
ABSTRACT
Pselaphotumulus Owens and Carlton, gen. nov., is described as the sixth genus in the tribe Pselaphini known from New Zealand. Three new species are described: Pselaphotumulus aorerei, sp. nov., Pselaphotumulus dubius, sp. nov. and Pselaphotumulus unus, sp. nov.. Three species, Pselaphus cavelli (Broun 1893), Pselaphus oviceps (Broun 1917) and Pselaphus urquharti (Broun 1917) are transferred to Pselaphotumulus, nov. combs. Lectotypes from type series in the New Zealand Broun Collection (Natural History Museum, London) are designated for these three species. Habitus photographs, distributional maps and line drawings of diagnostic characters are provided for each species. A key to species is provided. Searches of museum collections have not yielded representatives outside of New Zealand, suggesting that this is the first endemic genus in the tribe Pselaphini described from the New Zealand’s main islands, specifically, the South Island. Pselaphotumulus species exhibit restricted distribution patterns that approximately coincide with the Pacific/Indo Australian Plate boundary.
Introduction
The supertribe Pselaphitae is well represented in both tropical and temperate regions (Newton and Chandler Citation1989). Within this group, the tribe Pselaphini exhibits high levels of generic and species diversity in the Australian, Oriental and Western and Eastern Palearctic regions, while the Neotropical fauna is notably depauperate. A generic revision of the tribe in Australia (Chandler Citation2001) resulted in the description of five new genera from the region, including four monotypic to Australia as a result of the transfer of several species previously placed in the paraphyletic genus Pselaphus Herbst. Currently, five genera of Pselaphini are described from New Zealand (NZ) (Nomura and Leschen Citation2006). About one-third of the NZ species described in the genus Pselaphus will be transferred to Pselaphaulax, and the remainder will be reassigned among several newly-described genera and the genus Pselaphogenius (first author, unpublished data). As a part of the current efforts towards a resolution of the tribe in NZ we describe a new genus, Pselaphotumulus, bringing the total number of genera of Pselaphini in NZ to six.
Materials and methods
External morphological characters of all dry specimens were examined using an Olympus SZH10 dissecting microscope. Dissections were prepared for male specimens to allow detailed examination of aedeagal characters using an Olympus BX50 compound microscope fitted with a drawing tube.
For dissection, point-mounted individuals were removed from insect pins and relaxed in hot water for 10–15 minutes. Specimens were separated from points using an insect pin to tease away residual adhesive. Insect pins were then used to separate the terminal abdominal segments and tease out the aedeagus. Aedeagi were placed in glycerin on slide mounts for examination and drawing. After examination, aedeagi were removed from slide mounts and washed in several changes of 95% EtOH. Specimens were re-mounted with the body point mounted and the aedeagus glued to the point, dorsal side up.
Descriptions of external morphology follow Chandler (Citation2001) except meso-, meta- and abdominal sternites are referred to as ventrites. Plates were prepared and edited using Adobe Photoshop® (version cc2015). Habitus photographs were taken using a Syncroscopy® Automontage imaging system and edited using Adobe Photoshop®.
Specimens were obtained from the following institutions:
Verbatim label data are provided for all specimens examined and they are organised alphabetically by the New Zealand Entomology regions two-letter codes (Crosby et al. Citation1998). Two slashes (‘//’) are used to indicate label breaks. Sex and specimen numbers are indicated preceding label data (i.e. ‘(2 males, 1 female)’), and lending institution is indicated following label data (i.e. ‘(NZAC)’). All type specimens are deposited in the NHM or NZAC as indicated.
Lectotype designations
Syntypic series of most of the Coleoptera species described by Thomas Broun are deposited in the Natural History Museum, London as a result of New Zealand’s status as part of the British Empire during the period. Many of these are represented by short series labelled with numerical codes that correspond to numbers in his descriptions. Although many bear type labels, these do not constitute primary types under the ICZN because they were not specifically designated in his descriptions. In fact, evidence suggests that these type labels were added later, and not by Broun himself (Lord and Leschen Citation2014). Some of these series include mixed species, so designating lectotypes is necessary to fix the species to appropriate type specimens.
Taxonomy
Genus Pselaphotumulus (Owens and Carlton), gen. nov.
Type species. Pselaphotumulus cavelli (Broun Citation1893)
Etymology
The genus name is derived from the combination of the prefix ‘pselaph-’, which is commonly used for genera within the tribe Pselaphini, and ‘tumulus’, the Latin word for mound, referring to the broad, mound-like form of the gular process found in all species of this genus.
Diagnosis. Males. Head
eyes rounded, composed of 10–20 facets. Vertex convex; shallow sulcus beginning at base of vertex, extending to level of vertexal foveae; vertexal foveae large, set into margins of vertexal depression parallel to middle of eyes; vertexal depression widest between eyes, confluent with sulcus extending to apex of rostrum. Rostral sulcus bounded by lateral, flattened ridges extending to antennal acetabula. Frontal margin of rostrum in dorsal view typically pointed, bearing patch of modified, flattened setae. Antennal scape as long as antennomeres 2–3, antennae otherwise unmodified. Gular mound modified, slightly rounded, glabrous, expanded to occupy most of underside of head; area of head posterior to gular mound flattened, nude, gular foveae present, paired. Maxillary palpomere I elongate, 1/2 as long as palpomere II; palpomere II narrow, enlarged distally, shorter than head; maxillary palpomere III quadrate; maxillary palpomere IV typically longer than head, widened in distal 1/3–1/4, apical area smooth to slightly roughened, v-shaped sensory patch present. Thorax: pronotum widest at midpoint, narrowed anteriorly and posteriorly; basal sulcus present, delimited by lateral antebasal foveae, median antebasal fovea present, variable in size, basolateral foveae absent or reduced to small depressions. Prosternum in front of coxae slightly convex, anteriorly with modified flattened setae; lateral procoxal foveae present. Mesoventrite with median shield bearing patch of spongious setae posteriorly towards margin of mesocoxae; lateral mesoventral foveae present in large cavities filled with dense setae, meeting internally; lateral mesocoxal foveae absent; median mesoventral foveae present, paired, meeting internally; mesocoxae separated by extension of meso- and metaventrite. Metaventrite afoveate, produced medially into bulge with median depression, parallel lateral sulci bearing modified setae; metacoxae separated by metacoxal process of ventrite I. Elytra with two basal foveae, single sutural fovea; discal striae absent, sutural striae present; sparse apical fringe of setae present. Abdomen: tergites of usual form for tribe, unmodified; T2 without obvious basal foveae, lateral basal sulcus extending along width of ventrite, obscured by dense band of setae along posterior edge. Ventrite I covered in dense, shining setae; ventrite II with flattened area medially, apically with small median tubercle; ventrites otherwise unmodified.
Females
similar to males except eyes may be smaller, composed of fewer facets; secondary sexual modifications of the second visible ventrite reduced.
Comments
Prior to this study, the most recent new genus of Pselaphini described from NZ was Pselaphotheseus Park 1964 from Campbell Island. Aside from the recognition of Pselaphaulax Reitter from the North and South Islands, Pselaphotumulus is the first new genus record noted and the first new genus described from the NZ mainland ().
