
ABSTRACT
We determined the seasonal diet of the variable hawk, Geranoaetus polyosoma, in a hyperarid and threatened habitat in the Atacama Desert, northern Chile, by analysing the contents of the bird's pellets. We estimated the biomass and number of individuals of each prey species consumed. We compared our results with those of other studies on G. polyosoma in South America. In general, the diet was characterised by extremely low number of prey, low diversity of species consumed and high dietary breadth. In contrast with other biomes of Chile, in our study reptiles formed the dominant food item and accounted for the highest percentage of biomass consumed from autumn to spring, with rodents being a seasonally significant major food item. Arthropods were also consumed, but the biomass ingested was negligible. Statistically significant differences were noted across seasons in the prey items consumed, reflecting a flexible and opportunistic response to the scarcity of prey available. Our findings add to the data present on the basic natural history of G. polyosoma and can aid in its conservation in the Atacama Desert.
Introduction
The variable hawk, Geranoaetus polyosoma (Quoy and Gaimard, 1824), is a medium-sized raptor that occupies a vast area of South America, ranging from Colombia to Argentina and Chile (Jiménez Citation1995; Rau Citation2014). This species inhabits both arid and Mediterranean environments and can be found from sea level up to 4000 m above sea level (Rau Citation2014). In northern Chile, G. polyosoma is a common bird that is resident throughout the year (Figueroa et al. Citation2003) and is primarily diurnal in habits (Jiménez Citation1995). It is a generalist predator, consuming a wide range of prey, including rodents, reptiles, insects and birds (Schlatter et al. Citation1980; Jiménez Citation1995; Travani et al. Citation2012).
Little work has been published on the dietary ecology of G. polyosoma. In South America most studies have been conducted in Mediterranean and temperate ecosystems, and have shown that its diet chiefly comprises birds, micromammals, lagomorphs, reptiles and, occasionally, arachnids (Schlatter et al. Citation1980; Jimenez Citation1995; Figueroa et al. Citation2003; Baladron et al. Citation2009; Travaini et al. Citation2012). However, research on its diet in the hyperarid zones (biomes with a rainfall of <1 mm) of South America has been limited and has not considered seasonal variations in diet. At present, only one study has been published on the diet of G. polyosoma in the Atacama Desert, indicating that this species consumes a low diversity of prey and does not select prey by body size (Valladares et al. Citation2015).
In this context, the ecosystem of Pampa del Tamarugal in northern Chile offers a particularly valuable scenario for the study of the trophic habits of G. polyosoma because its habitat is within the Atacama Desert, the oldest and most arid desert in the world (Hartley et al. Citation2005). The climatic conditions of this environment are extreme, with highly variable temperatures, low rainfall and scarce vegetation, which limit the quantity and variety of prey available for raptors (Carmona & Rivadeneira Citation2006; Carevic et al. Citation2013). Thus, we hypothesised that G. polyosoma will show a low diversity of prey in its diet and generalist feeding behaviour across the seasons. Our findings will also provide a better understanding of how predators are able to adapt to the extreme conditions found in the Atacama Desert.
As with many habitats across their range, the Atacama Desert has been subject to increasing anthropogenic modification and the effect of this on bird populations is unclear. For example, one of the main obstacles to the establishment of seedlings in Pampa del Tamarugal has been the decline in the aquifer water table in the past few decades (Rojas & Dassargues Citation2007). This trend is associated with increasing use of water by the mining industry and the urban population growth. This loss of water affects tree species and potentially affects nesting birds (Calderon et al. Citation2015). In fact, the Prosopis tree species dominant in this area has been classified as endangered (Carevic Citation2014). Therefore, quantifying the diet of raptors, such as the variable hawk, can provide a baseline for monitoring future ecological changes or management options in the Pampa del Tamarugal.
In the present study, we analysed the seasonal feeding habits of G. polyosoma in the hyperarid ecosystem of Pampa del Tamarugal and compared the results of our study with those of studies conducted on other biomes of Chile and South America.
Materials and methods
Our research was conducted in the Pampa del Tamarugal area located 60 km from the coastal city of Iquique, in Tarapacá Region of Chile. This ecosystem presents scarce and homogenous vegetation, mainly represented by forests of Prosopis spp. and less dominant plant species such as Atriplex atacamensis, Tessaria absinthiodes and Distichlis spicata (Carmona & Rivadeneira Citation2006).
