
ABSTRACT
A sample of feral Hokonui, Campbell Island and Raglan sheep were captured after living feral > 40 years and managed on North and South Island farms alongside domesticated breeds for 28 years to understand relative performance in various traits. Fleece weight was significantly different (P < 0.001) between breeds (Hokonui, 1.48 kg; Campbell Island, 1.50 kg; and Raglan, 2.16 kg), lower than both domesticated Saxon Merino (3.26 kg) and Perendale (3.29 kg) in the North Island. Live weight was significantly different (P < 0.001) between flocks (Hokonui, 39.4 kg; Campbell Island, 38.6 kg; and Raglan, 43.0 kg), but intermediate between Saxon Merino (35.5 kg) and Perendale (51.6 kg). Reproductive rate depended on live weight and was also intermediate between the domesticated breeds, being significantly greater in the South Island (P < 0.001). Decreased fleece weight was common to farmed feral sheep from three different origins, suggesting this trait may have been an advantage during a feral existence.
Introduction
With domestication and the associated selective breeding procedures imposed by humans, it has been possible to modify farmed livestock at both a phenotypic and a genetic level to enable them to perform under intensive farming conditions. In the case of sheep, this has resulted in increased productivity as defined by live weight, fleece weight and reproductive rate. Breeding has also brought about changes in the appearance and performance of the various products derived from sheep that are used by humans such as meat, wool and milk. As a result of selective breeding, different breeds have either evolved or been developed; where a ‘breed’, as used here, is defined in the Shorter Oxford English Dictionary (Stevenson Citation2007) as a line of descendants perpetuating particular hereditary qualities. The use of prophylactic veterinary medications is likely to have reduced selection pressure on animal health traits such as resistance to parasites and pathogens, while more emphasis has been directed towards increasing productivity.
When farmed animals escape to the wild, different animals may succeed and reproduce than a farmer would choose when seeking to develop an animal for production, as humans would no longer be available to assist the feral animal in overcoming any physical or environmental challenges it may encounter. While it is anecdotally assumed that feral animals will revert to the form they had before selection was imposed by humans, this assumes that the animals return to an environment similar to that in which they evolved and that genes have not been lost from the population. Live weight and reproduction of feral sheep while in a feral environment have been reported by Orwin & Whitaker (Citation1984), Réale & Boussès (Citation1999), Réale et al. (Citation2000) and Van Vuren & Bakker (Citation2009). Although fleece growth and loss have been noted in feral environments (Wilson & Orwin Citation1964; Van Vuren & Bakker Citation2009), fleece weight has not previously been reported.
Three flocks of feral sheep were captured from different locations and returned to a common farmed environment, following a feral existence of between 40 and 75 years. These flocks differed in inherent fleece type and associated characteristics sufficiently to consider each flock as a distinct breed. The objective of the study reported here was to determine whether live weight, fleece weight or reproductive rate had permanently changed while the three flocks were feral. If the flocks had changed permanently, did the three breeds respond similarly? For comparison, the same parameters were also measured in two other breeds of sheep which were ancestrally related to the feral flocks, but had meanwhile been managed with human intervention.
Materials and methods
Description of the feral flocks
It has become common practice in New Zealand to name flocks of feral sheep by the name of the location in which they lived. Hence the three flocks included in this study will be described here as Hokonui, Campbell Island and Raglan. Hokonui sheep are considered to have derived from the Saxon Merino, Campbell Island sheep from the Saxon Merino crossed with British long wool breeds and Raglan sheep from Romney (British long wool breed), possibly crossed with other British long wool breeds (Appendices 1 and 2). The three feral breeds were thus representative of one of the three broad types that the New Zealand flock is commonly separated into. The types, based on the mean fibre diameter of their fleeces are: fine wool (< 24.4 µm); medium wool (24.5–31.4 µm); and coarse wool (> 31.5 µm). Fine wool is produced by Merino sheep and coarse wool by sheep from the British long wool breeds. Medium wool is generally produced from a cross between fine and coarse wool breeds. Many aspects related to the growth and production of meat and wool differ between these broad groupings (Ryder & Stephenson Citation1968). The breeds are listed in the text and Tables in order of increasing mean fibre diameter, according to their wool type classification; namely, Hokonui (fine wool), Campbell Island (medium wool) and Raglan (coarse wool).
At the time of capture in 1976, two of the feral flocks were due to be exterminated in their wild habitat on account of damage to the ecosystem in which they were living. These were the ‘Campbell Island’ captured on a subantarctic island (52°33′S, 169°09′E) 590 km south of Stewart Island and ‘Raglan’ captured on the Paritata Peninsula (37°46′S, 174°56′E) in Raglan (Whaingaroa) Harbour 30 km west of Hamilton in the Waikato region. The third flock, ‘Hokonui’, from the Hokonui Hills (45°95′S, 168°62′E) 45 km northeast of Invercargill in the Southland region, remains in a wild state in the habitat from which they were caught (Appendix 1). The foundation stock captured in 1976, and their descendants, were maintained on several Ministry of Agriculture and Fisheries farms with a description of the wool production and reproduction rate of the three flocks during their first 6 years following capture reported by Bigham & Cockrem (Citation1984). A photo of each breed group at the time is shown in . The three flocks were grazed together and managed identically, except during mating when they were kept as separate flocks to retain genetic purity. No analysable production data were collected from the three flocks until 1990 when they were returned to the Whatawhata Research Centre (37°48′S, 175°5′E) in the Waikato region of the North Island. They were then later transferred to the Winchmore Research Station (43°47′S, 171°48′E) in the Canterbury region of the South Island in 2000. All descendants of the foundation stock captured in 1976 and present in 1990 were used in this study. Collection of production data ceased in 2004 and the three flocks were dispersed.
Figure 1. Photos taken in 1982 at the Ruakura Animal Research Station of the descendants of the sheep captured 6 years earlier from flocks that had been grazing in a feral state for in excess of 40 years. A, Hokonui ewes; B, Campbell Island ewes; C, Raglan ewes. Photos: D. H. B. McQueen.

Managing inbreeding
Between 1976 and 1990 each flock was joined with two rams born within that flock. One ram was joined with the ewes for approximately 4 weeks before being replaced by the second ram for approximately 3 weeks. Parentage of each lamb was assessed on the basis of date of birth. In 1990, with concerns for potential inbreeding and the limited pedigree information available, the ewes in each flock were randomly allocated into three age-balanced groups, subsequently referred to as family lines. These ewes retained their family line allocation for their life in the flock, as did subsequent ewes born within each family line. Rams born in one family line (e.g. Line A) were joined with ewes born in one of the other family lines for two seasons (e.g. Line B), and then with ewes in the other family line for two seasons (e.g. Line C) making a 4-year rotational cycle in interbreeding between the three family lines within each flock. No individual rams were ever joined with ewes in the family line in which they were born. A total of nine feral rams were used each year, with one ram joined with each of the three family lines in each of the three flocks. The basis for selecting potential sires is described below. Where possible, each ram was only used for one breeding season.