Figure 1. Habitus, male Pselaphotumulus oviceps (Broun).

This genus is apparently endemic to NZ. Species in Pselaphotumulus are united primarily by the following characters: (1) the frontal margin of the rostrum pointed with a patch of spongious setae, (2) modification of a gular process into a glabrous, ‘mound-like’ form that occupies almost the entire gular area (, a–b), (3) the presence of an antebasal sulcus on the pronotum margined by laterobasal foveae, (4) the median antebasal fovea present with basolateral foveae present as small depressions, (5) the mesoventrite with a median shield bearing a posterior patch of setae (, a–b), (6) the metaventrite produced medially into bulge with median ovate sulcus, lateral margins of sulcus sometimes with spongious setae, and (7) the absence of discal striae on the elytra. In NZ, this genus most closely resembles Pselaphaulax; however, the latter lacks the spongious patch on the apex of the rostrum, the broadly expanded gular mound, and the setose sulci on the metaventrite. Visually, these two genera may be easily separated by the previously mentioned characters, although they occur in similar habitats and both overlap in distribution on the South Island. Undescribed species likely exist and may be discovered with additional sampling.
Figures 2a,b–3a,b. ) Meso-/metaventrite and visible ventrites I and II, female Pselaphotumulus oviceps (Broun), (i) median mesoventral setose pit, (ii) lateral mesoventral foveae, (iii) mesoventral shield, (iv) mesocoxal cavities, (v) metaventrite, (vi) metacoxal cavities, (vii) basal setose band, (viii) visible ventrite ii, (ix) visible ventrite ii basal depression; ) Gular region and underside of head, female Pselaphotumulus oviceps (Broun), (i) mandibles, (ii) maxillary palpi, (iii) labium, (iv) gular mound, (v) paired gular foveae.
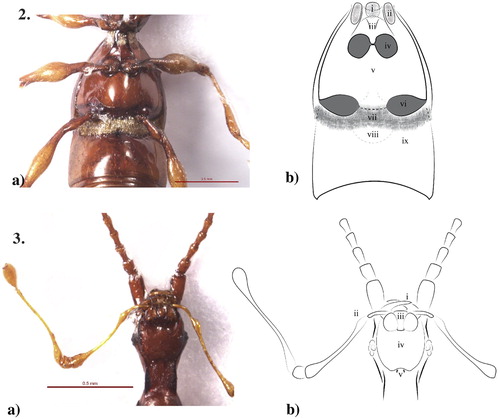
Currently, species of Pselaphotumulus are known only from the South Island. No specimens have been observed from collection localities on the North Island or on offshore island groups with distinct faunas, such as the Chathams or the subantarctic islands. This exclusively South Island distribution is not typical of other mainland genera of NZ Pselaphini (Pselaphaulax, Pselaphogenius, Pselaphophus), which may be found on both the North and South Islands.
Checklist of Pselaphotumulus species
Pselaphotumulus (Owens and Carlton), gen. nov.
Type species. Pselaphotumulus cavelli (Broun Citation1893)
Distribution. South Island NZ: NN, BR, NC, WD, MC, MK, FD.
Number of species. Six.
Pselaphotumulus aorerei (Owens and Carlton), sp. nov.
Type depository. NZAC.
Distribution. NZ: NN.
Pselaphotumulus cavelli (Broun Citation1893)
Pselaphus cavelli (Broun Citation1893: 1414)
Type depository. NHM.
Distribution. NZ: NN, BR.
Pselaphotumulus dubius (Owens and Carlton), sp. nov.
Type depository. NZAC.
Distribution. NZ: FD.
Pselaphotumulus oviceps (Broun Citation1917), comb. nov.
Pselaphus oviceps (Broun Citation1917: 380).
Type depository. NHM.
Distribution. NZ: NN, BR, WD, FD.
Pselaphotumulus unus (Owens and Carlton), sp. nov.
Type depository. NZAC.
Distribution. NZ: NC, WD.
Pselaphotumulus urquharti (Broun Citation1917), comb. nov.
Pselaphus urquharti (Broun Citation1917: 379)
Type depository. NHM.
Distribution. NZ: BR, NC, WD, MC, MK, FD.
Key to the species of Pselaphotumulus (males)
Unlike many genera of Pselaphini, Pselaphotumulus males exhibit external morphological characters useful for species-level identification. While the form of the male genitalia may be used exclusively for identification, reliance upon secondary sexual characters in males can aid identification without dissection. Due to the absence of observable external or genitalic characters, identification of females is only possible via association with male specimens.
Metaventrite tumosity with two lateral patches of spongious setae 2
1’ Metaventrite otherwise modified, not with two lateral patches of spongious setae3
2(1) Ventrite II flattened medially, apical margin produced into small tubercle
P. oviceps (Broun Citation1917), comb. nov.
2’ Ventrite II with two lateral carinae along median depression
P. urquharti (Broun Citation1917), comb. nov.
3(1’) Metaventrite tumosity with single median patch of spongious setae
P. unus (Owens and Carlton), sp. nov.
3 Metaventrite tumosity otherwise modified, not with single median patch of spongious setae4
4(3) Metaventrite flattened medially, forming shining ovate depression margined posteriorly by short setaeP. dubius (Owens and Carlton), sp. nov.
4’ Metaventrite formed as a rounded tumosity5
5(4’) Metaventrite with two patches of shortened setae posteriorlyP. cavelli (Broun Citation1893)
5’ Metaventrite with two low lateral tubercles, posteriorly P. aorerei (Owens and Carlton), sp. nov.
Pselaphotumulus aorerei (Owens and Carlton), sp. nov.
(, , a–b)
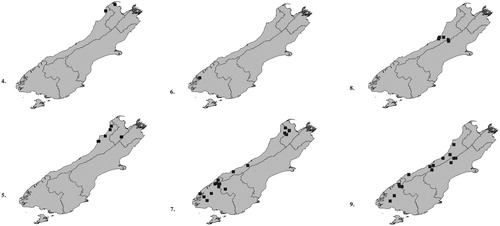
Figures 4–9. Distribution maps of Pselaphotumulus species. ) Pselaphotumulus aorerei, sp. nov.; ) Pselaphotumulus cavelli (Broun); ) Pselaphotumulus dubius, sp. nov.; ) Pselaphotumulus oviceps (Broun); ) Pselaphotumulus unus, sp. nov.; ) Pselaphotumulus urquharti (Broun).