The study of the diet of G. polyosoma was based on the analysis of 63 pellets collected in the summer (January to March), winter (July) and spring (November) of 2015. The samples were collected under three nesting sites in Pampa del Tamarugal (). Then, each pellet was moistened and crumbled to analyse its contents, removing tegumentary, dental, skeletal and chitinous remains. Each prey species was determined at the highest possible taxonomic level with the use of specialised keys (Donoso-Barros Citation1966; Peña Citation1992) and the reference collection of the Zoology Laboratory in Universidad Arturo Prat (Iquique, Chile). Most vertebrates in the pellets were identified on the basis of skulls, mandibles or dentary pairs, The number of individuals was determined using maximum count of left and right mandibles, skulls or dentary pairs. Insects were quantified by counting pairs of elytras.
Figure 1. Map of South America showing northern Chile and sites of pellet collection in Pampa del Tamarugal.
To estimate the quality of the collected information we performed two analyses. First, a species rarefaction curve, based on 100 randomisations (EstimateS 9.10; Colwell Citation2005), was constructed to determine whether the number of pellets sampled was enough to characterise the dietary diversity of G. polyosoma. Second, we estimated the expected total number of taxa in the diet of G. polyosoma using the non-parametric Chao1 index (EstimateS 9.10; Colwell Citation2005). The proportion between the observed and expected number of prey was an indicator of the completeness of our sampling, where values close to unity indicated a complete inventory.
To analyse the contribution of each prey species to biomass consumption, the number of individuals of each species in each pellet was multiplied by the average mass (weight in g) of each species. The mass value was obtained on the basis of studies conducted by Schlatter et al. (Citation1980), Donoso-Barros (Citation1996) and Carevic et al. (Citation2013). Seasonal differences in prey items were evaluated by the χ2 test. For this analysis, all prey groups found in the diet were included. To characterise the diet the following trophic statistics were used: (a) geometric mean mass of prey (GMMP) consumed (sensu Marti Citation1987); (b) Levins Index according to Simpson (Citation1949). For this index, we included all taxonomic categories of prey (maximum value was 4); (c) standardised Levins index by Colwell & Futuyma (Citation1971). This index was used to compare the trophic niche breadth in the Tamarugal across seasons where the supply of available prey for the variable hawk can be different; (d) diversity of consumed prey expressed through the Shannon–Wiener index (Shannon Citation1948), which quantifies the total diversity of a sample, being influenced by two main components: richness and equity. In order to compare Shannon indices between seasons in Pampa del Tamarugal a bootstrap approach with 1000 random samples was used according to Hammer & Harper (Citation2005). Approximate confidence intervals for Shannon–Wiener indices were also calculated with the bootstrapping procedure (1000 random samples were produced). All statistical analyses were performed with the program Past v3.0 (Hammer et al. Citation2001).
We compared the prey diversity by Shannon and evenness niche breadth indices (Pielou Citation1975) of the variable hawk at Pampa del Tamarugal with other sites in Chile and South America, based on an exhaustive literature search. All those studies followed similar sampling protocols and pooled dietary data across all seasons, which make them directly comparable to our results. These studies described the diet of the variable hawk at eight sites across Chile and Argentina, covering c. 30° of latitude and a wide range of variation in primary productivity, measured as the mean annual rainfall (Lima et al. Citation2002). Shannon and evenness indices were recalculated for each site considering taxa determination at the species, genus, family and, in a few cases, order level. To evaluate differences we performed a diversity permutation analysis by bootstrapping procedure with the program Past v3.0 (Hammer et al. Citation2001).
Results and discussion
Sixty-three pellets were collected in the Pampa del Tamarugal ecosystem. The composition of the diet of G. polyosoma for this zone revealed a preference for vertebrates, with a low consumption of insects. The reptile Microlophus theresioides was the most commonly consumed vertebrate, accounting for 45.3% of the total sample. The rodent Phyllotis darwini accounted for 40.4% of the total diet items, whereas insects only accounted for 13.5% of the total diet items and were represented by the families Tenebrionidae and Scarabaeidae, with 11.5% and 1.9%, respectively ().
Table 1. Seasonal description of Geranoaetus polyosoma diet in Pampa del Tamarugal (Atacama Desert).
Statistically significant differences were noted at the seasonal level in the prey items consumed by G. polyosoma (χ2 = 31.07, d.f. = 2, P < 0.01). Variable hawks mainly preyed on P. darwini (93.8%) in summer; the consumption of this rodent was 40% in autumn and none in spring. Microlophus theresioides was the most commonly preyed reptile in spring and autumn (71.4% and 53.3%, respectively), whereas it was not consumed in summer. The consumption of insects was mainly concentrated in the spring and autumn (28.6% and 6.7%, respectively; ). In terms of biomass, M. theresioides was the chief prey (> 68%), followed by P. darwini (> 30%) and insects (0.4%). The biomass provided by insects was low for all the stations analysed in Pampa del Tamarugal. The contribution of P. darwini was higher in summer than in autumn and spring. Microlophus theresioides was an important contributor to biomass consumed in all the seasons, particularly during autumn and spring. The average geometric weight of the prey reached a value of 45.1 g, with the values being higher in summer and autumn.
