
ABSTRACT
Deforestation, increasing urban development and associated infrastructure have resulted in widespread loss of roosting habitat for New Zealand’s long-tailed bat (Chalinolobus tuberculatus, pekapeka-tou-roa). As a result, artificial roosts are increasingly proposed as a short to medium-term mitigation measure, despite the absence of specific protocols for the installation, maintenance, and suitability for New Zealand bats. As of 2021, more than 110 artificial roosts had been installed in urban parks within Hamilton City, New Zealand. To determine their uptake and suitability for long-tailed bats, 74 Kent style roosts of varying ages were monitored for 1 year, with 24 (32%) roosts observed occupied by long-tailed bats. Occupation was generally infrequent, with no clear seasonal pattern or specific areas of occupation. However, two older roosts were consistently occupied, with numbers notably increasing during the summer maternity season. Distance to stand edge was the only factor that could be used to predict occupation, with logistic regression indicating that long-tailed bats may prefer roosts in less exposed areas with stable microclimates, within tree stands or sheltered landscape features. Future artificial roost installations should site roosts away from the stand edge in sheltered locations, ideally near natural or already occupied roosts to facilitate discovery and uptake.
Introduction
Globally, deforestation and land use change have been identified as major drivers of declining bat populations through the loss of roosting and foraging habitat (Mering and Chambers Citation2014; Weier et al. Citation2019). Short to medium-term mitigation of roost habitat loss has favoured the implementation of artificial roosts, also known as ‘bat boxes’ (Rueegger Citation2016). Bat boxes vary in design with the most common being rectangular ‘Kent’ style wooden boxes which consist of two or three stepped chambers or a single large chamber, which are accessed by narrow slots at the bottom (Rueegger Citation2016; Arias et al. Citation2020). Cylindrical, ‘Schwegler’ style boxes constructed from woodcrete™ with a single opening one-third of the way up the chamber have also been widely employed (Rueegger Citation2016). However, previous bat box programmes have rarely considered roost microclimate characteristics during the design and implementation of bat boxes, resulting in highly generalised guidelines concerning installation, maintenance, and reported suitability (Mering and Chambers Citation2014; Rueegger Citation2016; Griffiths et al. Citation2017). For example, lesser long-eared bats (Nyctophilus geoffroyi) had historically low rates of bat box use in voluminous-type boxes but were found to use boxes with narrow fissure-type cavities (15-20 mm) at a high frequency (Rueegger et al. Citation2020). This design had not been previously employed in Australia which may explain the limited use of bat boxes by this species, as maternity colonies generally select roosts with narrow-fissure type entrances (Rueegger et al. Citation2020; Lumsden et al. Citation2021). Effective provision of bat boxes for mitigation purposes requires targeting and understanding species-specific roost preferences.
Fragmented populations of the critically threatened long-tailed bat (Chalinolobus tuberculatus, pekapeka-tou-roa) inhabit forest remnants across both islands of New Zealand, as well as urban and peri-urban habitat in central North Island cities such as Hamilton, Rotorua, and Auckland (O'Donnell et al. Citation2018; Dumbleton and Montemezzani Citation2020; O'Donnell et al. Citation2021). Long-tailed bats are selective in their choice of roost trees based on their functional characteristics such as topography, tree size, presence of suitable hollows, and degree of surrounding vegetation, making them particularly vulnerable to roost habitat loss by tree felling for intensive land use (O'Donnell and Sedgeley Citation1999). While bat boxes have been proposed as partial mitigation for the loss of roost trees (AECOM Citation2019; Wildlands Citation2020), research concerning the suitability of bat boxes for New Zealand long-tailed bats has been minimal, and guidance on their preferred design and installation characteristics has not been thoroughly developed (Smith et al. Citation2017; Davidson-Watts Citation2019; O’Sullivan Citation2021; AECOM Citation2022).
Long-tailed bats have consistently been detected in Hamilton City’s southern gully systems and riparian margins of the Waikato River, with infrequent detections in the central, northern, and western parts of the city (Le Roux and Le Roux Citation2012; Dekrout et al. Citation2014; Mueller et al. Citation2017; van der Zwan and Mueller Citation2019; Dumbleton and Montemezzani Citation2020; Aughton Citation2021). Beginning in 2011, approximately 30 bat boxes, primarily of Kent design (), were installed in several of Hamilton City’s southern parks by Project Echo, an advocacy group for bats in the Waikato Region, in order to encourge roosting and enhance bat activity within the city boundary (Robinson Citation2022). These boxes remained largely unmonitored, and there were no reports of occupation until an individual was radio-tracked to a Sandford Park box in 2018 (Davidson-Watts Citation2019). In response to concerns from conservation advocates over the loss of long-tailed bat habitat from the Southern Links Roading Project (AECOM Citation2019), a further 80 artificial roosts (70 Kent, 10 Schwegler; ) were installed within the southern parks of Hamilton City during the summer of 2019–2020.
