Abstract
Three new species: Grandicrepidula hemispherica (Nukumaruan, S Hawke's Bay), Pelicaria granttaylori (Mangapanian-early Nukumaruan, Wanganui – Hawke's bay), Pelicaria arahura (Waipipian-early Mangapanian, Westland and Hawke's Bay). Drawings of marine species in Smith's (1874) three plates of New Zealand molluscan types are republished. Further Australian molluscs in Wanganui Basin: Sabia australis (Lamarck), Clanculus plebejus (Philippi), both early Nukumaruan. Further northern New Zealand mollusc in Wanganui Basin: Stephopoma roseum (Quoy & Gaimard), OIS 13, 9. Distinctive gastropods extinct at end Nukumaruan: Struthiolaria frazeri (Hutton), Taxonia suteri (Marwick). Taniella planisuturalis (Marwick) (Opoitian-Nukumaruan, southern NZ) and Trivia (Ellatrivia) zealandica (Kirk) (Nukumaruan, Hawke's Bay-Wanganui; Castlecliffian, North Canterbury) occur in Castlecliffian (OIS 15?) rocks at Whakatane. Cantharidella tessellata (A. Adams) and Risellopsis varia (Hutton), formerly Haweran, are recorded from Nukumaruan and Castlecliffian rocks, respectively. New fossil late Nukumaruan-early Castlecliffian records listed from Mikonui-1 offshore well, Westland, include Malluvium calcareum (Suter) and 10 other species. Other biostratigraphically useful gastropods: Calliostoma (Maurea) nukumaruense (Laws) (Mangapanian-OIS 17); Argobuccinum pustulosum (Lightfoot), Semicassis labiata (Perry) (both earliest in OIS 7). New synonymy: Zeacumantus perplexus (Marshall & Murdoch) = Z. lutulentus (Kiener); Pelicaria vermis (Martyn) = all named Nukumaruan-Recent forms (other than P. rugosa (Marwick) and P. granttaylori n. sp.); Trivia flora Marwick = T. zealandica Kirk. Taxonomy revised: Zelippistes benhami (Suter) (OIS 13 & 9 at Wanganui), distinguished from Lippistes and Separatista; Stiracolpus species, informally; Maoricrypta profunda (Hutton), Waipipian-early Castlecliffian (- OIS 19); M. radiata (Hutton) (= incurva Zittel, = hochetteriana Woods, = wilckensi Finlay), (Middle Miocene?) Tongaporutuan-Opoitian; Eunaticina cincta (Hutton), holotype illustrated.
Introduction
This paper is the third part of a revision of the taxonomy and biostratigraphy of New Zealand fossil Mollusca of the last two million years, with comments on related Recent and earlier Pliocene taxa. The first report (Beu Citation2004) reconsidered the generic positions or specific determinations of some New Zealand late Neogene to Recent molluscs, including migrants from eastern Australia and the tropical Pacific that appeared only briefly in New Zealand during interglacial periods. The second (Beu Citation2006) reconsidered time ranges and taxonomy of bivalves over this period, including further warm-water migrants. The present report is a reconsideration of the time ranges and taxonomy of some gastropods over the same period, with time ranges recorded in oxygen isotope stages (OIS) where possible. Because of space considerations, a number of further gastropods requiring revision have been deleted from this work (partially listed in Part 4). A similar paper on “higher” Gastropoda (Ptenoglossa, Neogastropoda and Heterobranchia) and a summary paper are in press. Time ranges have been compiled from the detailed faunal lists provided for the formations of Wanganui Basin by Fleming (Citation1953), assigned to individual oxygen isotope stages in the correlation diagram of cycles in Wanganui Basin compiled by Carter & Naish (Citation1999). The faunas included in this overview, their ages and their oxygen isotope stage correlations were summarized in Part 1 of this series of papers (Beu Citation2004), modified slightly in the introduction to Part 2 (Beu Citation2006), and are not repeated here. Once again this work is founded on the seminal works on Wanganui Basin by Fleming (Citation1947, 1953), and much of what the present ‘Wanganui Basin working group’ has published (particularly Beu Citation2004, Citation2006; Abbott et al. Citation2005; Naish et al. Citation2005; Pillans et al. Citation2005) is based on Fleming's brilliant foundation.
Fleming (Citation1953) listed a number of possibly new taxa in his faunal lists from Wanganui Pliocene–Pleistocene formations, based on material in GNS. Some of Fleming's material is surprising for temperature and migration interpretations, particularly the records (below) of Sabia and Clanculus from early Nukumaruan rocks in Opawe Stream, Pohangina valley, in eastern Wanganui Basin outside Fleming's (1953) mapped area. However, many of the putative new taxa listed by Fleming (Citation1953) are based on very few specimens, and possibly are undescribed but require confirmation from more material (e.g. species he assigned to Scissurella, Nobolira, Scrobs and Aupouria). Other potential new species now seem not to be different from Recent species that are more variable than Fleming realized. A single Pervicacia specimen with a much larger protoconch than Recent ones is a typical case of more material being required to confirm the characters of an apparently unnamed species (GS4057, R22/f6388, Kaikokopu Shellbed, OIS 19, Castlecliff coastal section). Still others, such as the several new species of Nuculidae that Fleming intended to name, are well founded but would be better to revise on the basis of the Recent fauna, where all have proved to be common (BA Marshall NMNZ pers. comm.). They have not been included in these papers. The section on Gastropoda has been divided into two papers, and a biostratigraphy section on gastropods, similar to that presented in the earlier parts 1 and 2 (Beu 2004, 2006), will be included in part 4, the second gastropod part.
Smith's (1874) illustrations of New Zealand Mollusca
One of the most useful but now hard to obtain of early publications on New Zealand Mollusca is EA Smith's (Citation1874) three plates illustrating type specimens in the Natural History Museum, London (part of the British Museum in 1874) of common New Zealand shallow-water Mollusca. The illustrations seem to have been prepared at an earlier date, apparently for JE Gray before the publication of Gray (1843). Smith's (1874:1) introduction to a brief text of only seven pages stated: ‘The four following excellent plates will be very acceptable to Conchologists, as they contain figures of several species (about twenty) which, until the present time, have been known only by descriptions, in some instances very short and insufficient, and also the more interesting as, in thirty-one instances, the actual type specimens are delineated. [New line] Plates 1 to 3 are devoted exclusively to species belonging to the New Zealand fauna. Four of them are new to science, and three, described some years since, are unrecorded in Von Martens’ [1873] very useful “Critical list of the Mollusca of New Zealand”. [New line] Plate 4 includes, with one exception (Helix menkeana), figures of some Australian Land and Freshwater Mollusca of which two appear to be as yet undescribed. [New line] All the species with the exception of Bulimus antipodum and Vermetus cariniferus are in the British Museum, and those preceded by a * were presented by Sir John Richardson, M.D., F.R.S.’. Nearly all the terrestrial snails are Australian; none of the non-marine taxa illustrated by Smith is included here. The figures of marine molluscs on the first three of Smith's (1874) plates are republished here, scanned from the published originals, approximately as oriented by Smith but rearranged, to make them more widely available. A few small figures on the plates are unnumbered and are not referred to in Smith's text or captions, and are not included here.
Determining which of Smith's illustrations show type specimens is complicated, as only 25 of the captions state ‘Type’, ‘Type figured’, or ‘Type in the British Museum’, rather than the 31 stated by Smith. All types are still present in the Natural History Museum, London. Therefore, it is assumed in the captions presented here that almost all New Zealand species, and certainly Smith's four new species, are illustrated from type material. The status of the specimens labelled ‘type’ should be treated with care, in view of ICZN Recommendation 73F, ‘avoidance of assumption of holotype’. In many cases the figured specimen is known to be one of several syntypes, and in others some original syntypes might now be lost. All are, therefore, considered to be syntypes.
Some comments are necessary on some of the drawings, or on some of the names used by Smith (1874) in his accompanying text.
-
Smith's (1874, pl. 1, ) illustration (M) is identified as ‘Trichotropis clathrata (A Adams Ms.) Hab. New Zealand (Dr Sinclair, Colonel Bolton and Dr Lyall, RN). The above name is attached to a specimen in the Cumingian collection, without any locality, which agrees in every particular with specimens from New Zealand collected by the gentlemen quoted above. The name appears in Messrs. Adams’ list in the “Genera of recent Mollusca” vol. I., p. 280; but as I cannot find a description of any species so designated, I append the following:’ [description follows]. Smith (1874:4) noted that ‘The figure representing this species is not characteristic, for it is taken from a young example, and the form of the mouth is very different from that of the adult shell’. Smith's illustration evidently is of a different species from the adult syntype, as the illustration shows Xymene plebeius (Hutton, 1873) (Muricidae). Suter (Citation1913:296, pl. 44, fig. 9) adopted T. clathrata in place of the prior T. inornata Hutton, 1873, the common Pliocene–Recent species usually known as Trichosirius inornatus, on the grounds that Hutton's species had not been illustrated, an unacceptable criterion even in 1913. He did not mention the Xymene illustration. Even Iredale (Citation1915:456), when reinstating Hutton's name, did not mention the misidentified illustration. Suter (Citation1913:296) clarified the situation by attributing the name T. clathrata to GB Sowerby II (Citation1874). Sowerby's (1874, pl. 2, ) illustration is at least of a Trichotropis (sensu lato) species, even if not very clearly of T. inornatus—the columella is too strongly excavated in Sowerby's drawing. The specimen illustrated by Sowerby (1874, pl. 2, ) is here designated the lectotype of Trichotropis clathrata, to avoid this name being attached to Xymene plebeius. This publication (August 1874) is accepted here as earlier than Smith's (1874, undated as to month), and the name is a later synonym of Trichosirius inornatus. Smith (1874) or an earlier draughtsman evidently illustrated Xymene plebeius in error. Petit (2009:97) confirmed the listing of Trichotropis clathrata by H Adams & A Adams (1854:280), ‘where it is a nude name’.
-
Vermetus cariniferus Gray, Citation1843 (Smith 1874, pl. 1, figr. 23) (T) is based on a polychaete tube rather than a gastropod, the abundant intertidal zone-forming tube-worm formerly known as Pomatoceros caeruleus (Schmarda, 1861), currently known as Spirobranchus cariniferus (Gray, 1843). This illustration presumably shows a syntype.
-
The limpet illustration captions are muddled; both Patella redimiculum Reeve, 1854 and Patella denticulata Martyn, 1784 are labelled in the text as applying to Smith's (1874, pl. 1, Fig. 24) (V). Smith's (1874, pl. 1, Fig. 25) (X) illustration is labelled ‘Patella antipodum, sp. n.,’ and Smith's (1874, pl. 1, Fig. 26) (S) illustration has no caption or text, but clearly shows Cellana ornata (Dillwyn, 1817). Pl. 1, Fig. 24 (V) shows the southern New Zealand species Cellana strigilis (Hombron & Jacquinot, 1841), of which P. redimiculum was ranked a subspecies by Powell (Citation1979:45) and Goldstien et al. (Citation2006). Apparently the illustration of Cellana ornata (Smith's Fig. 26) (O) was misidentified as P. denticulata. Suter (Citation1913:79) followed a suggestion by WH Dall in using the name ‘Helcioniscus antipodum’ for the eastern Australian species previously known as Patella tramoserica Martyn, 1784 (now consistently known as Cellana tramoserica [Holten, 1802]; Wilson Citation1993:36, pl. 1, figr. 23; Lindberg Citation1998:647, .36C, D; pl. 22 1, 2; Goldstien et al. Citation2006). However, Smith (Citation1915:78) stated that H. antipodum (Smith) is a synonym of Cellana radians (Gmelin, Citation1791).
-
The drawing of Vanganella taylori (Smith 1874, pl. 2, ; B) stated by Smith to be of a ‘full-grown’ specimen, not the immature ‘type specimens presented by the Rev. R. Taylor to the British Museum’) shows the species now universally known as Resania lanceolata Gray, 1853. J-P Rocroi (MNHN pers. comm. from P Bouchet, 17 April 2008) stated that Gray (Citation1853a), where the name Resania lanceolata was established, was published in January 1853, whereas Gray (Citation1853b), where the name Vanganella taylori was established, was published on 29 April 1853; R. lanceolata has priority.
-
Venus oblonga Gray, 1843 is illustrated (Smith 1874, pl. 2, ) (C), presumably a syntype despite the lack of such a label, whereas the earlier synonym Dosina zelandica Gray, Citation1835 seems at first sight not to be illustrated, despite Smith (1874:6) noting that ‘the type’ is in the British Museum. However, presumably as a result of a typographical error, an apparently unlabelled figure (Smith 1874, pl. 3, ) (E) is referred to within the third line of Smith's (1874:6) synonymy as if the reference were within Gray's (1835) publication; this illustration clearly represents a syntype of Dosina zelandica. As Smith (1874) noted, it is a short specimen of the same species as Venus oblonga, and the earlier name for this species.
-
Paphies ventricosa (Gray, 1843): Smith's (1874, pl. 3, ) illustration (K) shows the remaining syntype, identified as ‘Taria ventricosa’, BMNH 1842.5.17.189, ‘North shore, Cook's Strait, New Zealand’, that is, the Paraparaumu–Waikanae coast of western Wellington Province (where it can still be collected), collected by Dieffenbach. The specimen was illustrated as the ‘holotype’ by Beu & de Rooij-Schuiling (1982, –24) but this does not constitute designation of a lectotype under ICZN Article 74.6, as it has not been accepted that a single type specimen exists. Comparison of this figure with Beu & de Rooij-Schuiling's (1982, –24) photographs demonstrated that Smith's drawing is inaccurate, the posterior end being shown shorter and the umbonal-ventral ridge more prominent than they actually are, although other details of the hinge and growth irregularities around the posterior end agree well.
Fig. 1 Reproductions of illustrations of New Zealand marine molluscan type specimens, from Smith (1874), at published size. (A) Smith's pl. 1, fig. 10, Haminoea zelandiae (Gray, 1843), “type figured”. (B) Smith's pl. 1, fig. 11, Bulla quoyii Gray, 1843, “type figured”. (C) Smith's pl. 1, fig. 8, Eudoxochiton nobilis (Gray, 1843), presumed syntype. (D,E) Smith's pl. 1, fig. 6, Polydonta tuberculata Gray, 1843 [= Coelotrochus viridis (Gmelin, 1791)], “type figured”, that is, syntype of P. tuberculata. (F,G) Smith's pl. 1, fig. 14, Diloma subrostrata (Gray, 1843), presumed syntype. (H,I) Smith's pl. 1, fig. 12, Cantharidus sanguineus (Gray, 1843), “type figured” [see Williams et al. (2010) for adoption of Coelotrochus and Cantharidus]. (J,K) Smith's pl. 1, fig. 15, Chlorostoma undulosum A. Adams, 1853 [= Diloma subrostrata (Gray, 1843)], presumed syntype of C. undulosum. (L) Smith's pl. 1, fig. 9, Rhyssoplax aerea (Reeve, 1847), “type in museum collection”. (M) Smith's pl. 1, fig. 21, supposedly Trichotropis clathrata Smith, 1874, “young example,” not a type [apparently Xymene plebeius (Hutton, 1873)]. (N) Smith's pl. 1, fig. 13, Chiton (Plaxiphora) terminalis Smith, 1874, syntype [= Plaxiphora (Maorichiton) caelatus (Reeve, 1847)]. (O,Q) Smith's pl. 1, fig. 16, “Haliotis gibba Philippi?” [actually Haliotis virginea Gmelin, 1791], not a type. (P) Smith's pl. 1, fig. 20, Cerithidea bicarinata Gray, 1843 [= Zeacumantus lutulentus (Kiener, 1841)], “type figured”. (R) Smith's pl. 1, fig. 17, Chiton (Sypharochiton) sinclairi (Gray, 1843), “the type is figured”. (S) Smith's pl. 1, fig. 26, labelled “fig. 24”, supposedly Patella denticulata Martyn, 1784 [actually Cellana ornata (Dillwyn, 1817)], not a type. (T) Smith's pl. 1, fig. 23, Vermetus cariniferus Gray, 1843 [that is, Spirobranchus cariniferus, Polychaeta], “type figured”, despite Smith (1874:1) stating type not in British Museum. (U) Smith's pl. 2, fig. 13, Neilo australis (Quoy & Gaimard, 1835), not a type. (V) Smith's pl. 1, fig. 24, Cellana strigilis redimiculum (Reeve, 1854), presumed syntype. (W) Smith's pl. 2, fig. 3, Irus (Notirus) reflexus (Gray, 1843), “type figured”. (X) Smith's pl. 1, fig. 25, Patella antipodum Smith, 1874 [= Cellana radians (Gmelin, 1791)], a syntype of P. antipodum, established in this work. (Y) Smith's pl. 2, fig. 14, Leionucula strangei (A. Adams, 1856), syntype BMNH 20040712 (Beu 2006:fig. 3E, F). (Z) Smith's pl. 2, fig. 12, Zelithophaga truncata (Gray, 1843), “type figured”.
![Fig. 1 Reproductions of illustrations of New Zealand marine molluscan type specimens, from Smith (1874), at published size. (A) Smith's pl. 1, fig. 10, Haminoea zelandiae (Gray, 1843), “type figured”. (B) Smith's pl. 1, fig. 11, Bulla quoyii Gray, 1843, “type figured”. (C) Smith's pl. 1, fig. 8, Eudoxochiton nobilis (Gray, 1843), presumed syntype. (D,E) Smith's pl. 1, fig. 6, Polydonta tuberculata Gray, 1843 [= Coelotrochus viridis (Gmelin, 1791)], “type figured”, that is, syntype of P. tuberculata. (F,G) Smith's pl. 1, fig. 14, Diloma subrostrata (Gray, 1843), presumed syntype. (H,I) Smith's pl. 1, fig. 12, Cantharidus sanguineus (Gray, 1843), “type figured” [see Williams et al. (2010) for adoption of Coelotrochus and Cantharidus]. (J,K) Smith's pl. 1, fig. 15, Chlorostoma undulosum A. Adams, 1853 [= Diloma subrostrata (Gray, 1843)], presumed syntype of C. undulosum. (L) Smith's pl. 1, fig. 9, Rhyssoplax aerea (Reeve, 1847), “type in museum collection”. (M) Smith's pl. 1, fig. 21, supposedly Trichotropis clathrata Smith, 1874, “young example,” not a type [apparently Xymene plebeius (Hutton, 1873)]. (N) Smith's pl. 1, fig. 13, Chiton (Plaxiphora) terminalis Smith, 1874, syntype [= Plaxiphora (Maorichiton) caelatus (Reeve, 1847)]. (O,Q) Smith's pl. 1, fig. 16, “Haliotis gibba Philippi?” [actually Haliotis virginea Gmelin, 1791], not a type. (P) Smith's pl. 1, fig. 20, Cerithidea bicarinata Gray, 1843 [= Zeacumantus lutulentus (Kiener, 1841)], “type figured”. (R) Smith's pl. 1, fig. 17, Chiton (Sypharochiton) sinclairi (Gray, 1843), “the type is figured”. (S) Smith's pl. 1, fig. 26, labelled “fig. 24”, supposedly Patella denticulata Martyn, 1784 [actually Cellana ornata (Dillwyn, 1817)], not a type. (T) Smith's pl. 1, fig. 23, Vermetus cariniferus Gray, 1843 [that is, Spirobranchus cariniferus, Polychaeta], “type figured”, despite Smith (1874:1) stating type not in British Museum. (U) Smith's pl. 2, fig. 13, Neilo australis (Quoy & Gaimard, 1835), not a type. (V) Smith's pl. 1, fig. 24, Cellana strigilis redimiculum (Reeve, 1854), presumed syntype. (W) Smith's pl. 2, fig. 3, Irus (Notirus) reflexus (Gray, 1843), “type figured”. (X) Smith's pl. 1, fig. 25, Patella antipodum Smith, 1874 [= Cellana radians (Gmelin, 1791)], a syntype of P. antipodum, established in this work. (Y) Smith's pl. 2, fig. 14, Leionucula strangei (A. Adams, 1856), syntype BMNH 20040712 (Beu 2006:fig. 3E, F). (Z) Smith's pl. 2, fig. 12, Zelithophaga truncata (Gray, 1843), “type figured”.](/cms/asset/921151c7-5acb-40a3-81a6-571fca9773ef/tnzr_a_500717_o_f0001g.gif)
Fig. 2 Reproductions of illustrations of New Zealand marine molluscan type specimens, from Smith (1874), bat published size. (A) Smith's pl. 2, fig. 2, Cyclomactra ovata (Gray, 1843), “the type is figured” (hinge inaccurate). (B) Smith's pl. 3, fig. 7, Talochlamys zelandiae (Gray, 1843), “type figured”. (C) Smith's pl. 3, fig. 8, Felaniella (Zemysia) zelandica (Gray, 1835), “type figured”. (D) Smith's pl. 2, fig. 10, Crassula aequilatera (Deshayes in Reeve, 1854) (see Petit 2007:81), “type in Brit. Mus”. (E) Smith's pl. 2, fig. 6, Irus (Notopaphia) elegans (Deshayes, 1854), “type in Brit. Mus.”. (F) Smith's pl. 2, fig. 7, Tellinota edgari (Iredale, 1915), “type in Brit. Mus.”, that is, syntype of Tellina glabrella Deshayes, 1854 (junior primary homonym of T. glabrella Delle Chiaje, 1830), renamed T. edgari. (G,H,I) Smith's pl. 2, fig. 8, Pholadidea tridens (Gray, 1843) and interior and exterior of siphonoplax, “type figured”. (J) Smith's pl. 2, fig. 11, Gari (Psammobia) lineolata (Gray, 1843), presumed syntype. (K) Smith's pl. 3, fig. 6, Paphies ventricosa (Gray, 1843), remaining syntype, BMNH 1842.5.17.189.

Fig. 3 Reproductions of illustrations of New Zealand marine molluscan type specimens, from Smith (1874), at published size. (A) Smith's pl. 3, fig. 3, Irus (Notirus) reflexus (Gray, 1843), not a type, “Fig. 3 on pl. 3 appears to represent a species intermediate between reflexa and V. siliqua Desh.”. (B) Smith's pl. 2, fig. 5, Vanganella taylori Gray, April 1853 [= Resania lanceolata Gray, Jan. 1853], not a type. (C) Smith's pl. 2, fig.1, Dosina zelandica Gray, 1835, presumed syntype of Venus oblonga Gray, 1843. (D) Smith's pl. 2, fig. 4, supposedly Mactra discors Gray, 1843, not a type [“the … figure is taken from a very young specimen” of M. murchisoni Deshayes in Reeve, 1854]. (E) Smith's pl. 3, fig. 5, Dosina zelandica Gray, 1835, presumed syntype of Dosina zelandica, “the type… is in the British Museum”. (F) Smith's pl. 3, fig. 1, Solemya (Zesolemya) parkinsoni E. A. Smith, 1874, syntype; established in this work.
![Fig. 3 Reproductions of illustrations of New Zealand marine molluscan type specimens, from Smith (1874), at published size. (A) Smith's pl. 3, fig. 3, Irus (Notirus) reflexus (Gray, 1843), not a type, “Fig. 3 on pl. 3 appears to represent a species intermediate between reflexa and V. siliqua Desh.”. (B) Smith's pl. 2, fig. 5, Vanganella taylori Gray, April 1853 [= Resania lanceolata Gray, Jan. 1853], not a type. (C) Smith's pl. 2, fig.1, Dosina zelandica Gray, 1835, presumed syntype of Venus oblonga Gray, 1843. (D) Smith's pl. 2, fig. 4, supposedly Mactra discors Gray, 1843, not a type [“the … figure is taken from a very young specimen” of M. murchisoni Deshayes in Reeve, 1854]. (E) Smith's pl. 3, fig. 5, Dosina zelandica Gray, 1835, presumed syntype of Dosina zelandica, “the type… is in the British Museum”. (F) Smith's pl. 3, fig. 1, Solemya (Zesolemya) parkinsoni E. A. Smith, 1874, syntype; established in this work.](/cms/asset/29edaba1-bd10-4faf-aa70-2a67d0c50543/tnzr_a_500717_o_f0003g.gif)
Abbreviations and conventions
Collections and institutions housing specimens listed here are abbreviated:
Dimensions of specimens are cited in millimetres, in the consistent order H (= height), D (=diameter).
The standard abbreviations for points of the compass (N = north, S = south, E = east, W = west, NE, SSW, etc.) are used to abbreviate locality details.
Unless otherwise stated, I have examined the type material cited below under ‘Type material’.
Locality details
Detailed locality records are listed below for the less common species, for which localities help to establish accurate time range. Full locality data are not listed for common, widespread species.
Fig. 4 Reproductions of illustrations of New Zealand marine molluscan type specimens, from Smith (1874), at published size. (A) Smith's pl. 3, fig. 4, Austrovenus stutchburyi (Wood, 1828), presumed syntype. (B,F) Smith's pl. 3, fig. 9, 10, Pododesmus (Monia) zelandica (Gray, 1843), “type figured” (syntype, Fig. 4F, Smith's fig. 10; and interior of byssal foramen enlarged). (C) Smith's pl. 3, fig. 2, Dosinia (Phacosoma) subrosea (Gray, 1835), “the type… is here figured”. (D) Smith's pl. 2, fig. 9, Soletellina nitida (Gray, 1843), presumed syntype. (E) Smith's pl. 3, fig. 11, Circomphalus yatei (Gray, 1835), “The type … is figured”.

New records from Mikonui-1 well, off Westland
In an obscure report (Beu in Morgans Citation1981:12–13, table 1), I recorded Mollusca of bathyal aspect and late Nukumaruan–early Castlecliffian age from the Mikonui-1 well, a wild-cat oil well drilled off the west coast of the South Island, west of Hokitika, by the ‘Sedco 45’ drill ship, at 42°41′15″S, 170°08′45″E. The diverse molluscs were all from well cuttings from the upper part of the core. Depths originally were reported well depths, including 170 m for water depth plus height of the drilling platform above the water; 170 m has therefore been deducted from the depths to arrive at those reported here. Also, the uppermost 170 m drilled in the well was not recovered. Seven samples from near the top of the well (fossil record no. SE42170/f01; extending from 170 to 520 m below the seabed; at 170–180 m, 240–250 m, 270–280 m, 300–310 m, 360–370 m, 450–460 m, and 510–520 m) contained diverse, well-preserved molluscs, including such taxa as Paracomitas gemmea (Murdoch, Citation1900) (see in Part 4) and Propebela cf. abnormis (Hutton, 1885) (Propebela is adopted in Part 4), indicating a probable latest Nukumaruan–early Castlecliffian age. Fragments of other taxa such as Falsilunatia, Splendrillia and Scaphander contribute to the interpretation of this fauna as having lived in the upper bathyal zone, in water depths of 200–400 m, or more. The few fragments present of shelf taxa such as Austrofusus glans indicate down-slope transport of some specimens into the deposition site.
Several of the species present in these samples had not previously been recorded as fossils*, but were described in the Recent fauna of New Zealand, and the new records and a few other uncommon species are recorded here:
* Saccella hedleyi (Fleming, 1951), fragments (240–250 m);
*Austrotindaria benthicola Dell, 1956 (240–250, 300–310 m);
Poroleda lanceolata (Hutton, 1885) (240–250 m);
*Neilo cf. wairoana delli Marshall, 1978, fragments (all samples);
Neilo cf. sublaevis Marwick, Citation1926, fragments (240–250 m);
*Lissotesta cf. errata Finlay, 1926 (240–250 m);
*Tectisumen clypidellaeformis (Suter, 1908) (450–460 m);
*Rissoa (Haurakia) subsuturalis (Dell, 1956) (almost all samples);
*Rissoa (Haurakia) otagoensis (Dell, 1956) (510–520 m);
*Malluvium calcareum (Suter) (Suter, 1909) (240–250 m);
Galeodea triganceae Dell, 1953, fragments (240–250, 270–280 m);
Sassia kampyla (Watson, 1885), fragments (170–180, 240–250, 270–280 m);
*Cominella (Eucominia) powelli (Fleming, 1948) (240–250, 270–280, 360–370 m);
Penion benthicolus Dell, 1956 (240–250, 270–280, 300–310 m);
Paracomitas gemmea (Murdoch, Citation1900) (240–250 m) (see below);
Taranis nexilis (Hutton, 1885) (170–180, 240–250, 270–280 m);
Aoteadrillia wanganuiensis (Hutton, 1873) (170–180, 240–250 m);
Antimelatona buchanani (Hutton, 1873) (270–280 m);
Propebela cf. abnormis (Hutton, 1885) (240–250 m);
*Ringicula aff. delecta Murdoch & Suter, 1906 (240–250 m).
Taxonomy
Phylum Mollusca
Class Gastropoda
Remarks
The taxonomic order here follows that of Bouchet et al. (Citation2005). Authorship and dates of all taxa at subfamily level and above also can be found in Bouchet et al. (Citation2005).