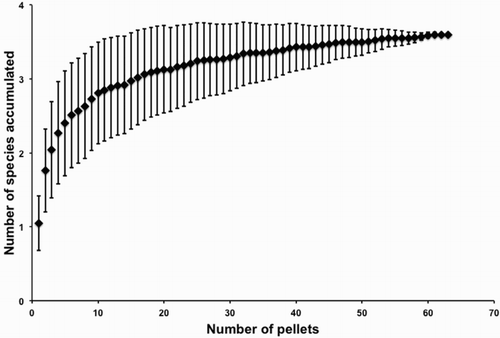
Rarefaction analysis showed an asymptotic curve, where a total of four prey were reached after sampling c. 60 pellets (). The extrapolated total richness estimated with Chao1 index was 3.25 prey items, very close to the observed value. The ratio between observed and expected total was 0.81, suggesting that the sampling has recorded most of the dietary diversity existing in the zone.
Figure 2. Rarefaction curve for the species richness found in the pellets of the variable hawk (mean and standard deviations) based on the number of replicates sampled. The asymptotic curve indicates that total species richness in the Pampa del Tamarugal was reached after about 60 samples. Calculation was performed using the EstimateS 9.10 program.
The trophic niche amplitude of the variable hawk as calculated by the Levins index (B and Bsta) showed a lower trophic amplitude value in summer than in autumn and spring. The diversity of prey was significantly higher in autumn and spring than in summer (bootstrapped Shannon–Wiener index, P < 0.01). This pattern can be explained because in cold seasons rodents are not available at the same proportion than in temperate seasons and, therefore, variable hawks may need to consume alternative prey such as reptiles and insects to compensate for the scarcity of food.
This is the first study on the diet of G. polyosoma in the hyperarid ecosystem of the Atacama Desert, northern Chile. The diet of this bird mainly comprised vertebrates, which accounted for 85% of the individuals found in the pellets. This result was in accordance with those of previous studies of G. polyosoma (Travaini et al. Citation2012; Rau Citation2014). In trophic terms, the consumption of reptiles and rodents classifies this species as a bird with diurnal–crepuscular habits because of the activities of these prey species. In particular, the results indicate that the consumption of reptiles is higher than that of micromammals (45% vs 40%). These results are in accordance with those of Valladares et al. (Citation2015), who reported that, in northern Chile, the consumption of reptiles is higher than that of micromammals. However, it should be noted that the number of micromammals found was lower than that in the present study (40% vs 20%). Furthermore, the results of the present study differ from studies in central and southern Chile, where the consumption of micromammals was reported to be higher than that of reptiles (Schlatter et al. Citation1980; Jiménez Citation1995; Figueroa et al. Citation2003). Studies in other South American countries have shown similar patterns of predation on small mammals and reptiles (Monserrat et al. Citation2005; Baladrón et al. Citation2009; Travaini et al. Citation2012). The differences could be explained by the fact that there is a higher abundance of reptiles in desert areas (Donoso-Barros Citation1966).
The diet of G. polyosoma in the Pampa del Tamarugal had low biodiversity, mainly consisting of reptiles, followed by small mammals and a small amount of insects. In comparison with more distant areas of Chile and Argentina, the biodiversity tended to be higher (A). Diversity measured with the Shannon index was lower in the Tamarugal area compared to all sites in Chile and Argentina (with the exception of Dehesa site), indicating differences in diversity of prey consumed (C). Thus, it is possible that the low primary productivity in the hyperarid ecosystem of Tamarugal would affect the variability of faunal populations, resulting in a low species prey diversity available for the variable hawk (Noy-Meir Citation1974; Carmona & Rivadeneira Citation2006; Carevic et al. Citation2013).
Figure 3. Variation of prey diversity consumed by Geranoaetus polyosoma in Chile and Argentina across the rainfall gradient. Species data and mean annual rainfall were obtained from: this study (Tamarugal); Valladares et al. (Citation2015) (Atacama); Schlatter et al. (Citation1980) (Dehesa); Figueroa et al. (Citation2003) (P. Tranquilo); Baladrón et al. (Citation2009) (B. Aires); Monserrat et al. (Citation2005) (Neuquén); and Travaini et al. (Citation2012) (Junín and Bosque P.). A, Diversity of prey measured with Shannon index; B, diversity measured with evenness index. Error bars indicate standard error for indexes; C, Shannon (H) and evenness (E) indices were evaluated to establish statistical differences among sites by using diversity permutation analysis with the software Past v3.0. *Indicates significant differences, and ns indicates no significant differences at P < 0.05 among sites and indices (H/E), respectively.
