
ABSTRACT
Four observations of what was probably a single vagrant adult female whitetip reef shark Triaenodon obesus were made at Raoul and Meyer Islands, Kermadec Islands, New Zealand, over a 12 month period in 2014/15. This is the most southerly occurrence of the whitetip reef shark and ambient winter temperatures were close to the assumed thermal minimum for the species. The closest potential source population occurs at Minerva Reefs, about 598 km northwest of Raoul Island.
Introduction
The whitetip reef shark Triaenodon obesus (Rüppell 1837), grey reef shark Carcharhinus amblyrhynchos (Bleeker 1856) and blacktip reef shark Carcharhinus melanopterus (Quoy and Gaimard, 1824) are characteristic Indo-Pacific coral reef species (Randall Citation1977; Johnson Citation1984). The ranges of all three species extend from East Africa to the central Pacific Ocean, with the whitetip reef shark also occurring in the tropical eastern Pacific (Whitney et al. Citation2012b). Paradoxically for such widespread species they typically exhibit limited home range sizes and strong site fidelity (Randall Citation1977; Johnson Citation1984; Papastamatiou et al. Citation2009; Heupel et al. Citation2010; Field et al. Citation2011; Fitzpatrick et al. Citation2011; Barnett et al. Citation2012; Whitney et al. Citation2012a; Chin et al. Citation2013). In the blacktip reef shark this is associated with high genetic structure across its range but genetic structuring is weak in grey reef shark and whitetip reef shark populations, suggesting that rates of long-distance dispersal may be higher than observed in most tracking studies (Whitney et al. Citation2012b; Vignaud et al. Citation2014; Momigliano et al. Citation2015).
The whitetip reef shark usually inhabits shallow coral reef lagoons and outer reef slopes from the intertidal zone to at least 330 m depth but it also occurs on rocky reefs and seamounts (Last and Stevens Citation2009). It is normally nocturnal and rests on the bottom during the day, often under ledges or in caves (Randall Citation1977; Fitzpatrick et al. Citation2011). In the western Pacific it is distributed from the Ryukyu Islands (27°46′N) and Ogasawara Islands (27°40′N) south to the Kermadec Islands (29°17′S) (Last and Stevens Citation2009; Duffy and Ahyong Citation2015). Its occurrence at the Kermadec Islands was first reported by Duffy and Ahyong (Citation2015) based upon a photograph of an individual taken in Boat Cove, Raoul Island (29°16.721′S, 177°53.694′W) in April 2014. No other information accompanied the record. Here we report details of that sighting and three others made at Raoul Island between February 2014 and February 2015, as well as relevant environmental data. The biological implications of these sightings are briefly discussed and the whitetip reef shark is formally reported from Minerva Reefs, Tonga, for the first time.
Methods
All sightings were made opportunistically by the authors (NF, TG, SH) between 5 February 2014 and 28 February 2015. Total length (TL) was visually estimated. Sex was determined by examining video and still photographs for the presence–absence of claspers. The claspers of mature male whitetip reef sharks are large and conspicuous, almost reaching the anal fin origin (Whitney et al. Citation2012a). Sea surface temperature was recorded by deploying a bucket thermometer from Fishing Rock, Raoul Island; monthly sample sizes ranged from two to five readings (average 4.25, mode 4.0).
Results and discussion
Three sightings were made at 1–5 m depth in Boat Cove, Raoul Island, and one in Boat Harbour, Meyer Islands (, ). Boat Harbour is located about 5.4 km by water from Boat Cove, and 1.8 km off the northern coastline of Raoul Island. Male whitetip reef sharks mature at 112–116 cm TL and females at 114–122 cm TL, maximum size is 170 cm TL (Randall Citation1977; Last and Stevens Citation2009). All observations were therefore of a mature individual or individuals. Sex could only be determined on two occasions, neither sighting was of an obviously pregnant female (; Whitney et al. Citation2012a). Although whitetip reef sharks can be individually identified by the pattern of dark spots on their flanks and the combination of fins with white tips, no images suitable for individual photo-identification were obtained (Randall Citation1977; Whitney et al. Citation2012a). The total number of sharks observed is therefore unknown; however, the proximity of the sightings to each other, estimated length and combination of white-tipped fins (first dorsal fin, upper caudal lobe) suggest that all sightings were of the same individual.