Clade Vetigastropoda
Family Trochidae
Genus Cantharidella Pilsbry, 1889
Cantharidella Pilsbry 1889a:197. Type species (by original designation): Gibbula picturata A. Adams & Angas, 1864, Recent, SE Australia.
Remarks
Molecular studies of Trochoidea and Turbinoidea are in the process of revitalizing their classification at present, and Williams & Ozawa (Citation2006), Williams (Citation2007) and Williams et al. (Citation2008) have recognized several new families. Cantharidella has usually been placed in a subfamily Gibbulinae, but Williams et al. (Citation2008:15) pointed out that the blue-green nacre that characterizes Trochidae Cantharidinae, as compared with the more usual colourless to pearly nacre of other trochoideans, is also present in Gibbula and Cantharidella. In awider molecular study, Williams et al. (2010) showed that Gibbula and Cantharidella belong in the Cantharidinae.
Cantharidella tessellata (A. Adams, Citation1853) (A,D)
Fig. 5 (A,D) Cantharidella tessellata (A. Adams), GS12790, V20/f038, Tangoio Limestone (late Nukumaruan), Petane Pa, Tangoio, Hawke's Bay; SEM, 2 views of abraded specimen, width 6.3 mm. (B,G,I) Calliostoma (Maurea) nukumaruense (Laws), 2 specimens, GS4115, R22/f6435, Nukumaru Brown Sand (late Nukumaruan), Nukumaru Beach, Wanganui; B, G, H 16.1, D 17.7 mm; I, height 20.3 mm. (C,F,H) Clanculus plebejus (Philippi), GS2784, T23/f6491, 3-10 m above Torlesse greywacke, Opawe Stream, Pohangina Valley, early Nukumaruan; 3 views of 1 shell, width 7.1 mm. (E) Risellopsis varia (Hutton), GS15119, R28/f062, 1855 uplifted beach, Cape Turakirae, Wellington; SEM; width 4.7 mm (see also B).

Margarita tessellata A Adams Citation1853:191.
Gibbula nitida A Adams & Angas Citation1864:36; Hutton Citation1880:102.
Chrysostoma inconspicua (sic). Hutton Citation1873a:36.
Gibbula inconspicua. Hutton 1880:102.
Photinula nitida. Suter Citation1913:134, pl. 38, ; Finlay Citation1926b:358.
Talopena sublaevis Finlay Citation1924c:520, ; Cernohorsky Citation1972:241, .
Cantharidella tessellata. Finlay Citation1926b:358; Powell Citation1979:57, pl. 18, Fig. 25; Spencer & Willan Citation1996:14; Spencer et al. Citation2009:203; Maxwell Citation2009:239.
Type material
Margarita tessellata, three syntypes BMNH 1968107, without locality. Gibbula nitida, 18 syntypes BMNH 1870.10.26.144, from ‘Hokianga [Harbour, Northland], New Zealand’. All these specimens are conspecific, and are the New Zealand species known by the name Cantharidella tessellata, and lectotypes are not needed. Chrysostoma inconspicuum, four syntypes NMNZ M.210 (Marshall Citation1996:13), without locality. Talopena sublaevis, holotype, AIM AK70756 (Cernohorsky Citation1972:241, ), a juvenile specimen from ‘oyster scrapings, Bluff’, Foveaux Strait (not seen).
Other material examined
Nukumaruan: GS12816, V20/f0147, Darkys Spur Formation, Waipunga Road, central Hawke's Bay (late Nukumaruan, OIS 67–69?; 1); GS12790, V20/f038, Tangoio Limestone, cutting on track 500 m N of Petane Pa, N of Napier, central Hawke's Bay (late Nukumaruan, OIS 65?; one; A,D). Haweran: GS12783, R22/f0151, Putiki Shellbed, main highway in Awarua valley, E of Wanganui (early Haweran, OIS 9; 1); also occurs in Brunswick Formation at Mt Jowett (GS4151, R22/f7416, 1) and at Sewell Road, Wanganui (OIS 9); Denby Shellbed at Waihi Beach, Hawera, South Taranaki (OIS 7); Rapanui Terrace, Waitotara, west of Wanganui (OIS 5e); Cape Wanbrow, Oamaru (OIS 5e); and Holocene deposits at Cape Kidnappers and Mahia Peninsula (common in many samples from the transects through Holocene terraces at Table Cape).
Distribution
Cantharidella tessellata occurs now in the intertidal zone of exposed rocky shores throughout the three main islands of New Zealand, but not at the Chatham Islands (Powell Citation1979:57). Fossils are recorded from late Nukumaruan (OIS 67–69?) rocks to Recent, but are rare other than in a few localities in the Petane Group in Hawke's Bay, in Brunswick Formation (OIS 9) around Wanganui, and in Holocene terrace cover beds.
Dimensions
Margarita tessellata, syntypes H 6.7, D 8.3 mm; H 6.0, D 7.6 mm; H 6.6, D 6.8 mm; RM3443, coast E of Island Bay, Wellington: H 8.2, D 8.8 mm; RM915, Waimarama, E Wairarapa: H 8.3, D 6.9 mm (unusually tall and narrow).
Remarks
Cantharidella tessellata is easily recognized by being almost the only one of the common, small, shallow-water New Zealand trochids that is deeply and obviously, if narrowly, umbilicate. Combined with the low, wide, closely spaced spiral cords, the umbilicus is a readily recognized character. Care has to be taken to distinguish C. tessellata from umbilicate specimens of Cantharidus species, particularly C. tenebrosus (A. Adams, 1853), which Marshall (Citation1998:126, –23) showed retains an open umbilicus at all sizes in all but the narrowest individuals. However, C. tenebrosus is easily distinguished from C. tessellata by its larger size, taller spire, and much more prominent, narrow, well-separated spiral cords. In C. tessellata, the height is roughly equal to the diameter (slightly taller than the diameter in some specimens), the shape is almost spherical, and the surface is unusually smooth for a New Zealand trochid, with low, closely spaced spiral cords, lightly polished in many specimens. The syntypes of Margarita tessellata are all patterned in paler and darker grey, in a checkerboard pattern, whereas the syntypes of Gibbula nitida are the more usual multicoloured assortment of checkerboard and spiral patterns.
This is one of several species that Fleming (Citation1953) thought appeared first in Brunswick Terrace cover beds (OIS 9) in Wanganui Basin, at Mt Jowett, Wanganui, and therefore was regarded until recently as a potential Haweran index species. Along with several other such species, it is now clear that there are much earlier records elsewhere (including Risellopsis varia (Hutton, 1873) [see below], Mytilus galloprovincialis Lamarck, Citation1819 and Protothaca crassicosta (Deshayes, Citation1835); Beu Citation2006). These species occupy intertidal rocky shores and therefore have a fossil record that is highly unreliable for biostratigraphy. The common New Zealand shallow offshore to sandy shoreface fossil localities of Castlecliffian–Nukumaruan age rarely contain such taxa. C. tessellata now can be recorded from late Nukumaruan (OIS 67–69?) rocks of the Petane Group in Matapiro Syncline, central Hawke's Bay. I am not aware of records earlier than late Nukumaruan, but this apparent first occurrence is most unlikely to be reliable for biostratigraphy. Most fossil specimens are severely abraded, reflecting transport from their intertidal environment.
Genus Clanculus Montfort, 1810
Clanculus Montfort 1810:190. Type species (by monotypy): ‘Trochus pharaonicus’ (= Trochus pharaonius) Linné, 1758, Recent, Indo-West Pacific.
Mesoclanculus Iredale 1924:224. Type species (by original designation, Iredale 1924:182): Clanculus plebejus Philippi, Citation1851, Recent, southern Australia.
Remarks
Wilson (Citation1993:83) pointed out that Cotton & Godfrey's (1934) generic subdivision of Clanculus can be applied only to the southern Australian temperate species, and breaks down with the addition of tropical Indo-West Pacific species. Mesoclanculus is therefore regarded as a synonym of Clanculus, which also has many other synonyms. Jansen (1995b) also used no subgenera in her revision of Australian Clanculus species.
Clanculus plebejus (Philippi, Citation1851) (C,F,H)
Trochus plebejus Philippi 1851:41; Philippi 1855 (in 1846–1855), p. 326, pl. 46, ; Fischer 1878 (in Citation1875–Citation1880), p. 243, pl. 83, , 2a.
Clanculus nodiliratus A Adams 1853:163; Angas 1865:179; Angas Citation1871:96; Tenison Woods Citation1878:40; Tenison Woods Citation1880:69.
Clanculus angeli Tenison Woods Citation1877:144 (synonym according to Tenison Woods 1878); Hardy Citation1915:62.
Clanculus dominicanus Tenison Woods 1877:144 (synonym according to Tenison Woods 1878); Tenison Woods 1878:40; Tenison Woods 1880:69; Hardy Citation1915:65.
Gibbula multicarinata Tenison Woods 1877:142 (synonym according to Tenison Woods 1878; Iredale 1924:224); Tenison Woods 1878:42; Tenison Woods 1880:70; Petterd Citation1879:354; Hardy Citation1915:68.
Trochus muscarius Fischer 1878 (in 1875–1880), pp. 243, 244 (in synonymy of ‘Trochus plebejus var. β’; synonym according to Iredale 1924:224).
Trochus nodoliratus. Whitelegge Citation1889:271.
Trochus (Clanculus) plebejus. Pilsbry Citation1889b:79, pl. 10, –22; pl. 13, , 2.
Clanculus plebejus. Tate & May Citation1901:401; May Citation1921:37; May Citation1923:41, pl. 18, ; Macpherson & Chapple Citation1951:113; Macpherson Citation1958:22, pl. 18, ; Monger Citation1984:140, illus.; Wilson Citation1993:88, unnumbered fig.; Jansen Citation1995:59, pl. 3, –18.
Trochus dominicanus. Pilsbry Citation1889b:59.
Trochus angeli. Pilsbry 1889b:60.
Clanculus plebeius (sic). Pritchard & Gatliff Citation1902:122; Verco Citation1908:6; Verco Citation1912:202; Hedley Citation1916a:178; Hedley Citation1916b:700; Verco Citation1935:6.
Trochus (Clanculus) takapunaensis Webster Citation1906:309, pl. 39, , 2a.
Eurytrochus plebejus Hedley Citation1918:M42.
Mesoclanculus plebeius. Iredale 1924:182, 224.
Clanculus (Mesoclanculus) plebejus. Cotton & Godfrey Citation1934:89; Singleton 1937:389; Cotton & Godfrey Citation1938:4; Kershaw Citation1955:304; Macpherson & Gabriel Citation1962:73; Macpherson Citation1966:212; Ludbrook & Gowlett-Holmes Citation1989:558, .16n-p.
Mesoclanculus plebejus. Powell Citation1958b:89; Cotton Citation1959:138, fig. 73.
Clanculus (Mesoclanculus) plebeius. Ludbrook Citation1978:9.
Type material
Trochus plebejus Philippi, type material presumably in Philippi's collection, in Museo Nacional de Historia Natural, Santiago, Chile; not available for examination. Clanculus nodiliratus A. Adams, no type material present in BMNH (K Way, BMNH pers. comm. 14 Jan 2008), location unknown; without locality. Gibbula multicarinata TASM TM5566, one syntype; Clanculus angeli TASM TM5564, one syntype; and C. dominicanus TASM TM5558, one syntype, all of Tenison Woods (1877) (Hardy Citation1915:62, 65, 68) (not seen). Trochus muscarius Fischer, no type material in MNHN; as this name was proposed in the synonymy of Trochus plebejus var. β and has never been used as a valid name, type material presumably was never selected. Trochus (Clanculus) takapunaensis Webster, holotype AIM AK70778 (not seen), an empty shell picked up on Takapuna Beach, Auckland; Powell (Citation1958b:89) when synonymizing this name with Mesoclanculus plebejus suggested that it was an ‘accidentally dropped shell’.
Other material examined
Nukumaruan: 3–10 m above Torlesse greywacke, Opawe Stream, Pohangina valley, E Wanganui Basin, with Glycymeris shrimptoni Marwick, 1923, Myadora waitotarana Powell, Citation1931, Sabia australis (Lamarck, 1819) (see below), Diloma, Micrelenchis, Austrolittorina, and diverse Rissooidea, an unusual shallow-water assemblage that obviously was deposited near an exposed rocky shore (GS2784, T23/f6491, one specimen). Recent: 35 m, off Taranga I, Hauraki Gulf, Auckland, C. A. Fleming collection (RM1001, two small, abraded, the larger incomplete).
Distribution
Clanculus plebejus is an abundant shallow-water trochid of exposed rocky shores in SE Australia, from Sydney, New South Wales, to Cape Naturaliste, S Western Australia (Wilson Citation1993:88). Macpherson & Gabriel (Citation1962:73) stated that C. plebejus is ‘the commonest species of the genus in Victoria obtained in countless numbers on the whole coastline’. A single shell from Takapuna Beach, Auckland, the holotype of Trochus takapunaensis Webster, was considered by Powell (Citation1958b) to belong in C. plebejus, although it is much more finely sculptured than the specimen illustrated by Wilson (Citation1993:88). Charles Fleming's boyhood collection also contains two small, abraded specimens of ‘Clanculus takapunaensis’ from Hauraki Gulf, Auckland. These are typical specimens of C. plebejus, with six uniform, rather coarsely nodulose spiral cords on the upper surface of the last whorl. Another New Zealand specimen is recorded here, from early Nukumaruan rocks in Opawe Stream, Pohangina valley, eastern Wanganui Basin, but all new Zealand records presumably result from accidental rafting events (or, for the Recent specimens, might be merely mislocalized Australian shells). Specimens evidently were rafted to New Zealand at least once, and possibly three times.
Dimensions
GS2784, Opawe Stream: H 5.1, D 7.1 mm; normal adults 7×10 mm (Cotton Citation1959:139).
Remarks
The newly recorded Opawe Stream specimen is highly aberrant and does not agree exactly with any southern Australian Clanculus species. Firstly, it has an immature aperture, with thin lips, so the final apertural armament is unknown. The umbilical wall also has been perforated by an epibiont, probably a bryozoan formerly attached there. However, it is clear that it is a species of one of the Clanculus species groups with a relatively simply sculptured umbilicus and inner lip, and an essentially smooth lining of the umbilicus. It differs from the restricted northeastern New Zealand Recent species C. peccatus (Finlay, 1926) (Powell Citation1979:60, pl. 17, ) in its much lower spire angle, its strongly convex rather than flat whorl profile, its obviously channelled rather than weakly impressed suture, its fewer, more prominent spiral cords, its more finely sculptured base, and its much less elaborately sculptured aperture and umbilicus. The Opawe Stream specimen has a unique wide, shallow depression occupying the inner half of the base over the last eighth of a whorl, between the umbilicus and a medial angulation along the outer edge of the depression, and it is clear that whatever species it belongs in, it is an aberrant specimen and cannot be expected to agree in all characters with normal specimens. The spiral sculpture on the upper (adapical) surface of early spire whorls consists of three equally prominent, quite coarsely nodulose cords, but on the penultimate whorl the uppermost cord begins to subdivide, producing on the last half-whorl a wide, shallow subsutural channel, followed by a depressed zone with six equally low, closely spaced, finely beaded spiral cords, followed below by two very prominent, widely spaced, coarsely nodulose cords continuing unchanged from the earliest teleoconch whorl; the lowest of these is weakly subdivided by a groove, and the lowest, almost smooth subdivision of this cord forms the periphery. The base bears seven smooth, narrow spiral cords in addition to the peripheral one, with a fine secondary thread between the second and third ones in from the margin. A smooth, low tooth at the base of the columellar margin of the umbilicus is set well back from the aperture; the inner lip is otherwise smooth. The entire surface is crossed by narrow, well-raised axial lamellae, coarse and prominent on the upper surface but thin and low on the base. The general appearance is therefore intermediate between those of Webster's (1906, pl. 39, ) and Wilson's (1993:88) drawings of C. plebejus, with a very similar base apart from the lack of nodules around the outer edge of the umbilicus, but with a protruding lower half of the upper whorl surface, a depressed upper half, and a wide, shallow sutural channel. This very aberrant specimen seems nearest to C. plebejus, and perhaps was aberrant as a result of surviving very rare larval transport to New Zealand and finding an unsuitable habitat here. It seems more likely, however, that this specimen was rafted to New Zealand in an algal holdfast or similar floating object. It is interesting that Webster's (1906) specimen was also considered by Powell (Citation1958b:90) to fall within the variation of southern Australian specimens of C. plebejus, suggesting the possibility that Webster's specimen was also a rafted shell. As noted above, the two Fleming collection specimens from off Taranga Island, Auckland, are also very small, abraded specimens of C. plebejus, so this species that is so abundant in SE Australia evidently is the trochid most commonly rafted from Australia to New Zealand. Jansen (Citation1995:48) recorded that the more westerly species C. consobrinus Tate, Citation1893 (Adelaide to Kalbarri, Western Australia) is similar to C. plebejus but has fewer spiral cords, is paler in colour, does not have nodules on all spiral cords, and lacks a columellar tooth, so in some ways the fossil seems closer to C. consobrinus than C. plebejus. However, the aberrant fossil is difficult to compare, and the further westerly distribution makes C. consobrinus an unlikely migrant to New Zealand, so an identification as C. plebejus seems more reasonable.
The sole previous fossil record of a southern Australian trochoidean from New Zealand is the specimen of Bankivia fasciata recorded by Beu (Citation2004:187). That also was considered likely to have been rafted. B. fasciata is also an abundant species in southern Australia. The minute demersal larvae of trochoideans float well and have been identified as a possible means of dispersal by Hadfield & Strathmann (Citation1990) and Hickman (Citation1992). It certainly appears that some trochoidean species have wide distributions in the Indo-West Pacific similar to those of planktotrophic caenogastropods such as Cypraea and Monoplex species. However, molecular phylogeny has demonstrated that some apparently widespread Indo-West Pacific turbinoideans (recognized as a superfamily separate from Trochoidea by Williams & Ozawa Citation2006) such as Astralium rhodostoma (Lamarck, 1822) (Meyer et al. Citation2005) are actually diverse species complexes, with restricted species inhabiting each island group, making it obvious that genetic exchange of turbinoideans (and so, by analogy, trochoideans) between island groups has not been as great as shell morphology suggests. This and the fact that not one species of trochoidean is now in common to Australia and New Zealand (BA Marshall NMNZ pers. comm.) makes a rafting origin for southern Australian trochoideans in Wanganui Basin considerably more likely than transport as larvae. The occurrence of another rocky shore intertidal trochoidean, Diloma nigerrima (Gmelin, 1791) abundantly in both Chile and New Zealand as well as at Juan Fernandez (Donald et al. Citation2005) demonstrates another case of rafting of a trochoidean, so their transport on algal holdfasts is possibly more common than has been appreciated previously. Waters (Citation2008) recently argued for a significant role of algal rafting in southern temperate marine biogeography.
Family Calliostomatidae
Genus Calliostoma Swainson, Citation1840
Calliostoma Swainson 1840:218, 351. Type species (by subsequent designation, Herrmannsen 1846:154): Trochus conulus Linné, 1758, Recent, Mediterranean and northeastern Atlantic.
Remarks
A synonymy of the genus and discussion of its status and synonyms were provided by Marshall (Citation1995a). Calliostomatidae was established firmly as a family of Trochoidea by Williams et al. (Citation2010).
Subgenus Maurea Oliver, Citation1926
Maurea Oliver Citation1926:108. Type species (by original designation): Trochus tigris Gmelin, 1791, Pleistocene and Recent, New Zealand [20 December 1926].
Mauriella Oliver Citation1926:109. Type species (by original designation): Trochus punctulatus Martyn, 1784, Pleistocene and Recent, New Zealand [20 December 1926; species name available, ICZN Opinion 479 1957].
Calliotropis Oliver Citation1926:110. Type species (by original designation): Trochus cunninghami Gray, 1834 (= Trochus selectus Dillwyn, Citation1817), Pleistocene and Recent, New Zealand [20 December 1926] (junior homonym of Calliotropis Seguenza, 1903).
Mucrinops Finlay Citation1926b:360. Type species (by original designation): Ziziphinus spectabilis A. Adams, Citation1855, Recent, southern New Zealand [23 December 1926].
Venustas Finlay Citation1926b:360. Type species (by original designation): Trochus tigris Gmelin, 1791, Pleistocene and Recent, New Zealand [23 December 1926; name officially rejected, ICZN Opinion 479 (1957:373); junior homonym of Venustas Allan, 7 December 1926].
Calotropis Thiele Citation1929:49. Replacement name for Calliotropis Oliver, Citation1926, preoccupied.
Calliostoma (Maurea) nukumaruense (Laws, 1930) (B,G,I)
Modelia nukumaruensis Laws 1930:550, pl. 90, ; Fleming Citation1966:40.
Calliostoma (sensu lato) nukumaruense. Beu & Maxwell Citation1990:405.
Calliostoma (Maurea) nukumaruense. Maxwell Citation2009:239.
Type material
Modelia nukumaruensis, holotype AIM AK70483 (not seen), from ‘Nukumaru’, that is, Nukumaru Brown Sand, Nukumaru Beach, late Nukumaruan (Fleming Citation1953:143); ‘two juvenile paratypes (Kai Iwi)’ (early Castlecliffian, horizon unknown).
Other material examined
Fleming (Citation1953:139, 143, 174, 178, 181, 182) recorded ‘Modelia nukumaruensis’ from Nukumaru Limestone (GS4183, R22/f6468, Ototoka Stream), Nukumaru Brown Sand (GS4115, R22/f6435, ‘member f’, Nukumaru Beach), four localities in Butlers Shell Conglomerate, OIS 31 (GS4081, R22/f6412; GS4083, R22/f6413; GS4109, R22/f6432, on the Ototoka coast, and GS4157, R22/f6458, in Kai-Iwi Stream), from the basal conglomerate member of Lower Okehu Siltstone, OIS 29 (Mowhanau Formation; Abbott & Carter Citation1999) on the Ototoka coast (GS4082, R22/f6532; GS4108, R22/f6431, as ‘Maurea (Mucrinops) nukumaruensis’), Okehu Shell Grit, OIS 27 (GS4066, R22/f5387; GS4071, R22/f6402), and the basal conglomerate member of Upper Okehu Siltstone, OIS 27 (Ostrea-Dosina Bed; Abbott & Carter Citation1999) (again as ‘Maurea (Mucrinops) nukumaruensis’; from any of five collections, not listed individually). Specimens recorded by Fleming (Citation1953:209) from undifferentiated formations of Kai-Iwi Group (GS4235, R21/f8479, W side Whangaehu Valley; GS4375, S22/f6498, head of Kaukatea valley) are not assignable to a precise formation or age. Material in the GNS reference collection: ‘Kai-Iwi’, unlocalized to formation, CA Fleming collection (four; abraded, probably from Kaimatira Pumice Sand, OIS 25); Nukumaru Brown Sand, Nukumaru Beach (GS4115, R22/f6435, 18; GS4117, R22/f6437, four; GS4184, R22/f7433, one); GS4066, R22/f6397, Okehu Shell Grit, Okehu Beach (OIS 27; four); Kaimatira Pumice Sand, OIS 25 (GS4139, R22/f6449, one; GS4140, R22/f6450, one; GS4148, R22/f7414, three; GS4064, R22/f6395, nine; GS4163, R22/f6459, one; GS15190, S22/f0155, one abraded); Kaikokopu Shellbed, OIS 19 (GS4134, R22/f7408, four; GS15191, S22/f0156, one fragment); Kupe Formation (OIS 17), Castlecliff coast (GS4047, R22/f6378, two; GS4121, R22/f6521, one, with one C. punctulatum; GS14150, R22/f0128, one).
Distribution
Calliostoma nukumaruense is recorded only from late Nukumaruan and early Castlecliffian rocks of Wanganui Basin; the youngest record is from Kupe Formation (OIS 17).
Dimensions
Modelia nukumaruensis, holotype: H 18, D 15 mm (Laws 1930:551); GS4115, Nukumaru Brown Sand, Nukumaru Beach: H 20.3, D 19.3 mm; H 16.4, D 17.0 mm; H 16.9, D 16.9 mm; H incomplete, D 26.9 mm (largest seen, apex missing, identity slightly in doubt); GS4121, Kupe Formation, Castlecliff: H 16.2, D 17.8 mm.
Remarks
Laws (1930) was impressed by the similarity of this species in its turbiniform shape, evenly and strongly convex whorl profile, and relatively coarse, even, nodulose sculpture to Modelia granosa (Martyn, 1784) (Turbinidae), and described it under the name Modelia nukumaruensis. M. granosa was described from the Recent New Zealand fauna but is recorded intermittently from rocks of Tongaporutuan (late Miocene) to Pleistocene age. Laws compared the new species only with this member of the Turbinidae. CA Fleming published a position in Calliostoma in several places in species lists (Fleming Citation1953), and discussed it with PA Maxwell and me many years ago. Initially this position was based on the observation that no calcareous opercula characteristic of the Turbinidae have been collected anywhere in the formations in which this species occurs, although the shells are common at some horizons within Nukumaru Brown Sand. The teleoconch is much smaller (to ca 27 mm high, although rarely more than 20 mm, compared with to ca 80 mm high in M. granosa) and a little more finely sculptured than in M. granosa. It also has a general appearance much more nearly like those of New Zealand species of Calliostoma (Maurea) with evenly and strongly inflated whorls, a deeply impressed suture, and evenly granulose sculpture, such as the Pliocene–Recent species C. granti (Powell, Citation1931) and the Pleistocene–Recent species C. punctulatum (Martyn, 1784) (see Marshall Citation1995a). There is no doubt that C. nukumaruense is closely related to these Calliostoma species. It is recognizable by its consistently small size (few specimens >20 mm high), its thick shell, its strongly and evenly inflated whorls, and its consistently coarse sculpture.
Compared with the development of teleoconch spiral cords on Calliostoma (Maurea) granti and C. punctulatum, described by Marshall (Citation1995a:89,92, Fig. 28, 29), C. nukumaruense is distinguished by lacking the strongly convex fifth teleoconch whorl of C. punctulatum. All secondary cords appear almost simultaneously (S3 slightly before S1 and S2 on most specimens) on the fourth teleoconch whorl and all enlarge rapidly to be as prominent as the primary cords over the fifth whorl of C. nukumaruense. Secondary cords appear on the fifth teleoconch whorl on C. punctulatum, leaving an unusually obvious channelled suture on early spire whorls that disappears once the outline is filled in by S3 on the fifth whorl. In contrast, all secondary cords appear earlier and S3 appears particularly early on the fourth whorl, so the suture is filled and a distinctly wider fifth whorl is not developed on C. granti. Obvious, narrow axial ridges uniting nodules on neighbouring spiral cords of the first three teleoconch whorls are present on both C. granti and C. nukumaruense, but not C. punctulatum. Some specimens of C. nukumaruense have a weakly developed sutural channel on early spire whorls, less obvious (as it extends a shorter way down the teleoconch) than in C. punctulatum, but more marked than in C. granti. C. nukumaruense is therefore more nearly similar to C. granti than C. punctulatum, but differs from C. granti in the later development of S3, all three main secondary cords developing almost simultaneously over the fourth whorl, and the consequent development of a narrow sutural channel on teleoconch whorls 2–3, as well as in its small size, thick shell, and consistently coarse sculpture. Details of the protoconch and first teleoconch whorl are not visible on any material I have seen.
Clade Sorbeoconcha
Superfamily Cerithioidae
Family Cerithiidae
Genus Taxonia Finlay, 1926
Taxonia Finlay Citation1926b:384. Type species (by monotypy): Ataxocerithium suteri Marwick, 1924, Mangapanian-Nukumaruan (late Pliocene–early Pleistocene), New Zealand.
Taxonia suteri. (Marwick, 1924) (A,E,H)
Fig. 6 (A,E,H) Taxonia suteri (Marwick), late Nukumaruan, Hawke's Bay, SEM; A, sectioned paratype, GS1063, V21/f8476, roadside 1.6 km NW of Okawa Stream, Matapiro syncline; height 16.0 mm; E,H, GS14833, T24/f062, 2 specimens, table-top hill on Waitio Station, Makuri-Coonoor Road; E, height 10.4 mm; H, height 15.2 mm. (B) Risellopsis varia (Hutton), second specimen from same locality as Fig. 5E; width 5.4 mm. (C) Stiracolpus vigilax (Marwick), holotype, TM1686, GS4003, R22/f7394, Landguard Sand (Haweran, OIS 9), Landguard Bluff, Wanganui; height 15.3 mm. (D,F,G,J) Zeacumantus lutulentus (Kiener); D,J, “perplexus form”, GS14833, T24/f062, table-top hill on Waitio Station, Makuri-Coonoor Road, Nukumaruan; height 30.6 mm; F,G, RM3289, uncorroded Recent specimen, head of Pauatahanui arm, Porirua Harbour, W Wellington; height 32.0 mm. (I) Stiracolpus symmetricus (Hutton), TM5375, paralectotype of Turritella (Zaria) tricincta Hutton, 1873 (preoccupied) identified as Stiracolpus delli murdochi by Marwick (1957b), Shakespeare Cliff Siltstone (Castlecliffian, OIS 11), “Shakespeare Cliff”, Wanganui; height 30.7 mm. (K) Stiracolpus huttoni (Cossmann), Marwick's (1957b, pl. 5, fig. 12) illustrated specimen, GS2877, J33/f7076, Lake Kaniere Road, Westland, Opoitian; height 18.3 mm.