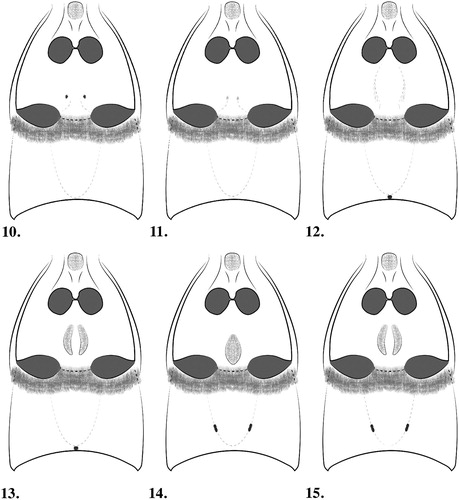
Figures 10–15. Meso-/metaventrites and tergites I–II, male Pselaphotumulus species. ) Pselaphotumulus aorerei, sp. nov.; ) Pselaphotumulus cavelli (Broun); ) Pselaphotumulus dubius, sp. nov.; ) Pselaphotumulus oviceps (Broun); ) Pselaphotumulus unus, sp. nov.; ) Pselaphotumulus urquharti (Broun).
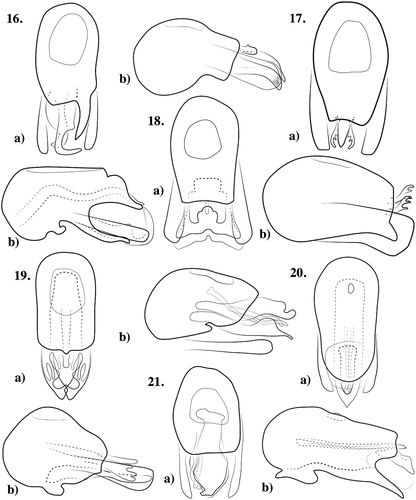
Figures 16a,b–21a,b. Aedeagi of Pselaphotumulus species. ) Pselaphotumulus aorerei, sp. nov. (a) dorsal, (b) lateral; ) Pselaphotumulus cavelli (Broun) (a) dorsal, (b) lateral; ) Pselaphotumulus dubius, sp. nov. (a) dorsal, (b) lateral; ) Pselaphotumulus oviceps (Broun) (a) dorsal, (b) lateral; ) Pselaphotumulus unus, sp. nov. (a) dorsal, (b) lateral; ) Pselaphotumulus urquharti (Broun) (a) dorsal, (b) lateral.
Type material. Holotype: (male)
Aorere Valley, Collingwood, Nelson 18.iv.63 J. I. Townsend (NZAC). The holotype is deposited in the NZAC.
Etymology
This species is named after the collection locality of the type series in the Aorere Valley.
Diagnosis
Externally, the combination of the metaventrite tumosity bearing two low lateral tubercles and ventrite II bearing two small lateral rows of punctures distinguishes this species from all others in the genus. The absence of basolateral foveae in combination with the smaller enlarged distal area of maxillary palpomere IV (enlarged in distal ¼ versus distal 1/3) are also unique. Additionally, the form of the aedeagus is diagnostic, particularly the form of the hook-shaped median lobe.
Description
MALE holotype. Integument: head, pronotum, elytra, abdomen, and appendages light brown, palpi lighter in colour. Head: wide, width between eyes greater than ½ length of head from base of vertex to apex of rostrum. Eyes rounded, composed of 14 facets. Frontal margin of rostrum in dorsal view pointed, with apical patch of modified, flattened setae. Gular area glabrous and mound-like, occupying most of gular area, gular foveae present. Maxillary palpomere IV longer than head, widened at base and distal 1/4, v-shaped sensory patches present. Thorax: pronotum with basal sulcus present, delimited by lateral antebasal foveae; median antebasal fovea present. Mesoventrite with median shield bearing patch of spongious setae in small concavity posteriorly. Metaventrite tumosity with two low, lateral tubercles posteriorly. Legs unmodified. Elytra with two basal foveae present, single sutural fovea; sutural striae present; apical fringe of setae sparse. Abdomen: tergites of usual form for tribe. Ventrite II bulging in distal 1/3, with two lateral rows of fine punctures. Genitalia: aedeagus asymmetrical. Median lobe hook-shaped. Ventral process curved to the right. Dorsal process well-developed. Lateral parameres present, paired, as long as median lobe.
Females
Unknown.
Distribution
Pselaphotumulus aorerei is known only from the type locality and a second collection event, both in NN.
Material examined. (2 males) New Zealand NN Scotts Hill Lookout Kohaihai// Sifted forest litter 2 Jan 2011 (JN).
Pselaphotumulus cavelli (Broun Citation1893)
(, , a–b)
Type material examined
After observing the type series of Pselaphus cavelli in the NHM and reviewing Broun’s original description, we designate the following specimen from the NHM as the lectotype: (1 male) TYPE (red/orange label)// 2642.// Charleston Westland// New Zealand Broun Coll. Brit. Mus. 1922–482 (designated lectotype). The additional specimen in the type series is a female specimen that is also a member of the species P. cavelli, collected from the same locality as the lectotype and is hereby designated as a paralectotype. Lectotype designations are necessary to fix the taxon to a name-bearing type. Broun did not specifically identify type specimens, and many of his syntype series include multiple species. All lectotype and paralectotype specimens are deposited in the NHM.
Diagnosis
Pselaphotumulus cavelli may be distinguished from other species in the genus via the combination of a bilobed dorsal and median process of the aedeagus.
Description
MALE lectotype. Integument: head, pronotum, elytra, abdomen, and appendages light brown, palpi lighter in colour. Head: wide, width between eyes greater than 1/2 length of head from base of vertex to apex of rostrum. Eyes rounded, composed of 14 facets. Frontal margin of rostrum in dorsal view pointed, with apical patch of modified, flattened setae. Gular area glabrous and mound-like, occupying most of gular area, gular foveae present. Maxillary palpomere IV longer than head, widened at base and distal 1/4, v-shaped sensory patches present. Thorax: pronotum with basal sulcus present, delimited by lateral antebasal foveae; median antebasal fovea present, basolateral foveae present as small depressions. Mesoventrite with median shield bearing patch of spongious setae in small concavity posteriorly. Metaventrite tumosity bearing two small tufts of setae posteriorly. Legs unmodified. Elytra with two basal foveae present, single sutural fovea; sutural striae present; apical fringe of setae sparse. Abdomen: tergites of usual form for tribe. Visible ventrite II slightly flattened medially throughout length. Genitalia: aedeagus symmetrical. Median lobe short, blunt. Ventral process broadly bilobed, with small striae apically. Ventral process associated with median lobe short, bilobed. Parameres present, as long as median lobe.
Females. Integument, head, thorax, abdomen
similar to male except eyes slightly smaller and metaventrite and visible ventrite II evenly rounded, lacking secondary sexual characters.
Distribution
This species is known from localities in NN and BR.