The ewes in each feral flock were retained for as long as they were deemed to have a constitution sufficient to rear a lamb during the subsequent season, regardless of their family line. The oldest ewe was retained in the flock for 12 production seasons. This protocol, which is not normal farm practice, enabled the size of each feral flock to gradually increase.
Human intervention
The sheep were subjected to common farm management practices to control feeding, reproduction and wool harvesting at the two farms, as described below. While at Whatawhata the three feral flocks were grazed conjointly with approximately 1500 experimental ewes of diverse breeds involved in several separate experiments. The large flock was managed as a single mob and only separated into its component sub-flocks over joining and lambing to facilitate recording of an accurate pedigree for all progeny. At Winchmore, the three feral flocks were combined and run separate from other mobs of experimental sheep, except at joining and lambing when they were separated into individual mating groups. All the ewes were weighed and their live weight recorded post-joining in April/May at Whatawhata and May/June at Winchmore.
Yearling sheep were shorn in September/October at both grazing locations, with the adult ewes being shorn in August (pre-lambing) at Whatawhata using a standard shearing comb. After the flocks were transferred to Winchmore, a ‘winter (or cover)’ shearing comb designed to leave a slightly longer residual fleece (Dabiri et al. Citation1995) was used in 2000 for the first ewe shearing in August. From 2001 onwards the ewes at Winchmore were shorn in December to alleviate the risk of exposing them to late spring storms. Later shearing in December necessitated removal of wool from the belly and breech ‘pre-lambing’ to assist the ewes over lambing and make it easier for the new born lamb to access the udder. Each fleece was weighed and the greasy fleece weight recorded following shearing.
Lambs were individually identified at birth, and their dam, sex and live weight were recorded. All ram lambs were left entire. Lambs born at Whatawhata were weaned, their live weight recorded and they were shorn as a precaution against flystrike in December, whereas lambs born at Winchmore were weaned and their live weight recorded in January and they were shorn in February. Thereafter the lambs were managed as single-sex mobs until their live weight was recorded as yearlings in September/October. The yearlings were then shorn and their greasy fleece weight recorded. A potential sire, plus a reserve, born in each family line were selected on the basis of their live weight at yearling shearing. The remainder of the rams were culled after yearling shearing.
Statistical analysis
A linear mixed model was fitted using residual maximum likelihood (REML) in GenStat (Payne et al. Citation2009), with ewe and year of record as random effects for all continuous variables. Fixed effects associated with flock, grazing location, ewe age and, where appropriate, the number of lambs born and number of lambs weaned, were included.
Recorded greasy fleece weights were adjusted to the greasy fleece weight grown in 365 days, by proportionally adjusting individual fleeces for the interval between the previous shearing and the date of shearing. Further adjustments were made for the seasonal pattern of wool growth (Bigham et al. Citation1978; Sumner et al. Citation1994), the use of a ‘winter’ shearing comb in 2000 and the different shearing policy at Winchmore when the ewes were shorn in December.
Sex, birth date, birth rank, rearing rank, dam age and age at weaning were included as fixed effects for the lamb analyses. Age at yearling live weight was added to the lamb model for analysing yearling live weight data, and interval between lamb shearing and yearling shearing were added to the lamb model for analysing yearling greasy fleece weight.
A measure of the efficiency of wool growth in terms of the kilograms of live weight associated with the production of one kilogram of greasy wool was calculated for ewes as the fleece-free post-join live weight of each ewe, divided by their greasy fleece weight. For yearlings this was calculated as the live weight at yearling shearing divided by their yearling greasy fleece weight. These variates were subjected to a log10 transformation for analysis.
Ewe reproductive performance was quantified as ‘ewe reproductive rate’, the number of lambs weaned (LW) per ewe joined (EJ). This comprised four measured components (Turner Citation1969):
Each of these components can be measured individually as:
where EPL = number of ewes present at lambing, EL = number of ewes lambing and LB = number of lambs born. Fixed effects associated with flock and ewe age were included. Ewe survival, ewe fertility and lamb survival were subjected to a logit transformation for analysis. Ewe survival in this context is only for the reproduction period between joining and lambing, and given the culling procedures in the farming environment outlined above it does not reflect annual survival or ability to survive during a feral existence.
A Merino industry measure of ewe reproductive performance calculated as the total weight of lamb weaned per kilogram of all ewes present at joining, was calculated as LW/EJ multiplied by lamb weaning weight, divided by the post-join live weight of each ewe (Safari et al. Citation2005). The weight of lamb weaned by ewes that did not rear a lamb was deemed to be zero.
Description of the domesticated flocks
Included in the experimental flocks grazed conjointly with the feral flocks at Whatawhata were a flock of Saxon Merino sheep (Sumner & Henderson Citation2013) and a flock of Perendale sheep (Sumner et al. Citation2012). Saxon Merino sheep are of a fine wool type ancestrally related to the Hokonui (Appendix 1), with ancillary data presented in Appendix 2 supporting the conjecture that Campbell Island sheep were established by crossbreeding Merino and British long wool breeds. The Saxon Merino flock was dispersed in 1994. Perendale sheep are a coarse wool type (British long wool breed) phenotypically similar to the Raglan sheep. The Perendale flock was relocated to Winchmore in 1997 and dispersed in 2004.
Live weight, fleece weight and reproductive rate data from the Saxon Merino and Perendale flocks were used to compare with the feral flock data. The flocks are listed in the text and Tables in order of increasing mean fibre diameter. Differences in the designs of the trials reported by Sumner & Henderson (Citation2013), Sumner et al. (Citation2012) and the present study preclude data from the five flocks being combined into a single statistical analysis.
Results
To facilitate a visual comparison between the three feral flocks and the two domesticated flocks, data for the domesticated flocks with an estimate of the standard error of the mean (SEM) derived from Sumner & Henderson (Citation2013) and Sumner et al. (Citation2012) are included in the following Tables reporting the statistical analysis of data from the three feral flocks.