Ataxocerithium suteri Marwick Citation1924b:195, pl. 17, ; Finlay Citation1924a:474, pl. 51, a, b.
Taxonia suteri Finlay Citation1926b:384; Fleming Citation1966:48, fig. 929; Beu & Maxwell Citation1990:318, pl. 41i; Maxwell Citation2009:240.
Type material
Ataxocerithium suteri, holotype TM6769, GS1063, V21/f8476, Okawa Stream, central Hawke's Bay, late Nukumaruan. (Care is needed to distinguish Okawa Stream, north of the Ngaruroro River in Matapiro Syncline, from Okauawa Stream, in the Kereru district, south of the Ngaruroro River and west of Mason Ridge).
Other material examined
Mangapanian: GS6215, M34/f7660, 50 m above Waiauan limestone, Weka Creek, 160 m downstream from junction with Weka Pass Stream, N Canterbury (two; with Cominella (Eucominia) aff. elegantula (Finlay, 1926), Polinices waipipiensis (Marwick, 1924), Glycymerita (Manaia) manaiaensis (Marwick, 1923)). Nukumaruan: The few records present in the GNS reference collection are: Okawa Stream, central Hawke's Bay (GS1063, V21/f8476, 18 paratypes); ‘Petane limestone, Petane’, Hawke's Bay (that is, Tangoio Limestone) (GS691, V20/f8476, five); upper band of limestone, Petane, Hawke's Bay (GS720, V20/f8482, six); summit of table-top hill, Waitio Station, west of Makuri-Coonoor Road, southern Hawke's Bay, in estuarine fauna with Austrovenus stutchburyi (Wood, Citation1828) form crassitesta and Zeacumantus lutulentus (Kiener, Citation1842) (GS14833, T24/f062, 22); GS12711, V20/f045, Tangoio Limestone, roadside quarry on Waipunga Road, 1 km N of junction with Turnbull Road, central Hawke's Bay (seven); GS12712, V20/f047, Te Ngaru Mudstone?, Waipunga Road, near top of ascent from Esk valley, thin sand beds with Paphies and mud partings (two); GS12790, V20/f038, Tangoio Limestone, cutting on track 500 m N of Petane Pa, N of Napier, central Hawke's Bay (one).
Distribution
All records of Taxonia suteri are Nukumaruan, apart from the single Mangapanian record from Weka Pass, N Canterbury. Although there are no records from the youngest Nukumaruan rocks of Wanganui Basin (Fleming Citation1953) or, indeed, many from Wanganui Basin at all, it is widespread and abundant in late Nukumaruan rocks of central to southern Hawke's Bay. It could be expected to occur in small numbers in Tewkesbury Formation and Pukekiwi Shellbed in Wanganui Basin. I am not aware of records from Wairarapa district and know of only the one Mangapanian one from North Canterbury, suggesting that T. suteri was a warm-water species.
Dimensions
Ataxocerithium suteri, holotype: H (incomplete) 19.0, D 6.1 mm; paratypes, GS1063, Okawa Creek, Matapiro syncline: H (incomplete) 18.9, D 5.9 mm; H (incomplete) 16.0, D 5.3 mm (sectioned specimen); GS6215, Weka Creek, N Canterbury: H (incomplete) 17.0, D 6.7 mm.
Remarks
Taxonia suteri is highly distinctive and easily recognized. It is a tall-spired, flat-whorled, narrow shell reaching ca 30 mm in height, with the entire exterior sculptured with three rows per whorl of large, closely spaced, smooth, circular nodules, flattened in the plane of the shell surface. As pointed out by Finlay (Citation1926b:384) and Marwick (Citation1948:28) its most distinctive character is the six unusual, prominent, wide, spiral ridges within the aperture: three inside the outer lip (two high up and one near the base; all formed as enlargements of the internally protruding exterior spiral interspaces), two on the parietal area, and one on the base of the columella. One of the paratypes (A) was sectioned longitudinally by Marwick, apparently in 1924, and confirms that the largest spiral ridge, low on the columella, and the spiral ridge on the top of the columella extend down the entire teleoconch.
Taxonia seems to have been an estuarine genus (as are many of the other warm-water ‘mud creepers’ in this superfamily at present) and so there are relatively few records before Nukumaruan time, when this facies became better represented in the fossil record than it had been previously. Marwick (Citation1948:28, pl. 7, ) described T. tesserata from the Waipipian (Pliocene) estuarine fauna of Otahuhu well, Auckland on the basis of a single specimen with smaller, more spherical nodules and a more deeply impressed suture than in T. suteri. The Laws collection material in GNS contains one more specimen. A tubeful of small juvenile cerithioids from Otahuhu well received by GNS in CR Laws’ collection is labelled ‘Taxonia tesserata Marwick, paratypes’ (TM1984–1988, nine specimens) and these all have almost complete apices and one has a complete protoconch. However, the spire angle and sequence of development of spiral cords agree precisely with other specimens assigned to Zeacumantus delicatus Laws, 1950 (that is, Pyrazus ebeninus (Bruguière, Citation1792); Beu Citation2004:200) and there is no doubt that they have been misidentified. Unfortunately, this means that I have not seen a protoconch of Taxonia. Wells (Citation1986:313, a–c) described Taxonia gudrunae from a Tongaporutuan (late Miocene) deposit in northern Wairarapa. T. gudrunae was a member of an estuarine fauna including Austrovenus stutchburyi, Melanopsis trifasciata Gray, 1843, Zeacumantus cf. tirangiensis (Marwick, Citation1926), Zefallacia ruamahunga Wells, Citation1986, and Cominella (Josepha) graemei Wells, Citation1986, which closely resembles the Nukumaruan–Recent estuarine species C. glandiformis (Reeve, 1847). Specimens probably representing Taxonia gudrunae occur also in mudstone lenses within Tongaporutuan conglomerate underlying Hurupi Formation in Hurupi and Putangirua Streams, South Wairarapa, again associated with Zeacumantus cf. tirangiensis. Quite similar specimens have also been collected recently from Chatton Formation (Duntroonian, Oligocene) in a pit on Cosy Dell farm, near Waimumu, Southland (in Geology Department, Otago University). The long gaps in this time range indicate that records of Taxonia are unreliable for biostratigraphy because of its estuarine habitat and concomitant poor fossil record. However, the genus is one of many whose late Nukumaruan extinction provides a biostratigraphically reliable definition of the Nukumaruan–Castlecliffian boundary.
Family Batillariidae
Genus Zeacumantus Finlay, 1926
Zeacumantus Finlay Citation1926b:380. Type species (by original designation): Cerithidea subcarinata G. B. Sowerby II, 1855, Pliocene to Recent, New Zealand.
Zeacumantus lutulentus (Kiener, Citation1842) (P,6D, F, G, J)
Cerithium lutulentum Kiener 1842:63, pl. 22, .
Cerithium bicarinata (sic) Gray 1843:241 (junior primary homonym of Cerithium bicarinatum Lamarck, 1804).
Cerithium (Pirenella) bicarinata. A Adams 1855:84.
Cerithium bicarinatum. GB Sowerby II 1855:888, pl. 186, fig. 288.
Cerithidea bicarinata GB Sowerby II 1866, pl. 4, Fig. 27; EA Smith 1874:3, pl. 1, (repeated here, P); Tryon Citation1887:163, pl. 35, ; Suter Citation1913:238, pl. 39, .
Ataxocerithium perplexum Marshall & Murdoch Citation1919:254, pl. 20, , 6 (new synonym).
Cerithidea perplexa. Marwick Citation1924b:194; Finlay Citation1924a:477.
Zeacumantus perplexus. Finlay Citation1926b:380; Fleming Citation1966:47; Lozouet Citation1986:199; Beu & Maxwell Citation1990:406; Maxwell Citation2009:240.
Zeacumantus lutulentus. Finlay Citation1926b:380; Laws Citation1940b:51; Fleming Citation1966:47; Morton & Miller Citation1968:444–445, pl. 17, ; Powell Citation1979:131, pl. 31, ; Lozouet Citation1986:198–199; Beu & Maxwell Citation1990:406; Spencer & Willan Citation1996:17; Spencer et al. Citation2009:204; Maxwell Citation2009:240.
Type material
Cerithium lutulentum, no type material present in MHNG, presumably lost (Y Finet MHNG pers. comm. 6 November 2007); from ‘les côtes de la Nouvelle Zélande’. Cerithium bicarinatum, three syntypes BMNH 1842.11.16.53–55, from ‘New Zealand’, Dr Stanger (not seen) (one syntype illustrated by EA Smith 1874:3, pl. 1, , repeated here, P); Gray (1843:241) also cited syntypes from the Bay of Islands (presented by Sinclair) but these have not been recognized if they are present in BMNH. K Way (BMNH pers. comm. 14 January 2008) stated that one of the three syntypes has been segregated from the others (perhaps the specimen illustrated by Smith?) but I am not aware of a published lectotype designation. The clear, if only life-size, illustration of a fresh, young, unworn specimen by Smith (1874, pl. 1, ; P) leaves no doubt of the identity of this name. The segregated syntype, BMNH 1842.11.16.53, is here designated the lectotype of Cerithium bicarinatum Gray, 1843 as well as the neotype of Cerithium lutulentum Kiener, 1842. The two types originally were from the same locality, ‘New Zealand’. A neotype designation is required to be certain that the name Cerithium lutulentum unambiguously is attached to the present species. Ataxocerithium perplexum, illustrated syntype TM6762, bearing the original Marshall label: ‘Type example with small area of sculpture’, is from ‘Nukumaru, in blue sandy clay’, that is, Tewkesbury Formation, late Nukumaruan; with one illustrated syntype, TM6763. The second syntype has a completely corroded, smooth surface; it was collected with the more complete syntype. These specimens have been labelled holotype (TM6762) and paratype in the GNS type collection, but in my opinion are syntypes; Marshall & Murdoch (Citation1919:254) did not select one as ‘the type’, or use any other similar term. The specimen with some sculpture remaining, TM6762, is therefore here selected as the lectotype of Ataxocerithium perplexum Marshall & Murdoch, Citation1919, as the other is unidentifiable.
Other material examined
Nukumaruan: almost all undoubted specimens of the form known as Zeacumantus perplexus Marshall & Murdoch, Citation1919 are from Nukumaru Brown Sand and Tewkesbury Formation at Wanganui (e.g. GS15348, R22/f6542A, shellbeds above and below Vinegar Hill Tephra (OIS 61), Brunswick Road, Kai-Iwi valley) and from estuarine sands in Hawke's Bay. Specimens in the GNS reference collection are from Nukumaru Beach (Tewkesbury Formation; GS1164, R22/f6348, 5, one without axial ridges down most of the spire) and summit of table-top hill on Waitio Station, west of the Makuri-Coonoor Road, southern Hawke's Bay (GS14833, T24/f062, many; some typical, some more strongly sculptured; with Taxonia suteri, see above); GS12712, V20/f047, Te Ngaru Mudstone?, Waipunga Road, near top of ascent from Esk valley, thin sandy beds with Paphies and mud partings (14, abraded). However, it is concluded here that Z. perplexus is part of the variation of Z. lutulentus (Kiener, 1842), which is widespread and abundant throughout New Zealand in Mangapanian to Haweran estuarine rocks and in the Recent fauna.
Fossils of Zeacumantus lutulentus in the GNS collection are from Nukumaru Brown Sand, Tewkesbury Formation, Waipuru Shellbed, Butlers Shell Conglomerate, Upper Okehu Siltstone, Kaimatira Pumice Sand, Kaikokopu Shellbed, Kupe Formation, Whangaehu shellbed (OIS 13, underlying Tainui Shellbed on the Whangaehu-Turakina interfluve) and Brunswick Formation at Wanganui, Te Ngaru Mudstone (Nukumaruan, about OIS 65) on Waipunga Road, Hawke's Bay; recorded as Z. perplexus at 345 m and 365 m and as Z. lutulentus at 314 m and 335 m in Kowai Formation in the Bexley testbore, Page's Road, Christchurch (Beu Citation1998; M35/f75 and M35/f69, Nukumaruan); GS14418, V15/f024, Manawahe Road, inland from Matata, Bay of Plenty (OIS 13?), and many younger localities such as Te Piki, near East Cape (OIS 7) and Holocene, Tauranga Memorial Park, Tauranga (GS12569, U14/f03).
Distribution
Zeacumantus perplexus has been thought of as a species restricted to late Nukumaruan rocks of Wanganui Basin, but it is a sculptural variant of Z. lutulentus (Kiener, 1842) rather than a distinct species of biostratigraphical utility. Z. lutulentus is widespread and abundant in estuarine environments throughout the main islands of New Zealand at present, and is a common fossil in Mangapanian to Haweran rocks of this facies.
Dimensions
Ataxocerithium perplexum, holotype: H 33.7, D 10.4 mm; paratype, H 35.0, D 11.3 mm; perplexus form, GS14833, Waitio Station, Hawke's Bay: H 30.6, D 9.3 mm; H 26.3, D 9.8 mm; GS1102, V21/f8492, Maraekakaho Stream, Ngaruroro River, central Hawke's Bay, Nukumaruan: H 35.9, D 12.2 mm; GS4084 re-collection, R22/f6414, Pukekiwi Shell Sand, Ototoka Stream mouth, Wanganui: H 42.0, D 13.3 mm; Recent, RM3289, excavation for bridge abutments, Pauatahanui, W Wellington: H 32.0, D 10.5 mm (F,G); H 31.3, D 10.9 mm; NMNZ M.17341, Parengarenga Harbour: H 32.1, D 11.5; H 34.2, D 10.6; H 30.7, D 11.0; NMNZ M.39886, Ohiwa Harbour, Whakatane: H 32.8, D 10.8; H 31.2, D 10.7.
Remarks
The lectotype of Zeacumantus perplexus is largely corroded, but the last quarter-whorl bears fresh sculpture. It consists of two rather weak peribasal angulations, formed by cords that are slightly more prominent than the others, and fine, close sculpture of low, narrow, closely spaced spiral cords covering the rest of the exterior from the suture to the canal tip. The cords alternate to some extent in prominence and spacing in some areas, particularly in the central part of the upper whorl slope; there are about 18 cords between the suture and the upper peribasal cord, three between the two peribasal cords, and a further 14 on the base. Several narrow, varix-like axial ridges cross the cords at irregular intervals, sinuate in conformity with the outer lip. At face value, this appears to be a very distinctive, restricted late Nukumaruan species, distinguished from the equally large Mangapanian to Recent species Z. lutulentus by its finer sculpture, with slightly more numerous, narrow, closely spaced spiral cords, much weaker axial ridges (fading out down the shell, rather than remaining prominent on to the last whorl as in normal specimens of Z. lutulentus) and slightly weaker peribasal cords. However, examination of GNS reference material showed that specimens agreeing closely with the holotype occur only at Nukumaru and on the table-top hill on Waitio Station, S Hawke's Bay; almost all were collected from Tewkesbury Formation. Similar specimens from other Nukumaruan localities have slightly more prominent spiral cords, and axial ridges continuing onto the last whorl. They indicate that Z. perplexus is based on unusual, weakly sculptured populations of Z. lutulentus that occurred only in the Nukumaru area and in southern Hawke's Bay. Comparison with Recent specimens (e.g. NMNZ M.25864, Te Hapua, Parengarenga Harbour, Northland, 11 clean shells) showed that unabraded and uncorroded Recent specimens have the same 18–20 fine spiral cords between the suture and the upper, prominent peribasal cord as on Nukumaruan fossils, and differ from the perplexus form only in retaining the axial ridges until the end of the last whorl. Z. perplexus is merely a finely sculptured variant of Z. lutulentus. Finlay (Citation1924a:477) long ago pointed out that Ataxocerithium perplexum Marshall & Murdoch belongs in ‘Cerithidea’, is very similar to C. bicarinata (that is, Z. lutulentus), and suggested that it ‘is possibly only this species with the keels rubbed off’. Zeacumantus perplexus is here considered to be a finely sculptured variant of Z. lutulentus in which the axial costae fade out over the last whorl, and to be of no taxonomic significance or biostratigraphical utility.
‘Zeacumantus’ delicatus Laws, 1950 (Otahuhu well, Auckland, Waipipian) was based on juvenile specimens of Pyrazus ebeninus (Bruguière, 1792) (Beu Citation2004). This leaves Zeacumantus containing only the lineage of Z. lutulentus (including Z. tirangiensis (Marwick, Citation1926) (Tongaporutuan) and the single species Z. subcarinatus (G. B. Sowerby II, 1855). Several unnamed Miocene species apparently ancestral to Z. lutulentus are represented in GNS collections. Z. subcarinatus is one of the many molluscs with an apparent Haweran first appearance in New Zealand, but as it is a high-tidal rocky shore species, its true time range is unknown and it is completely unreliable for biostratigraphy.
Family Siliquariidae
Genus Stephopoma Mörch, Citation1860
Stephopoma Mörch 1860:42. Type species (by subsequent designation, Cossmann 1912:134): Vermetus roseus Quoy & Gaimard, 1834, Pleistocene and Recent, New Zealand.
Lilax Finlay Citation1926b:387. Type species (by original designation): Stephopoma nucleogranosum Verco, 1904, Recent, southern Australia.
Stephopoma roseum (Quoy & Gaimard, 1834) (A–G)
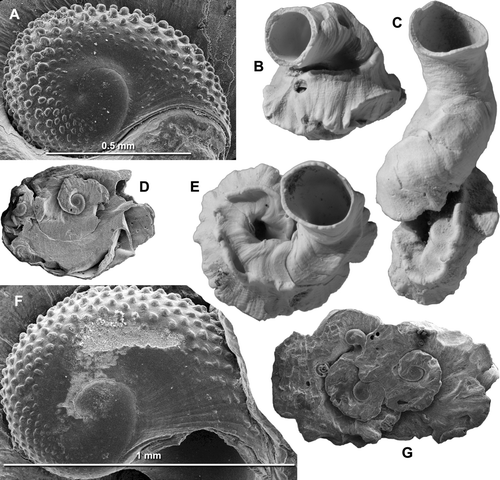
Fig. 7 Stephopoma roseum (Quoy & Gaimard). (A,D) GS15443, Y14/f505B, Te Piki, near East Cape, Haweran (OIS 7), SEM; D, basal view of 2 intergrown shells, protoconchs visible, width 7.0 mm; A, protoconch. (B,C,E) GS4013, R22/f6353, Tainui Shellbed (Castlecliffian, OIS 13), “the buttress”, Castlecliff, Wanganui, 3 specimens; B,C, lateral views, height 7.9 mm (B) and 17.8 mm (C); E, dorsal view, width 10.8 mm. (F,G) GS4013, R22/f6353, Tainui Shellbed (Castlecliffian, OIS 13), “the buttress”, Castlecliff, Wanganui, SEM; G, basal view of group of 3 shells, protoconchs visible, width 10 mm; F, protoconch.
Vermetus roseus Quoy & Gaimard 1834:300, pl. 67, –23.
Siliquaria rosea. ME Gray Citation1842, pl. 56, (copy of Quoy & Gaimard 1834, pl. 67, ); JE Gray 1850:83; JE Gray 1857:128.
Tenagoda rosea. H Adams & A Adams 1854:361 (as Tenagodus, p. xxxvii).
Stephopoma roseum. Mörch 1860:42; Mörch Citation1861:150; Hutton 1880:85; Suter Citation1913:261, pl. 15, , 6a (copy of Quoy & Gaimard 1834, pl. 67, ); Morton Citation1951:20,40, , 2, pl. 4, , 4–6; pl. 5, –14; pl. 6, –21, 23; Morton Citation1955:8, b; Morton & Keen Citation1960:27, , 12, 16; Powell Citation1979:129; Beu & Maxwell Citation1990:407; Bieler Citation1997:265, Fig. 25, 26.
Siliquaria laevigata. Hutton 1873a:31 (not Siliquaria laevigata Lamarck, 1818).
Vermetus (Stephopoma) roseus. Martens Citation1873:28; Tryon Citation1886:185, pl. 55, fig. 94 (copy of Quoy & Gaimard 1834, pl. 67, ).
Vermetus (Stephostoma) (sic) roseum. Clessin 1903:91, pl. 13, (copy of Quoy & Gaimard 1834, pl. 67, ).
Vermicularia rosea. Suter in Hutton Citation1904:76.
Vermicularia (Stephopoma) nucleogranosa. Suter Citation1906:328 (not Stephopoma nucleogranosum Verco, 1904).
Stephopoma nucleogranosum. Suter Citation1913:262, pl. 15, –7b (not S. nucleogranosum Verco, 1904).
Lilax nucleogranosa. Finlay Citation1926b:387.
Stephopoma rosea (sic). Fleming Citation1966:47; Spencer & Willan Citation1996:18; Spencer et al. Citation2009:207; Maxwell Citation2009:241.
Type material
Vermetus roseus seven syntypes (three dry shells in typothèque; four shells and seven animals in ethanol) MNHN moll. 22063 (not seen; Virginie Héros MNHN pers. comm. 14 July 2009); from ‘La rivière Tamise … Nouvelle Zélande’, ‘assez profondeur’, that is, Firth of Thames, Hauraki Gulf, New Zealand, at quite a great depth (Morton Citation1951:21).
Other material examined
Castlecliffian: Tainui Shellbed (OIS 13), abundant towards the top of the outcrop at ‘the buttress’, Castlecliff, some specimens attached to shells or large bryozoan colonies, many loose in the sediment; recorded by Fleming (Citation1953:229) as ‘Lilax n. sp. aff. nucleogranosum’ from GS 4013, R22/f6353, at ‘the buttress’ (11 in reference collection, five in later re-collection), and GS 4102, R22/f6516, at ‘the pinnacles’, both in the coastal outcrop; several early GNS collections from ‘Castlecliff’, including Suter collection no. 97 (four, one with tall, upright section); NMNZ M.27289, ‘Castlecliff’ (one), undoubtedly all from Tainui Shellbed. Haweran: GS4151, R22/f7416, Brunswick Formation (OIS 9), spur leading SE from summit of Mt Jowett, within Wanganui city (one broken, abraded specimen) (Fleming Citation1953:261); Te Piki, inland from Whangaparaoa, near East Cape (OIS 7; GS15443, Y14/f505B, 42 specimens, including several attached to Pecten shells, & some attached to Serpulorbis, in turn attached to Pecten shells; NMNZ M.15279, 10; M.40167, many); GS14777, O06/f067, intertidal flats 100 m SE of Waiparera church, NW Hokianga Harbour (OIS 7; 12, one with long, spiral upright section). Recent: 51 lots in NMNZ, ranging from the Three Kings Islands to the Bay of Plenty, and one specimen (protoconch only) from Mahia Peninsula (M.20929, no other data); one lot from the Chatham Islands (M.154146, 12–15 m, Ocean Bay, F Brook, four protoconchs). Specimens have been collected in depths from alive just below low tide to 188–228 m (M.113455, Rungapapa Knoll, Bey of Plenty, shell on rock, one specimen; possibly lived during a glacial lowered sea level) but this is the only sample collected in depths greater than 130 m, and few have been collected in more than 80 m. Most are beach shells, or were collected ‘under stones at low tide’ and on ‘rocks near low tide’.
One lot of New Zealand specimens (NMNZ M.94450, 24 km NW of Great I, Three Kings Islands, 710 m; seven protoconchs) has extremely fine granules on the protoconch, obviously finer than in Stephopoma roseum, and seems to represent another species, possibly S. nucleogranosum Verco, 1904, as illustrated by Bieler (Citation1997, –17; Bieler & Simone Citation2005, –8), although S. tricuspe Mörch, 1891 (Bieler Citation1997, –14) from eastern Australia is also very similar, differing in details of the opercular bristles (comparison with Australian specimens required). Another lot from the Kermadec Ridge (M.137514, seamount 130 km S of Esperance Rock, 32°28.30′S, 179°15.90′W, 538 m, on longline, one corroded teleoconch and four protoconchs) has markedly coarser and more elongate granules on the protoconch than in S. roseum, and represents a third, apparently unnamed species, not described by Bieler (Citation1997).
A further lot of New Zealand Holocene fossils (GS9151, Q07/f9602, Marsden Point oil installation water drillhole no. 3, Whangarei, Northland; 11 specimens; A–C) was assumed at first to belong in Stephopoma roseum, but SEM examination (B,C) showed that the protoconchs of these specimens, also, are much more finely granulose than those of S. roseum, suggesting they are also either S. nucleogranosum or S. tricuspe. The granules are not as obviously minute as in the Three Kings Islands specimens (NMNZ M.94450), and these two lots are possibly not conspecific. An adult teleoconch from the Marsden Point collection is also illustrated (A).
Fig. 8 (A–C) Stephopoma cf. nucleogranosum (Verco), GS9151, Q07/f9602, Marsden Point water well, Whangarei, Holocene; A, whole tube, height 19.4 mm; B,C, base of a tube, SEM. (D) Pelicaria zelandiae (Marshall & Murdoch), Marshall & Murdoch's figured syntype, TM8700, Waipipi, Waverley Beach, W of Wanganui, ex Marshall & Murdoch collection; H 36.4, D 26.8 mm. (E) Pelicaria arahura n. sp., paratype, TM8606, GS3046, J32/f9146, cutting S side Kumara-Inchbonnie Road, Big Hohonu River, Westland, Waipipian; height 43.8 mm.

Distribution
The molluscan faunal lists by Fleming (Citation1966) and Beu & Maxwell (Citation1990:407) included Altonian (early Miocene) records (queried) of Stephopoma roseum, but I do not know what this record was based on; no material is present in GNS. However, Laws (Citation1936:108; Citation1940a:441) recorded protoconchs of three apparently distinct species at Kaawa Creek, SW Auckland (Opoitian), differing in the size and spacing of the surface granules, indicating that early Miocene specimens are unlikely to be conspecific with S. roseum.
Other than the possible Altonian and Opoitian records, the fossil record is remarkable, as Stephopoma roseum is abundant in Tainui Shellbed (OIS 13) at Wanganui, and one specimen was collected by Charles Fleming from Brunswick Formation (OIS 9) on Mt Jowett, Wanganui, but there are no other records from Wanganui Basin. It is possible that the Brunswick Formation specimen is reworked from Tainui Shellbed, as it is incomplete and highly abraded. The only other fossil occurrences are within its present distribution, in the NE North Island warm province, at Te Piki and Hokianga Harbour. Other than a single record from the Chatham Islands, S. roseum is restricted to the NE North Island warm province at present, ranging from the Three Kings Islands to the Bay of Plenty and East Cape, with a single specimen from Mahia Peninsula. Morton & Miller (Citation1968:106–107, Fig. 38; pp. 135, 164, fig. 57; p. 274) described its occurrence in a narrow intertidal zone in shade under boulders, below the zone of the abundant tubicolous polychaete Spirobranchus cariniferus in the northern North Island. However, S. roseum is not a widespread zone-forming species, but has been found in scattered localities in samples ranging from one shell or a small number of shells to large masses of intertwined shells, up to 50×60×70 mm, mainly from Northland (Three Kings Islands to the Bay of Islands, Whangarei Heads, and offshore islands of Northland) to the Auckland beaches, including Takapuna reef, with several lots from the Bay of Plenty (e.g. off White Island; Motuhora or Whale Island), and a few localities extending as far south as East Cape (NMNZ M.20909, under boulders at low tide, Lottin Point, E of Cape Runaway, many short, intertwined shells in one mass ca 20×30×40 mm, with at least 22 brown, granulose protoconchs visible on the underside of the mass; M.154114, 79–83 m, Ranfurly Bank, East Cape, 1).
The Marsden Point drillhole fossils suggest that the fossil record of Stephopoma in New Zealand is more complex than has been suspected previously, and will repay fuller study.
Once again, fossils at Wanganui suggest significantly warmer sea temperatures than at present during interglacial periods, and planktotrophic larval transport from the NE North Island allowing range extension down the E North Island and through Cook Strait to Wanganui Basin during OIS 13 and possibly OIS 9 (the same stages as records of Zelippistes benhami). BA Marshall (NMNZ pers. comm. July 2009) pointed out that this is an established pattern for several molluscs (e.g. Micrelenchus rufozona A. Adams, 1853), Marshall Citation1998, fig. 63; Murexsul octogonus (Quoy & Gaimard, 1833), Marshall & Burch Citation2000, Fig. 37; Maoricrypta youngi Powell, Citation1940, Marshall Citation2003, ) although, in all three of these cases, a relict population remains living in W Cook Strait. Subsequent cooling of the SE North Island by northward extension of the cold Southland Current during glacial periods, which brought the Zygochlamys delicatula fauna north to Wanganui Basin early in Nukumaruan time, seems to have reduced the range of some of these species again to the NE North Island warm province. In some cases relict populations have remained in W Cook Strait, whereas in others the species disappeared from the southern North Island altogether. Stephopoma roseum, Maoricrypta costata, Zelippistes benhami, Prototyphis eos and possibly Stiracolpus vigilax can be viewed as examples of the group that became extinct altogether in the southern North Island.