Material examined. NN: (1 male)
New Zealand: NN: Nelson Lakes NP, N slope Mt. Robert 860 m 23-26.iii.80 Nothofagus spp. A. Newton, M. Thayer// berl., leaf and log litter, forest floor (FMNH); (3 specimens) New Zealand NN Taffytown Hill smt nr Little Wanganui 29-Jan-96// In forest litter (JN); (4 specimens) New Zealand NN Fenian Tck Oparara 26-iii-2011// Ground litter, red beech forest (JN); (1 male) L. Rotoiti 2800’ leaf litter 21-3-65 N. A. Walker (NZAC); (4 males, 2 females) New Zealand, NN, Denniston Saddle 10 Feb 1999, R. Leschen R. Hoare// Berlesate Nothofagus forest RL288 41°44′S, 171°48′E (NZAC); (2 males) Rotoiti// A. E. Brookes Collection (NZAC). BR: (1 male, 1 female) New Zealand: BR: 0.8 kmN Bullock Ck nr. Punakaiki 50 m 23.iii.80 broadlf-nikau palm-podocp. A. Newton, M. Thayer// berl., leaf and log litter, forest floor (FMNH); (2 specimens) New Zealand BR Mt Robert nr car park 12-ii-2014// Sifted ground litter, Nothofagus forest (JN); (1 male, 2 females) New Zealand: WD 1.8 km n Punakaiki, 50 m 19.xii.1984-20.i.1985 hdwd.-podo.-nikau for A. Newton/ M. Thayer 719// berl., leaf and log litter, forest floor (FMNH).
Pselaphotumulus dubius (Owens and Carlton), sp. nov.
(, , a–b)
Type material. Holotype: (male)
Doubtful Sound 1578A 5/4 1953 R. Hornabrook (NZAC). Paratypes: (2 males, 4 females) same as holotype locality and collection data (NZAC). The holotype and paratype specimens are deposited in the NZAC.
Etymology
The specific epithet is Latin for ‘doubtful’, referring to Doubtful Sound, the collection locality of the type series.
Diagnosis
Externally, the combination of the metaventrite depressed medially into a glabrous ovate depression and visible ventrite II medially flattened, bearing a small apical tubercle distinguishes this species from all others in the genus. Additionally, the larger enlarged distal area of maxillary palpomere IV (enlarged in distal 1/3 versus 1/4), distinguishes this species from all other Pselaphotumulus except for P. oviceps. The form of the symmetrical aedeagus with paired, symmetrical, branched dorsal processes is also diagnostic.
Description
MALE holotype. Integument: head, pronotum, elytra, abdomen, and appendages light brown, palpi lighter in colour. Head: wide, width between eyes greater than ½ length of head from base of vertex to apex of rostrum. Eyes rounded, composed of 13 facets. Frontal margin of rostrum in dorsal view pointed, with apical patch of modified, flattened setae. Gular area glabrous and mound-like, occupying most of gular area, gular foveae present. Maxillary palpomere IV longer than head, wider at base and distal 1/3, v-shaped sensory patches present. Thorax: pronotum with basal sulcus present, delimited by lateral antebasal foveae; median antebasal fovea present, basolateral foveae present as small depressions. Mesoventrite with median shield bearing patch of spongious setae in small posterior concavity. Metaventrite depressed medially into glabrous ovate depression, posteriorly with fringe of short, suberect setae. Legs unmodified. Elytra with two basal foveae present, single sutural fovea; sutural striae present; apical fringe of setae sparse. Abdomen: tergites of usual form for tribe. Ventrite II slightly flattened medially, apically with small median tubercle. Genitalia: aedeagus symmetrical. Median lobe short, blunt, barely exceeding length of basal bulb. Ventral process slightly ventrally curved, about as long as median lobe. Paired dorsal processes symmetrical, branching into three small distal processes. Parameres present, longer than median lobe.
Females. Integument, head, thorax, abdomen
similar to male except eyes slightly smaller and metaventrite and ventrite II evenly rounded, lacking secondary sexual characters.
Distribution
This species is only known from the type locality in FD.
Pselaphotumulus oviceps (Broun Citation1917), comb. nov.
Pselaphus oviceps (Broun Citation1917: 380)
(, , a–b)
Type material examined
After observing the type series of Pselaphus oviceps in the NHM and reviewing Broun’s original description, we designate the following specimen from the NHM as the lectotype: (1 male) TYPE (red/orange paper)// 3832.♂// New Zealand Broun Coll. Brit. Mus. 1922–482// Routeburn 16-2-1914 (NHM) (designated lectotype). The following three specimens are included in the type series in the NHM and have been determined to represent additional specimens of P. oviceps. We designate these as the paralectotypes: (1 male) New Zealand Broun Coll. Brit. Mus. 1922–482// 3832.♂// Routeburn 16-2-1912// Nov. Gen. oviceps Br. Cl. Besuchet det v 1968 (NHM) (designated paralectotype); (1 male) 3832.// New Zealand Broun Coll. Brit. Mus. 1922–482// Hollyford 19-2-1914 (NHM) (designated paralectotype); (1 female) 3832// Hollyford 19-2-1914// New Zealand Broun Coll. Brit. Mus. 1922–482 (NHM) (designated paralectotype). Lectotype designations are necessary to fix the taxon to a name-bearing type. Broun did not specifically identify type specimens, and many of his syntype series include multiple species. The lectotype and paralectotype specimens are deposited in the NHM.
Diagnosis
Externally, the two lateral patches of spongious setae of the metaventrite distinguish this species from all other Pselaphotumulus except for P. urquharti. The combination of the secondary sexual characters on the metaventrite in combination with the form of medially-flattened ventrite II and apical tubercle of ventrite II are unique. The form of the aedeagus, with paired dorsal processes, each of which possess three branches, in combination with a flattened ventral process that is distally bilobed, is also diagnostic.
Redescription
MALE lectotype. Integument: head, pronotum, elytra, abdomen, and appendages light brown, palpi lighter in colour. Head: wide, width between eyes greater than ½ length of head from base of vertex to apex of rostrum. Eyes rounded, composed of 13 facets. Frontal margin of rostrum in dorsal view pointed, with apical patch of modified, flattened setae. Gular area glabrous and mound-like, occupying most of gular area, gular foveae present. Maxillary palpomere IV longer than head, widened at base and distal 1/3, v-shaped sensory patches present. Thorax: pronotum with basal sulcus present, delimited by lateral antebasal foveae; median antebasal fovea present, basolateral foveae present as small depressions. Mesoventrite with median shield bearing patch of spongious setae in small concavity posteriorly. Metaventrite tumosity with two lateral patches of spongious setae. Legs unmodified. Elytra with two basal foveae present, single sutural fovea; sutural striae present; apical fringe of setae sparse. Abdomen: tergites of usual form for tribe. Ventrite II flattened medially, apically with small median tubercle. Genitalia: aedeagus symmetrical. Median lobe short, distally narrowed. Dorsal processes paired, symmetrical, with three branches. Ventral process flattened, broadly bilobed at apex. Parameres present, paired, slightly longer than dorsal and ventral processes.
Females. Integument, head, thorax, abdomen
similar to male except eyes slightly smaller and metaventrite and visible ventrite II evenly rounded, lacking secondary sexual characters.
Distribution
This species is known only from collection localities in the South Island: NN, BR, WD and FD.