Figure 1. Female Triaenodon obsesus, c.1500 mm total length, Boat Harbour, Meyer Islands, Kermadec Islands, 28 February 2015.

Table 1. Observations of Triaenodon obsesus at the Kermadec Islands, New Zealand.
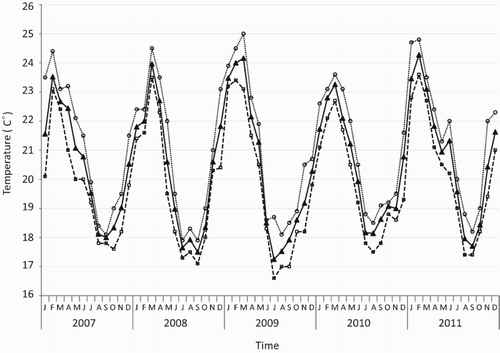
The whitetip reef shark is a tropical species that is most abundant at ambient temperatures above 26 °C (Nadon et al. Citation2012; Richards et al. Citation2012; Speed et al. Citation2012). These sightings represent the most southerly occurrence of the species. Before this, the southernmost records were from Leadsman Shoal, South Africa (27°48′S) and Flat Rock, Moreton Bay, Australia (27°25′S) (SAIAB, National Fish Collection Cat. No. 6269; Johnson Citation2010). Annual sea surface temperatures (SST) recorded at Raoul Island range from 16.6 to 25 °C, with average monthly SST seldom exceeding 24 °C in summer and regularly falling below 18 °C in winter (). Remote sensed mean monthly SST (derived from NOAA’s daily Optimum Interpolation SST) near Raoul Island during February, April and July 2014 and February 2015 were 25.2 °C, 23.8 °C, 19 °C and 23 °C, respectively; the minimum temperature recorded during winter 2014 was 18.4 °C (P. Sutton, NIWA, pers. comm.). The whitetip reef shark’s lower thermal limit is considered to be about 18 °C (Nadon et al. Citation2012).
Figure 2. Monthly maximum (open circles), average (solid triangles) and minimum (open triangles) sea surface temperatures recorded at Fishing Rock, Raoul Island, Kermadec Islands.
Scientific divers have visited the Kermadec Islands on at least 14 occasions between 1984 and 2014, making it unlikely that a resident but hitherto undetected population of whitetip reef sharks occurs there. The closest whitetip reef shark population to Raoul Island occurs at North and South Minerva Reefs (23°38.5′S, 178°54.7′W and 23°55.8′S, 179°6.2′W, respectively) on the northern end of the South Fiji Ridge about 598 km northwest of Raoul Island (Natural History New Zealand Ltd. archival footage, 27–29 May 1992). Intervening depths in the Havre Trough–Lau Basin exceed 2000 m. However, there are at least 12 seamounts rising to within 100 m of the surface between South Minerva Reef and Raoul Island that could act as stepping stones for dispersal of shallow-water species. Minimum distances between these seamounts range from 40 to 115 km, with the distance decreasing toward Raoul Island (Chart NZ14638, INT638).