Stephopoma roseum is abundant in the upper part of Tainui Shellbed at Castlecliff, but this is probably partly an environmental bias, reflecting the sediment-free, mainly shallow subtidal, cemented habit of this filter-feeding species. It is possible that this environment is otherwise not preserved at Wanganui, although the Brunswick Formation record shows than some specimens can be preserved out of their living habitat. S. roseum is a useful addition to the short list of species that extended their ranges briefly to Wanganui Basin from the northern North Island during Castlecliffian–Haweran time.
Dimensions
Tallest, ‘Wanganui’, Suter collection no. 97: H 22.5, D 8.8 mm; GS4013, Tainui Shellbed: H 6.2, D 11.1 mm; H 10.0, D 8.3 mm; H 17.8, D 8.3 mm (with tall, upright tube); H 5.9, D 9.2 mm; H 5.9, D 11.9 mm.
Remarks
The little irregularly coiled gastropod Stephopoma roseum has been illustrated only rarely previously; the only diagnostic illustration I am aware of is that by Suter (Citation1915, pl. 15, ). The shell was not illustrated by Morton (Citation1951). Suter's drawing gives a good idea of the distinctive low-coiled yet irregular shape, with a prominent flange extended ventrally from the whorls of most specimens to cement the shell to its substrate and, on many specimens, a further flange-like ridge around the upper side, facing away from the attachment area, forming a roughly rectangular cross-section. The attachment flange is irregularly but, on most specimens, quite coarsely ridged and puckered, copying the irregular substrate. Some specimens have many fine, low, closely spaced, slightly irregular spiral threads covering the interval between the two spiral flanges, and a few also have transverse ridges crossing the spiral ones, although most are smooth. Many specimens (including the majority of fossils in Tainui Shellbed) are essentially planispiral, with a final short upright section at right angles to the rest of the shell, but some large specimens develop a longer, straighter, cylindrical, upright but irregular section protruding well above the low initial coil. The illustration by Morton & Miller (Citation1968:164, fig. 57.7) shows a specimen in life orientation beneath a boulder, with no upright section and only the unattached side visible, whereas on most specimens the irregular flanges are more conspicuous.
Apart from the obvious, large flanges, the most distinctive character ensuring the identification of Stephopoma roseum is the small, almost planispiral protoconch of little more than one evenly rounded whorl, bearing rather closely spaced, large, hemispherical granules all over the exterior. On fresh specimens, including all live-collected ones, the protoconch is pale brown, but it fades to white on many specimens, and on some examined in NMNZ an initial brown protoconch has a white last half-whorl. Examination of the fossil specimens listed above showed that most retain a well-preserved protoconch on the base, surrounded by the attachment flanges. Several of the Tainui Shellbed specimens have two or three protoconchs on the base of one coiled shell, demonstrating that they are actually two or three closely intergrown specimens. All protoconchs examined have identical shape, size, and sculpture of large, hemispherical, closely spaced granules, in GS14777 (Hokianga), Te Piki and GS4013 (Tainui Shellbed). Examination of the large collection at NMNZ showed that all Recent lots from New Zealand have an identical protoconch, other than the one lot of a second species from 710 m off the Three Kings Islands, mentioned above, with a more finely granulose protoconch, and other than this sample and the Marsden Point drillhole specimens, all fossil and Recent specimens are considered to be conspecific. The single specimen from Brunswick Formation at Wanganui (GS4151) seems to be identical to the common form in Tainui Shellbed.
The original illustration of the shell and animal (Quoy & Gaimard 1834, pl. 67, ; copied by ME Gray 1842, pl. 56, , and by Suter Citation1913 [1915], pl. 15, ) shows a small, tall shell that is narrowly coiled for its entire length, the loose coil leaving a spiral groove between the whorls. Suter (Citation1913) had difficulties associating this shell with New Zealand material, and identified the more common, nearly planispiral shell form found under low-tidal boulders as the South Australian species S. nucleogranosum Verco, 1904. However, Morton (Citation1951:21, 22) matched the multifid opercular bristles of New Zealand material with the illustration by Quoy & Gaimard (1834) and distinguished the bristles and the finer protoconch sculpture of South Australian material of S. nucleogranosum, identifying all New Zealand material as S. roseum. The group of shells of S. nucleogranosum illustrated by Morton (Citation1951, pl. 7, Fig. 26) has wider, more upright and less obviously spiral growth than most New Zealand material, with no ridges on the evenly cylindrical upright sections, and more nearly resembles standard vermetids such as Serpulorbis than many New Zealand specimens do. Most specimens in the Marsden Point drillhole sample also lack obvious ridges on the exterior of the tube. However, many large lots in NMNZ consist of masses of densely intergrown tubes, twisted to varying degrees, and some are longer and more weakly coiled that the specimens illustrated by Morton (Citation1951, pl. 7, Fig. 26). Some of them resemble Quoy & Gaimard's (1834, pl. 67, ) drawing quite closely. In particular, NMNZ M.91895 (Doubtless Bay, Northland, from E. Willis of RV ‘Ikatere’, so presumably dredged offshore) is a large sample of small masses up to 60×40×40 mm of intergrown tubes, with many individual tubes up to at least 50 mm long, narrow, smooth, and weakly coiled, without an obvious attachment area or flanges, but with the diagnostic brown, granulose protoconch attached. They are considerably longer and more narrowly tapered than the tubes of S. nucleogranosum illustrated by Morton (Citation1951). Other masses of tubes collected subtidally are also quite large, up to ca 50×70×80 mm, and the degree to which tubes coil together into compact masses or are short and separate clearly depends on the environment; large masses and longer tubes presumably live in sheltered, enclosed habitats, whereas the more common, shorter, tightly coiled, flanged tubes have been collected under boulders low in the intertidal zone, or only slightly deeper. Some of the fossil specimens have a tall, narrowly coiled shell with only a small attachment area, a groove between the irregular whorls, and a weak flange up the entire tube. These also resemble Quoy & Gaimard's (1834, pl. 67, ) drawing quite closely.
Family Turritellidae
Remarks
The classification of the Turritellidae has always presented problems because of their abundance, their enormous diversity at the species level, and their simple shells with few taxonomic criteria. Authors who have contributed to their classification include Cossmann (1912), Guillaume (Citation1925), Dollfus (Citation1926), Douvillé (Citation1929:54–57), Merriam (Citation1941), Ida (Citation1952), Marwick (Citation1957a, Citationb, Citation1971), Kotaka (Citation1959), Olsson (Citation1964:185–194), Allison (Citation1965), Adegoke (Citation1967), Allison & Adegoke (Citation1969), Garrard (Citation1972, Citation1974, Citation1976), Golikov (Citation1986), Allmon (Citation1996) and DeVries (Citation2007). Kotaka (Citation1959:8–13) reviewed most of the classifications proposed before 1959, and Kantor & Sysoev (2006, pls. 22, 23) illustrated the two new genera proposed by Golikov (Citation1986). However, taxonomists not ‘specializing’ in turritellids have been reluctant to adopt a finely subdivided classification. Several North American authors have revised numerous taxa and used them for biostratigraphical zonation, but retained them in Turritella (including Bowles Citation1939; Palmer Citation1947; Saul Citation1983; extra-American zonations listed by Allmon Citation1988:259). The world classification proposed by Marwick (Citation1957a) does little to help resolve phylogenetic relationships within the family, as much because many of the world's species are not classified as because those that are classified are too finely subdivided. Marwick's review was limited to groups ‘that have already been named, so it is not at all comprehensive’ (Marwick Citation1957a:149). Also, Marwick (Citation1957a) commented favourably on Iredale's (Citation1924) proposal of Australian genera, and summarized his introduction by noting that ‘In a natural classification, [restricted dispersal ability] should be reflected by numerous localized genera’ (Marwick Citation1957a:149). Marwick's resulting assumption that similar turritellid outer lip profiles evolved independently in numerous parts of the world seems based on circular argument, and requires reassessment. It would be fair to say that most taxonomists since 1957 have disagreed with Marwick's conclusion, and continued to use Turritella for most or all species (e.g. Allmon Citation1988, Citation1992). Yet the type species of Turritella, T. terebra (Linné, 1758), has a distinctive, large, tall and narrow, strongly sculptured teleoconch with strongly and evenly inflated whorls and an unusually shallow outer lip sinus (although a closely related Pliocene species in Japan, T. perterebra Yokoyama, 1923, is much less extreme; Kotaka Citation1959, pl. 1, –7). Clearly, phylogenetic relationships in this family are expressed less than optimally if all species are placed in Turritella. I conclude that there are likely to be fewer, more widely distributed genera than Marwick (Citation1957a, Citationb, Citation1971) assumed, and that some species and genera are likely to have been dispersed widely around the world at various times, as in most other molluscan families, apart from a few extremely ‘holobenthic’ families such as the Volutidae (Bouchet & Poppe Citation1988:30). Even in Volutidae some Athleta species (especially Paleogene fossils) and some Calliotectum and Lyria species are exceptions to the ‘holobenthic’ rule, having dispersive, demersal larvae (Bouchet & Poppe 1995; Maxwell 2003).
Kotaka (Citation1978) pointed out that lineages placed in Stiracolpus (in New Zealand), Haustator (in Europe), Neohaustator (in Japan) and ‘Turritella’ (in California) evolved in parallel through a remarkably similar succession of morphological changes during Pliocene and Pleistocene time. Along with the close morphological similarity of all these groups, including a similar outer lip sinus, the closely parallel evolution might well be further evidence of a relationship between them at the generic level, the ‘parallelism’ resulting from some level of genetic exchange between them. If this is correct, this group, at least, probably belongs in one genus (perhaps with several subgenera), the earliest name for which is HaustatorMontfort, 1810. Marwick (Citation1957a:154) pointed out that both Thiele (1931 :181) and Wenz (1939:653) regarded Gazameda Iredale, 1924 as a synonym of Haustator, but Marwick (Citation1957a:154) kept them separate because of the distinct protoconch and early spiral cord development of Gazameda. ‘Gazameda is consequently far removed from Haustator in development, and is here given generic rank’ (Marwick Citation1957a:154), despite also pointing out that Australian middle Miocene species assigned to Gazameda have a turbiniform protoconch with a globose initiation (nucleus), as in most other turritellids including Haustator. One of the major themes stressed in the present series of papers is that such developmental differences are no longer regarded as characters distinguishing genera or subgenera; planktotrophy has been lost readily and independently by many gastropod species within originally planktotrophic clades. Marwick (Citation1957a) also pointed out the possibility that Zeacolpus Finlay, 1926 had developed from Haustator rather than from Leptocolpus Finlay & Marwick, Citation1937 (Paleocene, New Zealand): ‘the shallower sinuses of Leptocolpus … raises the question whether Zeacolpus is not closer to Haustator than to Leptocolpus’ (Marwick Citation1957a:156). Beu & Maxwell (Citation1990:156, pl. 14f) referred Zeacolpus grindleyi Marwick, Citation1971 (Duntroonian–Waitakian, late Oligocene, New Zealand) to Gazameda. G. grindleyi closely resembles the weakly sculptured Victorian Miocene species Gazameda victoriensis (Cotton & Woods, 1935), indicating that at least some New Zealand taxa are related phylogenetically to Australian Gazameda (= Haustator?) species.
This discussion suggests the possibility that there have been several separate range extensions to New Zealand by Haustator species, or at least by clades descended from Haustator: middle to late Eocene, to give rise to the earliest species assigned to Zeacolpus; late Oligocene, to give rise to ‘Gazameda’ grindleyi; and latest Miocene–basal Pliocene, to give rise to the group of species usually included in Stiracolpus. Rather than assigning all these species to Haustator at present, the traditional New Zealand genera are retained, keeping their probable close relationship to Haustator in mind. The family urgently requires a world-wide reassessment, and is another of the poorly understood families with few taxonomic criteria that will benefit greatly from comparion of DNA sequences. Only after a world revision, assessing the possible iterative evolution of lip profiles, will it be appropriate to change the status quo by adopting different genera in New Zealand. However, I have little doubt that the present classification bears little relationship to phylogeny.
A further example of the need to rethink the relationships of New Zealand turritellids is provided by the origin of the most abundant New Zealand Pliocene–Recent species, Maoricolpus roseus (Quoy & Gaimard, 1834). The true origins of this species can be seen in Garrard's (1972:324) statement about the relationships of Maoricolpus murrayanus (Tate, 1885). M. murrayanus is closely similar to M. roseus in all characters, although the spiral sculpture is slightly finer in M. murrayanus. The similarity is close enough that TA Garrard originally intended to synonymize the two species, and was only persuaded not to by me, because of the different time range (early to middle Miocene) and geographical range (southern Australia) from that of M. roseus (Pliocene–Recent, New Zealand). It is ironical, then, that M. roseus invaded D'Entrecasteaux Channel in SE Tasmania during the twentieth century, and is now abundant there (Greenhill Citation1965:67), as well as occurring at many other localities in Tasmania (Greenhill Citation1965; Bax et al. Citation2003; Probst & Crawford 2008) and is spreading northwards to inhabit the Furneaux Group islands in Bass Strait (Allmon et al. Citation1994), with records from Victoria and New South Wales as far north as Sydney (Scott Citation1997:25; Bax & Gowlett-Holmes Citation2000). M. roseus was not recorded from Tasmania by earlier authors such as May (Citation1921) (Bax et al. Citation2003). Garrard (Citation1972:322) recorded M. roseusalso from 110–150 m off Cape Moreton, S Queensland, but Bax et al. (Citation2003) doubted the reliability of this record, and collectors in S Queensland (A Limpus, Bundaburg; T Whitehead, Brisbane; pers. comm. June 2008) have been unable to substantiate it, so M. roseus evidently occurs only around Tasmania and in Bass Strait to Sydney, although the lack of natural predators seems to be making it possible for M. roseus to spread in Australia (Allmon et al. Citation1994). If the type species of Maoricolpus invaded New Zealand from Australia at the beginning of Pliocene time, what are the real phylogenetic relationships of the other New Zealand taxa referred by Marwick (Citation1971) to Maoricolpus? They seem unlikely to belong in the same clade. Marwick obviously realized that not all species assigned to the traditional New Zealand genera belong there, in a phylogenetic sense, and he apparently was aware that his classification was not final. Much remains unknown about the phylogeny of these taxa, and their inclusion by Marwick (Citation1971) in a few restricted New Zealand genera was a matter of practicality. Determining the phylogeny of world turritellids remains a task for molecular phylogeny in the future.
Genus Stiracolpus Finlay, 1926
Stiracolpus Finlay Citation1926b:389. Type species (by original designation): Turritella symmetrica Hutton, 1873, late Pliocene (Nukumaruan)–Recent, New Zealand.
Remarks
Within New Zealand Turritellidae, Marwick's (1957b, 1971) two monographs display a marked difference in approach, and consequently in biostratigraphical utility. The later monograph, with its careful stratigraphical segregation of species defined within relatively long lineages of Zeacolpus, Maoricolpus and apparently related larger turritellids (remembering the proviso about Maoricolpus pointed out above), resulted in species that are recognizable by subsequent workers, and are useful in biostratigraphy—within limits, because of the inherent similarity of most turritellids. In contrast, Marwick's (1957b) earlier monograph of Pliocene to Recent Stiracolpus species was hampered by a bewildering array of variation interpreted with a very narrow species definition, almost an unconsciously typological viewpoint. Re-collection of the Stiracolpus species in situ in the Castlecliffian rocks of Wanganui Basin and examination of Marwick's collections in GNS demonstrated that Marwick regarded the coarsely sculptured end-members of the population in each formation as one species, the finely sculptured end-members as another species, and all the intergrading, intermediate specimens as hybrids. The resulting plethora of species has proved impossible for subsequent workers to distinguish, and those familiar with (in particular) Pleistocene and Recent turritellids in New Zealand (RM Carter, James Cook University, Townsville pers. comm.; JA Grant-Mackie, University of Auckland pers. comm.; BA Marshall, NMNZ pers. comm.) agree with my opinion that few of Marwick's taxa are real biological species. The assessment of biostratigraphically useful taxa within Pliocene–Pleistocene turritellids therefore must commence with a new taxonomy of the species. Once again, this is nearly impossible with the characters visible on the shells, and will only be comprehensible after the taxonomy and variation of Recent species have been resolved by molecular techniques.
Nevertheless, some points seem reasonable to conclude, after 50 years’ experience with these shells:
-
The great majority of Marwick's (1957b) late Pliocene–Recent ‘species’, with numerous prominent spiral cords, are part of the variation of either Stiracolpus symmetricus (Hutton, 1873) (southern forms) or S. pagoda (Hutton, 1873) (all forms living now in central and northern New Zealand). While S. symmetricus is most strongly sculptured at present around Foveaux Strait and Stewart Island, where most specimens have three equally prominent, narrow spiral cords on a strongly convex whorl profile, very similar forms occur in Nukumaruan rocks in Wairarapa district (named Zeacolpus delli vellai Marwick, 1957), and an astonishing range of variants on the theme occurs throughout Nukumaruan and Castlecliffian rocks of Wanganui Basin, the East Coast Basin and North Canterbury. The more weakly sculptured Recent central New Zealand forms named (e.g.) Z. blacki and Z. delli by Marwick (Citation1957b) all intergrade, and are probably variants of the northern New Zealand species S. pagoda (Reeve, 1849). Names that apparently were based on trivial variants of S. symmetricus are Z. delli murdochi, Z. delli granti, Z. delli vellai, Z. robinae and Z. shepherdi, all of Marwick (Citation1957b). Names that seem to be based on trivial variants of S. pagoda are Zeacolpus ascencus, Z. blacki, Z. delli and presumably Z. knoxi, Z. knoxi tardior, Z. uttleyi and Z. uttleyi ruahinensis, all of Marwick (Citation1957b), Z. pagoda (Reeve, 1849), Z. pagoda powelli Marwick, 1957, Z. maorius (Powell, Citation1940), Z. mixtus (Finlay, 1930) and Z. waikopiroensis (Suter, Citation1917). More than one species is possibly hidden in the complex variation of S. pagoda (BA Marshall, NMNZ pers. comm. March 2006). Analysis of the ontogeny of development of the spiral cords, with a broader interpretation than Marwick's, could perhaps aid understanding of this group.
-
An exception to the Wanganui Basin ‘species’ all being synonyms of S. symmetricus is provided by Stiracolpus vigilax (Landguard Sand, OIS 9, and a few specimens from OIS 11; one specimen from Te Piki, near East Cape, OIS 7), which is smaller and much more strongly unicarinate than any other species at Wanganui. It is very similar to and possibly conspecific with S. ahiparanus (Powell, Citation1927) (Recent, NW North Island), although the Wanganui material reaches only about half the height of Recent material of S. ahiparanus. However, S. ahiparanus and S. vigilax are also both possibly further synonyms of S. pagoda.
-
The extremely finely sculptured specimens at Wanganui (most notably in Kupe Formation, OIS 17; a few records from shellbeds deposited during OIS 19 and 15) identified by Marwick (Citation1957b) as ‘Zeacolpus (Stiracolpus) cf. waikopiroensis’ possibly represent incursions of the northern warm-water S. pagoda species group into Wanganui Basin. This needs confirmation, as they might also merely be unusually finely sculptured end-members of S. symmetricus. However, it is also possible that S. waikopiroensis itself (that is, Hawke's Bay Nukumaruan weakly sculptured forms, assumed here to be finely sculptured end-members of the form identified by Marwick [Citation1957b] as Z. uttleyi) is based on a member of the S. pagoda species group; these specimens closely resemble the northern North Island Recent form known as S. maorius.
-
Stiracolpus huttoni (Cossmann, 1912) represents a different species group from the complex of S. symmetricus and S. pagoda, with only two prominent, widely spaced spiral cords, and no other obvious spiral sculpture. It is remarkably unvaried, implying that it had planktotrophic rather than direct development. I am not aware of records younger than late Nukumaruan.
-
Earlier (Opoitian–Mangapanian) species such as Stiracolpus kaawaensis (Laws, Citation1936), S. procellosus (Marwick, Citation1931), S. propagoda (Laws, 1940) and possibly S. quennelli (Marwick, 1957) and S. wiltoni (Marwick, 1957) seem to be distinct, much less variable species. This implies that (as with Pelicaria; see below) the inception of the lecithotrophic (probably direct) developmental mode that produced the enormously variable ‘species complex’ of S. symmetricus and S. pagoda was an early Pleistocene (early Nukumaruan) event. Cooling of the sea seems to have led to similar developmental changes and a marked increase in variation of shell characters during early Nukumaruan time by S. symmetricus, Pelicaria vermis and Aoteadrillia wanganuiensis (see below). S. huttoni seems to be an example of the other possibility, stenothermal warm-water taxa that simply became extinct when sea temperatures reached a critical threshold during late Mangapanian–early Nukumaruan glaciations.
This summary results in a group of four or five Pliocene (Opoitian–Mangapanian) species—those listed in the paragraph above plus Stiracolpus huttoni—and only a further four or five Nukumaruan–Recent species. One obvious biostratigraphically useful result is the extinction of S. huttoni late in (at the end of?) Nukumaruan time. Another is the apparent arrival of S. symmetricus (in the broad sense used here) in the southern North Island during early Nukumaruan time, along with the cold-water fauna dominated by Zygochlamys delicatula (Fleming Citation1944). Much further work is required to understand this taxonomically ‘difficult’ genus in New Zealand, and a detailed taxonomic revision obviously is not appropriate here.
Species I suggest, therefore, should be recognized in Stiracolpus are:
Stiracolpus ahiparanus (Powell, Citation1927), Recent, NW Northland (possibly = S. pagoda; possibly = S. vigilax).
Stiracolpus huttoni (Cossmann, 1912) (J), Opoitian–Nukumaruan, Westland, Wanganui and Hawke's Bay.
Stiracolpus kaawaensis (Laws, Citation1936), Opoitian, Kaawa Creek, SW Auckland.
Stiracolpus pagoda (Reeve, 1849), Recent, central and northern North Island (and Pleistocene, Wanganui?) (the species name is an indeclinable noun); probably = Z. huttoni ohopeus Marwick, 1957, Castlecliffian (OIS 15?), Ohope, Bay of Plenty.
Stiracolpus procellosus (Marwick, Citation1931), Opoitian, N Hawke's Bay and N Wairarapa.
Stiracolpus propagoda (Laws, 1940), Mangapanian, Wanganui Basin (possibly = S. symmetricus).
Stiracolpus symmetricus (Hutton, 1873) (H), Mangapanian?; Nukumaruan–Recent, central and southern New Zealand (= S. delli vellai, etc.).
Stiracolpus vigilax (Marwick, 1957) (possibly = S. ahiparanus) (I), OIS 11–9, Wanganui; one specimen in GS10910, Y14/f7505B, Te Piki member, road cut 6 km E of Whangaparaoa, near East Cape, OIS 7 (again suggesting that S. vigilax might be just a sculptural variant of S. pagoda).
A small list remains of possibly distinct species requiring further study, as their status is unknown:
Stiracolpus hurunuiensis (Marwick, 1957), Waipipian(?) redeposited fauna at Tormore, North Canterbury (this mixed fauna apparently resulted from redeposition of Waipipian fossils during Nukumaruan time; Beu Citation1977).
Stiracolpus nanulus (Marwick, 1957), Nukumaruan, Kereru, Hawke's Bay (= juvenile S. symmetricus?).
Stiracolpus quennelli (Marwick, 1957), Mangapanian, S Hawke's Bay (possibly = S. symmetricus).
Stiracolpus wiltoni (Marwick, 1957), Opoitian, N Wairarapa.
Type material
The opportunity is taken to illustrate Hutton's (1873b:13) ‘Shakespeare Cliff’ paralectotype of Turritella (Zaria) tricincta (TM5375; junior primary homonym of T. tricincta Borson, 1821, of T. tricincta Münster, 1841, and of T. tricincta Morris in Strzelecki, 1845) (almost certainly collected from Shakespeare Cliff Siltstone, where this form is common), as this specimen has never been illustrated previously (I; H 30.7, D 7.7 mm). Marwick (Citation1957b:32) recorded S. delli murdochi Marwick, 1957 as the form common in Shakespeare Cliff Siltstone, and Dell et al. (1964):448) identified Hutton's ‘Shakespeare Cliff’ syntype as S. delli murdochi. Hutton (1873b:13) recorded T. tricincta also from ‘Awatere; Waikari (small variety); Weka Creek; Mount Cookson’, and ‘var. B’ from ‘Kanieri; Hatter's Creek; Mount Cookson’, so it represents several different genera of Pleistocene to Oligocene age. The illustrated shell is a good example of a Wanganui Castlecliffian specimen of Stiracolpus symmetricus. However, it is not a name-bearing type. Although Harris (1897:241) proposed Turritella kanieriensis as a replacement name for T. tricincta Hutton, because it is a junior primary homonym, he did not mention a type specimen. Dell et al. (Citation1964) described the nomenclatural complications of this case; the International Commission on Zoological Nomenclature (Opinion 810 Citation1967) designated the specimen from ‘Kanieri’ illustrated by Suter (Citation1914, pl. 16, ) as the lectotype. This is a specimen of Hutton's (1873b:13) T. tricincta var. B. The name T. kanieriensis therefore applies to a latest Miocene–early Pliocene (Kapitean–Opoitian) species of Zeacolpus rather than to the Castlecliffian Stiracolpus species.
I also illustrate the holotype of Zeacolpus (Stiracolpus) vigilax Marwick, 1957 (C; TM1686, GS4003, R22/f 7394, Landguard Sand, Landguard Bluff, Wanganui, OIS 9; H 15.3, D 4.75 mm) and one of Marwick's (1957b):35, pl. 5, ) illustrated specimens of Stiracolpus huttoni, from Kaniere (K; TM1665, GS2877, J33/f7076, Lake Kaniere Road, opposite Kaniere powerhouse, Westland; H 18.3, D 7.5 mm; Opoitian; the locality from which Pelicaria parva (Suter, Citation1915) is listed below). It should be noted that Hawke's Bay Nukumaruan specimens of S. huttoni have a markedly narrower spire angle than Kaniere specimens, and Marwick (1957b:35) commented on the variable spire angle.
Clade Littorinimorpha
Family Littorinidae
Genus Risellopsis Kesteven, 1902
Risellopsis Kesteven 1902b:319. Type species (by original designation): Adeorbis varius Hutton, 1873, Pleistocene and Recent, New Zealand.
Risellopsis varia (Hutton, 1873) (Fig. 5E,6B)
Adeorbis varius Hutton 1873a:35.
Risella(?) varia Hutton Citation1878:27.
Fossarina varius. Hutton 1880:79; Hutton Citation1882:164.
Risellopsis varia Kesteven Citation1902:320, Fig. 29–32; Suter Citation1913:191, pl. 35, ; Odhner Citation1924:21; Finlay Citation1928:241; Powell Citation1937:67, pl. 9, ; Fleming Citation1966:42; Morton & Miller Citation1968:80,353, Fig. 23, 77; Pilkington Citation1974:411, ; Pilkington Citation1976:338, a–c; Powell Citation1979:88, , no. 7; Reid Citation1988:142, , 25–27; Spencer & Willan Citation1996:18; Spencer et al. Citation2009:206; Maxwell Citation2009:240.
Risellopsis varia var. carinata Kesteven Citation1902:321, Fig. 33–35; Suter Citation1913:192, pl. 35, ; Bucknill Citation1924:38, pl. 6, , 21a; Finlay Citation1928:241.
Type material
Adeorbis varius, lectotype (designated by Reid Citation1988:142) NMNZ M.160, from ‘Stewart's Island’; paralectotype NMNZ M.1822, from the Chatham Islands. Risellopsis varia var. carinata Kesteven, holotype AMS C.11422, from ‘New Zealand’ (Reid Citation1988:142) (the last not seen).