Number of observations
The number of records of lambs born, yearlings weighed and shorn, and ewes joined each year within the five flocks are shown in . Between 1991 and 2004 the Campbell Island flock was larger than the Raglan flock, which was larger than the Hokonui flock. The domesticated Saxon Merino flock was smaller than the three feral flocks and the Perendale flock was considerably larger. Feral ewes that were older than 8 years of age were combined in a ‘> 8’ age group. While it is a common farm practice to cull ewes in productive flocks at 5 or, in some cases, 6 years of age, some ewes in the domesticated flocks were retained beyond this age to conserve genetic resources. In both domesticated flocks, ewes that were older than 5 years of age were combined in a ‘> 5’ age group. The proportional distributions of ewes in each age group within each of the three feral and two domesticated flocks are shown in .
Table 1. Number of records of lambs born, individuals weighed as a yearling the following year and ewes joined each year across the two grazing locations within A, each of the three feral flocks and B, each of the two domesticated flocks that were grazed conjointly with the feral flocks.
Table 2. Proportional distribution (%) of ewe records in each age group at joining across the two grazing locations within A, each of the three feral flocks and B, each of the two domesticated flocks that were grazed conjointly with the feral flocks.
Human intervention to manage the farm resources restricted the size of each flock and precluded a meaningful analysis of death and culling rates over the whole year to indicate comparative longevity between the five flocks.
Ewe live weight and greasy fleece weight
Predicted means for ewe post-join live weight, 365-day greasy fleece weight and post-join live weight divided by their 365-day greasy fleece weight are shown in . Perendale ewes were heavier and Saxon Merino ewes were lighter than the three feral flocks, whereas Raglan ewes were heavier than Hokonui and Campbell Island ewes at both grazing locations. Ewes at Winchmore were heavier than ewes at Whatawhata. The feral ewes grew significantly less wool than either the Perendale or Saxon Merino ewes. Within the feral ewes, the Raglan ewes grew significantly more wool than the Campbell Island and Hokonui ewes. Ewes at Winchmore were heavier, but failed to grow significantly more greasy wool than ewes at Whatawhata. Assuming live weight as an indicator of feed intake, the combined effect of these two production indicators was that Saxon Merino ewes were the most efficient in growing wool per unit of food eaten, with Perendale ewes ranking second and the Raglan ewes being more efficient than the Hokonui and Campbell Island ewes which were similar. The efficiency ratios were not significantly different between the two grazing locations.
Table 3. A, Mean and standard error of difference (SED) of ewe post-join live weight, ewe fleece weight adjusted to 365 days of growth and ewe live weight per unit fleece weight as a measure of the efficiency of live weight farmed to produce a kilogram of wool for the three feral flocks; B, mean and standard error of the mean (SEM) of the same parameters for two domesticated flocks grazed conjointly with the feral flocks. The SEM for the domesticated flocks has been calculated as the SED reported in the original paper divided by √2, as appropriate for groups of similar size within the accuracy of the quoted SED as only one flock from each study are reported here. Bold text indicates significance at P < 0.05.
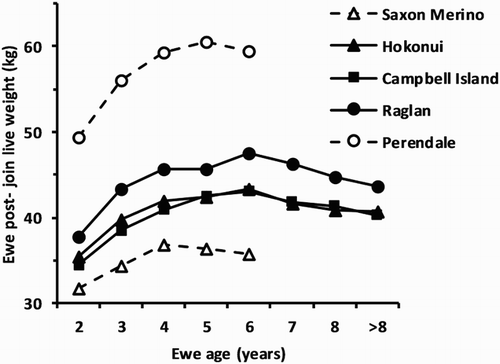
There was a significant curvilinear effect of age on post-join live weight (P < 0.001), when the influences of previous and impending pregnancies and lactations were least within each of the five flocks (). The ewes within each flock, with the possible exception of the Saxon Merino ewes, attained their maximum live weight at 5 to 6 years of age. The variability in mean response tended to increase with age due to the reducing number of ewes in the older age groups ().
Figure 2. Effect of ewe age on mean live weight post-joining in April/May pooled over grazing locations for the three feral flocks, and the two domesticated flocks: Saxon Merino (Sumner & Henderson Citation2013; R. M. W. Sumner, unpubl. data); and Perendale (Sumner et al. Citation2012) grazed conjointly with the feral flocks. Standard error of difference range = 0.6–2.8 kg.
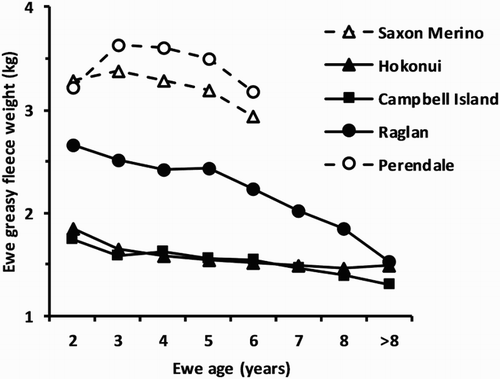
Greasy fleece weight of each of the feral flocks progressively decreased from 2 years of age at both grazing locations, with the rate of reduction of the Raglan sheep being greater than for the other two flocks which were similar (). The greasy fleece weight of the domesticated flocks appeared to plateau at 3 years of age, due to an artefact of the shorter period of growth of fleeces harvested from 2-year-old ewes, compared with adult ewes. Two-year-old ewes were previously shorn as yearlings at a different time to the adult ewes. As with post-join live weight, the variability in mean response for greasy fleece weight tended to increase with advancing age ().
Figure 3. Effect of ewe age on mean greasy fleece weight adjusted to 365 days growth pooled over grazing locations for the three feral flocks, and the recorded fleece weight for the two domesticated flocks: Saxon Merino (Sumner & Henderson Citation2013; R. M. W. Sumner, unpubl. data); and Perendale (Sumner et al. Citation2012) grazed conjointly with the feral flocks. Values for 2-year-old ewes are not comparable between the feral and domesticated flocks. Standard error of difference range = 0.07–0.24 kg.
Ewe reproductive performance
Predicted means for ewe reproductive rate (LW/EJ), ewe survival (EPL/EJ), ewe fertility (EL/EPL), litter size (LB/EL) and lamb survival (LW/LB) within each of the five flocks are shown in .
Table 4. A, Mean and standard error of difference (SED) of reproductive performance for the three feral flocks; B, mean and standard error of the mean (SEM) of the same parameters for two domesticated flocks grazed conjointly with the feral flocks. The SEM for the domesticated flocks has been calculated as the SED reported in the original paper divided by √2, as appropriate for groups of similar size within the accuracy of the quoted SED as only one flock from each study are reported here. Bold text indicates significance at P < 0.05.