Figure 1. Examples of typical bat boxes installed in green spaces of Hamilton City. Top row: Older (10 + years) single chamber (left) and triple chamber (right) Kent style boxes installed by Project Echo. Bottom row: A newer (2 + years) double chamber Kent box (left) and a Schwegler style box (right) installed by Hamilton City Council. Schwegler boxes were not included in the study due to the low number of boxes and difficulties with visually determining occupation by bats.

The installation of bat boxes in Hamilton City provided an opportunity to measure the uptake of bat boxes by long-tailed bats and assess their general suitability for roosting. The monitoring also sought to determine if there were specific areas of occupancy in the city, seasonal differences in occupancy, or environmental factors that influenced occupancy. Habitat surveys and geospatial analyses were undertaken to determine which factors could be used to reliably predict bat box occupancy. This research is intended to inform management practices regarding bat box use in New Zealand.
Materials and methods
Study area
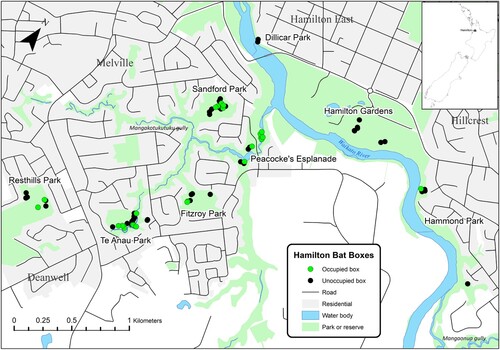
Hamilton City (11,080 ha) is located in the central North Island of New Zealand within the highly modified Hamilton Ecological District (159,376 ha) (Clarkson and McQueen Citation2004; Cornes et al. Citation2012). The Waikato River is a key landscape feature bisecting the city, with four major gullies extending from the river into the city, covering an area of approximately 750 ha (Clarkson and McQueen Citation2004; Le Roux and Le Roux Citation2012). The gullies are incorporated into several parks, planted with a mixture of native and exotic vegetation. A total of nine urban green spaces, with bat boxes installed either by Hamilton City Council or Project Echo, were included in this study (). The mean distance between the centroid of each group of boxes was 2.5 km, with seven of the green spaces in the southern part of the city connected by the Mangakotukuku gully system and the Waikato River. Donny Park was the only location situated in the northern precincts of Hamilton City, while Resthills Park was disconnected from the gully system by State Highway 3 and residential housing. Vegetation at each location predominately consisted of a mix of mature exotic and native tree species such as pines (Pinus radiata), Eucalyptus spp., lacebark (Hoheria populnea), and blackwood (Acacia melanoxylon) (Cornes et al. Citation2012).
Figure 2. Locations of the monitored bat boxes across eight parks in the southern suburbs of Hamilton City, New Zealand. Note that Donny Park is not presented as this location had a single unoccupied bat box.
Bat box monitoring
An initial survey of 118 bat boxes within Hamilton City and its peri-urban surrounds was conducted in March 2021. All boxes were installed on tree trunks 6–18 m above ground with predator proof metal banding above and below the box. Boxes with features such as cobwebbed entrances and an absence of faecal staining on the entrance, were deemed unlikely to be regularly utilised by bats and were excluded from further study. In addition, boxes that could not be safely accessed and those where the number of bats could not be reliably determined, such as the Schwegler boxes were also excluded. The remaining 74 bat boxes (11 project echo boxes and 63 newer Hamilton City Council boxes) were monitored once every 3 months from May 2021 for bat occupation using infrared (IR) cameras (Equinox Z2, Bushnell) for a 12-month period. Occupation was determined either by direct observation of bats within the bat box using the IR camera or by positioning the camera at the base of the tree and video recording the box entrance from sunset to 1-hour post-sunset. The video footage was then reviewed using Windows Media Player (Microsoft, v. 12) and the total number of bats roosting within the box was recorded. If bats were observed occupying a box, the monitoring frequency was increased to every 4-weeks. Covid-19 restrictions meant that the monthly monitoring of 22 bat boxes was delayed by 1 month from August to September 2021.