Although maximum observed dispersal distance in the whitetip reef shark is only 26.4 km, the species’ widespread Indo-Pacific distribution means that some individuals must occasionally engage in oceanic dispersal (Whitney et al. Citation2012a, Citation2012b). Genetic structuring of whitetip reef shark populations may occur along stretches of continuous or semi-continuous habitat, consistent with limited home range size, but little or no genetic structure is evident across the Pacific Ocean (Whitney et al. Citation2012b). This may be due to recent dispersal across the East Pacific Barrier and ongoing range extension in the eastern Pacific (Whitney et al. Citation2012b; Corgos and Rosende-Pereiro Citation2016). Whitney et al. (Citation2012b) suggested that long-distance dispersal by whitetip reef sharks may be driven by high levels of intraspecific competition in oceanic island populations. Although whitetip reef sharks are common at Minerva Reefs (T. Trnski, Auckland Museum, pers. comm.) no information is available on home range size and movements there. Whitney et al. (Citation2012b) suggested that long-distance dispersal by whitetip reef sharks may be passive (by drifting) or semi-passive, possibly involving association with flotsam. Although surface currents north of the Kermadec Islands are highly variable, mean flow is weakly eastward, creating a barrier to passive dispersal between Minerva Reefs and the Kermadecs (Sutton et al. Citation2012; Edgar et al. Citation2013). As there are no other up-current source populations it seems likely that movement of the shark observed at Raoul Island was active rather than passive.
Although Raoul Island represents marginal habitat for whitetip reef sharks the occurrence of a mature female there illustrates how new populations of elasmobranchs could be founded well outside their established range by dispersal of a single female. A species’ persistence is largely determined by geography, environmental gradients and its dispersal ability (Hanski Citation1998). The ability to successfully disperse across large distances may favour persistence through colonisation of new habitats and recruitment to isolated populations (Hargreaves and Eckert Citation2014). Conversely, high rates of dispersal may threaten the survival of small, isolated populations near the edge of a species range (Hanski Citation1998; Phillips Citation2012). Large body size and energetic efficiency favour long-distance dispersal by elasmobranchs and even apparently sluggish benthic species undertake seasonal migrations spanning hundreds of kilometres in each direction (Chapman et al. Citation2015; Bass et al. Citation2016). Viviparity, parthenogenesis and sperm storage are reproductive traits that also favour colonisation of new habitats by elasmobranchs (Thiel and Haye Citation2006; Robinson et al. Citation2011; Hargreaves and Eckert Citation2014; Bernal et al. Citation2015). Like most sharks the whitetip reef shark is viviparous, giving birth to one to five relatively large pups after a 10–12 month gestation period (Last and Stevens Citation2009). It is also one of five sharks in which parthenogenesis has been reported (Portnoy et al. Citation2014). Although the fitness of parthenogenetic elasmobranch offspring is uncertain, second-generation parthenogenesis has been reported in another shark species and ecological theory suggests that asexual reproduction may be selected for at range margins (Hargreaves and Eckert Citation2014; Straube et al. Citation2016). Range extension mediated by female dispersal in this way could be rapid and would result in low genetic diversity in the founder population and the sharing of genotypes across large distances (e.g. Benavides et al. Citation2011; Whitney et al. Citation2012b; Vignaud et al. Citation2014).
No whitetip reef sharks were observed during filming and research expeditions to Raoul Island in October 2015 and October 2016. More than 118 diver hours were spent at various locations around Raoul Island and its outlying islets during these expeditions, including 11 hours in Boat Cove and 42 hours at the Meyer Islands (T. Trnski, Auckland Museum; K. Scollay, Ocean Answers; C. Middleton, NIWA, pers. comm.). This apparent absence could have been due to chance, diver avoidance, mortality, movement into deeper water, or movement away from the islands. The latter could include return to the source population (Chapman et al. Citation2015).
Acknowledgements
We are grateful to Phil Sutton, National Institute of Water & Atmospheric Research (NIWA), for providing satellite SST data, to Dave Sanderson, Auckland Museum, who assisted with preparation of , and to Tom Trnski, Auckland Museum, and Libby Liggins, Massey University, who reviewed the manuscript before submission.
We thank Olly Rudd, Natural History New Zealand (NHNZ), for providing access to archival footage of Triaenodon obesus at Minerva Reefs (NHNZ; LR590, 27 May 1992; LR592, 28 May 1992; LR593, 29 May 1992). Associate editor: Dr Jonathan Banks.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Barnett A, Abrantes KG, Seymour J, Fitzpatrick R. 2012. Residency and spatial use by reef sharks of an isolated seamount and its implications for conservation. PLoS ONE. 7(5):e36574. doi: 10.1371/journal.pone.0036574
- Bass NC, Mourier J, Knott NA, Day J, Brown C, Guttridge T. 2016. Long-term migration patterns and bisexual philopatry in a benthic shark species. Marine and Freshwater Research. doi.org/10.1071/MF16122.