Perendale ewes had a higher reproductive rate than Campbell Island and Raglan ewes, which had a higher reproductive rate than Hokonui ewes, with the Saxon Merino having the lowest reproductive rate. Reproductive rate was significantly higher at Winchmore than at Whatawhata, principally due to a heavier post-join live weight in all flocks at Winchmore. Although litter size within the three feral flocks did not differ significantly at Whatawhata, there was a significant flock effect at Winchmore, where the litter size of the Hokonui ewes exceeded that of the Raglan ewes, which exceeded that of the Campbell Island ewes. Litter size in the Perendale ewes exceeded that in the feral flocks, while litter size in the Saxon Merino was less than in the feral flocks. Lamb survival within the Hokonui flock was significantly lower than that of the other two feral flocks, which were not significantly different but similar to the Perendale flock. The Saxon Merino had the lowest recorded lamb survival. There was no significant difference in ewe survival between the three feral flocks at, or between, locations, with both the Saxon Merino and Perendale having a similar recorded ewe survival.
Lamb performance
Predicted means for lamb birth weight and lamb weaning weight within each of the feral and domesticated flocks are shown in . Perendale lambs were the heaviest at birth, with the ranking for the other breeds being Raglan, Saxon Merino, Campbell Island and Hokonui. While the Perendale lambs remained heavier than the other flocks at weaning, the ranking for the other breeds was Raglan, Hokonui and Campbell Island, which were similar, with Saxon Merino being lightest. The Hokonui lambs gained more weight up to weaning than the Campbell Island lambs and the Saxon Merino lambs gained less weight than the other flocks. Ram lambs within each breed were heavier than ewe lambs at birth and at weaning, at both locations. Lamb birth and lamb weaning weights were not significantly different between the two grazing locations.
Table 5. A, Mean and standard error of difference (SED) of lamb birth weight and lamb weaning weight adjusted for age at weaning, yearling live weight adjusted for age at weighing, yearling wool production adjusted for age at shearing and adjusted yearling live weight per unit of adjusted yearling fleece weight as a measure of the efficiency of live weight farmed to produce a kilogram of wool for the three feral flocks; B, mean and standard error of the mean (SEM) of the same parameters for two domesticated flocks grazed conjointly with the feral flocks. The SEM for the domesticated flocks has been calculated as the SED reported in the original paper divided by √2, as appropriate for groups of similar size within the accuracy of the quoted SED as only one flock from each study are reported here. Bold text indicates significance at P < 0.05. Italic text indicates significance between P = 0.05 and P = 0.10.
Predicted means for total weight of lambs born per ewe, total weight of lambs weaned per ewe and total weight of lambs weaned per kilogram of live weight at joining within each of the feral and domesticated flocks are shown in . As a consequence of more twins being born to the Hokonui ewes at Winchmore, there was a significant location by flock effect for each of these derived characteristics. A similar effect was also reported for the Perendale (Sumner et al. Citation2012).
Table 6. A, Mean and standard error of difference (SED) of total weight of lambs born per ewe lambing adjusted for sex, total weight of lambs adjusted for sex weaned per ewe weaning lambs and total weight of lambs adjusted for sex weaned per unit of ewe post-join live weight for the three feral flocks; B, mean and standard error of the mean (SEM) of the same parameters for two domesticated flocks grazed conjointly with the feral flocks. The SEM for the domesticated flocks has been calculated as the SED reported in the original paper divided by √2, as appropriate for groups of similar size within the accuracy of the quoted SED as only one flock from each study are reported here. Bold text indicates significance at P < 0.05. Italic text indicates significance between P = 0.05 and P = 0.10.
The ranking of total weight of lambs born per ewe at Whatawhata was Perendale, Raglan, Saxon Merino, followed by Campbell Island and Hokonui which were not significantly different. By weaning, the ranking for total weight of lambs weaned per ewe at Whatawhata changed to Perendale, Raglan, Campbell Island and Hokonui which were not significantly different, with Saxon Merino sheep having the lowest total weaning weight. While Perendale sheep at Winchmore also had the highest total weight of lambs born per ewe and total weight of lambs weaned per ewe, Hokonui and Raglan sheep were not significantly different, with Campbell Island sheep being the least productive.
When considered as an industry measure of the efficiency of lamb production expressed as the total weight of lamb weaned per kilogram of all ewes present at joining, the ranking at Whatawhata was for the Perendale ewes to be the most efficient, followed by the Campbell Island ewes and then the Raglan, Hokonui and Saxon Merino ewes which were not significantly different. The Perendale ewes were also the most efficient at producing lambs at Winchmore, but the Hokonui, Campbell Island and Raglan ewes were not significantly different from each other.
Yearling live weight and greasy fleece weight
Predicted means for yearling live weight and yearling greasy fleece weight within each of the feral and domesticated flocks are shown in . Perendale yearlings were heavier than Raglan and Hokonui yearlings, which were not significantly different, with the Campbell Island yearlings being lightest and the Saxon Merino yearlings being intermediate between the Raglan and Hokonui, and Campbell Island yearlings. Yearling rams were significantly heavier than yearling ewes. Saxon Merino yearlings had a heavier greasy fleece weight than Perendale yearlings which in turn had a heavier greasy fleece weight than the feral yearlings. While there was no significant difference in yearling greasy fleece weight of the feral flocks between the two locations, the Raglan yearlings grew significantly more greasy wool than the Campbell Island yearlings, with the Hokonui yearlings being intermediate. Assuming yearling live weight as an indicator of feed intake, the combined effect of these two production indicators was that yearling Saxon Merino were the most efficient at growing greasy wool per unit of feed eaten. Yearling Perendale and Raglan sheep were similar and less efficient than Saxon yearlings, but more efficient than the yearling Hokonui and Campbell Island sheep which were not significantly different.
Discussion
All animals are continually subjected to selection pressure mediated via the continuously changing environment in which they live. Such changes tend to be subtle and difficult to measure in the short term. Although the time that these feral flocks spent in a feral state was relatively short in evolutionary terms, the recorded data indicate a measureable effect on their productivity relative to that of two continuously farmed ‘ancestrally related’ flocks.
There is an underlying epidemiological problem in a study such as that reported here. The challenges that feral sheep may have been exposed to in the wild were potentially different to those encountered during the farming operations before the sheep were released into the wild, and certainly after they were captured and subjected to farming conditions. It is also possible that, by chance, the sheep that originally became feral, or the small groups that were recaptured and then farmed, were genetically different in some way to the larger populations they were derived from at the respective times, but given similar trends in three populations we consider this unlikely. It is therefore reasonable to assume that because the sheep survived while feral, they may have also developed some other physiological or behavioural traits beyond a change in their live weight, fleece weight or reproductive performance that enabled them to withstand various challenges.