Habitat survey and geospatial analysis
Seventeen variables were initially selected for their potential influence on bat box occupation, factors such as number of chambers, estimated overstory density, distance to stand edge, percentage overlap with vegetation, terrain slope, hill shade, terrain elevation, minimum distance to water and aspect were included as they may influence roost habitat environmental and microclimatic conditions (Rueegger Citation2016). Minimum distance to the nearest building and minimum distance to streetlighting were used as a proxy for artificial light at night, which has been found to result in habitat avoidance in species such as the little forest bat (Vespadelus vulturnus), and delay the emergence time of some bat species, such as Geoffroy’s bat (Myotis emarginatus) (Boldogh et al. Citation2007; Haddock et al. Citation2019). The time taken for bats to use boxes has been found to vary considerably, and it has been suggested that discovery of new roosts is time consuming, and that new roosts are more likely to be discovered if they are near existing roosts (Ruczyński et al. Citation2009). Therefore, age of the box, distance to occupied roosts and nearest neighbouring box were included as variables. The tree diameter at breast height (DBH) each bat box was mounted on were recorded as there is evidence that tree age plays a role in roost selection by long-tailed bats, although the likelihood of cavity formation appears to be the main driver (Sedgeley and O'Donnell Citation1999, Citation2004). Mounting height of boxes was also included as a potential factor of influence, although previous studies have yet to establish clear evidence of box height preferences in bats (Rueegger Citation2016).
The approximate age of each bat box (years), species of tree each box was mounted on, and the number of internal chambers were provided by Hamilton City Council. Bat box mounting heights (m) were measured with either a laser distance measurer or a hypsometer (Haglöf, Sweden Vertex 5) if there was obstructing vegetation. Aspect was determined by a hand-held GPS receiver (Garmin, 64s), while distance to stand edge (m), defined as the distance each bat box was from the edge of the stand, and tree diameter at breast height (cm) (DBH) were measured in the field. Estimated overstory density (%) was calculated with a spherical convex densiometer (Forestry Suppliers, model 43887).
Geospatial analysis was conducted with QGIS (version 3.16.16) using shapefiles for building footprints, street light points, and vegetation provided by Hamilton City Council. The plugin ‘nearest-neighbour join’ was used to determine the closest distance (m) between bat boxes to buildings, streetlights, and surface water. To determine the minimum distance from a bat box to flowing water, New Zealand River topographic 1:150k polygons were converted to lines and the layer was merged with New Zealand topographic 1:150k river centrelines. Distance was then determined by the nearest neighbour analysis. The proportion of vegetative cover (%) within a 100 m radius of each bat box was calculated by applying a 100 m buffer around each bat box, which was then converted to a polygon shapefile. This layer was overlaid with the vegetation layer and the percentage overlap was determined using the plugin ‘Overlap Analysis Tool’. Terrain elevation (m), slope (degrees), and hill shade (relative scale) were determined from the Waikato 1 m LiDAR digital elevation model (DEM) file obtained from the LINZ data service. After merging to create a single raster geotiff file, terrain elevation for each site was determined by the plugin ‘Point Sampling Tool’ using the coordinates for each bat box. Similarly, slope of each site was calculated using the slope feature and extracted from the DEM using the point sampling tool. It was assumed that bat boxes were more likely to be occupied during summer as previous monitoring of selected Hamilton bat boxes recorded increased bat activity and box occupancy during this period (O’Sullivan Citation2021; GT unpublished data). Therefore, hill shade was calculated for the summer solstice at 3 pm when air temperature was likely to be at its maximum. It was assumed that this would represent the period of thermal maxima within each box, with variations in microclimatic conditions occurring between boxes due to differences in direct sun exposure, resulting in bats selecting boxes with optimal thermal conditions. This resulted in an azimuth of 295° and a vertical angle (altitude of the light) of 63° used to calculate hill shade with a Z factor of 1. Hill shade at each bat box was then extracted from the DEM using the ‘Point Sampling Tool’.
Statistical analysis
Analyses were conducted in GraphPad (Prism, v. 9.4). To test for independence of covariates, a correlation matrix was performed to exclude variables with correlation coefficients >0.7 as per Pschonny et al. (Citation2022). As correlation could only be computed between continuous variables, box aspect was omitted from this analysis. Instead, a chi square test was conducted to determine the likelihood of north facing boxes being occupied was different to random chance. Additionally, because most boxes were deliberately installed north facing, a Fisher’s exact test was conducted to determine if north-facing boxes were preferentially occupied. Analysis regarding the species of tree each box was mounted on was not performed, as O'Donnell and Sedgeley (Citation1999) concluded that long-tailed bats do not discriminate between tree species for potential roosts.
Multiple logistic regression with no interaction effects was conducted to determine if any of the variables could be used to reliably predict bat box occupation. The multiplicative effect of the odds ratios is considered more useful when interpreting the results of logistic regression because the parameter estimates illustrate how much the log odds change when the predictor variable changes (Peng et al. Citation2002). With each unit increase of a predictor variable, the odds of bat box occupation are multiplied by the value of its odds ratio. Odds ratios >1 indicate that when all other factors are held constant, with each unit increase of the predictor the likelihood of observed occupation increased. Conversely, predictors with odds ratios <1 indicate that with each unit increase of the predictor the odds of bat box occupation decreased. In addition, observations by O’Sullivan (Citation2021) one-month prior to the commencement of this research were incorporated into the logistic regression analysis to increase the sample size, this increased the total number of observed occupied boxes by one.