Other material examined
Castlecliffian: 1 m boulder of Teer Formation (about OIS 25) on beach, ca 1.1 km E of Whiskey Creek and 2 km W of Teer Creek, N coast of Cascade Point, S Westland (GS12293, E38/f13, few specimens; Beu in Sutherland et al. Citation1995:447). Haweran: Brunswick Formation (OIS 9), Mount Jowett, Wanganui (Fleming Citation1953:260) (GS4151, R22/f7416, one); Brunswick Formation (OIS 9), Sewell Road, N of Brunswick Road, W of Wanganui (GS12425, R22/f10, one); 8.4 m below sea-level in Waikanae Beach test bore, Gisborne, Holocene (Y18/f33, GNS, one); ‘paua kill’ site on 1855 intertidal slope ca 400 m east of Cape Turakirae, Wellington, Holocene (GS15119, R28/f62, ca 30); 1855 intertidal slope, 100 m north of scientific reserve boundary, Cape Turakirae, Wellington, Holocene (GS15121, R28/f072, one); ‘lepadomorph site’ beneath large boulder on 1855 intertidal slope, ca 400 m east of Cape Turakirae, Wellington, Holocene (GS15122, R28/f062A, 14). Specimens also occur in several Holocene samples from trenches dug through the cosiesmically uplifted terraces at Table Cape, Mahia Peninsula.
Distribution
Rissellopsis varia is one of several species with an apparent OIS 9 first appearance (around Wanganui, particularly at Mt Jowett, in Brunswick Formation), but which actually occur significantly earlier. Again, though, as it is a high-tidal rocky shore species, its first occurrence is highly unreliable for biostratigraphy. Specimens are now recorded from deposits as old as at least OIS 25 (Teer Formation, Cascade, South Westland).
Dimensions
RM704, Takapuna Beach, Auckland: H 3.1, D 5.0 mm; H 3.3, D 4.9 mm; H 3.6, D 5.1 mm; GS4151, Brunswick Formation, Mount Jowett, Wanganui: H 2.3, D 3.8 mm.
Remarks
Rissellopsis varia is a small (up to ca 6 mm diameter) relative of Bembicium, with a strongly depressed, coarsely spirally sculptured shell. Reid (Citation1988) reviewed the characters, range, ecology and variation, and little need be added here. It is limited to exposed high-tidal rocky shores throughout New Zealand. Many specimens shelter under dense sheets of the mussel Limnoperna pulex (Lamarck, 1819).
Family Struthiolariidae
Genus Pelicaria Gray, Citation1857
Pelicaria Gray 1857:77. Type species (by monotypy) ‘P. vernis’, that is, Buccinum vermis Martyn, 1784 (available, ICZN Opinion 479 Citation1957), Pleistocene and Recent, New Zealand.
Pelicaria granttaylori n. sp. (9A–E)
Fig. 9 Pelicaria granttaylori n. sp. (A,C) holotype, TM8594, GS11617, T22/f8537A, Hautawa Shellbed (early Nukumaruan), Hautawa Road, between Murimotu Valley, Rangitikei, and Turakina Valley; height 51.1 mm. (B) paratype, TM8701, GS4206, S22/f6444, Hautawa Shellbed (early Nukumaruan), Upokonui Stream, Parihauhau Road, NE of Wanganui; height 50.2 mm. (D,E) paratype, TM8702, GS13076, W19/f027, sandstone upstream from highway, Mohaka River, N Hawke's Bay, Mangapanian; height 56.3 mm.

Type material
Holotype (TM8594) and three paratypes (TM8595-7) from GS11617, T22/f8537A, Hautawa Shellbed (basal Nukumaruan), disused Hautawa Road, between West Road, Murimotu valley, N of Hunterville, Rangitikei valley, and Otiwhiti Station, Turakina Valley Road.
Other material examined
Mangapanian: GS4220, S22/f6450, lower Okiwa Group, Paparangi Road, N of Wanganui (five paratypes; a small form with narrow cords, otherwise resembling P. arahura n. sp.); GS4124, R22/f6543, Wilkies Shellbed, Wilkies Bluff, mouth of Waitotara River, W of Wanganui (two fragmentary paratypes, similar to P. zelandiae (Marshall & Murdoch, Citation1920) in sculpture); GS4224, R22/f6467, Wilkies Shellbed, Ohie Stream, W of Wanganui (two fragmentary paratypes, similar to P. zelandiae in sculpture); GS13076, W19/f027, brown sandstone upstream from highway in Mohaka River, N Hawke's Bay (one large paratype, TM8702, with many narrow cords, otherwise shaped as in GS4220; D,E). Recorded also by Fleming (Citation1953:120, 124, 129) from Makokako Sand (GS4221, R21/f9504, Kauarapaoa Valley, N of Wanganui); Wilkies Shellbed (GS4209, R21/f8501, Paparangi Road, N of Wanganui), and Te Rama Shellbed (GS4188, R22/f6471, near Te Rama trig. station, W of Wanganui). Early Nukumaruan: GS4206, S22/f6444, Hautawa Shellbed, Upokonui Stream, Parihauhau Road, 5 km NE of junction with Highway 4 (Parapara Road), NE of Wanganui (four paratypes, incomplete to almost complete, distorted; three resemble P. zelandiae in shape and cord prominence; one resembles holotype, illustrated, C, TM8701); GS7837, T22/f8504, above Hautawa Shellbed, Hautawa Road, between Turakina and Rangitikei valleys (one fragmentary paratype). Recorded also by Fleming (Citation1953:131,134) from Hautawa Shellbed (GS3096, T22/f8492, Hautawa Road; GS4204, S22/f6442, near Te Wharau trig. station, Mangamahu Stream, 12 km NE of Mangamahu, NE of Whangaehu valley) and upper Okiwa Group (GS4346, S22/f6471, Turakina River, 1 m above Hautawa Shellbed; GS4359, S22/f6484, Mangawhero Valley, near Klein Trig. Station, 17 m above Hautawa Shellbed; GS4354, S22/f6479, Hales Road, Turakina, 12 m below Ohingaiti Sand).
Distribution
Mangapanian and early Nukumaruan only. Pelicaria granttaylori n. sp. is common in Wilkies Shellbed and higher Mangapanian shellbeds in Wanganui Basin, and in Hautawa Shellbed; it tapers off in abundance higher in the succession, and is not known higher than 12 m below Ohingaiti Sand, still quite low in Nukumaruan rocks. I am aware of specimens only from Wanganui Basin (perhaps because of its restriction to near-shore facies), apart from a single large paratype from the Mohaka River, N Hawke's Bay.
Dimensions
Holotype: H 51.1, D (distorted) 33.1 mm; paratype GS4206: H (incomplete) 50.2, D 34.3 mm; paratype GS13076: H 56.3, D 39.7 mm.
Description
Shell characteristic of Pelicaria in overall appearance, size, and aperture shape and thickening. Protoconch not seen. Teleoconch of 5–5.5 whorls. Spire moderately tall to rather short for genus; last whorl occupying 2/3 to 3/5 of height. Suture moderately to deeply channelled, channel ranging from narrow, weakly concave, horizontal ramp (in holotype and some other specimens from Hautawa Shellbed) to deeply and narrowly excavated as in P. zelandiae (in older specimens, and some from Hautawa Shellbed). Outline below sutural channel straight, vertical in some specimens (which, therefore, resemble Pelicaria zelandiae in outline shape), or with weakly defined, outward-sloping sutural ramp, weakly defined but obvious peripheral angulation, and vertical whorl mid-section in other specimens; peribasal angulation weakly defined, rounded in some specimens, more sharply defined by single protruding spiral cord in others. Spiral sculpture of some specimens closely resembling that of P. zelandiae, with numerous low, narrow, closely spaced cords not affecting outline shape; in others (including holotype) of relatively few prominent, convex-crested spiral cords, with narrow interspaces each filled by 1–3 narrow, closely spaced secondary cords; three prominent cords on spire whorls and four on last whorl (including subsutural cord) and one secondary cord in each interspace on holotype, slightly more numerous cords on some larger specimens; 6–8 cords on base, slightly lower, narrower and more closely spaced than on whorl sides, decreasing in size anteriorly. Uppermost prominent spiral cord bearing weakly defined, low, irregular nodules in a few specimens. Exterior of well-preserved specimens closely covered with very fine spiral threads. Aperture large, relatively wide as in all Pelicaria species, with relatively narrow, evenly rounded terminal varix and smoothly thickened, narrow inner lip (similar in width to outer lip), weakly extended to left over previous whorl in some specimens; outer lip sinuous as in all Pelicaria species, deflected abaperturally below suture, followed by obvious, evenly convex adapertural flexure at mid-lip, followed again below by abapertural flexure leaving shallow, evenly concave anterior sinus below junction with columella. Inner lip of uniform thickness in many specimens, that is, without obvious parietal thickening; slight parietal thickening in a few specimens (including holotype) producing shallow, weakly defined posterior sinus.
Remarks
Pelicaria arahura n. sp. and Pelicaria granttaylori n. sp. share a similarity in shape to P. zelandiae (D; Beu & Maxwell Citation1990:294, pl. 36p) brought about by the shallow but consistent spiral depression above the peribasal spiral cord, the obvious sutural channel, and the adapically inward-sloping sutural ramp below the sutural channel. Most specimens of P. granttaylori n. sp. are taller and narrower than P. arahura n. sp., and P. arahura n. sp. lacks the narrow secondary spiral cords present on nearly all specimens of P. granttaylori n. sp. The inward-sloping sutural ramp and the consistent presence of the spiral depressed area between the second and third main spiral cords also distinguish both P. arahura n. sp. and P. granttaylori n. sp. from all younger species, particularly the early acuminata form of P. vermis (Martyn, 1784) (see below). P. arahura n. sp. also is shorter-spired and has more nearly parallel whorl sides, producing a more rectangular shell shape and a more stepped spire than in younger forms of P. vermis and most specimens of P. granttaylori n. sp., except for some specimens of the bradleyi form of P. vermis (Recent, N coast of the South Island) which are similar to P. arahura n. sp. in all characters except the much fewer, more prominent spiral cords of P. arahura n. sp.
Pelicaria zelandiae differs from P. granttaylori n. sp. in its straighter whorl sides and much finer and closer spiral sculpture. Most specimens also have a much more obvious, deeper and wider sutural channel than P. granttaylori n. sp., with a narrow rim, and a less obvious sutural ramp and almost straight whorl outlines between the margin of the sutural channel and the prominent, narrow, consistent peribasal angulation. Most specimens of P. zelandiae also have sculpture of many narrow, even, closely spaced spiral cords covering the sides and base of the last whorl, although it is a little coarser on a few specimens. In other words, P. granttaylori n. sp. is much more variable than P. zelandiae. P. zelandiae consistently has a wide sutural channel and vertical whorl sides, and varies mainly in the prominence of the spiral cords—weak, numerous, and closely spaced on most specimens, but fewer and more prominent on others—whereas P. granttaylori n. sp. is much more variable in outline shape, in the prominence of the sutural channel, and in the prominence of the spiral cords. P. zelandiae is particularly well-known from the Waipipi Shellbeds at Waverley Beach, Wanganui, but also occurs at Waihi Beach, Hawera (Waipipian), and at Whakino, South Taranaki (from P. Marshall's collection; Waipipian). Poorly preserved Opoitian specimens of a similar form occur in GS7193, N34/f6789, Omihi Creek, N Canterbury (two moulds with fine spiral sculpture, and with slightly more convex whorls than Waipipian specimens of P. zelandiae).
The other main Pliocene species that needs to be distinguished from Pelicaria granttaylori n. sp. is P. canaliculata (Zittel, 1864) (D; Beu & Maxwell Citation1990:293, pl. 36q), which differs in its more evenly convex whorl shape, with a less obvious peribasal angulation, sutural channel and sutural ramp than in P. zelandiae and P. granttaylori n. sp., and in having four more even, prominent, high, vertical-sided to undercut spiral cords on the sides of the last whorl. Specimens are best known from Starborough Formation in the Awatere district (Starborough Creek; lower Awatere River; coast 2 km N of Cable Station Road, at mouth of Blind River), but also occur less commonly in Greenwood Formation in the lower Waipara Gorge, Canterbury (GS4997, N34/f6192, one fragment); near Taueru, central Wairarapa (GS94, R27/f8495, three poor moulds); in the Mangahao district in N Wairarapa (Neef Citation1970); and rarely in the Waipipi Shellbeds, Waverley Beach, Wanganui (GNS: one collected by P Marshall, one collected by FD Chambers); all of Waipipian age. P. procanalis (Beu, Citation1970) from Opoitian rocks in the Awatere Valley seems likely to have been ancestral to P. canaliculata, and differs from it in its much lower spiral cords. P. arahura n. sp. also is similar to P. procanalis, again differing in its more prominent spiral cords, and in the presence of the obvious, inward-sloping sutural ramp.
Fig. 10 (A,B) Pelicaria arahura n. sp., holotype, TM8598, GS12289, J33/f066, conglomerate at top of Eight Mile Formation (Waipipian), head of E tributary of Greeks Creek, S side of Arahura Valley, Westland; height 41.4 mm. (C,E,F,H) Pelicaria vermis (Martyn); C, RM2974, extreme “bradleyi form”, Recent, Rabbit Island Beach, Nelson; height 36.6 mm; E, “convexa form”, GS10748, S28/f6206, Pukenui Limestone (early Nukumaruan), above junction Mangaopari Stream and Makara River, S Wairarapa; height 48.0 mm; F, holotype of Struthiolaria acuminata Marwick, TM7682, GS1040, S27/f8448, Pukenui Limestone (early Nukumaruan), White Rock Road, Makara River, S Wairarapa; height 42.9 mm; H, lectotype of Struthiolaria crenulata Lamarck and neotype of Buccinum vermis Martyn, Murex australis Gmelin, Struthiolaria inermis G. B. Sowerby I, and Struthiolaria tricarinata Lesson, MNHN moll.20522, Recent, New Zealand (Waikanae-Paraparaumu coast, W Wellington?), collected by Péron & Lesueur; height 40.8 mm (MNHN photo by V. Héros). (D) Pelicaria canaliculata (Zittel), specimen resembling Zittel's illustration, “Awatere”, ex Marshall & Murdoch collection, presumably from Starborough Formation (Waipipian), Seddon; height 42.8mm. (G) Pelicaria rugosa (Marwick), Neef's (1970) illustrated specimen, T24/f8049, Marima Sandstone (early Nukumaruan), tributary of Mangahao River E of Marima School, N Wairarapa; height 45.3 mm.

The final named Pliocene species requiring comparison is Pelicaria cestata Marwick (Citation1965:32, pl. 7, ). The holotype (TM3730), from GS1586, X19/f7480, sandstone above Tahaenui Limestone, Makeretu Stream, Mangapoike, N Hawke's Bay (Waipipian), with two paratypes, has prominent, evenly convex spiral cords closely resembling those of the much younger acuminata form of P. vermis (see below), but a little narrower, more widely spaced and smoother-surfaced than in any specimens of P. vermis, and with a more obvious sutural channel, a secondary spiral cord below the channel on most specimens, and distinctive, more prominent spiral cords on the base. It seems to result from an independent evolution of an acuminata-like form, although its origin is not obvious. Other specimens in GNS are from GS4887, V22/f8511A, Hickey's Stream, off Argyll Road, Te Aute, S Hawke's Bay, Waipipian (one, smaller than others); GS1541, X18/f9477, sandstone above Tahaenui Limestone, Te Reinga Falls, Wairoa River, N Hawke's Bay, Waipipian (one incomplete); and GS8000, X19/f7642, sandstone overlying Tahaenui Limestone, Mangapoike River below mouth of Haupatanga Gorge, N Hawke's Bay, Waipipian (one incomplete).
Comparison of the range of variation of Pelicaria granttaylori n. sp. with that of the highly variable P. vermis, as interpreted broadly here (see below), suggests that like P. vermis, P. granttaylori n. sp. was much more variable in most characters than other Pelicaria species are. Therefore, it probably had direct development. Direct development, apparently an adaptation to cooling marine temperatures, evolved separately in both the P. zelandiae-granttaylori and P. canaliculata-vermis lineages, but in the P. zelandiae lineage it was adopted earlier (during early Mangapanian time) than in the P. canaliculata-vermis lineage (during earliest Nukumaruan time). Nevertheless, the P. zelandiae lineage apparently was unable to adapt to the continued cooling of marine temperatures and became extinct quite early in Nukumaruan time (the youngest record of P. granttaylori n. sp. is in GS4354, S22/f6479, 12 m below Ohingaiti Sand, early Nukumaruan). In contrast, P. vermis is still a common, widespread species around New Zealand at present. The much greater range of variation than in P. zelandiae, indicating direct development, and the rarity of specimens closely resembling typical Waipipian specimens of P. zelandiae justify the recognition of P. granttaylori n. sp. as a separate species.
Etymology
It gives me great pleasure to honour, with the name for this species, the memory of that great character and ‘ideas man’, Thomas Ludovic Grant-Taylor (18 October 1923–9 May 1982; New Zealand Geological Survey 1950–1982) (Hornibrook Citation1982). Tom was a frequent collecting companion, along with Norcott Hornibrook, during my early years in North Island east coast stratigraphy, and helped collect many of the fossil localities where Pelicaria species are found.
Pelicaria arahura n. sp. (E, 10A, B)
Type material
Holotype (TM8598) and two incomplete paratypes (TM8599-8600) from GS12289, J33/f066, uppermost concretionary, blue-grey, shelly conglomerate at top of Eight Mile Formation, Blue Bottom Group, head of E tributary of Greeks Creek, S side of Arahura Valley, Westland, Waipipian; one paratype (TM8606) from GS3046, J32/f9146, S side Kumara-Inchbonnie Road, E of former Westbrook Hotel, 2.8 km W of Three Mile Creek bridge and 1.0 km E of Cape Terrace Road, Big Hohonu River, Westland, Waipipian; one paratype (TM8607) from GS11557, J32/f9809, re-collection of GS3046, Kumara-Inchbonnie Road, Westland, Waipipian.
Other material examined
Waipipian: GS2875, J33/f7075, top of Eight Mile Formation, Greeks Creek, Arahura valley (two incomplete); GS12354, J33/f063, locality as holotype, from soft sandstone underlying conglomerate (three incomplete); GS3049, J32/f9149, roadside E of former Westbrook Hotel, Kumara-Inchbonnie Road, Westland, 2.8 km W of Three Mile Creek bridge and 1.0 km E of Cape Terrace Road, overlying GS3046 (see above) (one fragmentary); GS12368, J32/f0167, third of three outcrops on Kumara-Inchbonnie Road near former Westbrook Hotel, 2.8 km W of Three Mile Creek bridge and 1.0 km E of Cape Terrace Road, Westland (one incomplete). A few specimens from Hawke's Bay Pliocene rocks apparently are conspecific: GS8164, V20/f8608, Napier-Taupo Highway 1.6 km S of Te Pohue (one incomplete). Almond (Citation1980:198, pl. 6, , 2) also recorded and illustrated this species from eight localities in Eight Mile Formation in the Kaniere-Arahura area, E of Hokitika in Westland (J33/f13, head of Foley Creek, off Blue Spur Road, between Arahura and Kaniere valleys; J33/f36A, lower part of E branch, Greek's Creek, Arahura; J33/f40, f41A, f42, f43, all from head of E branch, Greeks Creek; J33/f48, halfway up W branch, Greek's Creek; J33/f51A, small creek between Greeks Creek and Humphreys Gully, S side Arahura valley; material in Geology Department, University of Otago). Early Mangapanian?: GS12512, V19/f014, Waihua Valley, Wairoa (one small, with slightly narrower and more prominent spiral cords than all other specimens referred here).
Distribution
Pelicaria arahura n. sp. is widespread and not uncommon in Waipipian rocks of the Hokitika district, Westland, and rare in Waipipian and possibly early Mangapanian rocks of northern Hawke's Bay.
Dimensions
Holotype: H 41.4, D 28.0 mm; paratype TM8606: H 43.8, D 29.1 mm; paratype TM8607: H 41.7, D 30.3 mm.
Description
Shell characteristic of Pelicaria in overall appearance and aperture shape and thickening; moderate-sized for genus. Protoconch not seen. Teleoconch of 5.5–6.5 whorls. Spire moderately short for genus; last whorl occupying 2/3 to 3/5 of height. Suture deeply and widely channelled; outer border similar in width to major spiral cords lower on whorl sides. Outline below sutural channel with weakly defined, narrow, outward-sloping, weakly concave sutural ramp, bordered below by obvious peripheral angulation formed by single prominent spiral cord; peribasal angulation defined clearly by single prominent spiral cord in all specimens; vertical area between two angulations weakly concave on specimens with weak median spiral cord, almost straight and sloping slightly inward anteriorly in specimens with prominent median spiral cord (most specimens, including holotype). Spiral sculpture of few, prominent, wide, convex-crested spiral cords, with interspaces each equal in width to one cord; two prominent cords on spire whorls, three on main, vertical side of last whorl (the central one varying in prominence on different specimens, prominent on many but weakly developed on a few specimens), one prominent cord on base below peribasal angulation, followed below by 3–4 narrower cords decreasing in size anteriorly. Major spiral cords, particularly at peripheral angulation, bearing low, gently rounded, widely spaced nodules on most specimens; peribasal angulation bearing smaller, more numerous, more closely spaced nodules on some specimens. Spiral sulcus above peribasal cord well-developed on all specimens, forming posterior area of sutural channel on spire. Exterior of well-preserved specimens closely covered with very fine spiral threads. Aperture large, relatively wide as in all Pelicaria species, with relatively narrow, evenly rounded terminal varix and smoothly thickened, narrow inner lip, similar in width to outer lip; outer lip sinuous as in all Pelicaria species, shallowly deflected abaperturally below suture, followed by obvious, evenly convex but shallow adapertural flexure at mid-lip, followed again below by shallow abapertural flexure leaving evenly concave anterior sinus below junction with columella. Inner lip consistently narrow, not spreading over previous whorl; with slight parietal thickening in most specimens (including holotype) producing shallow, weakly defined posterior sinus.
Remarks
Specimens of Pelicaria arahura n. sp. from Waipipian localities in Westland and northern Hawke's Bay are similar to Pelicaria granttaylori n. sp. in many characters. However, as noted above, they differ in their consistently slightly smaller size, their shorter spire, and their much more consistent prominence and arrangement of the spiral cords, all lacking the narrow intermediate cords that are present on almost all specimens of P. granttaylori n. sp. P. arahura n. sp. is similar to P. zelandiae in consistently having a spiral depression between the median and lowest main spiral cords on the last whorl, although less obvious than that of P. zelandiae, and in many specimens having a weakly developed, inward-sloping sutural ramp above the uppermost main spiral cords, but it differs obviously from both P. zelandiae and P. granttaylori n. sp. in its much fewer, wider, more prominent spiral cords. Differences from other species are as for P. granttaylori n. sp. P. arahura n. sp. is relatively unvaried, certainly much less so than either P. granttaylori n. sp. or P. vermis, implying that it had good genetic exchange between distant populations, as it retained the brief demersal larval life of earlier Pelicaria species and Struthiolaria papulosa.
Etymology
Name based on the Arahura River, Westland, near the type locality of the species; a noun in apposition.
Pelicaria vermis (Martyn, 1784) (figs. 10C,E,F,H, A–H, A–E)
Fig. 11 Pelicaria vermis (Martyn). (A) holotype of Struthiolaria convexa Marwick, TM5763, GS1089, V21/f8482, Olrig Station, Okauawa Stream, Kereru Road, central Hawke's Bay, late Nukumaruan; height 42.9 mm. (B) “vermis/convexa form”, GS13709, S22/f023, Kupe Formation (Castlecliffian, OIS 17), Whangaehu Valley Road, E of Wanganui; height 51.1 mm. (C) holotype of Pelicaria wellmani Neef, TM4888, T24/f8022, Marima Sandstone (early Nukumaruan), Mangahao River upstream from Mangahao bridge, N Wairarapa; height 40.3 mm. (D) holotype of Struthiolaria media Marwick, TM7884, GS81, U26/f6446, Castlepoint, E Wairarapa, early Nukumaruan; height 35.8 mm. (E) nodulose Tainui Shellbed form, GS4022, R22/f6360, Tainui Shellbed (Castlecliffian, OIS 13), head of “the pinnacles” gully, Castlecliff; height 52.7 mm. (F) RM5559, Recent, deep-water “tricarinata form”, trawled off Castlepoint, E Wairarapa, 300-350 m; height 45.8 mm. (G) holotype of Pelicaria rotunda Vella, TM8587, S27/f7519, Whakarua Stream, Whangaehu River, S Wairarapa, early Nukumaruan; height 44.1 mm. (H) holotype of Pelicaria mangaoparia Vella, TM8584, GS2626, S27/f8465, Mangaopari Mudstone (early Nukumaruan), Mangaopari Stream, upstream from junction with Makara River, S Wairarapa; height 35.9 mm.

Fig. 12 (A–E) Pelicaria vermis (Martyn), Recent; A, RM4425, Te Arai Beach, Northland, height 51.1 mm; B, RM5302, “grahami form” with many weak spiral cords, trawled, 18-37 m, off East Cape; height 47.5 mm; C, RM5876, typical Wellington W coast “powelli form”, Waikanae Beach; height 51.6 mm; D, E, RM3588, 2 specimens, Tokerau Beach, Doubtless Bay, Northland; D, height 40.3 mm; E, height 45.5 mm. (F,G) Pelicaria monilifera (Suter), holotype, TM8699, “Awatere” (wrong), locality unknown, North Canterbury?, Opoitian-Waipipian?; height 48.3 mm.

Buccinum vermis Martyn 1784, vol. 2, pl. 53 (available, ICZN Opinion 479 1957).
Murex australis Gmelin 1791:3542; Dillwyn 1817:712; Martens Citation1872:20.
Struthiolaria inermis GB Sowerby I Citation1821, pl. 222, , 4; GB Sowerby II Citation1842:23, pl. 5, , 13, 19; Hutton 1880:68.
Struthiolaria crenulata Lamarck 1822b:148; Gray 1835:308; Quoy & Gaimard 1835:430, pl. 31, –9; Kiener Citation1838–1839a:5, pl. 2, , 3a; Reeve 1842 (in Citation1841–1842):201, pl. 245, , 4 (as S. inermis in plate caption; repeat of G. B. Sowerby I, 1821, pl. 222, , 4); ME Gray 1842 (in 1842–1859), pl. 5, ; 1850 (in 1842–1859), pl. 91, ; Deshayes 1843:535; JE Gray 1850:2,19,66; Paulucci Citation1877:225.
Struthiolaria tricarinata Lesson 1841:256; Hutton 1880:68; Suter in Hutton Citation1904:75; Marwick Citation1924a:186, pl. 15, , 7.
Struthiolaria australis. Reeve 1842:200, pl. 245, , 4; Reeve 1851, pl. 1, , 2; Martens 187326.
Struthiolaria scutulata Gray 1843:232; Hutton 1873a:24; Hutton Citation1873b:10 (incorrect use of the name Tylospira scutulata (Gmelin, 1791), Australian).
Struthiolaria vermis. Deshayes 1843:535, footnote; H Adams & A Adams 1854:282, pl. 27, ; Chenu Citation1859:263, fig. 1653; Tryon Citation1885:133, pl. 12, Fig. 35, 36; Hutton 1873a:24; Hutton Citation1873b:10; Paulucci Citation1877:229; Hutton Citation1893:61; Harris 1897:219; Suter in Hutton Citation1904:75; Cossmann Citation1904, pl. 8, ; Moss Citation1908:20, pl. 4, ; Suter Citation1913:276, pl. 40, ; Marwick Citation1924a:187, pl. 15, , 2; Bucknill Citation1924:44, pl. 4, ; Dell Citation1955:21, Fig. 32.
Struthiolaria vernis (sic). JE Gray Citation1850:66.
Pelicaria vernis (sic). JE Gray 1857:77.
Struthiolaria sulcata Hector 1886:50, (in part; junior primary homonym of S. sulcata Jonas, 1829).
Struthiolaria vermis tricarinata. Suter Citation1913:276.
Struthiolaria acuminata Marwick Citation1924a:185, pl. 15, , 12 (new synonym).
Struthiolaria media Marwick Citation1924a:187, pl. 15, , 14 (new synonym).
Struthiolaria convexa Marwick Citation1924a:188, pl. 15, , 4 (new synonym).
Struthiolaria fossa Marwick, Citation1924a:189, pl. 15, , 9 (new synonym).
Struthiolaria vermis var. tricarinata. Bucknill Citation1924:45, pl. 4, .
Pelicaria vermis. Finlay Citation1926b:391; Morton Citation1950:456, –6, 8; Penniket & Moon Citation1970:28, pl. 11, ; Morley Citation2004:92, upper fig.; Maxwell Citation2009:241.
Pelicaria acuminata. King Citation1933:340, pl. 39, Fig. 28; Vella Citation1953:44, pl. 4, ; Beu & Maxwell Citation1990:320, pl. 42e; Beu Citation1995:131, fig. 68i,j; Maxwell Citation2009:241.
Struthiolaria (Pelicaria) vermis. Powell Citation1937:73, pl. 14, Fig. 25; Powell Citation1946a:75, pl. 14, Fig. 25; Powell Citation1958a:100, pl. 14, Fig. 25; Powell Citation1962:92, pl. 14, Fig. 25; Morton & Miller Citation1968:491, fig. 182.4; p. 587, fig. 219.5.
