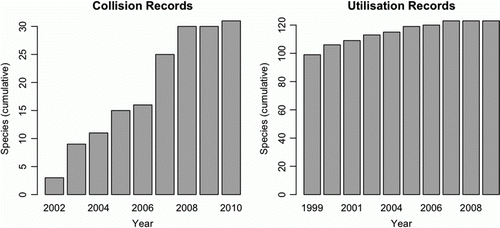
The utilisation study species accumulation curve began to asymptote after eight years from the start of surveys in 1999 (a). The carcass survey species list reflected the increase in survey effort in 2007 (when surveys were added and SBWF was commissioned), but it began to asymptote seven years after the start of BPWF (b). As this measured the addition of newly detected species, it indicated that the lists for both studies were close to complete by the end of 2010.
Figure 2 Species accumulation curves for the Bluff Point wind farm. A is the curve for the carcass monitoring surveys and B is the curve for the utilisation surveys.
Relationship between the collision and utilisation data
The lists of species detected during carcass surveys and bird utilisation surveys were weakly correlated at both sites (BPWF r s =0.14, P ~ 0.1, SBWF r s =0.16, P=0.06). Unsurprisingly, when both sites were combined a similar pattern was found (r s =0.19, P ~ 0.05). This suggests that the frequency of collision is not a simple function of abundance of species (indicated by diurnal surveys) onsite.
Characteristics of species that collided
The CART allocated the species to three terminal nodes, based on a combination of specific species family/superfamily and foraging strategies/zones (). The proportion of collided to not-collided species in each node was 94% (highest risk group), 33% (mid group) and 13% (lower risk group). The families of birds and foraging strategies/zones with the higher, mid and lower probability of colliding are listed in .
Table 4 The families and foraging strategies of birds most likely to collide with turbines.
The brown goshawk, collared sparrowhawk, grey goshawk and orange-bellied parrot were (mis)classified into the colliding group, while the Eurasian skylark and white-throated needletail were (mis)classified into the not-colliding group.
Spatial patterns
Passerines exhibited a significant deviation from the expected mortality counts at turbine circuits. Turbine circuits A and B (in predominantly open paddock) had a higher count than expected (assuming mortalities were distributed equally among turbines), while they had a lower count than expected at turbine chains C, D and E (χ 2 =14, P<0.01, d.f=4). No difference between circuits was found amongst non-passerines (χ 2=8.3, P<0.2, d.f.=4; ).
Table 5 Actual versus expected mortality counts for non-passerines, passerines and all species from the Bluff Point Wind Farm.
Discussion
The species accumulation curve for the BPWF indicated that both types of monitoring have been of sufficient duration to detect most species, particularly those that collided with turbines, even with the addition of turbines in 2004 and changes to survey effort in 2006 and 2007. This is broadly consistent with the findings of Smallwood and Thelander (Citation2008) who found that carcass monitoring needed to be conducted for at least three years to detect all species. The longer time to asymptote in the current study is likely to be an artefact of changing survey effort over time, and the staged development of the wind farm.
The number of species detected during carcass surveys was substantially less than that detected during utilisation surveys, with 21% of the species observed at the BPWF and 18% observed at the SBWF involved in collisions with turbines. Furthermore, the ranking of collision counts per species was weakly correlated with the rank of their abundance in utilisation records. This may be explained in part by the fact that the species most frequently colliding (short-tailed shearwater) was not observed during bird utilisation surveys, as these surveys were conducted during diurnal periods and most seabirds conduct their activity at colonies during nocturnal periods. It does indicate that presence and abundance onsite were poor predictors of collision risk and that specific factors were associated with which birds collided with turbines. This is consistent with overseas studies on raptors, which have recognised that there are specific features associated with collision risk beyond presence onsite (e.g. de Lucas et al. Citation2008).
The Procellariformes, particularly short-tailed shearwaters, were most commonly found in the collision records, for example at the BPWF they comprised 44.7% of records. There are estimated to be over 19 million pairs of short-tailed shearwaters (not including counts of burrows) nesting on islands within about 30 km of these wind farms (Skira et al. Citation1996; Brothers et al. Citation2001). Therefore, there is likely to be a very high number of movements of short-tailed shearwaters in this area, with an unknown proportion moving through the wind farms. The other seabirds recorded in the collision records have been observed on some of the surrounding islands (see Brothers et al. Citation2001; the Atlas of Living Australia http://www.ala.org.au/).