There are four key reference points in considering the relative performance of descendants of feral flocks captured after living in a feral state for a significant period of time. The first is relativity between the flocks when released into the wild, which was not possible to determine in this study. The second is the relativity between the flocks at the time of capture, but the small numbers of animals that were captured from each of the three feral populations limited the number of observations and there were no effective control flocks for comparison during this period (Bigham & Cockrem Citation1982, Citation1984). The third is their relativity to domesticated flocks of a breed with similar characteristics farmed continuously throughout the period the flocks were living in a feral state. The data from two ‘domesticated’ flocks provided a practical basis to compare and contrast. The fourth is the relativity between the flocks after being farmed for several generations following capture.
Live weight and fleece weight
International studies have reported that live weight of feral flocks in their feral habitat was lighter than comparative domesticated breeds (Réale & Boussès Citation1999; Réale et al. Citation2000; van Vuren & Bakker Citation2009). A small number of feral sheep were captured from Santa Cruz Island and raised on farms, and van Vuren & Bakker (Citation2009) reported that rams became heavier than those feral on the island, while ewes reached a similar weight. However, these were from the initial capture and not self-replacing flocks like those reported here. Pearson et al. (Citation1999) reported that Raglan ewes were heavier than the Hokonui and Campbell Island ewes 14 years after capture in 1976, and this was supported here. In the current study, adult ewes of the three feral flocks were heavier than the Saxon Merino but lighter than the Perendale sheep farmed under the same conditions. In contrast to other authors, we suggest that these flocks at least are intermediate and not smaller than comparative breeds.
Wool production of the feral flocks, on the other hand, was considerably less than either the Saxon Merino or Perendale flocks. Yearling fleece weight was markedly lower in feral than domesticated breeds, and likewise in adult ewes at both North Island and South Island locations. Campbell Island yearlings and adults in particular grew less than half the wool of either domesticated breed. Wool growth greater than necessary to maintain body temperature is effectively a waste of nutrients for a feral sheep. Furthermore, a short fleece can help maintain thermal equilibrium, while additional growth may hinder survival. The additional weight and volume of the fleece, particularly when wet, will require additional energy expenditure by the sheep for locomotion. In association with limited food availability this may result in inanition and death, particularly in combination with the fleece becoming entangled on trees and scrub as sheep forage in forest or scrubland rather than pasture. If the fleece grows continuously without moulting, this effect would compound over time as the fleece became longer. We might expect this would generate selection pressure for a sheep to shed its fleece as occurred before humans selected sheep for continuous wool growth (Ryder & Stephenson Citation1968). Indeed, in a flock of Wiltshire sheep, selection for increased fleece weight reduced annual fleece moulting normally observed in that breed (O’Connell et al. Citation2012). However, under farmed conditions none of the feral sheep studied here exhibited moulting and there are no reports of moulting in Hokonui or Raglan sheep when grazing in the feral state. This occurs despite the fact that all three flocks exhibit seasonal patterns of wool growth and follicle activity (Pearson et al. Citation1999). Wilson & Orwin (Citation1964) reported that, among 30 sheep that were shot on Campbell Island, all but one had lost at least part of their fleece. They noted a reduction in fibre diameter in the wool grown during the late autumn to early spring period when forage would have been scarce. Wool loss without any sign of seasonal moulting has also been reported from Santa Cruz Island sheep (Van Vuren & Bakker Citation2009). A sudden marked reduction in fibre diameter reduces the tensile strength of wool staples (Ryder & Stephenson Citation1968), enabling parts of the fleece to be rubbed off as the sheep traverse through vegetation rather than moulting per se. Loss of the fleece in this manner could reduce any selective advantage for increased moulting during a feral existence.
Simplistically, the live weight, greasy fleece weight and reproductive rate of sheep are each related to feed intake and the nutritive value of that feed. Thus, when planning the grazing management of sheep, farmers commonly use live weight, or desired growth rate, as an indicator of an appropriate feeding level. Hence the live weight and greasy wool production of both yearling sheep and adult ewes can be used to estimate the efficiency of greasy wool growth, measured as kilograms of live weight farmed over winter to produce one kilogram of greasy wool. There was a similar across-breed ranking for both sexes of yearlings and adult ewes. The feral sheep exhibited a lower efficiency of wool growth than the domesticated breeds, with a ranking of Saxon Merino, Perendale and Raglan, followed by Hokonui and Campbell Island, which were not significantly different and were the least efficient of all the flocks ( and ). Where commercial farmers wish to maximise the production of wool per unit of live weight, and hence per unit of dry matter eaten, farmed feral sheep suggest a lower efficiency of wool production might suit a feral existence. This trend could possibly be interpreted as an advantage for a reduced fleece weight sufficient to maintain insulation while sparing nutrients for other purposes in sheep exposed to long cold winters with limited feed availability. The mechanism underlying this process within the three feral flocks could be the reverse of the process associated with domestication and selection, and has been shown to result from a reduction in total wool follicle density and increased fibre seasonality associated with reduced follicle activity during winter (Pearson et al. Citation1999). Published international reports associated with feral sheep have not reported fleece weight, and this is disappointing since it is easy to measure.
Age trends in live weight and wool production
Similar to international reports (Réale & Boussès Citation1999; Van Vuren & Bakker Citation2009), sexual dimorphism was evident in all the feral flocks, with rams significantly heavier at birth, weaning and again at yearling, though the sexes were separated and grazed separately after weaning. With the exception of the Raglan ram lambs, all feral lambs were lighter at birth than either Saxon Merino or Perendale lambs (); however, by weaning the Saxon Merino were the lightest and the feral lambs were lighter than the Perendale. The Campbell Island yearling ewes were the lightest, and the Saxon Merino had regained their position around the middle of the range in live weight, with the Perendales clearly heavier as yearlings at Winchmore only. Having reached 25 kg to 29 kg at Whatawhata (), yearling feral ewes attained live weights between 38 kg and 43 kg as adult ewes, (), whereas Saxon Merino ewes grew from heavier yearlings (29.8 kg) to become lighter as adults (35.5 kg) in that environment, while the Perendale ewes were much heavier at both ages on the North Island farm (34.6 kg and 51.6 kg, respectively). This suggests different growth paths for the feral and domesticated breeds as noted by van Vuren & Bakker (Citation2009), yet in our results the Saxon Merino were not ultimately heavier and this difference may simply be choice of the breeds used for comparison in the two studies. Van Vuren & Bakker (Citation2009) also made the interesting observation that the adult body mass of other insular feral sheep populations derived from Merino converged towards a similar body size, ranging from 47 kg to 53 kg for rams and 34 kg to 40 kg for ewes. In the present study, live weight of both the Hokonui and Campbell Island ewes only slightly exceeded this range under the favourable lowland farming conditions at Winchmore.