Struthiolaria (Pelicaria) tricarinata. Powell Citation1937:73; Powell Citation1946a:75; Powell Citation1958a:100; Powell Citation1962:92.
Pelicaria mangaoparia Vella Citation1953:40, pl. 4, –10 (new synonym).
Pelicaria rotunda Vella Citation1953:41, pl. 4, , 7 (new synonym).
Pelicaria media. Vella Citation1953:44, pl. 4, .
Pelicaria aff. Media. Vella Citation1953:45, pl. 4, –3.
Pelicaria convexa. Vella Citation1953:45; Beu & Maxwell Citation1990:320, pl. 42b; Beu Citation1995:131, fig. 68a–h,k,l; Maxwell Citation2009:241.
Struthiolaria (Pelicaria) convexa fossa Beu Citation1965:143, pl. 2, figs. 15–17.
Struthiolaria (Pelicaria) acuminata. Fleming Citation1966:52, pl. 89, figs. 1054, 1056.
Struthiolaria (Pelicaria) convexa. Fleming Citation1966:52, pl. 90, figs. 1063, 1064.
Struthiolaria (Pelicaria) fossa. Fleming Citation1966:52, pl. 90, figs. 1061, 1062.
Struthiolaria (Pelicaria) mangaoparia Fleming Citation1966:52.
Struthiolaria (Pelicaria) media. Fleming Citation1966:52, pl. 90, figs. 1059, 1060.
Struthiolaria (Pelicaria) rotunda. Fleming Citation1966:52.
Struthiolaria (Pelicaria) vermis vermis. Fleming Citation1966:52, pl. 90, figs. 1067, 1068; Neef Citation1970:459, figs. 10, 11, 24, 24a; Powell Citation1976b:94, pl. 21, fig. 25; Powell Citation1979:143, pl. 7, fig. 4; pl. 30, ; Spencer & Willan Citation1996:21.
Struthiolaria (Pelicaria) vermis tricarinata. Fleming Citation1966:52, pl. 90, figs. 1065, 1066.
Struthiolaria (Pelicaria) vermis flemingi Neef Citation1970:462, , 13; Powell Citation1976b:94; Powell Citation1979:143, pl. 30, ; Spencer & Willan Citation1996:21 (new synonym).
Struthiolaria (Pelicaria) vermis convexa. Neef Citation1970:463.
Struthiolaria (Pelicaria) vermis cf. Convexa. Neef Citation1970:463, , 17.
Struthiolaria (Pelicaria) vermis grahami Neef Citation1970:465, , 21; Spencer & Willan Citation1996:21 (new synonym).
Struthiolaria (Pelicaria) vermis powelli Neef Citation1970:465, , 15; Spencer & Willan Citation1996:21 (new synonym).
Struthiolaria (Pelicaria) vermis bradleyi Neef Citation1970:467, , 19; Spencer & Willan Citation1996:21 (new synonym).
Struthiolaria (Pelicaria) vermis vellai Neef Citation1970:468, , 23 (new synonym).
Struthiolaria (Pelicaria) wellmani Neef Citation1970:472, Fig. 35, 36 (new synonym).
Pelicaria fossa. Beu & Maxwell Citation1990:321,407, pl. 42a; Maxwell Citation2009:241.
Pelicaria vermis vermis. Beu & Maxwell Citation1990:351,407, pl. 47u; Spencer et al. Citation2009:207.
Pelicaria vermis flemingi Beu & Maxwell Citation1990:407; Spencer et al. Citation2009:207.
Pelicaria vermis grahami. Beu & Maxwell Citation1990:407; Spencer et al. Citation2009:207.
Pelicaria vermis bradleyi. Spencer et al. Citation2009:207.
Pelicaria vermis powelli. Spencer et al. Citation2009:207.
Pelicaria flemingi. Maxwell Citation2009:241.
Type material
As with all of Martyn's illustrated specimens, the type material of Buccinum vermis Martyn has long been lost. Neef (Citation1970, Fig. 24, 24a) repeated Martyn's original illustrations, which shows the common form of Pelicaria vermis (Martyn, 1784) occurring around Auckland and in Northland, with relatively prominent nodules around the periphery. Neef (Citation1970:462) thought a locality near Queen Charlotte Sound likely for the holotype, because the type originated from Cook's voyages to New Zealand, and this was the place where Cook stayed longest and most frequently. However, Pelicaria vermis is rare in or, in most areas, absent from the Marlborough sounds, the illustrated shell is clearly an Auckland or Northland specimen, and a type locality in the Bay of Islands is much more likely.
Gmelin (1791:3542) cited as illustrations for Murex australis only Spengler's (Citation1782) drawings in ‘Spengl. Naturf. 17. t. 2. f. C. D.’ (see below), so Spengler's specimen would be the holotype of Murex australis if it still existed; unfortunately, it does not. A complication with this name is the usage of the same binomen for the large conoidean species now consistently known as Nihonia australis (e.g., Powell 1969:331, pl. 192, , 17; pl. 254, , 2). Tucker (Citation2004:101) attributed this name to Gmelin (1791:3542), stating incorrectly that Gmelin referred only to Chemnitz (Citation1795, pl. 190, fig. 1827, 1828). Obviously it is impossible for Gmelin (1791) to have referred to a work published in 1795, so it not surprising that Powell (1969:331) attributed this name to a later author, Roissy (Citation1805:72), where it was published as Pleurotoma australis. Roissy in turn referred to Chemnitz's (1795, pl. 190, figs. 1827, 1828) figures, among others. Powell (1969:331) made the situation clear by stating under Nihonia australis: ‘This is not Murex australis Gmelin, 1791, which refers to the New Zealand species Struthiolaria (Pelicaria) vermis (Martyn, 1784)’. The misunderstanding over this name probably arose because Roissy (1805:72) incorrectly included a reference to ‘Spengler, Naturf. 17, tab. 2, fig. C, D’ (= Pelicaria vermis) in the synonymy of Pleurotoma australis. There is no doubt that Gmelin (1791:3542) was referring to Pelicaria vermis, as his remarks include ‘Habitat in Oceano australi, stramineo affinis, testa quoque straminea … ’; Murex stramineus Gmelin, 1791 (= Struthiolaria papulosa (Martyn, 1784)) is the preceding species on the same page.
Struthiolaria inermis G. B. Sowerby I, 1821, no type material present in BMNH, and no material in the general collection is identified as S. inermis (K Way, BMNH pers. comm. 14 January 2008); location of type material unknown. This name has usually been credited to GB Sowerby II (1842), but it was referred to by Lesson (1841), and both Deshayes (1843) and Paulucci (1877) pointed out its earlier publication by GB Sowerby I (1821). None of Lesson's type material of Struthiolaria tricarinata is present in MNHN and its location is not known, although a photograph in GNS, sent from MNHN during the 1960s when Neef (Citation1970) was carrying out his research, purports to be of ‘the type’, and is labelled on the back ‘type de Struthiolaria tricarinata Lesson, Muséum Paris’. Lesson (1841:256) stated that it was obtained by the frigate ‘la Vénus’ (not, as was assumed by Neef [Citation1970]:468], Bay of Islands material collected by the ‘Coquille’), Lesson had seen ‘about 20 perfectly similar’ specimens from New Zealand (that is, there were originally ca 20 syntypes), ‘but it appears to be found solely on the beaches of the South Island [l’île Méridionale]’. The described specimen was 22 mm high and 14 mm wide, unusually small for an adult specimen of P. vermis. The locality, the size, and Lesson's description of the suture as ‘simple et peu profonde’ suggest that he was describing the ‘bradleyi form’ with a narrow sutural channel, named by Neef (Citation1970:467, , 19) from beaches near Nelson, northern South Island—common on Rabbit Island Beach after storms (C), but also seen from Marahau, Kaiteriteri Beach, and Golden Bay. No other form around New Zealand at present has a deep sutural channel, and Pelicaria is not cast ashore on any other beaches in the South Island. Specimens from Mahurangi estuary, southern Northland (GNS RM4420, many, collected by JR Penniket) are very similar to the ‘bradleyi form’ in their short, squat shape, with an obvious peripheral angulation and a prominent, flat, outward-inclined sutural ramp, but have a much shallower, narrower sutural channel than N South Island specimens. The specimen of P. vermis illustrated under the name S. crenulata by Kiener (1838–1839a, pl. 2, ) was also said to ‘Habite les plages de la nouvelle baie Tasman, dans le détroit de Cook, à la Nouvelle-Zélande’ [found on the beaches of the new Tasman Bay, in Cook Strait, at New Zealand], and clearly has a narrow sutural channel, and so is a specimen of the same form. The ‘variety’ illustrated by Kiener (1838–1839a, pl. 2, a) is shorter, lacks a sutural channel and has nodules around the periphery, and appears to be an Auckland-Northland specimen. Lesson (1841) discussed a species Struthiolaria oblita of Kiener, in relation to S. crenulata, that is, Pelicaria vermis, but this name does not appear in Kiener (1838–1839a). I conclude that no type material exists for any of the four early names Buccinum vermis, Murex australis, Struthiolaria inermis or Struthiolaria tricarinata, and at least two names listed by early cataloguers are nomina nuda (S. oblita ‘Kiener’ Lesson, Citation1841; S. canaliculata ‘Spengl.’ H Adams & A Adams 1854:282).
Struthiolaria crenulata, one syntype MNHN moll. 20522, H 40.8 mm, without locality, stated by Lamarck to be from ‘Nouvelle Zélande’, collected by Péron & Lesueur in 1803; labelled ‘Individu nommé par Lamk’, although it is considered to be a syntype under ICZN Recommendation 73F, as Lamarck (1822b:148) stated merely ‘coll. du Muséum’, without stating how many specimens he had before him. Photographs (H) kindly supplied by Virginie Héros (Malacologie, MNHN) show that this, too, possibly is a specimen of Pelicaria vermis of the ‘bradleyi form’, from northern beaches of the South Island, with a steep but clearly defined sutural ramp and an obvious but shallow sutural channel, although this specimen has a less prominently protruding peripheral cord than most other specimens of the ‘bradleyi form’, and is not the specimen of which photographs were supplied earlier as ‘the type’ of S. tricarinata Lesson, which is smaller and has a more prominent, nodulose peripheral cord. It might also possibly be from the beaches of western Wellington (Paekakariki to Otaki), where the taller form with a less obviously channelled suture (C) is cast ashore that was named S. vermis powelli by Neef (Citation1970:465, , 15). This form appears to be intermediate between the ‘bradleyi form’ and forms in the northern North Island. These intermediates and the Mahurangi estuary intermediates in Northland show that there is little justification for treating these forms as geographical subspecies. The syntype, MNHN moll. 20522, is here designated the lectotype of Struthiolaria crenulata Lamarck, 1822, in case other syntypes should come to light and are not conspecific with the remaining syntype. In order to attach all other early names permanently and unambiguously to this species rather than to S. papulosa (Martyn, 1784), the lectotype of Struthiolaria crenulata Lamarck, MNHN moll. 20522, is here designated also the neotype of Buccinum vermis Martyn, 1784, the neotype of Murex australis Gmelin, 1791, the neotype of Struthiolaria inermis G. B. Sowerby I, 1821, and the neotype of Struthiolaria tricarinata Lesson, Citation1841. These species were all stated to come from ‘New Zealand’, the locality of the neotype, and use of this specimen as the neotype renders all these names objective synonyms of Pelicaria vermis (Martyn, 1784).
Struthiolaria acuminata holotype TM7682 (F), GS1040, S27/f8448, sandstone member of Pukenui Limestone, Te Awaite cutting, Awhea Road, 8 km SE of Martinborough, S Wairarapa, with two paratypes, TM7683-4; early Nukumaruan, occurring with Zygochlamys delicatula. (The early locality ‘Te Awaite cutting’ [also written ‘Twaite's cutting’] was well-known; it was on an outer curve of White Rock [or Awhea, Te Awaite] Road overlooking Makara River about 1.3 km SW of the present ‘banana bridge’ over Huangarua River and 1.5 km N of the junction of Makara River and Mangaopari Stream, but was by-passed by road straightening many years ago.) Struthiolaria media holotype TM7884 (D), GS81, U26/f6446, Castlepoint, E Wairarapa; early Nukumaruan, with Z. delicatula. Struthiolaria convexa holotype TM5736 (A), GS1089, V21/f8482, Okauawa Formation (late Nukumaruan) (Erdman & Kelsey 1992), Okauawa Stream, Kereru Road, inland central Hawke's Bay. Struthiolaria fossa holotype TM8583, GS191, V21/f8463, ‘Shrimpton's’, that is, Kikowhero Stream, Matapiro Station, N of Ngaruroro River, central Hawke's Bay; late Nukumaruan, near the axis of Matapiro syncline. Pelicaria mangaoparia holotype TM8584 (H), V300,=GS2626, S27/f8465, with two paratypes, TM8585-6, Mangaopari Stream, 1.2 km upstream from Makara River, Awhea Road, SE of Martinborough, S Wairarapa; early Nukumaruan, underlying Z. delicatula (which, however, appears late in this succession because of the deep-water facies of the lower part of the section; Gammon Citation1997; Orpin et al. Citation1998, ); other paratypes from V34, V301. Pelicaria rotunda holotype TM8587 (G), V205, S27/f7519, junction Whakarua Stream and first tributary from S, upstream from junction of Whakarua Stream and Whangaehu River, S Wairarapa, early Nukumaruan; with paratypes from V7, V25, V59, V204, V216, V272, V274 and V278 (localities and fossil record numbers listed by Vella Citation1953:46–47); one paratype TM8588, GS5452, T27/f6529, Oreka Stream, W slope of Maungaraki Range, E of junction of Millars Road with Martinborough-Gladstone Road, S Wairarapa; early Nukumaruan. Pelicaria wellmani holotype TM4888 (C), with one figured paratype TM4889, G Neef's locality 1022, T24/f8022, listed by Neef (Citation1967, appendix 1) as Marima Sandstone, with three persistent beds of concretions, left bank Mangahao River 350 m NE of bridge at Marima, N Wairarapa, grid ref. T24/401743; age listed by Neef (Citation1970:472) as ‘lower Hautawan’, that is, early Nukumaruan.
Other material examined
Not listed; Pelicaria vermis is widespread and abundant in early Nukumaruan–Haweran rocks throughout most of New Zealand.
Distribution
Pelicaria vermis is interpreted here as a single highly variable species (as a result of its direct development) occurring widely in earliest Nukumaruan to Haweran rocks and in the Recent fauna. The many named forms all intergrade, and any evolutionary change during the time range of P. vermis apparently was anagenetic. In the Recent fauna it occurs in shallow water (allowing living specimens to be cast ashore after storms) throughout the eastern North Island, rarely on Ninety Mile Beach, NW Northland, and on both sides of Cook Strait, and occurs also on the continental shelf in slightly deeper water (ca 20–80 m) along the east coast of both islands from East Cape to Otago Peninsula (Graham Citation1962:57; dead specimens rare in Graham's zone 2, 9–55 m). A few specimens have also been collected in much deeper water off eastern New Zealand (e.g. RM5559, 300–350 m, trawled off Castlepoint, E Wairarapa; F) and some of them have relatively prominent spiral cords similar to the Nukumaruan P. acuminata form (also known as P. tricarinata). It is absent from the rest of New Zealand.
Dimensions
Lectotype of Struthiolaria crenulata Lamarck and neotype of Buccinum vermis Martyn, Murex australis Gmelin, Struthiolaria inermis G. B. Sowerby I and Struthiolaria tricarinata Lesson: H 40.8 mm, D 26.8 mm; holotype of P. acuminata: H 42.9, D 26.7 mm; holotype of P. media: H 35.8, D 23.6 mm; holotype of P. convexa: H 42.9, D 28.8 mm; holotype of P. rotunda: H 44.1, D 30.0 mm; paratype, GS5452: H 47.5, D 32.5 mm; holotype of P. mangaoparia: H 35.9, D 23.2 mm; holotype of P. wellmani: H 40.3, D 26.5 mm; large, elongate specimens of acuminata form, GS3111, T26/f6462, Taueru bridge, N of Masterton, Wairarapa: H 57.5, D 34.5 mm; H 53.5, D 33.6 mm; acuminata form, GS2611, S27/f8450, Makara River, 800 m upstream from Ruakokopatuna River junction, S Wairarapa: H 53.1, D 31.0 mm; convexa form, GS10748, hillside above Makara River–Mangaopari Stream junction: H 48.0, D 32.4 mm; large specimen of convexa form, Waikopiro, S Hawke's Bay, Suter collection: H 56.0, D 38.2 mm; large Recent specimens: RM5244, beach, N side Ayres Point, Te Arai Beach, N of Leigh, Northland: H 56.5, D 35.1 mm; RM5302, trawled in 18–37 m, off East Cape: H 55.4, D 35.2 mm; short ‘bradleyi form’, RM2974, Rabbit Island Beach, Nelson: H 36.6, D 28.3 mm.
Remarks
A few authors (Gmelin 1791:3542; Deshayes 1843:535; Martens Citation1872:20; 1873:26; Paulucci 1877) have recognized that Pelicaria vermis was first illustrated by Spengler (1782:31, pl. 2, figs. C, D), who, however, did not propose a name for this species. As mentioned by Iredale (Citation1956:83), Spengler was among the first to illustrate and describe shells brought to Europe by Cook. Spengler (1782) described and illustrated both Recent Struthiolaria species, using only the English vernacular names proposed by George Humphrey. The year 1782 is well before any publications by Humphrey, so the names were presumably suggested orally. Spengler stated that he obtained his specimens through Humphrey, who obtained them in turn from Sir Joseph Banks and Daniel Solander; that is, as seems necessary this early, they were collected during Captain James Cook's first visit to New Zealand. H Adams & A Adams (1854:282) listed Struthiolaria canaliculata ‘Spengl.’ in their catalogue of Struthiolaria species, but this name was not proposed in Spengler's (1782) paper, and was not listed by Sherborn (Citation1902). It seems to be a nomen nudum appearing only in H Adams & A Adams’ list. If this name were available, it would be a senior primary homonym of the well-known New Zealand fossil species Pelicaria canaliculata (Zittel) (1864:34, pl. 15, ), originally proposed in Struthiolaria; the varietal name S. canaliculata becki proposed by Neef (Citation1970:469, Fig. 25, 29) would take its place if Zittel's name is a junior homonym. However, P. canaliculata (Zittel) appears to be valid. As noted above, Spengler's illustrated specimen is also the holotype of Murex australis Gmelin (1791:3542). Kathe Jensen (ZMC pers. comm. August 2006) reported that no Pelicaria material illustrated by Spengler (1782) remains in Spengler's collection in ZMC. Pelicaria vermis is one of several New Zealand species named in Martyn's non-binominal work that were conserved in ICZN Opinion 479 (1957).
The great range of variation of forms of what is here identified as the single species Pelicaria vermis has long troubled paleontologists and neontologists alike, and a certain amount of ‘art’ has always been involved in distinguishing the intergrading forms traditionally recognized as fossil species. I previously (Beu Citation1995:119–121, fig. 68) discussed their occurrence in central Hawke's Bay, where they at first seemed to offer a similar biostratigraphically useful zonation to that proposed by Vella (Citation1953) in SE Wairarapa. However, I concluded that their arbitrary-looking occurrences resulted from ecological differences between the deposition sites, combined with a temperature control on the prominence of spiral cords, rather than being an evolutionary series. Local populations greatly dominated by the P. acuminata form, with a relatively tall (but highly variable) spire and very prominent spiral cords, are encountered in S Wairarapa and at Castlepoint. Only rare specimens of the P. convexa form, with a slightly shorter spire and no prominent spiral cords at all, have been found in S Wairarapa. I am aware of only two: one incomplete specimen was reported by Vella (Citation1953:45) from V311 (S27/f7520, sandstone downstream from junction Ruakokopatuna and Makara Rivers [that is, Hautotara Formation], SE of Martinborough, late Nukumaruan) and I collected one specimen (E; GS10748, S28/f6206, sandstone beneath Pukenui Limestone member B, above White Rock Road at the junction of Mangaopari Stream and Makara River, SE of Martinborough; early Nukumaruan, with Zygochlamys delicatula). In contrast, local populations dominated by the P. convexa form, and only uncommonly and early in the succession varying towards the P. acuminata form (Beu Citation1995, fig. 68c–f,i), occur throughout central and southern Hawke's Bay, as far south as the Totara Road area, near Kumeroa, E of Woodville. Unfortunately, this species group is not known from the geographically intermediate area of central and northern Wairarapa. Suitable facies in this area apparently do not extend this young. I (Beu Citation1965, Citation1995:121) described a ‘pseudoevolutionary’ succession of forms intergrading between P. convexa and P. fossa in the Devil's Elbow hill section, on Highway 2 N of Napier, in which the degree of expression of the sutural channel characteristic of the P. fossa form increases in succeeding siltstone formations up-section. However, alternating sandier units (with P. convexa and common Tawera) and muddier units (with P. fossa and more offshore taxa such as Amalda mucronata (G. B. Sowerby I, 1830)) in the succession exposed along Kereru Road, inland central Hawke's Bay, show that the P. fossa form occurred in more offshore units contemporaneously with the shallower-water P. convexa form. The Devil's Elbow succession evidently reflects water depth increasing gradually up-section during the deposition of siltstone units, rather than the gradual evolution of a distinct species. Despite deposition being affected by rhythmical glacio-eustatic sea-level change, the entire basin sank more rapidly than accommodation was filled with sediment. This confusion between evolutionary and ecophenotypic change is frequent and normal in fossils of the Pelicaria vermis species complex, and describes the succession in S Wairarapa (Vella Citation1953) as well as in Hawke's Bay. Neef (Citation1970) also described a number of local variants (interpreted by him as geographical subspecies) of P. vermis living at present in areas around the North Island, northern South Island, and off E Canterbury–N Otago, demonstrating that a similar range of ecological and geographical variation occurs in the Recent fauna.
Morton (Citation1950) described the development of Pelicaria vermis, in which ‘the embryos are completely incubated within the [brood] pouch, and are liberated as tiny replicas of the adult which at once take on a benthic existence’, an excellent example of direct development. In a popular account (Morton Citation1997a, Citationb) he compared the anatomy, ecology, life history and phylogeny of Pelicaria with Struthiolaria, Perissodonta, Tylospira, Aporrhais and other stromboideans, emphasizing the different developmental modes of Struthiolaria papulosa and Pelicaria vermis. Direct development and the consequent lack of genetic exchange between local populations by a significant, although relatively brief, demersal larval stage (as occurs in Struthiolaria papulosa) explains the maintenance of distinct forms in discrete areas by P. vermis, as well as the intergradation of the long series of subtly different forms of P. vermis recognized previously as species and subspecies by paleontologists. In contrast, S. papulosa exhibits simple clinal variation in spire height and peripheral nodule size over New Zealand, with smaller, shorter, prominently nodulose specimens in the far north of the North Island, much larger, taller, narrower specimens lacking nodules altogether at Stewart Island, and a complete morphological and geographical intregradation between these extremes. Pelicaria vermis is therefore interpreted here as a classic example of a single long-ranging, anagenetically changing, direct-developing species, highly plastic in its phenotypic expression. Although forms within it might be useful for biostratigraphy in local areas (one sub-basin or smaller area) over short time scales, my experience in Hawke's Bay and SE Wairarapa indicates that the ecological and temperature controls on the variation of P. vermis have produced a fossil record that confuses rather than aids the biostratigrapher. It has also always been impossible to distinguish some Castlecliffian forms that traditionally would have been identified as P. vermis (e.g. in Kupe Formation in Wanganui Basin; E) from the Nukumaruan form in Hawke's Bay traditionally identified as P. convexa. In my opinion this is one variable species, and it is time to study its interesting genetics and ecology rather than attempt to force it into apparent biostratigraphical utility that produces conflicting or simply incorrect results.
Neef (Citation1970) described an evolutionary series preserved in Waipipian–Mangapanian rocks in the Marima area, SW of Woodville in N Wairarapa, in which Pelicaria vermis (as interpreted broadly here) evolved gradually from P. canaliculata in a series of forms with progressively lower spiral cords with progressively less strongly undercut edges (Neef Citation1970, ; ‘period of co-existence’, ‘clarki probably grades into marima’, etc.). This, then, poses the question of where the boundary should be drawn of what is included in P. vermis. Neef's (1970) study is detailed enough to show that the succession of forms at Marima represents an evolving unidirectional series of species rather than the arbitrary-looking series of random walks (ecophenotypic adaptation to local environmental variation) adopted in different geographical areas by P. vermis. The early Nukumaruan form named P. wellmani by Neef (Citation1970:472, Fig. 35, 36) is closely similar to specimens identified as P. acuminata elsewhere in Wairarapa and Hawke's Bay, and is assumed here to represent the first specimens of the direct-developing species P. vermis. As the enormous range of variation of P. vermis commences with the P. acuminata form, and populations of what Neef (Citation1970) called P. canaliculata, P. clarki and P. marima are remarkably unvaried (even though apparently evolving gradually, that is, anagenetically), it seems likely that a demersal larval stage was abandoned and direct development adopted on evolving the P. acuminata form, at the beginning of Nukumaruan time. The forms intermediate between typical, early P. canaliculata (Waipipian) and P. acuminata deserve further study, and perhaps would be best recognized as the single species P. canaliculata, but are here segregated from P. vermis, which is limited here to the direct-developing, final species of the genus.
A further form that seems to fall within the range of variation of Pelicaria vermis is early Nukumaruan specimens in GNS identified as related to P. lacera (Marwick Citation1931:96, fig. 133, 134). This species with a few prominent, very narrow, widely spaced spiral carinae was named from poor, distorted, mouldic material from GS1244 (Z17/f6464, Mangatuna Quarry, main highway N of Tolaga Bay, N of Gisborne; Kapitean, late Miocene) and represents a small, wide, very early, latest Miocene species that is almost unknown elsewhere. The large early Nukumaruan specimens with narrow cords that have been thought related to P. lacera are essentially tall-spired specimens of the acuminata form with unusually narrow, widely spaced spiral cords, and are included here in P. vermis: GS2481, T24/f6408, Lower Kumeroa Formation, Tahoraite SD, Dannevirke, one large, incomplete; GS2651, T24/f7389, all data as last, one; GS11959, T24/f8797, gully 900 m ESE of Pahiatua trig. station, N Wairarapa, G Neef, two; GS11991, T24/f8051, left bank Mangahao River, N Wairarapa, G Neef, one incomplete; TM4896, T24/f8041, Neef's locality 1041, slip W side of Mangaramarama Stream, 2 km E of Totaranui Station, between Konini and Pahiatua, N Wairarapa; specimen illustrated by Neef (Citation1970, Fig. 37).
Pelicaria vermis provides an example of the type of highly variable complex of forms that commenced in early Nukumaruan time, and is also exemplified by Stiracolpus symmetricus and Aoteadrillia wanganuiensis.
Pelicaria rugosa (Marwick, 1924) (g)
Struthiolaria cingulata. Hutton 1873b:11 (in part, ‘Patea’ shell only, not S. cingulata Zittel, 1864); Suter Citation1914:18, pl. 1, (not S. cingulata Zittel, 1864).
Struthiolaria rugosa. Marwick Citation1924a:189, pl. 13, .
Struthiolaria (Pelicaria) rugosa. Fleming Citation1966:52, pl. 89, fig. 1058; Neef Citation1970:472, Fig. 38.
Pelicaria rugosa Beu & Maxwell Citation1990:321, pl. 42c; Maxwell Citation2009:241.
Type material
Struthiolaria rugosa, holotype TM4898, supposedly from ‘Patea’, S Taranaki (early NZ Geological Survey type-set label glued onto specimen), but this Waipipian locality is obviously an error; early Nukumaruan, apparently from North Canterbury (Beu & Maxwell Citation1990:321), possibly from Motunau Beach. The holotype of P. rugosa is the specimen that Hutton (1873b:11) recorded as S. cingulata from ‘Patea’, and that Suter (Citation1914:18, pl. 1, ) illustrated as S. cingulata. The printed label attached to the shell reflects this early confusion; both the locality and the identification were incorrect. Suter (Citation1914:18) followed Hutton's error and described Hutton's (1873b:11) ‘var. B. – Ribs sub-moniliform’ as the subspecies S. cingulata monilifera, that is, Pelicaria monilifera (F,G). Struthiolaria cingulata is a more prominently spirally ridged, typical Struthiolaria species limited to Kapitean–Opoitian rocks of the Awatere valley.