Material examined
NN: (5 males, 2 females) New Zealand: NN: Kahurangi N.P., Mt. Arthur Track, below Mt. Arthur Hut, 1200 m, 28.xi.2005, Nothofagus forest; FMHD#2005-048, berl., dead wood, D. Clarke; ANTM site 1157 95% ethanol Field Museum Nat. Hist. (FMNH EtOH); (3 males, 1 female) New Zealand: NN: N. P. Kahurangi, Cobb Ridge, above Cobb Reservoir, 1050 m, 41°06.351′S, 172∘41.658′E, 29.xi.2005, Nothofagus menzeisii and N. solandri cliffortioides forest; FMHD#2005-055, berl., sifted dryish moss on ground and rocks, A. Newton; ANMT site 1159 95% ethanol Field Museum Nat. Hist. (FMNH EtOH). BR: (1 male, 2 females) New Zealand: NN: Nelson Lakes N.P., Mt. Robert, Speargrass Track, 875 m, 41∘49.469′S, 172∘48.311′E, 17.xii.2005-110, berl., leaf and log litter, A. Solodovnikov, D. Clarke et al.; ANMT site 1161 95% ethanol Field Museum Nat. Hist. (FMNH EtOH). WD: (2 males, 1 female) New Zealand: WD 3.2 km NE Haast, 14 m Haast River Walk 43′52S 169′ 03E, #072// Nothofagus/Podocarp forest leaf litter berlese, 17 Jan 1998 (LSAM); (2 males, 1 female) New Zealand: WD 2 km SE Fox Glacier 225 m, #066 43′29S 170′01E// Weinmannia/Metrosideros leaf litter berlese, 16 Jan 1998 (LSAM); (1 male) New Zealand WD Taipo River Rd Taramakau Vly// sifted ground moss berlesate (JN). FD: (1 male) New Zealand: FD: Hidden Falls, Lower Hollyford R. Litter, 6 Jan 1967 A. K. Walker, 67/3 (LSAM); (3 males, 3 females) New Zealand: FD: Hollyford Tr, #109 44′42S 168′08 E/ Nothofagus coastal forest flood debris 22 Jan 1998 C. Carlton, R. Leschen (LSAM); (1 male slide-mounted, 1 female) New Zealand: FD: Hollyford Tr, #110 44′42S 168′08E/ Nothofagus coastal forest leaf litter berlese, 22 Jan 1998 C. Carlton, R. Leschen (LSAM); (2 males) New Zealand: FD: L. Howden Fork Key Summit, 720 m, litter 14 Dec 1966, 66/522 A. K. Walker, K. Z. Wilson (LSAM); (1 male) New Zealand: FD: Milford Rd Tutoko River, #112 44′41S 167′56E/ Coastal forest leaf litter berlese, 22 Jan 1998 C. Carlton, R. Leschen (LSAM); (1 male) New Zealand: FD: Milford Sound Litter, 1 Nov 1966 J. I. Townsend, 66/359 (LSAM); (1 male) New Zealand: FD: Murchison Mts., Point Burn, litter, 11 Dec 1972 A. C. Eyles, 72/272 (LSAM); (1 female) New Zealand: FD: Wilmot Pass, 400 m Litter, 23 Jan 1970 J. I. Townsend, 70/145 (LSAM); (3 males, 2 females) New Zealand: FD: Wilmot Pass, 450 m Litter, 23 Jan 1970 J. I. Townsend, 70/73 (LSAM); (1 male) New Zealand: FD: Wilmot Pass, 900 m, litter, 23 Jan 1970 G. W. Ramsay, 70/75 (LSAM); (1 male, 1 female) New Zealand: South Is. Fiordland, Hollyford Tr. Nothofagus/coastal forest flood debris, 20 Jan 1998 C. Carlton, R. Leschen, #109 (LSAM); (2 males) New Zealand FD Bauza Is. Doubtful Sd March 1986// Pitfall trap G. M. Bremner (JN). (2 males) New Zealand FD 400 m S of Borland Sdl 28 Jan 2011// Washed soil sample beach forest (JN); (2 males) Hollyford Valley, N. of L. Wakikarou Southland// T. Hall 19.2.1914 (NZAC).
Pselaphotumulus unus (Owens and Carlton), sp. nov.
(, , a–b)
Type material examined. Holotype: (1 male)
New Zealand: NC, 3 km N Arthur’s Pass, 900 m 42′55S 171′33E, #042// Nothofagus/Dracophyllum leaf litter Berlese, 11 Jan 1998 C. Carlton, R. Leschen (NZAC). The holotype is deposited in the NZAC.
Etymology
The specific epithet ‘unus’ is derived from the Latin word for ‘one’, referring to the presence of a single median longitudinal patch of spongious setae on the metaventrite of males of this species.
Diagnosis
Externally, the combination of the metaventrite with a single median, longitudinal patch of spongious setae and ventrite II medially impressed with two short carinae in distal 1/3 distinguishes this species from all others in the genus. Additionally, the aedeagus of P. unus is unique among species in the genus in the combination of a median lobe with the distal end narrowed and curved upwards, and a ventral process sharply narrowed and curved downwards, distally.
Description
MALE holotype. Integument: head, pronotum, elytra, abdomen, and appendages light brown, palpi lighter in colour. Head: wide, width between eyes greater than ½ length of head from base of vertex to apex of rostrum. Eyes rounded, composed of 13 facets. Frontal margin of rostrum in dorsal view pointed, with apical patch of modified, flattened setae. Gular area glabrous and mound-like, occupying most of gular area, gular foveae present. Maxillary palpomere IV longer than head, wider at base and distal 1/4, v-shaped sensory patches present. Thorax: pronotum with basal sulcus present, delimited by lateral antebasal foveae; median antebasal fovea present, basolateral foveae present as small depressions. Mesoventrite with median shield bearing patch of spongious setae in small concavity posteriorly. Metaventrite bearing single median patch of spongious setae. Legs unmodified. Elytra with two basal foveae present, single sutural fovea; sutural striae present; apical fringe of setae sparse. Abdomen: tergites of usual form for tribe. Visible ventrite II shallowly impressed medially, with two short carinae in distal 1/3. Genitalia: aedeagus asymmetrical. Median lobe with distal end narrowed and curved upwards. Ventral process sharply narrowed and curved downwards, distally, about as long as median lobe. Parameres present, as long as median lobe.
Females. Integument, head, thorax, abdomen
similar to male except eyes slightly smaller, metaventrite and visible ventrite II evenly rounded, and lacking secondary sexual characters.
Distribution
Pselaphotumulus unus is known from the type locality in NC, and several localities in WD.
Material examined
NC: (1 male, 1 female) New Zealand: NC: Arthur’s Pass NP, Bealey Vy. Tr. 840 m 18-21.iii.80 sub-alpine Nothofagus A. Newton, M. Thayer// berl., leaf and log litter, forest floor (FMNH). WD: (2 males, 2 females) New Zealand: WD: L. Mahinapua Scen. Res. 30 m 16-22.iii.1980 podocarp-mixed broadleaf A. Newton, M. Thayer// berl., leaf and log litter (FMNH). (2 males, 1 female) New Zealand: WD: Hokitika R. Gorge, S Kowhitirangi 100 m 17.iii.1980 podocarp-braodlf. A. Newton, M. Thayer// berl., leaf and log litter, forest floor (FMNH). (2 males, 3 females) New Zealand: WD Okuku Scenic Reserve, 9.2 km sse Kumara, 120 m, 8-19.i.1985 podocarp-hdwd. Forest A. Newton/ M. Thayer 731;// berl., leaf and log litter, forest floor (FMNH).