Results
Bat box occupation
Over the 12-month period, 24 (32%) of the monitored bat boxes were occupied by long-tailed bats, across six different parks (). Occupation was generally sparse and too infrequent to discern seasonal patterns from graphed results, with 21 (28%) boxes used by <5 bats at any one time and 15 (62.5%) of the boxes were occupied only once. Of the 63 newer Hamilton City Council boxes, 21 (33%) were occupied, with five boxes first observed to be in use during June 2021, approximately 18 months after installation. Comparatively, only three (27%) of the older project echo boxes were observed as occupied. However, the two older triple-chambered boxes in Sandford Park were consistently occupied, with the number of bats ranging from 9–29 over autumn, winter, and spring, and 30–53 bats over the summer (November–February), when they were used as maternity roosts, as confirmed by the observation of pups. A newer box in Peacockes Road Esplanade was also utilised as a maternity roost in December 2021, consisting of ten adults and five pups, but was unoccupied in subsequent observations.
Table 1. A total of 74 bat boxes were monitored across nine green spaces in Hamilton City, the percentages of occupied and unoccupied boxes, and the total number of long-tailed bats observed from May 2021–May 2022.
Habitat survey and geospatial analysis
Fifteen variables either quantified from field surveys or geospatial analysis, were initially selected for their potential influence on bat box occupation (). In addition, aspect of mounted box was analysed for the 74 monitored boxes, 74.3% were installed facing either cardinal north or northeast. A chi-square test indicated that the likelihood of north facing boxes being occupied was not statistically different to random chance (Chi-square, df = 1, P = 0.35). Similarly, a Fisher’s exact test determined there was no significant difference in occupation between all northern facing and southern facing boxes (Fisher’s two-tailed test, P = 0.74).
Table 2. Descriptive statistics of the 15 continuous variables selected for their potential to influence bat box occupation by long-tailed bats in Hamilton City.
A correlation matrix was performed on the 15 continuous variables to test for independence of covariates. Due to skewness in the data distribution, minimum distance to occupied roosts and distance to nearest neighbouring box were log natural transformed prior to analysis. Using a threshold value of 0.7, hill shade and terrain slope were classified as negatively correlated, while minimum distance to buildings from roost and minimum distance to streetlights from roost were positively correlated. Hill shade and minimum distance to buildings were excluded from further analysis, as the calculation of hill shade was based on a hypothetical light source, and building lights were considered less consistent sources of illumination compared to streetlights.
Multiple logistic regression was conducted to determine if any of the 13 remaining variables could be used to predict occupation. Distance to stand edge was the only significant (n = 71, df = 57, P < 0.027) predictor of box occupation (). The model deviance was 64.1 which was lower than the reference intercept model’s deviance of 90.8. From the classification table, the model correctly classified unoccupied boxes 91.5% of the time, whilst occupied boxes were correctly classified 50% of the time. This is supported when observed occupied and unoccupied box groups were plotted against the corresponding predicted values from the logistic model. The model did not perform as well when classifying the occupied boxes, as the predicted probabilities are more uniformly distributed (mean 0.55, median 0.52) compared to the unoccupied boxes (mean 0.22, median 0.22) (). The area under the receiver operating characteristic (ROC) curve was 0.83 indicating that the overall predictive performance of the model was reasonable (Pearce and Ferrier Citation2000). The odds ratio estimates of terrain elevation, distance to occupied roosts, distance to nearest neighbouring box, number of chambers, and distance to stand edge indicated that they had comparatively the largest influence on bat box occupation per unit change. However, the model found that most variables’ ability to predict occupation was no better than zero ().
Figure 3. Plotted observed versus predicted probability for Hamilton bat box occupation based on multiple logistic regression modelling of 13 variables with box occupation as the dependant variable. The more uniform distribution of points for the observed occupied boxes indicates that the model performed poorly in classifying these boxes.
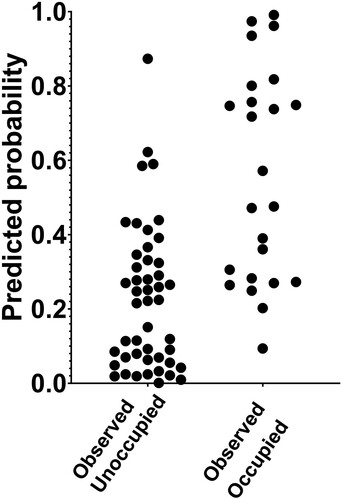
Table 3. Odds ratios from the multiple logistic regression with occupation status as the dependent variable and 13 independent variables selected as potential predictors of bat box occupation.