Other material examined
Nukumaruan: GS2855, U24/f6440, Waitahora Road, Mangaotoro, S Hawke's Bay (one; Beu & Maxwell Citation1990, pl. 42c); GS2419 (U24/f6337, Mangapuku Stream, 2 km E of Waitahora Valley Road and 2.5 km S of Dannevrke-Weber Road; few); GS2472 (U24/f6368, tributary of Mangamaire Stream, 2 km NE of Dannevirke-Weber Road and 5 km SE of Mangatoro; few); GS2483 (T24/f7375, hill slope above Highway 2, 1 km W of Waiaruhe, between Woodville & Dannevirke; few); and GS2758 (U24/f6436, Mangatoro Stream, 1 km SW of Mangatoro; few), all in Lower Kumeroa Formation of Dannevirke Subdivision (Fleming & Marwick in Lillie Citation1953, table 21); G Neef's fossil locality 1049, T24/f8049, 1 km SW of Marima, N Wairarapa (30 cm-thick P. rugosa shellbed in Marima Sandstone, tributary of Mangahao River 0.5 km SE of Naenae Road, grid ref. T24/391735; Neef Citation1967, appendix 1, table following p. 359; ‘a [30 cm] shell bed largely composed of P. rugosa … is unique … . Scattered P. rugosa shells occur up to [1.6 m] above the P. rugosa bed in otherwise unfossiliferous sandstone’; Neef Citation1967:183; 1970:472, Fig. 38); GS5490, S27/f8593, low in Pukenui Limestone (presumably the same horizon as the following one), stream beside Te Muna Road, W side Huangarua River, 2.2 km upstream from Martinborough-Masterton Road, 4 km SE of Martinborough, SE Wairarapa (one); common in a thin bryozoan-rich zone in Mangaopari Stream ca 10 m upstream from Birch Hill Station access bridge, ca 200 m upstream from junction with Makara River, 100 m S of White Rock Road, S Wairarapa; GS10965, S28/f6202A and re-collections, common in sandstone with abundant Zygochlamys delicatula where Whangaimoana Stream flows into beach sands at its mouth, E end of Whangaimoana Beach, between Lake Ferry and Hurupi Stream, Palliser Bay, S Wairarapa; abundant (with Z. delicatula) in the matrix of the enormous debris flows (Lewis Citation1976:539) at Motunau Beach, North Canterbury; common (with Z. delicatula) in the Gower River and nearby areas, Cheviot district, North Canterbury.
Distribution
Pelicaria rugosa occurs only in a brief basal Nukumaruan zone in S Hawke's Bay, N and S Wairarapa, and in the Cheviot district and at Motunau Beach in North Canterbury. It provides a very useful biostratigraphical index of earliest Nukumaruan time; all these localities seem to correlate closely in age. It occurs with Zygochlamys delicatula at all localities other than in S Hawke's Bay and N Wairarapa, where the facies seems to have been too shallow for Z. delicatula to occur (Z. delicatula lives now below 70 m, and almost all records are from water deeper than 200 m). This highlights an unusual aspect of the occurrences of P. rugosa: it occurs in moderately deep-water (outer shelf to upper bathyal) facies, deeper than most or all other Pelicaria species, and I am not aware of syntopic occurrences with P. vermis. Among other things, P. rugosa is helpful for identifying the age of most of the fossil penguins in concretions at Motunau Beach, North Canterbury (Marples Citation1960; Simpson Citation1972) as early Nukumaruan. I am not aware of any authentic records of P. rugosa from Wanganui Basin, again probably because depositional facies there were too shallow.
Dimensions
Holotype: H 40.9, D 28.0 mm; GS5490, Te Muna Road, S Wairarapa: H 44.8, D 31.5 mm; GS2855, Mangaotoro, S Hawke's Bay: H 45.3, D 32.0 mm; GS10965, Whangaimoana Stream mouth, Palliser Bay: H 34.2, D 22.9 mm.
Remarks
Pelicaria rugosa is the one Pelicaria species I am aware of in post-Mangapanian rocks that is distinct from P. vermis, apart from the early Nukumaruan records (above) of P. granttaylori n. sp. P. rugosa is rendered highly distinctive by its exterior sculpture, as it lacks the prominent, wide spiral cords (often labelled cinguli in this genus) so characteristic of P. arahura n. sp., P. granttaylori n. sp., P. canaliculata, and several early forms of P. vermis. Instead it bears numerous low, narrow, closely spaced, convex-crested spiral ridges, about 10 on spire whorls and 25 on the last whorl, with interspaces each about equal in width to one cord, crossed by numerous low, narrow axial ridges with the same prominence and spacing as the spiral cords, forming small, narrowly rounded nodules at the intersections and so producing the finely nodulose surface reflected in the species name. The shape is similar to that of the convexa form of P. vermis, with no clearly defined sutural ramp, with rounded shoulders and a gradually rounded base, and the smoothly rounded lips are very similar to those of all other Pelicaria species. P. rugosa is almost completely constant in its characters, implying again that it retained a demersal larval life that allowed genetic exchange throughout its range, as in earlier Pelicaria species and Struthiolaria papulosa. A little-known, more coarsely sculptured species, P. monilifera (Suter) (Citation1914:18, pl. 1, ) (Opoitian–Waipipian, rare in N Canterbury–Marlborough; holotype F,G), is similar apart from its fewer, larger nodules (caused by its fewer, wider spiral cords) and markedly taller spire, and possibly was the ancestor of P. rugosa.
Recognized Pelicaria species
This part of the present paper essentially is a revision of the Pliocene to Recent species of Pelicaria, so a list is provided of the taxa now recognized in the genus. The status of a few named species remains unclear. ‘Pelicaria’ incrassata (Powell) (Citation1931:101, pl. 13, Fig. 30–33) (Waipipian, Tangahoe Formation, Waihi Beach, Hawera) does not belong in any obvious lineage of Pelicaria, and the late PA Maxwell suggested to me that it is a small species of Struthiolaria. The relatively tall, narrow shape, the presence of only two spiral cords bordering an otherwise wide, concave, last whorl mid-section, the small aperture, the heavily thickened, weakly sinuous lips, and the unusual, large parietal tubercle all agree with this position, and I consider S. incrassata to be a species of Struthiolaria. P. pseudovermis (Bartrum & Powell) (1928:142, pl. 30, fig. 63, 64) (Opoitian, Kaawa Creek, SW Auckland) also has some similarities with Struthiolaria, particularly the unusually deep, prominent anterior sinus in the outer lip. However, Bartrum & Powell (1928:142, pl. 30, fig. 63, 64) argued for a relationship with P. nana (Marwick, Citation1926), the earliest species of Pelicaria (Beu & Maxwell Citation1990:262, pl. 29g), and although the last whorl lacks the characteristic Pelicaria median spiral cord, it is present on spire whorls. A relationship with P. nana appears feasible, and this species is retained in Pelicaria. The status of P. clarki and P. marima, N Wairarapa species recognized by Neef (Citation1970), remains to be clarified, and they are listed here despite apparently intergrading anagenetically with P. canaliculata. So although two new species are named here, the 19 species and subspecies listed by Beu & Maxwell (Citation1990:407) are reduced to 15, and probably in the long run will be reduced to 13.
Recognized species are:
Pelicaria arahura n. sp., E, 10A,B, Waipipian (–Mangapanian?), Westland, N Hawke's Bay.
Pelicaria canaliculata (Zittel, 1864), D, Opoitian?, Waipipian, Hawke's Bay–N Canterbury; rare in Waipipi shellbeds, Waverley Beach, W of Wanganui.
Pelicaria cestata Marwick, Citation1965, Waipipian, N Hawke's Bay.
Pelicaria clarki (Neef, Citation1970), Mangapanian, N Wairarapa (probably intergrades with P. canaliculata).
Pelicaria granttaylori n. sp., A–E, Mangapanian–early Nukumaruan, Wanganui & N Hawke's Bay.
Pelicaria lacera (Marwick, Citation1931), Kapitean, Gisborne district.
Pelicaria marima (Neef, Citation1970), Mangapanian, N Wairarapa (probably intergrades with P. canaliculata).
Pelicaria monilifera (Suter, Citation1914), F,G, (Opoitian?), Waipipian, N Canterbury.
Pelicaria nana (Marwick, Citation1926), Tongaporutuan, N Taranaki and S Wairarapa.
Pelicaria parva (Suter, Citation1915), Opoitian, holotype from unknown locality (others in GNS: GS1574, W19/f7481; GS1576, W19/f7483, both Wahanui Road, N Hawke's Bay, one at each locality; GS4936, W21/f8555, between shelter huts and Black Reef, Cape Kidnappers, three; GS2230, T25/f6521, Kaitawa Road, Makuri, E of Woodville, S Hawke's Bay, one unusually tall and narrow, possibly a distinct species; GS11519, J33/f088, outcrop west of McKay Creek mouth, opposite lower power house, Lake Kaniere Road, Westland, in a diverse fauna including Veprichlamys scandula (Hutton, 1873), Mesopeplum cf. waikohuense (Marwick, Citation1931), Marama murdochi (Marwick, 1927), Zeacolpus kanieriensis (Harris, Citation1897), Galeodea flemingi Beu & Maxwell, Citation1990, Aeneator n. sp., Polinices (Polinella) obstructus (Marwick, 1924); two specimens).
Pelicaria procanalis Beu, 1970, Opoitian, Awatere valley.
Pelicaria pseudovermis (Bartrum & Powell, Citation1928), Opoitian, Kaawa Creek.
Pelicaria rugosa (Marwick, 1924), G, early Nukumaruan, Hawke's Bay–N Canterbury.
Pelicaria vermis (Martyn, 1784), C,E,F,H, 11A–H, 12A–E early Nukumaruan–Recent, widespread.
Pelicaria zelandiae (Marshall & Murdoch, 1920), A, Opoitian, N Canterbury?; Waipipian, S Taranaki–W Wanganui.
Genus Struthiolaria Lamarck, Citation1816
Struthiolaria Lamarck 1816, pl. 431, a,b; ‘Liste des objets représentés dans les planches de cette livraison’ p. 7. Type species (by monotypy): Struthiolaria nodulosa Lamarck, 1816 (= Buccinum papulosum Martyn, 1784; available, ICZN Opinion 479 1957), Pliocene–Recent, New Zealand.
Struthiolaria frazeri Hutton, 1885 (fig. 13A,C–F)
Struthiolaria frazeri (ex Hector MS) Hutton Citation1885:329; Harris 1897:220, pl. 6, a, b; Speight Citation1913:31; Suter 1921:19; Marwick Citation1924a :181, pl. 13, , 10; Fleming Citation1966:51, fig. 1020, 1022; Beu & Maxwell Citation1990:318, pl. 42f; Maxwell Citation2009: 241.
Struthiolaria fraseri. Hector 1886:48, .1; Hutton 1893:61.
Struthiolaria papulosa. Marwick Citation1924a:180, pl. 12, only (in part not S. papulosa Martyn, 1784).
Struthiolaria tasmani King Citation1933:340 (in part, not pl. 39, Fig. 30); Fleming Citation1966:51, fig. 1029.
Type material
Marwick (Citation1924a:181) discussed the possible type material of Struthiolaria frazeri, and concluded that the holotype is the specimen then in the ‘Geological Survey collection at the Dominion Museum’. These type specimens were transferred to the New Zealand Geological Survey in the 1950s. The holotype (an unusually small specimen) is TM8284, GS231, V21/f8466, from ‘McLean's’, that is, Donald McLean's station at Maraekakaho, opposite ‘Shrimpton's’ on the S side of the Ngaruroro River, central Hawke's Bay; from fine-grained interbeds in Scinde Island Limestone of Mason Ridge. Specimens of S. frazeri in BMNH recorded by Harris (1897:221) are from the same collection, made by Hector in 1871. Struthiolaria tasmani King, Citation1933, holotype TM8703, the specimen illustrated by Marwick (Citation1924a, pl. 12, ), a severely abraded specimen from GS1094, U21/f8494, Mangatahi River, Kereru Road, central Hawke's Bay (late Nukumaruan), apparently a weakly sculptured form of S. frazeri.
Other material examined
Mangapanian: an ancestral Mangapanian–early Nukumaruan form of Struthiolaria frazeri occurs in shallow-water facies rocks of northern Hawke's Bay: GS12508, V19/f012, brown sand beneath three obvious major conglomerate beds, Matahorua Road, W of Tutira, N of Napier, Hawke's Bay (seven); GS13076, W19/f027, brown sands underlying Nukumaruan siltstone in Mohaka River, upstream from Highway 2 bridge, N Hawke's Bay (five); GS13079, W19/f031, as last, at small side waterfall 20 m further downstream (two). Early Nukumaruan: GS12865, V21/f095, roadside, Napier-Taihape Road, 2 km W of Sherenden hall, low in Nukumaruan rocks of Matapiro syncline (one); definitely Nukumaruan in age, underlying by about 30 m the Nukumaruan limestone-conglomerate succession of the Flag Range (Flag Range Limestone; Beu Citation1995:130). Late Nukumaruan: almost the only previous record of Struthiolaria frazeri from Wanganui Basin is that by Marshall & Murdoch (1920:123) ‘over a distance of [1.6 km] south [actually ESE] from the Nukumaru boat-landing’). Fleming (Citation1953:158) assigned this specimen to Tewkesbury Formation. Marwick (Citation1924a:182) recorded the specimen as ‘broken to pieces in transmission from Mr. Suter’, and a fragmentary but reglued specimen labelled ‘12 m below moa bed, Nukumaru’ in GNS is evidently this shell. The only complete Wanganui Basin specimen I am aware of is an unusually large specimen from the Suter collection in GNS labelled ‘Wanganui, Pliocene’. It has a yellow stain suggesting that it came from sands or Nukumaru Limestone facies high in the succession (e.g. from a horizon such as Waipuru Shellbed, or higher), rather than the grey stain of specimens from Tewkesbury Formation. Therefore, it is interesting that RM Carter (James Cook University, Townsville pers. comm. August 2003) reported an incomplete spire of S. frazeri in ‘shellbed f’ (of Abbott et al. [Citation2005], ], in the second cycle up from the base of Tewkesbury Formation, 18 m above the base of the section, deposited during OIS 71) on the Nukumaru coast, seen during a visit in August 2003. The only other Wanganui Basin record I am aware of is that by Fleming (Citation1953:148) from GS4341, S22/f6466, a ‘sub-Waipuru’ shellbed in the Turakina valley, E of Wanganui.
All other records are from central and northern Hawke's Bay. Struthiolaria frazeri is abundant in two shellbeds high in Okauawa Formation, cropping out in Okauawa Stream, Whanakino Stream, Poporangi Stream and nearby streams in the Kereru district, inland central Hawke's Bay, high in the Nukumaruan succession in this area (stratigraphy described by Erdman & Kelsey 1992): GS5439, V21/f8575, Okauawa Stream (six); GS10943, V21/f8575A, lower 200 m of Okauawa Stream (two); GS12868, U21/f028, road cutting W side Whanakino Stream (two); GS12866, U21/f033, Okauawa Stream (15); GS14584, V21/f096, Racham Station (one). Specimens also occur as scattered individuals at a range of other localities in Hawke's Bay, from ‘not far below the Scinde Island limestone at Maraekakaho, Ngaruroro River’, that is, in interbeds within Mason Ridge limestone, very near the type locality (Marwick Citation1924a:182); in several limestone beds in the Waipunga Road section, ascending from Esk Valley towards Devils Elbow; underlying the Devil's Elbow succession at the Aropaoanui Stream bridge on the Napier-Wairoa Highway; in Mairau Mudstone at ‘the watchman’ hill, Ahuriri, Napier (GS11465, V21/f8015A, two fragmentary); and in Darkys Spur Formation on Darkys Spur road, 5 km WNW of Devils Elbow (GS11225, V20/f8002, two; common but difficult to remove) to pockets in sandstone high in the Nukumaruan succession in Matahorua Gorge, on the Napier-Wairoa Highway, and scarce specimens in mudstone near the Mohaka River highway bridge, also in northern Hawke's Bay. It extends into late Nukumaruan rocks, in Tewkesbury and Okauawa Formations, although I am not aware of a record from the highest Nukumaruan fossiliferous unit in Wanganui Basin, Pukekiwi Shell Sand.
Distribution
Struthiolaria frazeri is a valuable, highly distinctive, Mangapanian–Nukumaruan index fossil. Most records are from late Nukumaruan rocks of central and northern Hawke's Bay, but four specimens are recorded from Wanganui Basin. I am not aware of any specimens from Wairarapa district or anywhere further south, so evidently it was a warm-water species. Intergrading ancestral specimens are also recorded here from Mangapanian–early Nukumaruan rocks of central and northern Hawke's Bay.
Dimensions
Struthiolaria frazeri, holotype, H 73.9, D 42.2 mm; large specimen, GS14584, Racham Station, Kereru Road: H 93.4, D 48.8 mm; Suter's ‘Wanganui Pliocene’ specimen: H 96.8 (apex incomplete; estimated originally 100 mm), D 52.9 mm; GS12865, Sherenden, Hawke's Bay, early Nukumaruan: H 81.2, D 43.6 mm; GS13076, Mohaka River, Mangapanian: H 69.6, D (slightly incomplete) 38.5 mm.
Remarks
Hector (1886:48) and Hutton (1893:61) spelled the species name ‘fraseri’, rather than the original and more usual spelling ‘frazeri’. The only likely source of the name is Charles Fraser (1823?–1886) (Breward Citation1990), who, as one of the founders of both the Philosophical Institute of Canterbury (secretary 1868–1869) and the Canterbury Museum, would have been well known to both Hector and Hutton. However, I know of no published evidence that the name was intended to honour Charles Fraser, so the original spelling frazeri should be retained.
Struthiolaria frazeri is a highly distinctive Nukumaruan index fossil. Firstly, it is the largest struthiolariid, reaching 100 mm in height. Also, it has much coarser and more prominent spiral cords than any other struthiolariid, and among the most prominent of all New Zealand gastropods. Its spire also is unusually tall, with a strongly stepped outline, and with a rather narrow but almost horizontal sutural ramp. The rounded shoulder angle protrudes a little beyond the weakly to quite strongly concave lateral shell outline anterior to the periphery, and on some specimens the shoulder is weakly or, in a few specimens, quite strongly nodulose. The few, very wide, strongly raised, flat-topped spiral cords are closely spaced around the shoulder angle and on the peribasal angle, but more widely spaced both above and below both angulations, becoming low and narrow over the central area of the base. The aperture is very prominently but rather narrowly callused for a Struthiolaria species. The varix is flat but unusually wide in the spiral plane, and corrugated at the back where it meets the spiral cords.
Specimens occurring in Mangapanian and early Nukumaruan rocks of central and northern Hawke's Bay (A–C) are interpreted as ancestral, intergrading specimens. They are similar to younger specimens of Struthiolaria frazeri in their tall, narrow shape, with a strongly stepped spire and relatively small last whorl with weakly concave outlines below the peripheral angle, in having more prominent spiral sculpture than other Struthiolaria species, and in their prominently but rather narrowly callused aperture. They differ from middle to late Nukumauan specimens of S. frazeri in their smaller size, and in particular, in their narrower, more numerous and more closely spaced spiral cords. While the cords are very clearly defined, with vertical sides and a flat crest as in later specimens of S. frazeri, there are 6–7 cords on the sutural ramp (three on late Nukumaruan S. frazeri), 5–7 cords between the peripheral and peribasal angles, with many fine interstitial cords on some specimens (3–4, without interstitial cords, on late Nukumaruan S. frazeri), and 20–30 fine, narrow, closely spaced cords, including many interstitial ones, on the base (six plus several faint, narrow ones in late Nukumaruan S. frazeri). These characters suggest that Mangapanian–early Nukumaruan specimens are intermediate between a more typical Struthiolaria species resembling S. papulosa (Martyn, 1784) and late Nukumaruan specimens of S. frazeri. However, the characters of late Nukumaruan specimens of S. frazeri also vary ecophenotypically and specimens from more offshore environments, e.g. Mairau Mudstone at ‘the watchman’ hill, Ahuriri, Napier, closely resemble Mangapanian–early Nukumaruan ones. The holotype of S. tasmani King seems to be a relatively short, weakly sculptured ecomorph of S. frazeri of this type; such specimens are common in a second shellbed near Kereru Road overlying the one with abundant S. frazeri, whereas the specimens from S Wairarapa identified by King (Citation1933:34, pl. 39, Fig. 30) as S. tasmani definitely are strongly sculptured specimens of S. papulosa. Beu & Maxwell (Citation1990:350) included S. tasmani in the synonymy of S. papulosa but, strictly speaking, following King's (1933) designation, the holotype is Marwick's illustrated specimen from near Kereru Road, inland Hawke's Bay. Pliocene species of Struthiolaria (sensu stricto) are poorly understood, and the ancestor of S. frazeri is not obvious. Assuming the early form is not simply ecophenotypic, S. frazeri seems to provide yet another example of an anagenetically evolving succession. As there is no point in this succession at which a subdivision into more than one species would be anything other than arbitrary, in my opinion the Mangapanian–Nukumaruan form is part of the variation of one species, S. frazeri. The earlier form possibly can be used to identify rocks of Mangapanian–early Nukumaruan age in shallow-water facies in Hawke's Bay, but must be used with care in view of the phenotypic variation observed in late Nukumaruan specimens.
Family Calyptraeidae
Genus Maoricrypta Finlay, 1926
Maoricrypta Finlay Citation1926b:393. Type species (by original designation): Crepidula costata G. B. Sowerby I, 1824, Pleistocene and Recent, New Zealand.
Zeacrypta Finlay Citation1926b:393. Type species (by original designation): Calyptraea monoxyla Lesson, 1831, Pleistocene and Recent, New Zealand.
Remarks
Marshall (Citation2003) recognized Maoricrypta as a valid genus for most New Zealand species that recently have been referred to Crepidula, on the basis of its two muscle scars; no muscle scars are visible in most species of Crepidula (but see Simone [Citation2006], fig. 122]; a single scar is present in some South American species of the C. plana Say, 1822 species complex). The generic distinction was confirmed by Collin (Citation2003) on the basis of molecular phylogeny. Marshall (Citation2003:118) also recognized that although most of the younger species in New Zealand have two muscle scars, one on each side of the columellar septal insertion, and belong in Maoricrypta (M. costata [see below], Castlecliffian–Recent; M. haliotoidea Marwick, Citation1926, Tongaporutuan, N Taranaki; M. kopua Marshall, Citation2003, Recent; M. monoxyla (Lesson, 1830), Recent; M. profunda (Hutton, 1873) [see below]; M. sodalis Marshall, Citation2003, Haweran [Landguard Bluff, OIS 9]–Recent; M. radiata (Hutton, 1873) [see below], the Tongaporutuan-Opoitian stacking species, and M. youngi Powell, Citation1940 [see below]). A few other species have only one muscle scar, on the right side, and belong in Grandicrepidula McLean, Citation1995 (see below).
McLean (Citation2009) recognized that still further genera of Calyptraeidae can be defined in the ‘Crepidula-group’ based on the number of muscle scars. He pointed out that they are separately evolved conditions. One scar (the presumed basal condition) is present in Bostrycapulus, Crepipatella, Grandicrepidula and an unnamed genus, two scars are present in Garnotia and Maoricrypta, and no scars are present in Crepidula (sensu stricto), Verticumbo and Siphopatella.
Dimensions
For dimensions of specimens of Maoricrypta and Grandicrepidula special abbreviations are required because of their atypical shape; H is used for greatest dimension (overall height of the specimen, parallel to the apertural plane), W for width, and D for depth (height normal to the apertural plane).
Maoricrypta costata (G. B. Sowerby I, 1824) (B)
Fig. 13 (A,C-F) Struthiolaria frazeri Hutton; A, GS12865, V21/f095, early Nukumaruan sandstone underlying Flag Range Limestone, Napier-Taihape Road W of Sherenden, Hawke's Bay; height 81.2 mm; C,D, GS13079, W19/f027, Mangapanian sandstone, Mohaka River, N Hawke's Bay; C, height 74.9 mm; D, height 64.5 mm; E,F, shallow-water Nukumaruan form, GS11225, V20/f8002, Darkys Spur Formation (late Nukumaruan), Darkys Spur, W of Devils Elbow, central Hawke's Bay; height 95.8 mm. (B) Maoricrypta costata (G. B. Sowerby I) attached to underside of Astraea heliotropium (Martyn), GS4014, R22/f6513, basal shellbed member of Shakespeare Cliff Sand (Castlecliffian, OIS 11), Castlecliff; width of Astraea 64 mm.

Crepidula costata GB Sowerby I 1824, part 23, pl. 152, ; Quoy & Gaimard 1835:414, pl. 72, , 12; Gray 1843:242; Reeve 1842, in 1841–1842:29, pl. 143, ; Reeve 1859, pl. 4, a,b; Martens Citation1873:28; GB Sowerby III Citation1883:67, pl. 452, fig. 113, 114; Suter Citation1913:287, pl. 44, , 6a; Iredale Citation1915:456; Hoagland Citation1977:370; Beu & Maxwell Citation1990:408; Bandel & Reidel 1994:341; Spencer & Willan Citation1996:21.
Crepidula costata Deshayes Citation1830:26 (synonymous junior primary homonym of Crepidula costata G. B. Sowerby I, 1824; junior primary homonym of Crepidula costata Morton, 1829); Hoagland Citation1983:2.
Crypta costata. Deshayes 1838 [in 1835–1845], p. 644; Hutton 1873a:32; Hutton 1873b:14; Hutton 1880:87; Hutton 1883:122, pl. 14, fig. A.
Crepidula (Crypta) aculeata. Tryon 1886:129, pl. 39, fig. 65 only (in part not Crepidula aculeata Gmelin, 1791).
Crepidula aculeata Suter in Hutton Citation1904:79 (not Crepidula aculeata Gmelin, 1791).
Maoricrypta costata. Finlay Citation1926b:393; Fleming Citation1966:50; Pilkington Citation1974:419; Powell Citation1979:149, pl. 30, ; Marshall Citation2003:118, , 2, 11, 25, 26, 66; Spencer et al. Citation2009:204; Maxwell Citation2009:242.
Crepidula (Maoricrypta) costata. Wenz 1940: 904, fig. 2661.
Type material
Crepidula costata G. B. Sowerby I, three ‘probable syntypes’ BMNH 1993118, from ‘New Zealand’, from the collection of H Cuming. The largest syntype appears to be that illustrated by Sowerby (1824, pl. 152, ) (as also was commented by Marshall Citation2003:119), demonstrating that at least this specimen is an authentic syntype, and by Reeve (1842:29, pl. 143, ; Reeve 1859, pl. 4, a,b); Reeve's illustrations are both reprintings of Sowerby's (1824) plate (Petit Citation2006, appendix 2). Crepidula costata Deshayes, six syntypes in MNHN (recorded, but not illustrated, by Hoagland Citation1983:2), from ‘Baie des Iles’ (Bay of Islands), northern North Island; all are normal, coarsely ribbed specimens of C. costata, with a muscle scar clearly visible on each side of the front margin of the columellar septum.
Other material examined
Castlecliffian: recorded in GNS files from: Kaimatira Pumice Sand (OIS 25), Castlecliff (GS4064, R22/f6395), Kaikokopu Shellbed (OIS 19), Turakina valley (GS15194, S22/f0159); Lower Castlecliff Shellbed (OIS 15), Turakina valley (GS15141, S23/f085); ‘Otapatu’ shellbed (OIS 13), between Kakariki and Onepuhi, E bank Rangitikei River (GS13720, S23/f022); Shakespeare Cliff Sand (OIS 11), Turakina valley (GS15182, S22/f0161). Haweran: Te Piki, near East Cape, OIS 7; Waipuna Conglomerate (OIS 7), coast E of Wanganui (GS14151, R22/f0129); Hauriri Terrace shellbed (OIS 5a), Waverley Beach (GS4174, Q22/f7464; Fleming Citation1953:273); Holocene, Mahia Peninsula, and Cape Kidnappers (GS13796, W21/f057, terrace at shelter hut, one fragment). Recorded also by Fleming (Citation1953:188,203,226,229,236,261) from Kaimatira Pumice Sand (OIS 25), Kupe Formation (OIS 17), Pinnacle Sand (OIS 15–14), Tainui Shellbed (OIS 13), Shakespeare Cliff Sand (OIS 11) and Brunswick Pebbly Sand (OIS 9; as ‘Maoricrypta aff. radiata’) at Wanganui, and Oturi shellbed, mouth of Wairoa Stream, Waverley Beach (OIS 5a; GS4174, Q22/f7464; Fleming 1953:273).
Distribution
OIS 25 to OIS 5a, Wanganui; Ohope Beach, Whakatane, OIS 15? (NMNZ M.43129, shellbed below old reservoir in stream at foot of Ohope-Whakatane road, four specimens; Marshall Citation2003:119); Te Piki, Whangaparaoa, near East Cape, OIS 7; Holocene; that is, a range of mid-Castlecliffian to Recent; Recent in the northeastern North Island only, as far south as Mahia Peninsula, living intertidally down to 47 m (Marshall Citation2003:119). Richardson (2002) recorded M. costata from Hawke's Bay Nukumaruan rocks, but this record is presumably based on relatively prominently ribbed specimens of M. profunda (see below). Boreham (in Kingma Citation1971, table p. 91, and table 10A) also recorded M. costata from at least 21 localities in early to late Nukumaruan rocks of central Hawke's Bay, but only M. profunda is present (along with what is apparently M. monoxyla (Lesson, 1830)) in all GNS collections from Nukumaruan rocks of Hawke's Bay, except for the record of Grandicrepidula hemispherica n. sp. (below) at Maharakeke Road, W of Waipukurau.