Pselaphotumulus urquharti (Broun Citation1917), comb. nov.
Pselaphus urquharti (Broun Citation1917: 379)
(, , a–b)
Type material examined
After observing the type series of Pselaphus urquharti in the NHM and reviewing Broun’s original description, we designate the following specimen from the NHM as the lectotype: (1 male) TYPE (red/orange paper)// 3831.♂// Oakden 23-9-1913// New Zealand Broun Coll. Brit. Mus. 1922–482 (NHM) (designated lectotype). The following three specimens are included in the type series in the NHM and have been determined to represent additional specimens of P. urquharti. We therefore designate these as paralectotypes: (1 male) 3831.♂// New Zealand Broun Coll. Brit. Mus. 1922–482// Oakden 23-9-1913 (NHM) (designate paralectotype); (1 male) 3831// New Zealand Broun Coll. Brit. Mus. 1922–482// Algidus 25-9-1913 (NHM) (designate paralectotype); (1 female) 3831// New Zealand Broun Coll. Brit. Mus. 1922–482// Oakden 23-9-1913 (NHM) (designate paralectotype). Lectotype designations are necessary to fix the taxon to a name-bearing type. Broun did not specifically identify type specimens, and many of his syntype series include multiple species. Lectotype and paralectotype specimens are deposited in the NHM.
Diagnosis
This species may be distinguished from all others in the genus except P. oviceps by the presence of two lateral patches of spongious setae on the metaventrite. This secondary sexual character, in combination with the presence of two median lateral carinae on visible ventrite II, is unique among Pselaphotumulus species. This species may also be identified by the form of the aedeagus, with both the ventral process and the median lobe simple, straight.
Redescription
MALE holotype. Integument: head, pronotum, elytra, abdomen, and appendages light brown, palpi lighter in colour. Head: wide, width between eyes greater than ½ length of head from base of vertex to apex of rostrum. Eyes rounded, composed of 13 facets. Frontal margin of rostrum in dorsal view pointed, with apical patch of modified, flattened setae. Gular area glabrous and mound-like, occupying most of gular area, gular foveae present. Maxillary palpomere IV longer than head, widened at base and distal 1/4, v-shaped sensory patches present. Thorax: pronotum with basal sulcus present, delimited by lateral antebasal foveae; median antebasal fovea present, basolateral foveae present as small depressions. Mesoventrite with median shield bearing patch of spongious setae in small concavity posteriorly. Metaventrite tumosity with modified spongious setae set into parallel sulci. Legs unmodified. Elytra with two basal foveae present, single sutural fovea; sutural striae present; apical fringe of setae sparse. Abdomen: tergites of usual form for tribe. Visible ventrite II slightly flattened medially, bearing two lateral carinae along margin of medial area. Genitalia: aedeagus symmetrical. Median lobe short, internal elements extending above sclerotised shelf. Ventral process simple, about as long as median lobe. Parameres present, paired, about as long as median lobe and ventral process.
Females. Integument, head, thorax, abdomen
similar to male except eyes slightly smaller and metaventrite and visible ventrite II evenly rounded, lacking secondary sexual characters.
Distribution
This species has been collected from localities in BR, NC, WD, MC, MK and FD.
Material examined. BR: (1 male) New Zealand: BR: Paparoa Ra., 929 m, L. nr Mt. Priestly, litter, 69/241 10 Dec 1969, J. McBurney (LSAM); (1 male) Pororari R. Westland 17-3-49// A. E. Brookes Collection (NZAC); (1 male, 1 female) Hokitika R. Gorge, S Kowhitirangi 100 m 17.iii.1980 podocarp-broadlf. A. Newton, M. Thayer/ berl., leaf and log litter, forest floor (FMNH). NC: (1 female) New Zealand: NC: Arthur’s Pass Devil’s Punch Bowl Litter, 16 Nov 1966 A. K. Walker, 66/412 (LSAM). (1 female) New Zealand: NC: Arthur’s Pass, McGrath Ck Moss, 12 May 1965 J. I. Townsend, 65/314 (LSAM). (2 females) New Zealand: NC, 2.9 km N Arthur’s Pass, 840 m Bealey Valley Trailhead 42′55S 171′33E, #036/ Nothofagus solandri leaf litter berlese 11 Jan 1998 C. Carlton, R. Leschen (LSAM). (1 male) New Zealand: NC, 3 km N Arthur’s Pass, 900 m 42′55′S 171′33E, #042/ Nothofagus/Draco-phyllum leaf litter berlese, 11 Jan 1998 C. Carlton, R. Leschen (LSAM). (1 male, 2 females) New Zealand: NC, 4.5 km SE Arthur’s Pass 725 m, #046 42′55S 171′33E/ Nothofagus solandri leaf litter/flood debris 11 Jan 1998 C. Carlton, R. Leschen (LSAM); (1 male, 1 female) Arthur’s Pass NP, Bealey Vy. Tr. 840 m 10-21.iii.80 sub-alpine Nothofagus A. Newton, M. Thayer/ berl., leaf and log litter, forest floor (FMNH); (1 male, 1 female) Okuku Ck., 11.3 km SSE Kumara 60 m 18-22.iii.1980 podocarp-broadlf. A. Newton, M. Thayer/ berl., leaf and log litter, forest floor (FMNH). WD: (2 males, 3 females) Okuku Scenic Reserve, 9.2 km sse Kumara, 120 m, 8-19.i.1985 podocarp-hdwd. Forest A. Newton/ M. Thayer 731/ berl., leaf and log litter, forest floor (FMNH); (3 males, 1 female) New Zealand: WD 2 km SE Fox Glacier 225 m, #066 43′29S 170′01E (LSAM). (2 males, 1 female) New Zealand: WD, 2.7 km S Franz Josef Glacier,#063 Trailhead of Alex Knob 43′25S 173′10E, 150 m (LSAM); (1 male) New Zealand: WD Ross Mikonui Tk 9 Nov 2005 R. Leschen, S. Nomura// ants in wood RL1015 (NZAC).; (2 males, 2 females) L. Mahinapua Scen. Res. 30 m 16-22.iii.1980 podocarp-mixed broadleaf A. Newton, M. Thayer/ berl., leaf and log litter, forest floor (FMNH). MC: (1 male) Oakden nr. Mt. Algidus Canterbury// 3831// Coll. T. Hall 23.9.1914 (NZAC). (1 male) Algidus 3-12-13// 3831// Broun (NZAC). MK: (1 male) Hooker Valley, Mt. Cook Southern Alps// Leaf mould// Collector Jan. 1943. A. E. Brookes (NZAC). FD: NEW ZEALAND: FD: Fiordland N.P., Borland Rd., Borland Saddle vic., 980 m, 45∘44.81′S, 167∘22.88′E, 10.xii.2005, Nothofagus menziesii forest in ravine; FMHD#2005-090, berl., leaf and log litter, A. Newton; ANMT site 1171 95% ethanol Field Museum Nat. Hist. (FMNH EtOH); (5 males, 3 females) New Zealand: FD: Fiordland N.P., Milford Sound Rd., Tutoko Track, 70 m, 44∘40.517′S, 167∘57.895′E, 9.xii.2005, very mossy Nothofagus spp. Forest, emergent podocarps; FMHD#2005-086, berl., leaf and log litter, A. Newton/M. Thayer; ANMT site 1168 95% ethanol Field Museum Nat. Hist. (FMNH EtOH); (6 males, 3 females) New Zealand: FD: Fiordland N.P., Milford Sound Rd., Gertrude Saddle car park area, 800 m, 44∘46.13′S, 168∘00.16′E, 9.xii.2005, very mossy Nothofagus menziesii forest; FMHD#2005-087, berl., leaf litter, A. Newton/M. Thayer; ANMT site 1169 95% ethanol Field Museum Nat. Hist. (FMNH EtOH); New Zealand: FD: Fiordland N.P., Borland Rd., Borland Saddle vic., 980 m, 45∘44.81′S, 167∘22.88′E, 10.xii.2005, Nothofagus menziesii forest in ravine; FMHD#2005-091, berl., sifted? Sphagnum