Raptors constituted 17% and 26% of the collision records at the BPWF and SBWF, respectively. Raptors are thought to be at-risk taxa at modern wind farms overseas, but less so than at the old style wind farms such as the Altamont Pass Wind Resource Area which has a predominance of raptor collisions (Strickland et al. Citation2011). Not all species of raptor present at the BPWF and SBWF were involved in collisions, with four from a total of nine species observed onsite involved in collisions, and with three species (all goshawks) being misclassified into the colliding group. This suggests that not all raptor species are of equal risk and it is most likely there are key morphological, ecological and behavioural factors at play, again reinforcing the importance of focusing on the species found to be at risk of collision, not just broad taxonomic groups.
There were distinct families/superfamilies and foraging strategies/zones associated with whether a bird collided with turbines, and a combination of the two that classified collision risk. This combination defined a single CART node that consisted of 94% of species that collided. Species in the families seabirds, raptors, parrots and rosellas, pigeons, pelicans, swifts, robins, larks, white-eyes, flycatchers, and fantails and storm-petrels which undertook the foraging strategies/zones aerial searching, shrub searching and feeding, aerial searching and pursuit in water or canopy searching were most likely to appear in this ‘high risk’ group. Only one family appeared in the high and low risk family allocations (the Petroicidae), indicating that some species in this family undertook foraging strategies/zones that increased their risk of collisions with turbines, while other species in the family did not.
The classifiers family/superfamily and foraging strategy/zone were, however, simplistic, ‘first-cut’ classifiers and overlapped in some respects, which has probably resulted in an over-tuning of the model restricting its utility as a fully predictive tool. However, the results do support the notion that there are particular functional groups associated with collision risk. Studies from overseas sites have identified particular species or functional groups and species-specific behaviours that are linked with collision risk (Osborn et al. Citation2000; Smallwood & Thelander Citation2005; Kuvlesky et al. Citation2007). As this study is the first of its kind in Australia, it is important that further investigations are conducted at other sites and in other habitats to refine the classifiers and to ascertain the level of variability in species collision risk (per de Lucas et al. Citation2008).
The key features of these particular families that make them collision prone may be morphological (relating to their ability to detect the moving turbine blades, e.g. see Martin Citation2011; Martin et al. Citation2012), their agility (ability to avoid a blade once detected), ecological (relating to the food and where and how they obtain it) and behavioural (how they move through the site and avoid turbines; the additional risk of nocturnal flights for species such as short-tailed shearwaters is included in this behavioural feature). While ecological factors potentially place the individuals in the high risk collision zone (i.e. flights through the rotor swept area) with the number of flights through this zone increasing the collision risk (Smallwood & Thelander Citation2005), birds have been found to actively avoid wind turbines, described as a species avoidance rate. Although very few studies have quantified the avoidance rate of species (but see Hull & Muir Citationin press), it is recognised that such rates do exist (see Chamberlain et al. Citation2006) and without which birds would have substantially higher collision rates than are currently found. Why species with a recognised avoidance rate sometimes fail in their avoidance behaviour is unknown and is a key area for future study.
While the findings from this and previous studies suggest that specific morphological and behavioural factors are involved in collision risk, some studies have also identified specific site factors that are also associated with collision risk. In the USA, prey abundance, foraging areas (canyons and ridge crests) and tower type were associated with collision risk; however, a great deal of interspecific variability in these factors was found (Smallwood & Thelander Citation2004). Other studies found the interaction of wind-relief and species-specific and seasonal factors (Barrios & Rodríguez Citation2004); species, turbine height and elevation above sea level in raptors (de Lucas et al. Citation2008); rain and fog (e.g. Osborn et al. Citation2000 and references therein); spatial, temporal, taxonomic and behavioural factors (de Lucas et al. Citation2008); and movement at night to roosting sites (Everaert Citation2008) associated with collision risk. The more evidence obtained, the more it appears that collision risk factors are likely to vary across sites (Thelander & Rugge Citation2000), reinforcing the need for assessments of site variability in Australia.
While it was not possible to conduct a detailed spatial analysis at these sites, coarse-level patterns were found. More passerine collisions were detected at circuits in open areas, compared to circuits within or adjacent to melaleuca vegetation. Given that the formal surveys (i.e. not ad hoc finds) were predominantly conducted at turbines surrounded by predator-proof fences and survey frequency was dictated by scavenger removal rates, the most parsimonious explanation for this pattern is that passerines from the high risk groups were more abundant in the open areas of the wind farm. This may be because they are foraging in the open areas (species such as skylarks) or moving through the open areas at turbine height (species such as silvereyes). Unfortunately, further assessment of spatial patterns could not be tested in the utilisation data as two datasets were not comparable in this respect. These findings highlight the importance of habitat variability, particularly which habitats may have more of the collision-prone bird groups, and so should be considered in risk assessments for new wind farms.