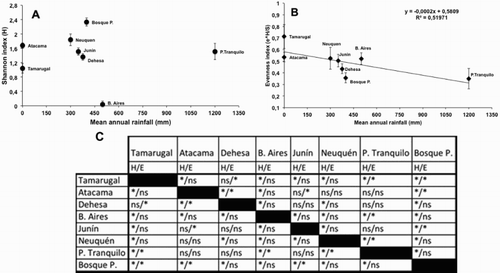
On the other hand, the evenness index calculated for sites from Chile and Argentina revealed a decreasing pattern across the rainfall gradient showing values from 0.71 to 0.34 (B). In fact, dietary evenness in the Tamarugal showed significant differences with Dehesa, P. Tranquilo and Bosques P. sites, which showed mean annual rainfall greater than 350 mm (). It should be noted that values close to 0 indicate diets dominated by a single kind of prey, while values close to 1 indicate diets where all prey species contribute an equal amount. Thus, we concluded that the diet of the variable hawk in Pampa del Tamarugal was generalist because it consumed a low diversity of prey (only four taxa), but an equal proportion of each. This pattern can be influenced by the scarcity of prey in the field, which forces G. polyosoma to take all available prey species in equal proportion to compensate for the scarcity of food. Thus, the comparison of diet diversity of G. polyosoma in the Pampa del Tamarugal with others populations in ecosystems of South America suggest there is a similar pattern in species prey richness, and evenness of the diet, which consistently show variation in the diet. This would indicate that G. polyosoma has a strong capacity for flexibility, because this predator can vary its diet depending on the environmental conditions (Jimenez Citation1995; Valladares et al. Citation2015) and its response to the availability of prey in the field (Jaksic et al. Citation1993).
In the study area, the quantity and quality of nesting sites for birds are quite low, particularly in Pampa del Tamarugal, because the plant density and diversity is low (Carevic et al. Citation2014). In addition, the development of seedlings, and thus recruitment of trees into the forests in Pampa del Tamarugal, has been declining because of water loss by natural causes, increased mining and urban population growth over the past few decades. This affects tree survival and potentially affects nesting sites for the variable hawks (Calderon et al. Citation2015). Therefore, trophic studies of raptors could contribute to understand how human impacts can affect the ecological properties of top predator species, especially in fragile desert ecosystems like Pampa del Tamarugal.
In summary, we found that the diet of G. polyosoma in Pampa del Tamarugal was particularly different from others reported in South America, because it was characterised by an extremely low number of prey, low diversity of species in the diet and high dietary breadth, suggesting that this top predator has a flexible, opportunistic and wide food strategy in response to the scarcity of prey available in the hyperarid Pampa del Tamarugal ecosystem. Further, our finding of seasonal variability in ecological indicators emphasises the necessity of conducting such studies at relevant seasonal time scales to assess the variability of diet of birds of prey across their range.
Acknowledgements
F.S. Carevic thanks the logistical support for fieldwork in National Reserve Pampa del Tamarugal provided by CONAF Tarapacá and Fondecyt project number 11130242. E.R. Carmona thanks the Dirección General de Investigación y Postgrado of Universidad Católica de Temuco, DGIP UCT project No. CD2010-01, and MECESUP UCT 0804 project.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Baladrón AV, Malizia AI, Bó MS. 2009. Predation upon the subterranean rodent Ctenomys talarum (tuco-tucos) by Buteo polyosoma (red-backed hawks) in coastal grasslands of Argentina. Studies on Neotropical Fauna and Environment. 44:61–65. doi: 10.1080/01650520902915022
- Calderon G, Garrido M, Acevedo E. 2015. Prosopis tamarugo Phil.: a native tree from the Atacama Desert groundwater table depth thresholds for conservation. Revista Chilena de Historia Natural. 88:18–25. doi: 10.1186/s40693-015-0048-0
- Carevic FS. 2014. The role of ecophysiological studies in the genus Prosopis: implications for the conservation of drought-prone species. Idesia. 32:77–81. doi: 10.4067/S0718-34292014000400010
- Carevic FS, Carmona ER, Muñoz-Pedreros A. 2013. Seasonal diet of the burrowing owl Athene cunicularia (Molina, 1782) (Strigidae) in a hyperarid ecosystem of the Atacama Desert in northern Chile. Journal of Arid Environments. 97:237–241. doi: 10.1016/j.jaridenv.2013.07.008
- Carevic FS, Delatorre J, Arenas J. 2014. Physiological strategies during frost periods for two populations of Prosopis burkartii, an endangered species endemic to the Atacama desert. Journal of Arid Environments. 114:79–83.