moss on ground, A. Newton; ANMT site 1171 95% ethanol Field Museum Nat. Hist. (FMNH EtOH); (3 females) New Zealand: FD: Fiordland N.P., Borland Rd., Mt. Burns Tops Track, above Borland Saddle, 1075 m, 45∘44.883′S, 167∘22.938′E, 10.xii.2005, subalpine tussockland; FMHD#2005-097, berl., unsifted litter at bases of shrubs (Dracophyllum, possibly filifolium), M. Thayer; ANMT site 1173 95% ethanol Field Musuem Nat. Hist. (FMNH EtOH); (1 male, 1 female) New Zealand: FD: Fiordland N.P., Borland Rd., Mt. Burns Tops Track, above Borland Saddle, 1035 m, 45∘44.82′S, 167∘22.87′E, 10.xii.2005, short ridgetop Nothofagus menziesii forest, near timberline; FMHD2005-098, berl., twiggy and rooty leaf and log litter, M. Thayer, ANMT site 1174 95% ethanol Field Museum Nat. Hist. (FMNH EtOH). (3 males, 6 females) New Zealand: FD: Fiordland N.P., Kepler Track, above Broad Bay, 600 m, 45∘24.37′S, 167∘39.105′E, 11.xii.2005, Nothofagus forest; FMHD#2005-102, berl., leaf and log litter, A. Solodovnikov/D. Clarke; ANMT site 1177 95% ethanol Field Museum Nat. Hist. (FMNH EtOH). (1 male) Hollyford leafmould (NZAC); (1 male) Key Summit 4/2/54 (NZAC).
Discussion
Biogeographic considerations
The limited information available concerning distributional relationships and patterns of Pselaphotumulus species suggests presumptive endemism along the main northeastern and southwestern slopes of the Southern Alps. With the addition of molecular data and increased geographic coverage, we believe Pselaphotumulus may be an interesting candidate for further exploration of the application of hypothesis regarding the factors influencing distributions of taxa with shared biogeographic histories in the South Island region of New Zealand. Similar distributions are exemplified by many other plant and animal species, such as other Coleoptera taxa, including genera of Carabidae (Johns Citation2005), Zopheridae (Marske et al. Citation2011) and euasthetine Staphylinidae (Clarke Citation2011). These distributions are hypothesised to be either the result of strike slip displacements on either side of the Alpine fault (Heads Citation1998; Heads and Craw Citation2004) or due to habitat restrictions resulting from complex local interactions influenced by climate and geological substrates, most notably Pleistocene glacial cycles (Wardle Citation1988).
The former tectonic vicariance model is suggested by sister group relationships that pair taxa displaced from each other to the northeast on the western side of the Alpine fault and to the southwest on the eastern side. Such disjunctions along the Alpine fault are found in over 200 plant and animal taxa (Heads Citation2017), including several members of the supertribe Faronitae (Staphylinidae: Pselaphinae): the genus Leschenea (Park and Carlton Citation2015) and Sagola turretensis (Park and Carlton Citation2014). While many taxa exhibit disjunct distributions, a number of others exhibit either continuous distributions from north to south or distributions that are localised in the less diverse central region of South Island. A notable feature of this central area is the ‘beech gap’, a region of forest in Westland that is devoid of most species of Nothofagus, which are common in other areas of South Island native forests (Leschen et al. Citation2008). These central distributions are typically hypothesised to be the result of range expansions and contractions via either several large (Willett Citation1951) or numerous smaller microrefugia (Mee and Moore Citation2014) during Pleistocene glacial cycles. In any case, such climatically and geologically influenced allopatry would reflect fine scale, mainly habitat-based allopatry, and should not show obvious concordance with the large-scale geological history of the region. Shared patterns resulting from Alpine climatic fluctuations and the disruptive effects of sea level rise at lower elevations might be revealed through character-based and deeper node molecular analyses and population level genetic divergence data.
Typical for many South Island taxa, Pselaphotumulus species appear to be mainly distributed along the northwestern and southwestern portions of the South Island, with concentrations in the forested regions of Nelson and Fiordland. Pselaphotumulus aorerei, P. cavelli, P. dubius and P. oviceps exhibit this distributional displacement and are absent in the drier, central regions of South Island. Of the two remaining species, P. urquharti has a more or less continuous distribution across western South Island, with a conspicuous absence in Nelson. P. unus appears to be present in the beech gap, with additional specimens in the Alpine region of Arthur’s Pass.
Generally, distributions consistent with classic Alpine fault endemism with populations that extend into Fiordland are thought to either (1) predate the Miocene orogeny of the Southern Alps (Winkworth et al. Citation2005), persisting during mountain building, or (2) have diversified relatively late during Alpine mountain building and subsequent strike slip displacements (Trewick and Wallis Citation2001; Heads and Craw Citation2004). Additionally, the presence of a South Island gap in some groups is thought to be consistent with the phenomenon of the restriction of previously widespread populations due to glaciation or Alpine uplift, constrained to several large refugia on the north and south ends of the South Island (Willett Citation1951). Conversely, the existence of species within the beech gap may be consistent with the theory of the existence of multiple microrefugia with metapopulations persisting through glaciations in small ecological islands (Heads Citation1994; Mee and Moore Citation2014; Buckley et al. Citation2015). Such distributions could also be indicative of the subsequent dispersal of populations back into previously glaciated areas following glacial retreats.
Despite examination of pselaphine specimens from major museum collections representing a number of collection localities across New Zealand, Pselaphotumulus specimens are still known only from a relatively small number of localities. As with many litter-associated organisms, this taxon holds promise for further investigation of larger-scale questions, but more extensive sampling across the South Island is necessary. Additionally, it is important to note that many South Island species share congeners on the North Island, providing additional reference points for understanding species diversification and dispersal, and discovery of Pselaphotumulus species on the North Island would not be unexpected. It remains to be seen whether further sampling provides additional specimens and increases geographic coverage. The collection of DNA quality material across the genus’ range is necessary for further exploration of broader questions.