In this study, post-join live weight of the feral flocks showed a curvilinear relationship that increased to a plateau at 5 to 6 years of age before declining, with the domesticated flocks beginning to decline a year earlier (). The proportional decline in live weight with age in the feral flocks aligned closely with that reported for farmed Merino (Brown et al. Citation1966) and Perendale sheep (Sumner et al. Citation2012), albeit at a lower mean live weight. This response may be associated with the link between body size, metabolic rate and longevity (Hulbert et al. Citation2007). In contrast Santa Cruz Island sheep which remained feral reached mature live weight faster than those that were farmed, with no indication of a significant decline in live weight up to 9 years of age (Van Vuren & Bakker Citation2009). Interestingly, wild-derived versus laboratory-adapted mice show similar differences (Miller et al. Citation2002).
Quoted values of greasy fleece weight for the feral and domesticated flocks at 2 years of age are not comparable. Fleeces shorn from the 2-year-old sheep grew over a 9 to 10 month period, rather than 12 months for the other age groups. Ignoring 2-year-old fleece weight, the greasy fleece weight of the domesticated flocks exhibited a curvilinear decline after 3 to 4 years of age, an effect also reported in a large flock of grazed Merino sheep (Brown et al. Citation1966). The rate of decline in Raglan sheep was greater than in the other two feral flocks (). The mean response in both live weight and greasy fleece weight was more variable at older ages due to the declining number of ewes in each increasing age group (). Clearly the domesticated flocks produced more wool, but the decrease in fleece weight with aging during the lifetime of all the flocks suggests that this characteristic may be a natural trend in the productivity of aging wool follicles which has been unaffected by domestication.
Reproductive performance in feral flocks
The limited data collected following capture suggested that reproductive rate of the three feral flocks was similar to that expected in commercial flocks at that time (Bigham & Cockrem Citation1984). This was reinforced in the current study by the lack of a significant effect on the key components of reproductive rate (). Lamb survival in farmed sheep is a complicated interaction that has been widely researched (reviewed in Hinch & Brien Citation2014). Lambs from singleton births tend to be more likely to survive than those from twin births, which may explain some of the low survival rates of Hokonui lambs at Winchmore. Birthweight within a breed can be very important to survival, but neonates of low birth weight might die from hypothermia, while those with high birth weight may perish from complications caused by dystocia, such that there is a parabolic relationship between birthweight and survival (Hinch et al. Citation1985). Dystocia is unlikely to occur in small lambs from large ewes, for example a 42 kg Hokonui ewe delivering a 3.6 kg of lamb compared to a 35.5 kg Saxon Merino ewe delivering a 3.7 kg lamb, but if the Hokonui ewe were delivering twins the chances of survival might be quite different. Maternal behaviour, dam to lamb bonding and neonatal behaviour have also been widely investigated (reviewed in Dwyer & Lawrence Citation2005), but how these traits were expressed during a feral existence would likely bear no resemblance to what might have occurred to the farmed feral sheep on research farms where the lambs were tagged and weighed at birth, particularly in comparison with domesticated breeds that may have been selected on the basis of this interaction with humans for many years. Saxon Merino sheep weaned fewer lambs per ewe joined than all other flocks, while Perendale sheep weaned more lambs per ewe joined, attributable to litter size and lamb survival. The lower lamb survival of the Saxon Merino and Hokonui flocks is a trend exhibited by other fine wool Merino flocks, associated with lighter birth weight (Hatcher et al. Citation2009). The total weight of lambs born per ewe lambing and the total weight of those lambs again at weaning () are other indices of reproduction used in farming, incorporating litter size and lamb weight. Although the Hokonui and Campbell Island ranked lowest at birth, they had outperformed the Saxon Merino in growth, if not survival (), at weaning.
It is of interest from an adaptive perspective that the feral flocks retained the ability to produce more lambs when offered more feed over the joining period (Smith Citation1991). Both ewe fertility and litter size improved in all three feral flocks when shifted from hard hill country conditions at Whatawhata to easy lowland conditions at Winchmore. Even though there was little difference between the feral flocks in the change in post-join live weight, the Hokonui ewes exhibited the most pronounced response in reproductive traits following relocation, while the Raglan exhibited an intermediate response. The reproductive rate of the Campbell Island ewes was the least sensitive to improved nutrition under lowland conditions. The environments in which the Hokonui and Campbell Island sheep lived during their feral state were arguably more severe than that experienced by the Raglan sheep in the Waikato; however, the only feral flock by location interaction of practical significance was that related to the litter size of the Hokonui ewes when relocated from Whatawhata to Winchmore.
Which traits might suit a feral environment?
A feral environment is harsh, with prevailing weather resulting in fluctuating feed supplies that affect mortality rates and population dynamics as seen in Soay sheep on the island of Hirta in the Outer Hebrides of Scotland (Owen-Smith & Marshal Citation2010). The heritability of body mass was considered a potential reason for the reduction in size of the Santa Cruz Island sheep (Van Vuren & Bakker Citation2009). However, in contrast to the observation of Van Vuren & Bakker (Citation2009), the farmed feral sheep studied here were intermediate in live weight between the domesticated breeds we used for comparison. The sheep used here were also intermediate in reproductive rate compared to domesticated flocks but they retained the fertility response to improved nutrition. Since reproductive rate has a low heritability (h2 = 0.01; Fogarty Citation1995) and is associated with live weight, changes in reproductive rate in feral populations may be less likely. Indeed, low heritability of reproductive traits was not considered as a reason for a lack of response in reproductive traits in the feral flock from Ile Longue (Réale & Boussès Citation1999; Réale et al. Citation2000). Greasy fleece weight is highly heritable (h2 = 0.45; Fogarty Citation1995); feral sheep from all three environments produced lower fleece weights than either domesticated breed they were farmed with, and we suggest that low fleece weight may have been an advantage during their feral existence.
Conclusion
Overall, the results of this study showed that three flocks of sheep that existed in a feral state for some 15 generations before being captured and subsequently subjected to normal sheep farming practices, had a lower fleece weight and an intermediate live weight and reproductive rate relative to two ancestrally-related domesticated breeds that were farmed continuously throughout the period that the feral sheep existed in a feral state. Each feral flock was representative of a different sheep type, had a different geographical origin, and yet exhibited a similar performance in most of the traits measured while they were farmed. These findings suggest that reduced fleece weight may be advantageous to feral sheep.