- Benavides MT, Feldheim KA, Duffy CA, Wintner S, Braccini JM, Boomer J, Huveneers C, Rogers P, Mangel JC, Alfaro-Shiguetol J, et al. 2011. Phylogeography of the copper shark (Carcharhinus brachyurus) in the southern hemisphere: implications for the conservation of a coastal apex predator. Marine and Freshwater Research. 62:861–869. doi: 10.1071/MF10236
- Bernal MA, Sinai NL, Rocha C, Gaither MR, Dunker F, Rocha LA. 2015. Long-term sperm storage in the brownbanded bamboo shark Chiloscyllium punctatum. Journal of Fish Biology. 86:1171–1176. doi: 10.1111/jfb.12606
- Chapman DD, Feldheim KA, Papastamatiou YP, Hueter RE. 2015. There and back again: a review of residency and return migrations in sharks, with implications for population structure and management. Annual Review of Marine Science. 7:547–570. doi: 10.1146/annurev-marine-010814-015730
- Chin A, Tobin AJ, Heupel MR, Simpfendorfer CA. 2013. Population structure and residency patterns of the blacktip reef shark Carcharhinus melanopterus in turbid coastal environments. Journal of Fish Biology. 82:1192–1210. doi: 10.1111/jfb.12057
- Corgos A, Rosende-Pereiro A. 2016. First record of the whitetip reef shark, Triaenodon obesus from the coast of Jalisco, western Mexico mainland. Marine Biodiversity Records. 9:42. doi: 10.1186/s41200-016-0069-7
- Duffy CAJ, Ahyong ST. 2015. Annotated checklist of the marine flora and fauna of the Kermadec Islands Marine Reserve and northern Kermadec Ridge, New Zealand. Bulletin of the Auckland Museum. 20:19–124.
- Edgar G, Stuart-Smith R, Thomson R, Freeman D, Southwood P. 2013. Reef life survey assessment of biodiversity in northern New Zealand marine reserves and associated coastlines. Report for New Zealand Department of Conservation. Hobart: Aquenal Pty Ltd and Reef Life Survey Inc. p. 52.
- Field IC, Meekan MG, Speed CW, White W, Bradshaw CJA. 2011. Quantifying movement patterns for shark conservation at remote coral atolls in the Indian Ocean. Coral Reefs. 30:61–71. doi: 10.1007/s00338-010-0699-x
- Fitzpatrick R, Abrantes KG, Seymour J, Barnett A. 2011. Variation in depth of whitetip reef sharks: does provisioning ecotourism change their behaviour? Coral Reefs. 30:569–577. doi: 10.1007/s00338-011-0769-8
- Johnson RH. 1984. Sharks of tropical and temperate seas. Singapore: Les Editions du Pacifique. p. 170.
- Hanski I. 1998. Metapopulation dynamics. Nature. 396:41–49. doi: 10.1038/23876
- Hargreaves AL, Eckert CG. 2014. Evolution of dispersal and mating systems along geographic gradients: implications for shifting ranges. Functional Ecology. 28:5–21. doi: 10.1111/1365-2435.12170
- Heupel MR, Simpfendorfer CA, Fitzpatrick R. 2010. Large–scale movement and reef fidelity of grey reef sharks. PLoS ONE. 5:e9650. doi: 10.1371/journal.pone.0009650
- Johnson JW. 2010. Fishes of the Moreton Bay Marine Park and adjacent continental shelf waters, Queensland, Australia. Memoirs of the Queensland Museum – Nature. 54(3):299–353.
- Last PR, Stevens JD. 2009. Sharks and Rays of Australia, 2nd ed. Collingwood: CSIRO Publishing. p. 644.