Discussion
Mitigation for lost bat roosting habitat has favoured the implementation of bat boxes, as they have been perceived as immediate substitutes for roost loss compared to more long-term exotic or native tree plantings which require substantial time to develop suitable roost cavities (Le Roux et al. Citation2016; Rueegger Citation2016; Rueegger et al. Citation2020). It is increasingly likely that bat boxes will be adopted for urban development projects in New Zealand. However, bat boxes have yet to be established as effective supplementary roosts due to substandard landscape placement or unsuitable box designs (Whitaker et al. Citation2006; Rueegger et al. Citation2020). The aim of this study was to determine the uptake of bat boxes at nine Hamilton City parks and identify factors which could influence occupation by long-tailed bats. Of the 74 monitored bat boxes, c. 32% were occupied infrequently, with no clear seasonal patterns of occupation. This is not unexpected as long-tailed bats are known to frequently switch roosts to meet changing social and thermoregulatory needs, inhibit ectoparasites, and reduce the energetic costs of commuting to foraging grounds (Lewis Citation1995; O'Donnell and Sedgeley Citation1999; Reckardt and Kerth Citation2007). In addition, despite observations of >50 bats in one roost (O’Sullivan Citation2021, current study) the Hamilton long-tailed bat population is unlikely to be abundant as the surrounding landscape has been highly modified to urban and agricultural land use, resulting in limited and fragmented habitat, which was reflected in the low box occupation rates. While the current study attempted to balance monitoring frequency with number of monitored sites, it is acknowledged that the monitoring frequency likely resulted in missed detections of bats in boxes. A parallel study of the newer Hamilton City bat boxes utilised a different combination of survey techniques including thermal imagery, box endoscope surveys, and eDNA, detecting the presence of long-tailed bats at 33 of 80 boxes (AECOM Citation2022). The most effective method of detection was eDNA with 25 positive indications, and accounted for the overall difference of nine detections between the two studies (AECOM Citation2022). However, eDNA provides an indication of past occupancy rather than a real-time measure, it may also represent a brief visitation rather than actual roosting activity. Factors such as predation risk, availability of natural roosts and box microclimate may also influence box occupation but were not examined in this study (Sedgeley Citation2001; Goldingay and Stevens Citation2009; Pschonny et al. Citation2022). Overall, the uptake of the newer Hamilton City Council boxes occurred more rapidly than similar overseas studies would indicate (Mering and Chambers Citation2014; Rueegger Citation2016). The age and previous use of the two original Kent style Project Echo boxes in Sandford Park (installed 2012) likely facilitated the uptake of nearby newer boxes, as previous exposure to bat boxes has been found to corresponded with quicker uptake (Rueegger Citation2016). The two Sandford Park boxes were first observed to be occupied in 2018 and have consistently been used as maternity roosts since that time (Davidson-Watts Citation2019; O’Sullivan Citation2021).
Fidelity to roosts that provide suitable environmental conditions for reproduction is adaptive, as it reduces the costs of searching for alternative roosts with the required qualities (Humphrey Citation1975; Lewis Citation1995). Breeding females require warm, thermally stable microclimates to facilitate foetal growth and conserve energy while remaining normothermic, as torpor in breeding females will delay foetal development or inhibit milk production (Humphrey Citation1975; Sedgeley Citation2001; Bergeson et al. Citation2021). Communally roosting females can increase roost temperatures by 5–10°C compared to unoccupied roosts, reducing the need for torpor (Sedgeley Citation2001; Borkin and Parsons Citation2011; Hałat et al. Citation2020). In contrast, solitary roosting males and subadults may tolerate a wider range of thermal conditions as they can more readily enter torpor (Sedgeley Citation2001; Borkin and Parsons Citation2011). An assessment of factors that may facilitate suitable maternal roosts was outside the scope of this study. However, two and three chambered Kent style bat boxes appear to be of sufficient size and configuration to facilitate clustering, allowing breeding females and pups to derive energetic benefits from social warming (Sedgeley Citation2001; Hałat et al. Citation2020). Further investigation of the microclimatic conditions in bat boxes used as communal or solitary roosts should be undertaken to better understand the requirements of long-tailed bat maternity colonies. This is of increased importance for urban colonies, where the relative sparseness of mature trees can further restrict the availability of suitable maternity roosts (Dekrout Citation2009).