The results from the current study provide an opportunity to compare the actual impact of these operating wind farms to that estimated during risk assessments, given that questions have recently been raised about the adequacy of such assessments (Ferrer et al. Citation2012). The Development Proposal and Environmental Management Plan (DPEMP; Hydro Tasmania Citation2000) conducted for these wind farms identified all species of birds found during carcass surveys, with the exception of seven (Australian gannet, common diving petrel, short-tailed shearwater, white-faced storm petrel, fairy prion, Wilson's storm petrel and grey-backed storm petrel). However, while the short-tailed shearwater was not observed during daytime utilisation surveys, it was recognised as being at possible risk of collision after having been found to collide with wind monitoring towers. Presumably the seabirds were not observed during the utilisation surveys because they entered the wind farms during crepuscular periods or at night (therefore outside the survey periods), especially when local breeders returned to their colonies. This needs to be considered in risk assessments for new wind farm proposals. While the vast majority of species present onsite were documented in the DPEMP, the focus of the risk assessment was threatened species and what were assumed to be at-risk groups. Some of these assumed at-risk groups, such as waterbirds and some migratory passerines, were either found not to collide or not to be in the high risk group.
The analysis in this study was restricted to comparing species that collided with those that did not, in relation to the presence of birds onsite. If a bird utilisation study was designed as part of a collision monitoring study, there would be greater opportunities to explore patterns in the two data sets to provide greater insights into the factors involved in collision risk.
This study identified some of the key factors associated with the likelihood of birds colliding with turbines at these two wind farms. Simple presence onsite was a poor indicator of turbine collision risk; rather, specific families/superfamilies and foraging strategies/zones (which are probably proxies for morphological, ecological and behavioural factors) were associated with turbine collisions. It is important that similar analyses are conducted at wind farms in other habitats and sites in Australia to determine the level of variability and to refine the classifiers for Australian species.
Acknowledgements
The bird utilisation surveys were conducted by Biosis Research, Ecology Partners and Wildspot Consulting, and the carcass searches were conducted by Wildspot Consulting, for which we thank them. We would like to thank Stuart Muir, Stephen Ross and two anonymous referees for comments on the manuscript.
References
- Barrios , L and Rodríguez , A . 2004 . Behavioural and environmental correlates of soaring-bird mortality at on-shore wind turbines . Journal of Applied Ecology , 41 : 72 – 81 . doi: 10.1111/j.1365-2664.2004.00876.x
- Bay K 2010 Updated summary of bird and bat fatalities from wind-energy facilities . In : Savitt Schwartz S ed. Proceedings of the Wind Wildlife Research Meeting VIII. October 2010 , Lakewood Colorado. Prepared for the National Wind Coordinating Collaborative by RESOLVE Inc ., Washington , DC . 10 – 13 .
- Bender , EA , Case , TJ and Gilpin , ME . 1984 . Perturbation experiments in community ecology: theory and practice . Ecology , 65 : 1 – 13 . doi: 10.2307/1939452
- Breiman L , Friedman JH , Olshen RA , Stone CJ 1984 . Classification and regression trees . Belmont , CA , Wadsworth . 358 .
- Brothers N , Pemberton D , Pryor H , Halley V 2001 . Tasmania's Offshore Islands: seabirds and other natural features . Tasmanian Museum and Art Gallery , Hobart , Tasmania .
- Chamberlain , DE , Rehfisch , MR , Fox , AD , Desholm , M and Anthony , SJ . 2006 . The effect of avoidance rates on bird mortality predictions made by wind turbine collision risk models . Ibis , 148 : 198 – 202 . doi: 10.1111/j.1474-919X.2006.00507.x
- de Lucas , M , Janss , GFE , Whitfield , DP and Ferrer , M . 2008 . Collision fatality of raptors in wind farms does not depend on raptor abundance . Journal of Applied Ecology , 45 : 1695 – 1703 . doi: 10.1111/j.1365-2664.2008.01549.x
- Erickson WP , Johnson GD , Strickland MD , Young DP Jr , Sernka KJ , Good RE 2001 . Avian collisions with wind turbines: a summary of existing studies and comparisons with other sources of avian collision mortality in the United States . Western EcoSystems Technology, Inc. prepared for the National Wind Coordinating Collaborative , Washington , DC . 62 .
- Everaert J 2008 . Effecten van windturbines op de fauna in Vlaanderen Onderzoeksresultaten, discussie en aanbevelingen . Effects of turbines on fauna in Flanders Study results, discussion and recommendations . Brussels , Belgium , Instituut voor Natuur – en Bosonderzoek . 174 .