Acknowledgements
To Drs Murray Bigham and the late Francis Cockrem who had the foresight to preserve these sheep flocks for scientific study, and to field and technical staff at Whatawhata Research Centre and Winchmore Research Station for assistance in management of the sheep and collection of field data. Associate Editor: Associate Professor Adrian Paterson.
Disclosure statement
No potential conflict of interest was reported by the authors.
References
- Bell BD, Taylor RH. 1970. Expedition’s study of the wild sheep of Campbell Island. Forest and Bird. 187:6–10.
- Bigham ML, Cockrem FRM. 1982. Feral sheep: have they any value. New Zealand Journal of Agriculture. 144:3–4.
- Bigham ML, Cockrem FRM. 1984. Observations on wool growth and other characteristics of feral sheep. In: Blair HT, editor. Coloured sheep and their products. Proceedings of the World Congress on Coloured Sheep; 1984 Jan 20–31; New Zealand: Lincoln College and Massey University; p. 302–305.
- Bigham ML, Sumner RMW, Elliott KH. 1978. Seasonal wool production of Romney, Coopworth, Perendale, Cheviot and Corriedale wethers. New Zealand Journal of Agricultural Research. 21:377–382. doi: 10.1080/00288233.1978.10427422
- Brown GH, Turner HN, Young SSY, Dolling CHS. 1966. Vital statistics for an experimental flock of Merino sheep. III. Factors affecting wool and body characteristics, including the effect of age of ewe and its possible interaction with method of selection. Australian Journal of Agricultural Research. 17:557–581. doi: 10.1071/AR9660557
- Dabiri N, Holmes CW, McCutcheon SN, Parker WJ, Morris ST. 1995. Resistance to cold stress in sheep shorn by cover comb or standard comb. Animal Science. 60:451–456. doi: 10.1017/S1357729800013321
- Department of Conservation. 2014. Offshore islands and conservation; [cited 2014 June 11]. Available from: www.doc.govt.nz/conservation/land-and-freshwater/offshore-islands/new-zealands-subantarctic-islands/campbell-island/
- Dwyer CM, Lawrence AB. 2005. A review of the behavioural and physiological adaptations of hill and lowland breeds of sheep that favour lamb survival. Applied Animal Behaviour Science. 92:235–260. doi: 10.1016/j.applanim.2005.05.010
- Fogarty NM. 1995. Genetic parameters for live weight, fat and muscle measurements, wool production and reproduction in sheep: a review. Animal Breeding Abstracts. 63:101–143.
- Hatcher S, Atkins KD, Safari E. 2009. Phenotypic aspects of lamb survival in Australian Merino sheep. Journal of Animal Science. 87:2781–2790. doi: 10.2527/jas.2008-1547
- Hinch GN, Brien F. 2014. Lamb survival in Australian flocks: a review. Animal Production Science. 54:656–666. doi: 10.1071/AN13236
- Hinch GN, Crosbie SF, Kelly RW, Owens JL, David GH. 1985. Influence of birth weight and litter size on lamb survival in high fecundity Booroola-Merino crossbred flocks. New Zealand Journal of Agricultural Research. 28:31–38. doi: 10.1080/00288233.1985.10426996
- Hulbert AJ, Pamplona R, Buffenstein R, Buttemer WA. 2007. Life and death: metabolic rate, membrane composition and life span of animals. Physiological Reviews. 87:1175–1213. doi:10.1152/phyrev.00047.2006
- Johns A. 1980. Hokonui wild Merinos a unique gene pool. New Zealand Farmer. 101:156–158.
- Kerr IS. 1976. Campbell Island, a history. Wellington: AH & AW Reed. 182 p..
- McKenzie GW, Abbott J, Zhou H, Fang Q, Merrick N, Forrest RH, Sedcole JR, Hickford JG. 2010. Genetic diversity of selected genes that are potentially economically important in feral sheep of New Zealand. Genetics Selection Evolution. 42:43–50. doi: 10.1186/1297-9686-42-43
- Meurk CD. 1982. Regeneration of subantarctic plants on Campbell Island following exclusion of sheep. New Zealand Journal of Ecology. 5:51–58.
- Miller RA, Harper JM, Dusko RC, Durkee SJ, Austad SN. 2002. Longer life spans and delayed maturation in wild-derived mice. Experimental Biology and Medicine. 227:500–508. doi: 10.1177/153537020222700715
- O’Connell D, Scobie DR, Hickey SM, Sumner RMW, Pearson AJ. 2012. Selection for yearling fleece weight and its effect on fleece shedding in New Zealand Wiltshire sheep. Animal Production Science. 52:456–462. doi:10.1071/AN11281.
- Orwin DFG, Whitaker AH. 1984. Feral sheep (Ovis aries L.) of Arapawa Island, Marlborough Sounds, and a comparison of their wool characteristics with those of four other feral flocks in New Zealand. New Zealand Journal of Zoology. 11:201–224. doi: 10.1080/03014223.1984.10423759
- Owen-Smith N, Marshal JP. 2010. Definitive case studies. In: Owen-Smith N, editor. Dynamics of large herbivore populations in changing environments: towards appropriate models. Chichester: Wiley-Blackwell; p. 5–8.
- Payne RW, Murray DA, Harding SA, Baird DB, Soutar DM. 2009. GenStat for Windows, 12th edition. Introduction. Hemel Hempstead: VSN International. 256 p.; [cited 2016 Nov 8]. Available from: http://www.vsni.co.uk/downloads/genstat/release12/doc/introguide.pdf
- Pearson AJ, Ashby MG, Nixon AJ, Sumner RMW. 1999. Seasonal wool growth patterns and the follicle and fibre characteristics of New Zealand feral sheep. Proceedings of the New Zealand Society of Animal Production; 1999 June 28–July 1; Mosgiel, New Zealand; 59; p. 34–38.
- Réale D, Boussès P. 1999. Effects of summer and winter birth on growth of lambs in a population of feral sheep. Journal of Mammalogy. 80:1028–1037. doi: 10.2307/1383273
- Réale D, Boussès P, Pisanu B, Chapuis J. 2000. Biannual reproductive cycle in the Kerguelen feral sheep population. Journal of Mammalogy. 81:169–178. doi: 10.1644/1545-1542(2000)081<0169:BRCITK>2.0.CO;2
- Ryder ML, Stephenson SK. 1968. Wool growth. London: Academic Press. 805 p.
- Safari E, Fogarty NM, Gilmour AR. 2005. A review of genetic parameter estimates for wool, growth, meat and reproduction traits in sheep. Livestock Production Science. 92:271–289. doi: 10.1016/j.livprodsci.2004.09.003
- Smith JF. 1991. A review of recent developments on the effect of nutrition on ovulation rate (the flushing effect) with particular reference to research at Ruakura. Proceedings of the New Zealand Society of Animal Production; 1991 Feb 11–15; Dunedin, New Zealand: Otago University; 51; p. 15–23.