Multiple logistic regression was conducted to identify variables which could be used to predict box occupation. Of the thirteen analysed variables, distance to stand edge was the only statistically significant variable, with the model odds ratio indicating that the chance of a box being occupied increased with distance from the stand edge. This suggests that bat boxes should be sited in the interior of tree stands rather than on stand fringes or exposed ridge sites. Boxes installed on exposed marginal features may be more subject to environmental extremes, whereas sheltered roosts are likely to have more stable microclimates, allowing greater energy conservation during winter and avoiding overheating during summer (Hamilton and Barclay Citation1994; Borkin and Parsons Citation2011; Hoeh et al. Citation2018). As a result, aspect may have little influence on box occupation in sheltered areas, explaining why there was little difference in occupation rate between northern and southern facing boxes. However, sex-specific roost selection has been identified in long-tailed bats, and box aspects that provide ample sun exposure may be an important factor to consider for maternity roosts in more southern areas of New Zealand (Borkin and Parsons Citation2011; Tillman et al. Citation2021).
Although the remaining variables were not statistically significant, odds ratio estimates of minimum distance to occupied roosts and nearest neighbouring box indicated that box occupation may decrease with increasing distance. Long-tailed bats prefer to roost in clusters of boxes, as this facilitates their fission-fusion roosting behaviour and enables them to choose boxes that suit their social and thermoregulatory needs (Sedgeley and O'Donnell Citation1999; O'Donnell et al. Citation2021). The greater proportion of newer double-chambered Hamilton City Council boxes may also account for the negative influence of number of chambers on box occupation as previous studies suggest that multiple large chambers would be beneficial as they enable clustering, and may provide a temperature range within the box that allows bats to move to areas with suitable temperatures throughout the day (Sedgeley Citation2001; Rueegger Citation2019). Hoeh et al. (Citation2018) found that three-chambered bat boxes on warm clear days were warmer at the top of the chambers than the bottom, as the wood boards likely trapped heat near the top of the roost. Although it should be noted that the bats were excluded from the roosts during temperature recordings and boxes were installed on the edge of wooded areas with no canopy closure (Hoeh et al. Citation2018).
The findings from this study reflect a sample of bat boxes that were clustered within a relatively limited geographic extent in Hamilton City. Different behavioural responses and habitat preferences may exist in other geographic areas of New Zealand. The relatively limited distribution of the Hamilton bat boxes means that observations are unlikely to be completely independent, with box occupancy potentially resulting in reduced chance of nearby boxes being occupied. In addition, the initial exclusion of boxes deemed unlikely to be occupied for monitoring purposes, although necessary for logistical reasons, may have resulted in some bias and led to an underestimation of factors that make a bat box a desirable roost. Similarly, due to monitoring frequency, missed detections of bats in boxes resulted in falsely identifying a box as unoccupied. Future research will need to address these limitations by employing multiple survey methods to ensure an accurate depiction of factors that influence box occupation by long-tailed bats is gathered. Nonetheless, future installations of bat boxes for long-tailed bats should consider placing boxes within sheltered tree stands or landscape features such as gullies which provide protection from weather extremes creating a more thermally stable roost environment. New roosts should also ideally be installed near natural or already occupied roosts to facilitate discovery and uptake. Bat boxes which fail to provide suitable conditions, especially during the maternity season, may result in the loss of local bat populations following the removal of natural roosts. It is therefore critical that future research investigates boxes used as maternity roosts to understand the factors that influence their occupation so these can be applied to box installation guidelines in New Zealand.
Acknowledgements
The authors are grateful for the support from many people in preparing this journal article. We would like to thank the enthusiastic volunteers and those that provided invaluable advice including Aman Sharma, Katherine Rowe, Sarah Endres, and Kerry Borkin.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- AECOM. 2019. Environmental management and monitoring plan (EMMP) Southern Links Project - Hamilton City Council section. Client report prepared for Hamilton City Council and New Zealand Transport Agency by AECOM New Zealand Limited. p. 68–70.
- AECOM. 2022. Bat box monitoring report 2022 Southern Links Project, Hamilton. Client report prepared for Hamilton City Council by AECOM New Zealand Limited. p. 17–19.
- Arias M, Gignoux-Wolfsohn S, Kerwin K, Maslo B. 2020. Use of artificial roost boxes installed as alternative habitat for bats evicted from buildings. Northeastern Naturalist. 27(2):201–214. doi:10.1656/045.027.0203.
- Aughton H. 2021. Project Echo 2021 Hamilton City wide bat survey. Report prepared by Project Echo. 7–8.
- Bergeson SM, Brigham RM, 'Keefe O, M J. 2021. Free-ranging bats alter thermoregulatory behaviour in response to reproductive stage, roost type, and weather. Journal of Mammalogy. 102(3):705–717. doi:10.1093/jmammal/gyab049.
- Boldogh S, Dobrosi D, Samu P. 2007. The effects of the illumination of buildings on house-dwelling bats and its conservation consequences. Acta Chiropterologica. 9(2):527–534. doi:10.3161/1733-5329(2007)9[527:TEOTIO]2.0.CO;2.
- Borkin KM, Parsons S. 2011. Sex-specific roost selection by bats in clearfell harvested plantation forest: Improved knowledge advises management. Acta Chiropterologica. 13(2):373–383. doi:10.3161/150811011X624848.