Notes on symmetry of genitalia
Pselaphotumulus, as a genus, is unconventional among most other Pselaphinae in the inclusion of species that possess both asymmetrical and symmetrical genitalia. Within the subfamily, symmetry patterns of genitalia are generally considered to be characters conserved among genera and have been used to distinguish genera that are otherwise superficially similar (Kurbatov and Sabella Citation2008). While this phenomenon of varying symmetry among species is known to occur among some other genera (Chandler Citation1991), the vast majority of genera in the subfamily do not display this level of genitalic polymorphism. Conventional ideas about the evolution of the aedeagus presume symmetrical forms to be plesiotypic. This implies that asymmetrical forms arise from either single or independent origins within the genus. Within Pselaphotumulus, only two species, P. aorerei and P. unus, possess asymmetrical genitalia. Determination of origins of this asymmetry requires detailed homology of aedeagal structures, which is complicated by the highly derived and complex form of the morphology of male genital structures in Pselaphinae. Such analysis is beyond the scope of this paper, but may prove an interesting case study in future explorations of the evolution of variations within these complex sexual organs.
Acknowledgements
We thank the curators at museums and institutions, especially Victoria Bayless (LSAM), Rich Leschen (NZAC) and Max Barclay (NHM) for providing feedback and access to specimens. We especially thank John T. Nunn (JTN) for access to valuable specimens from his extensive personal collection of New Zealand Pselaphinae.
Disclosure statement
No potential conflict of interest was reported by the authors.
References
- Broun T. 1893. Descriptions of new Coleoptera from New Zealand. Annals and Magazine of Natural History (6th series). 12:163–195.
- Broun T. 1917. Descriptions of new genera and species of Coleoptera, part V. New Zealand Institute Bulletin. 1:347–474. John Mackay, Wellington.
- Buckley TR, Krosch M, Leschen RAB. 2015. Evolution of New Zealand insects: summary and prospectus for future research. Austral Entomology. 54:1–27. doi: 10.1111/aen.12116
- Chandler DS. 1991. The Lucifotychus of Eastern North America (Coleoptera: Pselaphidae). Psyche. 98:47–56. doi: 10.1155/1991/79759
- Chandler DS. 2001. Biology, morphology and systematics of the ant-like litter beetle genera of Australia (Coleoptera: Staphylinidae: Pselaphinae). In: Gupta Virendra, editor. Memoirs on entomology international Vol. 15. Gainesville (FL): Associated Publishers; p. 8–9.
- Clarke DJ. 2011. A revision of the New Zealand endemic rove beetle genus Agnosthaetus Berhnhauer (Coleoptera: Staphylinidae). The Coleopterists Bulletin Monograph. 10:1–118.
- Crosby TK, Dugdale JS, Watt JC. 1998. Area codes for recording specimen localities in the New Zealand subregion. New Zealand Journal of Zoology. 25:175–183. doi: 10.1080/03014223.1998.9518148
- Heads M. 1998. Biogeographic disjunction along the alpine fault, New Zealand. Biological Journal of the Linnean Society. 63:161–176. doi: 10.1111/j.1095-8312.1998.tb01512.x
- Heads M. 2017. Biogeography and evolution in New Zealand. Boca Raton (FL): Taylor & Francis/CRC. 635 pp.
- Heads MJ. 1994. Biogeography and biodiversity in Hebe, a South Pacific genus of Scrophulariacea. Candollea. 48:19–60.
- Heads M, Craw R. 2004. The alpine fault biogeographic hypothesis revisited. Cladistics. 20:184–190. doi: 10.1111/j.1096-0031.2004.00009.x
- Johns PM. 2005. Field guide to South Island carabid beetles of conservation interest (Coleoptera: Carabidae). Christchurch (NZ): Department of Conservation. 1–74 pp.
- Kurbatov SA, Sabella G. 2008. Revision of the genus Atychodea Reitter with a consideration of the relationships in the tribe Tychini (Coleoptera, Staphylinidae, Pselaphinae). Transactions of the American Entomological Society of America. 134:23–68. doi: 10.3157/0002-8320(2008)134[23:ROTGAR]2.0.CO;2
- Leschen RA, Buckley TR, Harman HM, Shulmeister J. 2008. Determining the origin and age of the Westland beech (Nothofagus) gap, New Zealand, using fungus beetle genetics. Molecular Ecology. 17:1256–1276. doi: 10.1111/j.1365-294X.2007.03630.x
- Lord NP, Leschen RAB. 2014. Illustrated catalogue and type designations of the New Zealand Zopheridae (Coleoptera: Tenebrionidae). Zootaxa. 3809:1–127. doi: 10.11646/zootaxa.3809.1.1
- Marske KA, Leschen RAB, Buckley TR. 2011. Reconciling phylogeography and ecological niche models for New Zealand beetles: Looking beyond glacial refugia. Molecular Phylogenetics and Evolution. 59:89–102. doi: 10.1016/j.ympev.2011.01.005
- Mee JA, Moore J-S. 2014. The ecological and evolutionary implications of microrefugia. Journal of Biogeography. 41:837–841. doi: 10.1111/jbi.12254
- Newton AF, Chandler DS. 1989. World catalog of the genera of Pselaphidae (Coleoptera). Fieldiana: Zoology. 53:1–93.
- Nomura S, Leschen RAB. 2006. Faunistic review on the Pselaphinae species known from New Zealand (Insecta, Coleoptera, Staphylinidae). National Science Museums Monographs. 34:239–272.
- Park J-S, Carlton CE. 2014. Pseudostenosagola, a new genus from New Zealand (Coleoptera: Staphylinidae: Pselaphinae: Faronitae). Annals of the Entomological Society of America. 107:734–739. doi: 10.1603/AN14025
- Park J-S, Carlton CE. 2015. Pseudoexeirarthra, a new genus from New Zealand (Coleoptera, Staphylinidae, Pselaphinae), with descriptions of seven new species. ZooKeys. 491:95–118. doi: 10.3897/zookeys.491.9164
- Trewick SA, Wallis GP. 2001. Briding the “beech gap”: New Zealand phylogeography implicates Pleistocene glaciation and Pliocene isolation. Evolution. 55:2170–2180.
- Wardle P. 1988. Effects of glacial climates on floristic distribution in New Zealand 1. A review of the evidence. New Zealand Journal of Botany. 26:541–555. doi: 10.1080/0028825X.1988.10410660
- Willett RW. 1951. The New Zealand Pleistocene slow line, climactic conditions, and suggested biological effects. New Zealand Journal of Science and Technology. 1:18–48.
- Winkworth RC, Wagstaff SJ, Glenny D, Lockhart PJ. 2005. Evolution of the New Zealand mountain flora: origins, diversification and dispersal. Organisms, Diversity & Evolution. 5:237–247. doi: 10.1016/j.ode.2004.12.001