- Stevenson A. 2007. Shorter Oxford English dictionary on historical principles. London: Oxford University Press. 3742p.
- Sumner RMW, Clarke JN, Pearson AJ, Speedy PM. 1994. Repeatability of seasonal wool growth in Merino sheep. Proceedings of the New Zealand Society of Animal Production; 1994 Feb 8–11; Palmerston North, New Zealand: Massey University; 54; p. 163–165.
- Sumner RMW, Forrest RHJ, Zhou H, Henderson HV, Hickford JGH. 2013. Association of the KRT33A (formerly KRT1.2) gene with live-weight and wool characteristics in yearling Perendale sheep. Proceedings of the New Zealand Society of Animal Production; 2013 Jul 2–4; Hamilton, New Zealand; 73; p. 158–164.
- Sumner RMW, Henderson HV. 2013. Relative performance of three strains of fine-woolled Merino sheep farmed on Waikato hill country. Proceedings of the New Zealand Society of Animal Production; 2013 Jul 2–4; Hamilton, New Zealand; 73; p. 136–142.
- Sumner RMW, Scobie DR, O’Connell D, Henderson HV. 2012. Relative performance of Wiltshire and Perendale sheep. Proceedings of the New Zealand Society of Animal Production; 2012 Jul 2–5; Canterbury, New Zealand: Lincoln University; 72; p. 28–34.
- Turner HN. 1969. Genetic improvement of reproductive rate in sheep. Animal Breeding Abstracts. 37:545–563.
- UNESCO. 2014. The World Heritage Convention; [cited 2014 Jun 11]. Available from: whc.unesco.org/en/list/877
- Van Vuren DH, Bakker VJ. 2009. Rapid morphological change in an insular population of feral sheep. Journal of Zoology. 277:221–231. doi: 10.1111/j.1469-7998.2008.00531.x
- Wilson PR, Orwin DFG. 1964. The sheep population of Campbell Island. New Zealand Journal of Science. 7:460–490.
AppendicesAppendix 1. History of the three feral flocks
Hokonui
Currently a population of sheep remain unattended in the Hokonui Hills (45°95′S, 168°62′E) 45 km northeast of Invercargill. These sheep are believed to be of Merino origin, released into the wild during the 1860s. The predominant Merino at the time was the fine wool Saxon type (Johns Citation1980). Six rams and five ewes were caught in the Hokonui Hills in 1976 and transferred to the Whatawhata Research Centre near Hamilton.
Campbell Island
Campbell Island (52°33′S, 169°09′E), a subantarctic island 590 km south of Stewart Island, was discovered in 1810. It is a hilly remnant of an eroded volcanic cone with an area of 112.7 km2 being indented by many narrow inlets. The largest inlet nearly bisects the island. There were several introductions of sheep, and a few cattle, to the island beginning in 1895 under a pastoral grazing lease issued to James Gordon by the New Zealand government. The lease was transferred to Captain W. H. Tucker in 1901, with a few shepherds living on the island (Kerr Citation1976). Three importations of 1000 ewes each were undertaken between 1901 and 1902. They were Merino × British long wool sheep purchased in Southland, with the Merino probably being a fine wool Saxon type (Wilson & Orwin Citation1964; Kerr Citation1976). Although farming conditions were extensive and difficult in the harsh climate, the sheep population reached a peak of 6800 adults in 1916. Thereafter the number gradually dwindled as the most palatable plants were eaten out. Farming on the island collapsed in 1931 due to unreliable transport and the economic effects of the Great Depression. The remaining settlers vacated the island leaving approximately 4000 sheep and 30 cattle to run wild (Kerr Citation1976). In 1970 the island was fenced into two equal sized blocks at its narrowest point and all but an estimated 15 of the 1300 sheep cut off to the north of the fence, were shot (Bell & Taylor Citation1970). The two halves of the island were then used for an ecological study of the effect of grazing by sheep on subantarctic flora (Meurk Citation1982), as proposed by Wilson & Orwin (Citation1964). In February 1976 four mixed age rams and six mixed age ewes were caught on Campbell Island () and transported to the Whatawhata Research Centre. The remaining sheep on Campbell Island were systematically culled to protect the natural vegetation of the island, with the population being exterminated by 1991 (Department of Conservation Citation2014). Campbell Island is now inscribed in the United Nations Educational, Scientific and Cultural Organisation (UNESCO) World Heritage list (UNESCO Citation2014). At the time of writing a small flock of Campbell Island sheep were farmed by a member of the Rare Breeds Conservation Society in Canterbury.
Raglan
The Raglan sheep were descendants of a flock of Romney type sheep that were isolated when the Paritata Peninsula in Raglan (Whaingaroa) Harbour (37°46′S, 174°56′E) 30 km west of Hamilton was fenced off and the land on the peninsula abandoned in the 1930s (Bigham & Cockrem Citation1982). In April 1976 two mixed age rams and four mixed age ewes were caught on the peninsula and transported to the Whatawhata Research Centre. In December 1976 a further eight mixed age ewes were caught on the peninsula and transported to the Whatawhata Research Centre prior to the removal of all livestock from the peninsula for conservation purposes. At the time of writing two small flocks of Raglan sheep were farmed by members of the Rare Breeds Conservation Society in Canterbury.
Figure A1.1. Photo taken in 1976 on Campbell Island of some of the captured Campbell Island rams and ewes before they were transported to mainland New Zealand. The rams were shorn with blades prior to the sea voyage. Photo: W. R. Regnault.

Appendix 2. Gene frequency studies
While natural selection occurred during the period the feral flocks were grazing in the wild without human intervention, recent genetic studies of material from the Hokonui and Campbell Island flocks have indicated that there was insufficient selection pressure imposed during this phase to affect the degree of homozygous individuals. At least this was not evident for the presence or frequency of alleles associated with genes KRTAP1-1 (encodes a protein commonly found in wool), KRT33 (encodes wool keratin), ADR83 (associated with neonatal mortality) or DQA2 (associated with the immune response to pathogens and parasites) (McKenzie et al. Citation2010). No material from the Raglan flock was evaluated. A comparison of the frequency of the KRT33 allele in a reference Merino flock (McKenzie et al. Citation2010) and in a sample of Perendale sheep used in the current study (Sumner et al. Citation2013) showed there was a marked difference between the Merino and Perendale sheep and a similar marked difference between the Hokonui and Campbell Island sheep. This concurs with the belief that the Campbell Island sheep were established by crossbreeding Merino and British long wool breeds.