- Carmona ER, Rivadeneira MM. 2006. Food habits of the barn owl Tyto alba in the National Reserve Pampa del Tamarugal, Atacama Desert, north Chile. Journal of Natural History. 40:473–483. doi: 10.1080/00222930600699904
- Colwell RK. 2005. EstimateS: Statistical estimation of species richness and shared species from samples. Version 7.5. User’s Guide and application. http://purl.oclc.org/estimates.
- Colwell RK, Futuyma DJ. 1971. On the measurement of niche breadth and overlap. Ecology. 52:567–576.
- Donoso-Barros R. 1966. Reptiles de Chile. Santiago (Chile): Ediciones Universidad de Chile; p. 458.
- Figueroa RA, Corales ES, Alvarado S. 2003. Diet of the red-backed hawk (Buteo polyosoma) in a forested area of the Chilean Patagonia and its relation to the abundance of rodent prey. Hornero. 18:43–52.
- Hammer Ø, Harper DAT. 2005. Paleontological data analysis. Malden (MA): Wiley-Blackwell; p. 368.
- Hammer Ø, Harper DAT, Ryan PD. 2001. PAST: paleontological statistics software package for education and data analysis. Palaeontologia Electronica 4, 9. http://palaeo-electronica.org/2001_1/past/issue1_01.htm.
- Hartley AJ, Chong G, Houston J, Mather AE. 2005. 150 million years of climate stability: evidence from the Atacama Desert, northern Chile. Journal of the Geological Society. 162:421–424. doi: 10.1144/0016-764904-071
- Jaksic FM, Feinsinger P, Jiménez JE. 1993. A long-term study on the dynamics of guild structure among predatory vertebrates at a semi-arid neotropical site. Oikos. 67:87–96. doi: 10.2307/3545099
- Jiménez J. 1995. Historia natural del aguilucho Buteo polyosoma: una revisión. Hornero. 14:1–9.
- Lima M, Stenseth NC, Jaksic FM. 2002. Food web structure and climate effects on the dynamics of small mammals and owls in semi-arid Chile. Ecology Letters. 5:273–284. doi: 10.1046/j.1461-0248.2002.00312.x
- Marti CD 1987. Raptor food habits studies. In: Pendleton BAG, Millsap BA, Kline KW, Bird DM, editors. Raptor management techniques manual. 1st edn. Washington (DC): National Wildlife Federation; p. 67–80.
- Monserrat AL, Funes MC, Novaro AJ. 2005. Dietary response of three raptor species to an introduced prey in Patagonia. Revista Chilena de Historia Natural. 78:425–439. doi: 10.4067/S0716-078X2005000300006
- Noy-Meir I. 1974. Desert ecosystems: higher tropic levels. Annual Review of Ecology and Systematics. 5:195–214. doi: 10.1146/annurev.es.05.110174.001211
- Peña L. 1992. Introducción a los insectos de Chile, tercera ed. Santiago (Chile): Editorial Universitaria; p. 255.
- Pielou E. 1975. Ecological diversity. New York: John Wiley & Sons; p. 165.
- Rau JR. 2014. Papel ecológico de las aves rapaces: del mito a su conocimiento y conservación en Chile. Santiago (Chile): Ediciones Gráfica Metropilitana; p. 38.
- Rojas R, Dassargues A. 2007. Groundwater flow modelling of the regional aquifer of the Pampa del Tamarugal, northern Chile. Hydrogeology Journal. 15:537–551. doi: 10.1007/s10040-006-0084-6
- Schlatter RP, Yañez J, Jaksic F. 1980. Food-niche relationships between Chilean eagles and red-backed buzzards in central Chile. Auk. 97:897–898.
- Shannon CE. 1948. A mathematical theory of communication. Bell System Technical Journal. 27:379–423.
- Simpson EH. 1949. Measurement of diversity. Nature. 163:688.
- Travaini A, Santillán MA, Zapata SC. 2012. Diet of the red-backed hawk (Buteo polyosoma) in two environmentally contrasting areas of Patagonia. Studies on Neotropical Fauna and Environment. 47:25–32. doi: 10.1080/01650521.2011.649948
- Valladares P, Álvarez N, Urrutia N, Olivares F, Alvarado S. 2015. Dieta del aguilucho común Geranoaetus polyosoma (Quoy & Gaimard 1824) en la Región de Atacama, Chile. Gayana. 79:121–127.