- Clarkson BD, McQueen JC. 2004. Ecological restoration in Hamilton City, North Island, New Zealand. In Proceedings of the 16th International Conference, Society for Ecological Restoration, August 24-26, 2004, Victoria, Canada.
- Cornes TS, Thomson RE, Clarkson BD. 2012. Key ecological sites of Hamilton City: Volume 1. Client report prepared for Hamilton City Council by Centre for Biodiversity and Ecology Research, School of Science, University of Waikato. p. 5-19.
- Davidson-Watts I. 2019. Long-tailed bat trapping and radio tracking baseline report 2018 and 2019. Client report prepared for AECOM by Davidson-Watts Ecology (Pacific) Ltd. p. 28-32.
- Dekrout A. 2009. Monitoring New Zealand long-tailed bats (Chalinolobus tuberculatus) in urban habitats: ecology, physiology, and genetics [PhD thesis]. Auckland: University of Auckland.
- Dekrout AS, Clarkson BD, Parsons S. 2014. Temporal and spatial distribution and habitat associations of an urban population of New Zealand long-tailed bats Chalinolobus tuberculatus. New Zealand Journal of Zoology. 41(4):285–295. doi:10.1080/03014223.2014.953551.
- Dumbleton H, Montemezzani W. 2020. Hamilton City long tailed bat survey annual monitoring report 2019-2020. Client reported prepared for Project Echo by 4Sight Consulting Ltd. p. 2-10.
- Goldingay RL, Stevens JR. 2009. Use of artificial tree hollows by Australian birds and bats. Wildlife Research. 36(2):81–97. doi:10.1071/WR08064.
- Griffiths SR, Bender R, Godhino LN, Lentini PE, Lumsden LF, Robert KA. 2017. Bat boxes are not a silver bullet conservation tool. Mammal Review. 47(4):261–265. doi:10.1111/mam.12097.
- Haddock JK, Threlfall CG, Law B, Hochuli DF. 2019. Light pollution at the urban forest edge negatively impacts insectivorous bats. Biological Conservation. 236:17–28. doi:10.1016/j.biocon.2019.05.016.
- Hałat Z, Dechmann DKN, Zegarek M, Ruczyński I. 2020. Male bats respond to adverse conditions with larger colonies and increased torpor use during sperm production. Mammalian Biology. 100(6):611–620. doi:10.1007/s42991-020-00071-5.
- Hamilton IM, Barclay RMR. 1994. Patterns of daily torpor and day-roost selection by male and female big brown bats. Canadian Journal of Zoology. 72(4):744–749. doi:10.1139/z94-100.
- Hoeh JPS, Bakken GS, Mitchell WA, 'Keefe O, M J. 2018. In artificial roost comparison, bats show preference for rocket box style. Plos One. 13(10):e0205701.
- Humphrey SR. 1975. Nursery roosts and community diversity of Neartic bats. Journal of Mammalogy. 56(2):321–346. doi:10.2307/1379364.
- Le Roux D, Ikin K, Lindenmayer D, Bistricer G, Manning A, Gibbons P. 2016. Effects of entrance size, tree size and landscape context on nest box occupancy: considerations for management and biodiversity offsets. Forest Ecology and Management. 366:135–142. doi:10.1016/j.foreco.2016.02.017.
- Le Roux D, Le Roux N. 2012. Hamilton City bat survey 2011-2012. Unpublished report prepared for Project Echo by Kessels and Associates. p. 2-15.
- Lewis SE. 1995. Roost fidelity of bats: A review. Journal of Mammalogy. 76(2):481–496. doi:10.2307/1382357.
- Lumsden LF, Griffiths SR, Silins JE, Bennett AF. 2021. Roosting behaviour and the tree-hollow requirements of bats: Insights from the lesser long-eared bat (Nyctophilus geoffroyi) and Gould's wattled bat (Chalinolobus gouldii) in south-eastern Australia. Australian Journal of Zoology. 68(6):296–306. doi:10.1071/ZO20072.
- Mering ED, Chambers CL. 2014. Thinking outside the box: a review of artificial roosts for bats. Wildlife Society Bulletin. 38(4):741–751. doi:10.1002/wsb.461.
- Mueller H, Ulrich C, Purcell A. 2017. Hamilton City long-tailed bat survey 2016–2017. Client report prepared by Kessels Ecology for Project Echo.
- O'Donnell C, Borkin K, Christie J, Lloyd B, Parsons S, Hitchmough R. 2018. Conservation status of New Zealand bats, 2017 (New Zealand Threat Classification Series, 21). New Zealand Department of Conservation.
- O'Donnell C, Borkin K, Parsons S. 2021. Families Vespertilionidae and Mystancinidae (King CM, Forsyth DM, editors. 3rd ed.). Dunedin: Otago University Press.