- Momigliano P, Harcourt R, Robbins WD, Stow A. 2015. Connectivity in grey reef sharks (Carcharhinus amblyrhynchos) determined using empirical and simulated genetic data. Scientific Reports. 5:202. doi: 10.1038/srep13229
- Nadon MO, Baum JK, Williams ID, McPherson JM, Zgliczynski BJ, Richards BL, Schroeder RE, Brainard RE. 2012. Re-creating missing population baselines for Pacific reef sharks. Conservation Biology. 26:493–503. doi: 10.1111/j.1523-1739.2012.01835.x
- Papastamatiou YP, Lowe CG, Caselle JE, Friedlander AM. 2009. Scale-dependent effects of habitat on movements and path structure of reef sharks at a predator-dominated atoll. Ecology. 90:996–1008. doi: 10.1890/08-0491.1
- Phillips BL. 2012. Range shift promotes the formation of stable range edges. Journal of Biogeography. 39:153–161. doi: 10.1111/j.1365-2699.2011.02597.x
- Portnoy DS, Hollenbeck CM, Johnston JS, Casman HM, Gold JR. 2014. Parthenogenesis in a whitetip reef shark Triaenodon obesus involves a reduction in ploidy. Journal of Fish Biology. 85:502–508. doi: 10.1111/jfb.12415
- Randall JE. 1977. Contribution to the biology of the whitetip reef shark (Triaenodon obesus). Pacific Science. 31:143–164.
- Richards BL, Williams ID, Vetter OJ, Williams GJ. 2012. Environmental factors affecting large-bodied coral reef fish assemblages in the Mariana Archipelago. PLoS ONE. 7(2):e31374. doi: 10.1371/journal.pone.0031374
- Robinson DP, Baverstock W, Al-Jaru A, Hyland K, Khazanehdari KA. 2011. Annually recurring parthenogenesis in a zebra shark Stegostoma fasciatum. Journal of Fish Biology. 79:1376–1382.
- Speed CW, Meekan MG, Field IC, McMahon CR, Bradshaw CJA. 2012. Heat-seeking sharks: support for behavioural thermoregulation in reef sharks. Marine Ecology Progress Series. 463:231–245. doi: 10.3354/meps09864
- Straube N, Lampert KP, Geiger MF, Wei JD, Kirchhauser JX. 2016. First record of second-generation facultative parthenogenesis in a vertebrate species, the whitespotted bambooshark Chiloscyllium plagiosum. Journal of Fish Biology. 88:668–675. doi: 10.1111/jfb.12862
- Sutton P, Chiswell S, Gorman R, Kennan S, Rickard G. 2012. Physical marine environment of the Kermadec Islands region. Science for Conservation 318. Wellington: Department of Conservation. p. 16.
- Thiel M, Haye PA. 2006. The ecology of rafting in the marine environment. III. Biogeographical and evolutionary consequences. Oceanography and Marine Biology: An Annual Review. 44:323–429. doi: 10.1201/9781420006391.ch7
- Vignaud TM, Mourier J, Maynard JA, Leblois R, Spaet JLY, Clua E, Neglia V, Planes S. 2014. Blacktip reef sharks, Carcharhinus melanopterus, have high genetic structure and varying demographic histories in their Indo-Pacific range. Molecular Ecology. 23:5193–5207. doi: 10.1111/mec.12936
- Whitney NM, Pyle RL, Holland KN, Barcz JT. 2012. Movements, reproductive seasonality, and fisheries interactions in the whitetip reef shark (Triaenodon obesus) from community-contributed photographs. Environmental Biology of Fishes. 93:121–136. doi: 10.1007/s10641-011-9897-9
- Whitney NM, Robbins WD, Schultz JK, Bowen BW, Holland KN. 2012. Oceanic dispersal in a sedentary reef shark (Triaenodon obesus): genetic evidence for extensive connectivity without a pelagic larval stage. Journal of Biogeography. 39:1144–1156. doi: 10.1111/j.1365-2699.2011.02660.x