- Ferrer , M , de Lucas , M , Janss , GFE , Casado , E , Muñoz , AR , Bechard , MJ and Calabuig , CP . 2012 . Weak relationship between risk assessment studies and recorded mortality in wind farms . Journal of Applied Ecology , 49 : 38 – 46 . doi: 10.1111/j.1365-2664.2011.02054.x
- Handbook of Australian, New Zealand and Antarctic Birds 1990–2006 . 1–7 . Melbourne , Australia , Oxford University Press .
- Hull , CL and Muir , S . 2010 . Search areas for monitoring bird and bat carcasses at wind farms using a Monte-Carlo model . Australasian Journal of Environmental Management , 17 : 77 – 87 . doi: 10.1080/14486563.2010.9725253
- Hull CL , Muir SC in press. Behavior and turbine avoidance rates of eagles at two wind farms in Tasmania , Australia . Wildlife Society Bulletin.
- Hydro Tasmania 2000 . West Coast Wind Farm Woolnorth , north-west Tasmania Development Proposal and Environmental Management Plan . 1–2 .
- Kuvlesky , WP Jr , Brennan , LA , Morrison , ML , Boydston , KK , Ballard , BM and Bryant , FC . 2007 . Wind energy development and wildlife conservation: challenges and opportunities . Journal of Wildlife Management , 71 : 2487 – 2498 . doi: 10.2193/2007-248
- Langston RHW , Pullan JD 2003 . Windfarms and birds: an analysis of the effects of windfarms on birds, and guidance on environmental assessment criteria and site selection issues . Birldlife International for Bern Convention Directorate of Culture and of Cultural and Natural Heritage , Strasbourg , France . 56 .
- Madders , M and Whitfield , DP . 2006 . Upland raptors and the assessment of wind farm impacts . Ibis , 148 : 43 – 56 . doi: 10.1111/j.1474-919X.2006.00506.x
- Martin , G . 2011 . Understanding bird collisions with man-made objects: a sensory ecology approach . Ibis , 153 : 239 – 254 . doi: 10.1111/j.1474-919X.2011.01117.x
- Martin , G , Portugal , SJ and Murn , CP . 2012 . Visual fields, foraging and collision vulnerability in Gyps vultures . Ibis , 154 : 626 – 631 . doi: 10.1111/j.1474-919X.2012.01227.x
- Osborn , RG , Higgins , KF , Usgaard , RE , Dieter , CD and Neiger , RD . 2000 . Bird mortality associated with wind turbines at the Buffalo Ridge wind resource area, Minnesota . The American Midland Naturalist , 143 : 41 – 53 . doi: 10.1674/0003-0031(2000)143[0041:BMAWWT]2.0.CO;2
- Skira , I , Brothers , NP and Pemberton , D . 1996 . Distribution, abundance and conservation status of short-tailed shearwaters Puffinus tenuirostris in Tasmania, Australia . Marine Ornithology , 24 : 1 – 14 .
- Smales I 2006 . Wind farm collision risk for birds . Cumulative risks for threatened and migratory species. Prepared for the Australian Government Department of the Environment and Heritage by Biosis Research. Project no. 5182 . Canberra , Australia , Department of Environment and Heritage . 225 .
- Smallwood KS , Thelander CG 2004 . Developing methods to reduce bird mortality in the Altamont Pass Wind Resource Area . Final report by BioResource Consultants to the California Energy Commission, Public Interest Energy Research-Environmental Area, Golden, CO . 363 .
- Smallwood KS , Thelander C 2005 . Bird mortality at the Altamont Pass Wind Resource Area March 1998–September 2001 . Report to the National Renewable Energy Laboratory, US Department of Energy , Oak Ridge , TN . 403 .
- Smallwood , KS and Thelander , C . 2008 . Bird mortality in the Altamont Pass Wind Resource Area, California . Journal of Wildlife Management , 72 : 215 – 223 . doi: 10.2193/2007-032
- Strickland MD , Arnett EB , Erickson WP , Johnson DH , Johnson GD , Morrison ML , Shaffer JA , Warren-Hicks W 2011 Comprehensive guide to studying wind energy/wildlife interactions . Prepared for the National Wind Coordinating Collaborative , Washington , DC . 282 .
- Thelander CG , Rugge L 2000 . Examining relationships between bird risk behaviors and fatalities at the Altamont Pass Wind Resource Area: a second year's progress report . In : Proceedings of the National Avian Wind Power Planning Meeting IV , Golden , CO . 5 – 14 .