- O'Donnell C, Sedgeley J. 1999. Use of roosts by the long-tailed bat, Chalinolobus tuberculatus, in temperate rainforest in New Zealand. Journal of Mammalogy. 80(3):913–923. doi:10.2307/1383260.
- O’Sullivan A. 2021. Limitations of locating long-tailed bat Chalinolobus tuberculatus roosts with acoustic recorders in an urban park, Hamilton, New Zealand. Unpublished report, School of Science, University of Waikato. p. 8-24.
- Pearce J, Ferrier S. 2000. Evaluating the predictive performance of habitat models developed using logistic regression. Ecological Modelling. 133(3):225–245. doi:10.1016/S0304-3800(00)00322-7.
- Peng CYJ, Lee KL, Ingersoll GM. 2002. An introduction to logistic regression analysis and reporting. The Journal of Educational Research. 96(1):3–14. doi:10.1080/00220670209598786.
- Pschonny S, Leidinger J, Leitl R, Weisser WW. 2022. What makes a good bat box? How box occupancy depends on box characteristics and landscape-level variables. Ecological Solutions and Evidence, 3(1). doi:10.1002/2688-8319.12136.
- Reckardt K, Kerth G. 2007. Roost selection and roost switching of female Bechstein's bats (Myotis bechsteinii) as a strategy of parasite avoidance. Oecologia. 154(3):581–588. doi:10.1007/s00442-007-0843-7.
- Robinson H. 2022. The uptake of artificial roosts by long-tailed bats (Chalinolobus tuberculatus) in Hamilton City. [Master's thesis]. Hamilton: University of Waikato, Hamilton. p. 98.
- Ruczyński I, Kalko EK, Siemers BM. 2009. Calls in the forest: a comparative approach to how bats find tree cavities. Ethology. 115(2):167–177. doi:10.1111/j.1439-0310.2008.01599.x.
- Rueegger N. 2016. Bat boxes—a review of their use and application, past, present and future. Acta Chiropterologica. 18(1):279–299. doi:10.1111/j.1439-0310.2008.01599.x.
- Rueegger N. 2019. Variation in summer and winter microclimate in multi-chambered bat boxes in eastern Australia: potential eco-physiological implications for bats. Environments. 6(2):13. doi:10.3390/environments6020013.
- Rueegger N, Goldingay R, Law B, Gonsalves L. 2020. Testing multichambered bat box designs in a habitat-offset area in eastern Australia: Influence of material, colour, size and box host. Pacific Conservation Biology. 26(1):13–21. doi:10.1071/PC18092.
- Sedgeley J. 2001. Quality of cavity microclimate as a factor influencing selection of maternity roosts by a tree-dwelling bat, Chalinolobus tuberculatus, in New Zealand. Journal of Applied Ecology. 38(2):425–438. doi:10.1046/j.1365-2664.2001.00607.x.
- Sedgeley J, O'Donnell C. 1999. Factors influencing the selection of roost cavities by a temperate rainforest bat (Vespertilionidae: Chalinolobus tuberculatus) in New Zealand. Journal of Zoology. 249(4):437–446. doi:10.1111/j.1469-7998.1999.tb01213.x.
- Sedgeley J, O'Donnell C. 2004. Roost use by long-tailed bats in South Canterbury: examining predictions of roost-site selection in a highly fragmented landscape. New Zealand Journal Ecology. 28(1):1–18.
- Smith D, Borkin K, Jones C, Lindberg S, Davies F, Eccles G. 2017. Effects of land transport activities on New Zealand's endemic bat populations: reviews of ecological and regulatory literature. NZ transport Agency research report. 249.
- Tillman FE, Bakken GS, O'Keefe JM. 2021. Design modifications affect bat box temperatures and suitability as maternity habitat. Ecological Solutions and Evidence. 2(4). doi:10.1002/2688-8319.12112.
- van der Zwan W, Mueller H. 2019. Hamilton City long-tailed bat survey, 2018-2019. Annual Monitoring Report prepared for Project Echo. 11–15.
- Weier SM, Linden VMG, Grass I, Tscharntke T, Taylor PJ. 2019. The use of bat houses as day roosts in macadamia orchards South Africa. PeerJ. 7:e6954. doi:10.7717/peerj.6954.
- Whitaker JO, Sparks DW, Brack Jr V. 2006. Use of artificial roost structures by bats at the Indianapolis International Airport. Environmental Management. 38(1):28–36. doi:10.1007/s00267-005-0117-2.
- Wildlands. 2020. Bat management plan for Gleeson Quarry, Huntly. Contract report No. 5208e prepared for Gleeson Quarries Huntly Ltd and Gleeson Managed Fill Ltd by Wildland Consultants Ltd. p. 9-13.