Dimensions
Crepidula costata G. B. Sowerby I, largest (figured) syntype: H 42.6, W 31.1, D 15.5 mm; Crepidula costata Deshayes, largest syntype: H 37.0, W 26.4, D 12.8 mm; RM2523, Takapuna Beach: H 46.6, W 34.9, D 18.4 mm; NMNZ M.16454, dredged off Takapuna, Auckland: H 55.8, W 32.4, D 19.8 mm; H 42.5, W 32.7, D 18.8 mm.
Remarks
Hoagland (Citation1977:371) suggested that Crepidula lineolata Deshayes (1830:26) might be a further synonym of C. costata Sowerby, but later (Hoagland Citation1983:4) she showed that the three syntypes (in MNHN) include one specimen of C. excavata (Broderip, 1834) (Chile) and two specimens of the widespread North Atlantic species C. fornicata (Linné, 1758). Hoagland (Citation1977) also pointed out that the New Zealand species was named Crepidula costata independently twice, first by GB Sowerby I (1824) and later by Deshayes (1830).
Maoricrypta costata is the large, abundant, consistently prominently costate, externally convex Maoricrypta species of northern New Zealand at present. It lives only in the northeastern North Island warm-water province, from Cape Maria van Diemen to East Cape, and sparsely as far south as Mahia Peninsula. The only recorded fossils are from Castlecliff, Wanganui (OIS 25, and a continuous record from OIS 19 to 5a), from the Castlecliffian of Ohope Beach, Whakatane (Marshall Citation2003:119) and the late middle Pleistocene of Te Piki, Whangaparaoa, near East Cape (OIS 7; Marshall Citation2003:119). This is therefore another warm-water immigrant at Wanganui from the northern North Island. The record of M. costata from upper Okiwa Group (early Nukumaruan) at Wanganui by Fleming (Citation1953:134) is presumably based on a strongly sculptured specimen of M. profunda (Hutton, 1873) (GS4346, S22/f6471, siltstone 1 m above Hautawa Shellbed, Turakina River), but I have not been able to find this specimen in GNS collections. Specimens in Fleming's collections from nearby localities definitely are relatively coarsely sculptured specimens of C. profunda. A few specimens in GNS from Target Gully Shellbed, Oamaru (Altonian, early Miocene) represent an unnamed species resembling C. costata, but with thinner, sharper radial ridges. The protoconch of Recent specimens was illustrated by Marshall (Citation2003, Fig. 25, 26) and consists of about one smooth whorl, with a large, rounded, inflated initiation and an obvious spiral suture.
Maoricrypta profunda (Hutton, 1873) (A–D, G–I)
Crypta profunda Hutton 1873b:14.
Crepidula monoxyla Hutton 1893:62 (in part not Calyptraea (Crepidula) monoxyla Lesson, 1830).
Crepidula crepidula. Suter Citation1914:5 (in part; not Patella crepidula Linné, 1767).
Maoricrypta radiata. Marwick Citation1948:30; Fleming Citation1953:109–209; Fleming Citation1966:50 (in part not Pilaeopsis radiatus Hutton, 1873).
Crepidula radiata. Beu & Maxwell Citation1990:259,408, pl. 29j (in part not Pilaeopsis radiatus Hutton, 1873).
Crepidula youngi. Beu & Maxwell Citation1990:259 (in part not Maoricrypta youngi Powell, Citation1940).
Maoricrypta profunda. Marshall Citation2003:122, , 6; Maxwell Citation2009:242.
Type material
Crypta profunda, lectotype (A,B; designated by Beu & Maxwell Citation1990:259) TM8223, supposedly from ‘Shakespeare Cliff’, Wanganui; suggested by Marshall (Citation2003:122) to actually be from Tangahoe Formation at Waihi Beach, Hawera (Waipipian), a provenance confirmed by the soft grey sandstone inside the shell.
Other material examined
Not listed, abundant in all Waipipian–early Castlecliffian (to OIS 19, perhaps younger) rocks throughout New Zealand. Exceedingly abundant in the Otahuhu well, Auckland (Waipipian). Recorded by Fleming (Citation1953:109–113, 118, 124, 131, 134, 139, 144, 146, 148, 151, 158, 174, 178, 181, 182, 188, 198, 209) as Maoricrypta radiata, from Pepper Shell Sand, Rangikura Sandstone, Wapipi Formation (Waipipi shellbeds), Waverley Formation, Mangapani Shell Conglomerate, Wilkies Shellbed, Hautawa Shellbed, upper Okiwa Group (recorded as M. costata, p. 134), Nukumaru Limestone, Nukumaru Brown Sand, Mangamako Shellbed, undifferentiated Nukumaru Group, Waipuru Shellbed, Tewkesbury Formation, Butlers Shell Conglomerate (OIS 31; listed in parentheses, indicating ‘known or suspected to be derived from older sediments’; Fleming Citation1953:175), basal conglomerate member of Lower Okehu Siltstone (OIS 29; recorded as ‘Maoricrypta aff. radiata’), Okehu Shell Grit (OIS 27; listed in parentheses), basal conglomerate member of Upper Okehu Siltstone (OIS 27; not listed in parentheses), Kaimatira Pumice Sand (OIS 25; not listed in parentheses), Kaikokopu Shellbed (OIS 19; listed as ‘[Maoricrypta aff. radiata]’; indicating that this record was ‘known or suspected to be derived from older sediments’), and undifferentiated Kai-Iwi Group (not listed in parentheses, that is, thought to be in situ). This includes almost all formations in Wanganui Basin of Waipipian to early Castlecliffian age, up to OIS 19. Also recorded by Fleming (in Kingma Citation1971:98) from Maraetotara Sand (early Castlecliffian) at Cape Kidnappers, Hawke's Bay (GS5378, V22/f8768, as Maoricrypta radiata).
Distribution
Maoricrypta profunda is now adopted as the name for the large, abundant, widespread Maoricrypta species in New Zealand mid-Pliocene–early Pleistocene rocks, succeeding M. radiata (Hutton, 1873), the common Tongaporutuan–Opoitian species that formed tall, irregular to spiral stacks of specimens living on top of each other. Stacks were not formed by Waipipian–early Castlecliffian specimens. The known time range is Waipipian to early Castlecliffian (OIS 19), but it is difficult to evaluate whether some of the Opoitian records here assigned to M. radiata might actually be specimens of M. profunda; collection and examination of further juvenile specimens with protoconchs will be needed to clarify the taxonomy of this genus further. Protoconchs as described by Marshall (Citation2003:119, Fig. 28, 29) for the ‘smaller’ species aff. M. costata, with a clearly visible protoconch suture, have been confirmed on material assigned to M. profunda from: GS3528, R11/f7014, Otahuhu well, Auckland (Waipipian); GS11465, V21/f6015A, Mairau Mudstone, ‘the watchman’ hill, Ahuriri, Napier (Nukumaruan); GS10858, U22/f09, Makateru Mudstone (Thomson Citation1926), Ashcott Road, W of Waipukurau (Nukumaruan); and GS4084 re-collection, R22/f6414, Pukekiwi Shell Sand, Ototoka Beach, Wanganui (latest Nukumaruan). Specimens assigned to M. radiata from GS8101, Y14/f7618A, Oweka Creek, between Whangaparaoa and Hicks Bay, East Cape (Opoitian) and Kaawa Creek, SW Auckland (AUGD G5739, holotype of Crypta opuraensis Bartrum & Powell) have a markedly longer spiral suture visible on the protoconch, similar to that of M. costata (Marshall Citation2003, Fig. 25, 26) but still more strongly spiral (G).
Dimensions
Lectotype, TM8223: H 31.5, W 17.5, D 16.7 mm; GS14094, Q21/f6501A, Tangahoe Formation (Waipipian), Waihi Beach, Hawera: H 65.7, W 41.6, D 29.1 mm; GS4124, R22/f6543, Wilkies Shellbed (Mangapanian), Wilkies Bluff, W of Wanganui: H 63.2, W 34.0, D 24.6 mm; GS4117 re-collection, R22/f6437, Nukumaru Brown Sand (Nukumaruan), Nukumaru Beach, Wanganui: H 50.0, W 32.3, D 24.5 mm.
Remarks
The taxonomy of Cenozoic fossil Maoricrypta species in New Zealand is complex and as yet very poorly understood (Beu & Maxwell Citation1990:259; Marshall Citation2003). Marshall (Citation2003) pointed out that Beu & Maxwell (Citation1990:259) were wrong to suggest that M. profunda was an earlier name for M. youngi Powell, Citation1940, as they have different protoconchs—narrower at the end, joining the teleoconch (130 µm in M. profunda, 330–430 µm in M. youngi)—and the matrix in the lectotype of M. profunda is not that of any Castlecliff formation, but is a soft grey sandstone closely resembling Tangahoe Formation at Waihi Beach, Hawera (Waipipian). Waihi Beach is here taken to be the type locality, implying (as noted by Marshall Citation2003) that this is the earliest name for the abundant, widespread Maoricrypta species in New Zealand Pliocene–early Pleistocene rocks. Most specimens are relatively long and narrow (‘boat-shaped’) with a weakly recurved apex, and the lectotype agrees with this common form. Specimens are almost as strongly variable in the prominence of the spiral sculpture as is M. radiata (see below), from completely smooth to prominently ribbed, but none is as prominently ribbed and spinose as are normal specimens of M. costata. A range of variants is illustrated here. Marshall (Citation2003:119, Fig. 28, 29) described and illustrated two late Pliocene species of Maoricrypta from Mangahao, N Wairarapa, one with and one without a protoconch suture visible, but lacked comparative material to decide which species they were—although it should be noted that at least Grandicrepidula hemispherica n. sp. (below) is quite variable in the extent to which the suture is visible. Examination of juvenile specimens from many localities has demonstrated that the species with the protoconch suture clearly visible is M. profunda, although most protoconchs of pre-Pliocene Maoricrypta specimens from New Zealand have proved to be similar to this. A second species without the suture visible has proven to be much more elusive, although it is possibly Grandicrepidula hemispherica n. sp. (below).
A few large specimens from Tangahoe Formation (Waipipian) at Waihi Beach, Hawera (NMNZ M.276921, three specimens) bear obvious scars on the dorsum formed by the anterior end of other specimens that formerly lived attached to them, demonstrating that some early specimens of M. profunda formed short stacks of 2–3 similar-sized specimens, but I have not seen any specimens that were collected attached to each other in stacks, so this was evidently not their normal mode of life.
The significant point about Maoricrypta profunda for molluscan biostratigraphy is the age of its youngest occurrence. Following long-accepted time ranges (e.g. Fleming Citation1966:50), I have previously assumed that M. profunda (that is, the Pliocene–early Pleistocene non-stacking species previously known as M. radiata, with the protoconch suture clearly visible on juveniles) occurred last in late Nukumaruan rocks. The explanation usually assumed for the common early Castlecliffian specimens has been that they have been reworked from Nukumaruan or older rocks. However, it is now clear that specimens of M. profunda in early Castlecliffian shellbeds in Wanganui Basin are too abundant and too little-abraded to be reworked Nukumaruan specimens (particularly abundant in GS15338, S22/f0164, S22/096385, shellbed, Turakina Valley Road, east of Wanganui, ca 600 m S of Mangara Stream bridge and ca 200 m N of Glenroy Station; early Castlecliffian, OIS 43–41?). They indicate that M. profunda definitely survived into early Castlecliffian rocks. Its upper age limit is uncertain at present, as the reality of Castlecliffian occurrences has only recently been accepted, but seems likely to be above OIS 25 (Kaimatira Pumice Sand) and probably as young as Kaikokopu Shellbed (OIS 19). As usual, so many of the molluscs in Kaikokopu Shellbed are abraded and evidently reworked from older rocks as to make the reality of unreworked Kaikokopu occurrences likely but uncertain.
The later specimens (Nukumaruan–Castlecliffian) of Maoricrypta profunda are significantly shorter, wider and more inflated than Waipipian–Mangapanian specimens, and most are not prominently sculptured, but dimensions overlap greatly, and the change (if indeed it represents anagenetic evolution rather than just phenotypic variation) is very gradual. All Waipipian–early Castlecliffian specimens are interpreted as one species here but, as usual, more research is required to clarify this genus. It seems likely that younger specimens merely inhabited a different substrate from the earlier ones, but the fossil history could also be interpreted as a gradual withdrawal from the stacking habit to living individually on flat surfaces through early Pliocene (Opoitian–Waipipian) time.
Maoricrypta radiata (Hutton, 1873) (F,K, A–I, A)
Fig. 14 (A-D, G–I) Maoricrypta profunda (Hutton); A,B, lectotype, TM8223, “Shakespeare Cliff” (wrong; Tangahoe Formation (Waipipian), Waihi Beach, Hawera); height 31.5 mm; C,D, 2 specimens, GS4253, Q22/f7544, Upper Waipipi Shellbed (Waipipian), Waverley Beach, W of Wanganui; C, height 45.1 mm (radial costae on early part only); D, height 38.5 mm (narrow radial costae all over); G, GS4356, S22/f6481, Nukumaru Brown Sand (late Nukumaruan), Mangawhero Road, W of Wanganui; height 40.4 mm; H, I, GS14094, Q21/f6501A, Tangahoe Formation (Waipipian), Waihi Beach, Hawera; large smooth specimen, height 64.6 mm. (E,J) Maoricrypta youngi Powell, Castlecliff fossils; E, GS4014, R22/f6513, basal shellbed member of Shakespeare Cliff Sand; left side view, height 14.4 mm; J, GS4022, R22/f6360, Pinnacle Sand (OIS 14), head of “the pinnacles” gully; dorsal view, height 15.2 mm. (F,K) Maoricrypta radiata (Hutton), holotype of Crypta opuraensis Bartrum & Powell, AUGD G5739, Kaawa Creek, SW Auckland, Opoitian; height 23.5 mm.

Fig. 15 Maoricrypta radiata (Hutton). (A,B) holotype of Crypta turnialis Bartrum & Powell, AUGD G5737, Kaawa Creek, SW Auckland, Opoitian; height 31.6 mm. (C,D) dorsal and lateral views, specimen formerly identified as holotype of Pilaeopsis radiata (not a type), TM8580, “Awatere”, Kapitean-Opoitian; height 57 mm. (E) GS14892, P29/f0395, Upton Brook, Awatere Valley, Kapitean; 2 smooth and 2 sculptured specimens attached to each other, width 69 mm. (F) lectotype of Crepidula hochstetteriana Wilckens (tentatively synonymised with M. radiata), TM2608, GS13, O32/f8025, Haumuri Bluff, Marlborough, “Late Cretaceous” (Middle Miocene); dorsal view of partial slab with lectotype mould, height 50 mm. (G) AUGD G7130, Kaawa Creek, SW Auckland, Opoitian; juvenile specimen showing protoconch (terminating at line at arrow tip); SEM, height 2.4 mm. (H) GS10616, S28/f6169, Hurupi Formation (Tongaporutuan), Whatarangi cliffs, E Palliser Bay; 4 specimens attached to each other, width 90 mm. (I) holotype of Pilaeopsis radiata Hutton, TM8581, Awatere Valley, Kapitean?; whole block 34 mm wide, unwhitened to show original labels.
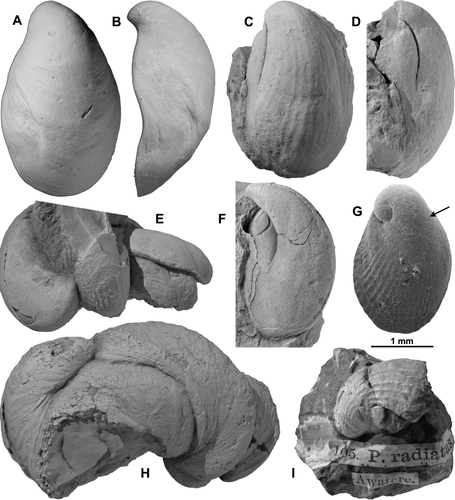
Fig. 16 (A) Maoricrypta radiata (Hutton), GS10950, P29/f6581, Upton Brook, Awatere Valley, Kapitean; 2 stacked, coarsely sculptured specimens, height 41 mm. (B-E, G-I) Grandicrepidula hemispherica n. sp., GS10857, U23/f7049, Maharakeke Mudstone (Nukumaruan), Maharakeke Road, W side Pukeora Hill, W of Waipukurau, Hawke's Bay; B,D,G, largest paratype, TM8689; height 35.0 mm, dorsal, ventral and lateral views; C,E,H,I, holotype, TM8688, height 24.3 mm; C,E,H, lateral, dorsal and posterior views; I, protoconch in H enlarged. (F) Zelippistes benhami (Suter), type species of Zelippistes Finlay, 1926, NMNZ M.100507, Recent, King Bank, Three Kings Islands, 123-128 m, 32°57.40'S, 172°19.40'E; dorsal view, SEM, diameter 10.8 mm.

Crepidula incurva Zittel 1864:44, pl. 15, ; Harris 1897:248 (junior primary homonym of Crepidula incurva Orbigny, 1841).
Crepidula sp. Zittel 1864:44, pl. 15, .
Pilaeopsis radiatus Hutton, 1873b:14.
Crypta striata Hutton 1873b:14 (proposed for specimen illustrated by Zittel 1864:44, pl. 15, ).
Hipponyx radiatus. Hutton Citation1887:218.
Hipponix radiatus. Suter Citation1914:20, pl. 1, .
Crepidula gregaria. Ortmann Citation1902:184 (in part not C. gregaria G. B. Sowerby II, 1846, South American); Suter Citation1914:20, pl. 2, a,b.
Crepidula crepidula. Suter Citation1914:5 (in part; not Patella crepidula Linné, 1767).
Crepidula hochstetteriana Wilckens Citation1922:5, pl. 1, a,b (new synonym).
Crepidula wilckensi Finlay Citation1924d:101 (replacement name for C. incurva Zittel, 1864, not C. incurva Orbigny, 1841).
Crepidula radiatus. Finlay Citation1924d:100.
Crepidula radiata. Powell Citation1924:283; Beu & Maxwell Citation1990:259,408, pl. 29j (in part not Pilaeopsis radiatus Hutton, 1873).
Crypta turnialis Bartrum & Powell 1928:144, pl. 27, Fig. 23, 24 (new synonym).
Crypta opuraensis Bartrum & Powell 1928:145, pl. 27, Fig. 25 (new synonym).
Maoricrypta radiata. Finlay Citation1926b:394; Marwick Citation1948:30; Fleming & Marwick in Lillie Citation1953, tables 21, 22; Fleming Citation1966:50 (in part not Pilaeopsis radiatus Hutton, 1873); Maxwell Citation2009:242.
Maoricrypta striata. King Citation1934:23.
Maoricrypta wilckenseni (sic). Dell Citation1952:74, .
Maoricrypta wilkinsi (sic). Fleming & Marwick in Lillie Citation1953, table 21.
Maoricrypta opuraensis. Fleming Citation1966:50; Maxwell Citation2009:242.
Maoricrypta turnialis. Fleming Citation1966:50; Maxwell Citation2009:242.
Crepidula turnialis. Grant-Mackie Citation1990:399, fig. 330–333; Beu & Maxwell Citation1990:408.
Crepidula opuraensis. Beu & Maxwell Citation1990:408.
Maoricrypta wilckensi. Maxwell 2009:242.
Type material
Crepidula incurva Zittel (not of Orbigny), holotype NHMW 1959/335/29, labelled ‘Awatere-Thal’, probably from the Kapitean–Opoitian succession in Upton Brook, Awatere valley, where stacked specimens of Maoricrypta radiata are common, particularly in Kapitean rocks. Zittel's (1864:44, pl. 15, ) other illustrated but unlabelled specimen, the holotype of Crypta striata Hutton, is NHMW 1959/335/30, also from ‘Awatere-Thal’, that is, the same locality as C. incurva.
Pilaeopsis radiatus, supposed holotype TM8580 (C,D), ‘Awatere’, apparently also from the Kapitean–Opoitian succession of Upton Brook (Marwick Citation1948:30). This specimen bears a label in J Marwick's handwriting, pointing out that there are areas of glue on the original wooden tablet that do not match those formerly attaching the supposed holotype, and ‘it may be that this specimen was wrongly glued to the label after Suter had handled it’. On the back of the label Marwick has also written: ‘Hutton gave the dimensions of his Pilaeopsis radiatus as 0.65×0.65 [inches]. Obviously this is not his specimen [the supposed holotype is 56.8 mm long and 45.5 mm wide, or about 2.2×1.8 inches]. Equally obviously it is a Crepidula with the platform showing. Hutton could not have missed that’. I can add that the original wooden tablet is not large enough to accommodate the specimen without it extending beyond the edges of the tablet, whereas all other original Hutton wooden tablets remaining in GNS are about 20–40% larger than the attached specimen. Therefore, there is little doubt that TM8580 is not Hutton's holotype, and it has been wrongly attached to the original labelled type tablet. It appears that TM8580 was not an original type specimen.
Another original Hutton specimen in GNS, TM8581 (I), is much smaller (an incomplete, strongly ribbed Maoricrypta shell about 23 mm long and 15 mm high, or about 0.8×0.6 inches, in a block of concretionary sandstone 34×33 mm, the same size as the glue patches on the type tablet for Pilaeopsis radiatus) bearing an original type-set label, glued onto the shell, reading ‘105. P. radiatus Awatere’. It seems likely that this specimen is the holotype. The number ‘105’ is the species’ number in Hutton (1873b:14); the type face precisely matches the style, size and letter thickness of that in Hutton (1873b:14), and the glued label clearly was cut from an original printed copy of Hutton's text, as was also done for several other Hutton types still in GNS (but no other material I am aware of), strengthening the interpretation that this is the holotype. The shell is present on the remaining fragment so that the septum is not visible, and anyone not familiar with the sculpture of M. radiata would not immediately recognize this specimen as belonging in Maoricrypta, so it is possible that Hutton considered it to be a patellogastropod limpet. Hutton's (1973b:14) description reads ‘High, curved, with about twenty-four radiating ribs, which are much narrower than the intermediate spaces’, and this applies well to TM8581, except that it has only about 18 radial ribs (which, however, are narrow, closely spaced and difficult to count near the margins). Hutton cited only ‘Awatere’ as a locality. The block also contains fragments of Struthiolaria sp. similar to S. cincta Hutton, 1873, confirming the locality as likely to be late Miocene rocks of the Awatere valley. Apparently Marwick (Citation1948:30) did not notice that confirmation that this specimen is indeed the holotype is provided by Suter's (1914), pl. 1, ) illustration of this specimen, TM8581, as ‘Hipponix radiatus (Hutton). Holotype. Awatere’. Another clean, complete, smooth specimen of the same species was illustrated by Suter (Citation1914), pl. 1, a,b) as ‘Crepidula gregaria Sowerby. Plesiotype. Awatere’, so it is certain that Suter did not realize that these specimens are conspecific. TM8581 is accepted here as the holotype of Pilaeopsis radiatus.
Crepidula hochstetteriana, lectotype (selected here, Wilckens’ figured syntype, TM2608; F) and undiagnostic, fragmentary paralectotype from GS13, O32/f8025, Haumuri Bluff, Marlborough, internal moulds in beach concretions from the Miocene succession overlying the Cretaceous rocks (see below). Crypta turnialis Bartrum & Powell, holotype AUGD G5737 (F,K), with one paratype G5738, from Kaawa Creek, coast S of Waikato Heads, SW Auckland, Opoitian. Crypta opuraensis Bartrum & Powell, holotype AUGD G5739 (B,C), with one paratype G5740, from Kaawa Creek, coast S of Waikato Heads, SW Auckland, Opoitian. Maoricrypta haliotoidea Marwick, Citation1926, holotype TM8582, from GS1135, R19/f8007, Tirangi Stream, N Taranaki, Tongaporutuan; smooth and low, possibly an unusually shaped specimen of M. radiata, but probably distinct.
Distribution
Extremely abundant and widespread in all middle Miocene–early Pliocene (Tongaporutuan–Opoitian, and probably earlier) shallow-water rocks throughout New Zealand; usually found as piles or stacks of large specimens attached to each other. The taxonomy and phylogeny of early Miocene and older species in New Zealand is almost completely unstudied, but none of them formed obvious, large stacks.
Dimensions
Supposed ‘holotype’ of Pilaeopsis radiatus, TM8580: H 56.8, W not measurable, D 14.0 mm; true holotype of P. radiatus, TM8581: H (highly incomplete) 22.8, D 14.4 mm; lectotype of C. hochstetteriana: H 49.9, D 20.5 mm; holotype of C. incurva Zittel: H 42.5, W 25.5 mm; holotype of C. striata Hutton, NHMW 1959/335/30: H 25.6, W 18.3, D 8.1 mm (a very low specimen); holotype of C. haliotoidea, TM8582: H 39.0, W 25.3, D 14.7 mm; holotype of Crypta turnialis: H 31.6, W 19.8, D 11.6; holotype of Crypta opuraensis: H 23.5, W 15.0, D 9.1 mm; ‘Waipara’, N Canterbury, Suter Collection no. 558 (yellow sandy matrix; apparently from Double Corner Shellbeds, Lower Waipara Gorge, Waiauan): H (incomplete) 76.4, W 49.2, D 33.0 mm; GS1037, S28/f6446, Hurupi Stream, Palliser Bay, Tongaporutuan: H 52.5, W 30.2, D 20.7 mm; GS5485, S28/f6568B, Putangirua Stream, Palliser Bay: H 53.3, W 35.1, D 18.8 mm.
Remarks
Hutton's (1873b) decalcified supposed holotype of Pilaeopsis radiatus has a groove (a mould of the former septum) across the centre and clearly belongs in Maoricrypta, rather than being the patellogastropod limpet suggested by the generic name Pilaeopsis. This was first pointed out by Powell (Citation1924):283; quoting correspondence from J Marwick) although, as noted above, this is not the holotype selected by Hutton. The true holotype also clearly is a very incomplete specimen of a strongly sculptured specimen of Maoricrypta. Hutton (1973b:14) named Crypta striata at the same time, based on Zittel's (1864:44, pl. 15, ) illustrated specimen of ‘Crepidula sp.’, from rocks of the same age in the same area as Pilaeopsis radiatus (late Miocene, Upton Brook, Awatere valley). Smooth and radially ribbed specimens occur in the same stacks of specimens in Upton Brook (E) and these names are certainly synonyms. Crepidula incurva Zittel (1864:44, pl. 15, a,b) also was described from late Miocene rocks of the Awatere valley, and is again conspecific, but thinking that this name is a junior primary homonym of C. incurva Broderip, 1834, Finlay (Citation1924d:101) provided the replacement name Crepidula wilckensi for it. This was unnecessary, as Pilaeopsis radiatus Hutton is available for the same species. Marwick (Citation1948:30) provided a synonymy for M. radiata (although he did not include Suter's usage and illustration of Hipponix radiatus) and pointed out that ‘C. incurva Broderip’ was proposed in Calyptraea, but that Crepidula incurva Orbigny, 1841 is a senior primary homonym of C. incurva Zittel, 1864. King (Citation1934:23) thought M. striata a species separate from M. radiata, but Marwick (Citation1948:30) pointed out that this species is highly variable in sculpture, and M. striata is part of the variation of M. radiata. As first reviser, I select the name Pilaeopsis radiatus Hutton, 1873 as the valid name to be used for the species named both Pilaeopsis radiatus and Crypta striata by Hutton (1873b:14). This abundant Tongaporutuan–Opoitian (and earlier?—apparently at least Waiauan, possibly all middle Miocene) stack-forming species therefore should be known as Maoricrypta radiata, with synonyms C. incurva Zittel, 1864 (not C. incurva Orbigny, 1841), C. striata Hutton, 1873, C. wilckensi Finlay, 1924, and probably C. hochstetteriana Wilckens, Citation1922. M. haliotoidea Marwick (Citation1926:318, pl. 73, ) possibly is a further synonym, based on a low, wide specimen with its apex curled laterally more than usual. A specimen in GS1037, S28/f6446, from Hurupi Stream, Palliser Bay (Tongaporutuan) is shaped very similarly to the holotype of M. haliotoidea, and occurs with abundant specimens of normal M. radiata; it appears possibly to be merely a specimen of M. radiata that lived on a smooth, flat substrate rather than in a stack of other specimens. However, another specimen (GS11014, R19/f8059, stream 3.2 km N of Pohokura Saddle, 16 km W of Whangamomona, inland Taranaki; Tongaporutuan) is more complete, with a relatively tall, spirally coiled spire of one whorl and the septum commencing unusually well back in the shell, demonstrating that M. haliotoidea probably is a distinct, uncommon species. This has been deleted from the synonymy above, therefore. More complete specimens (again, with protoconch) are required to be certain of the status of M. haliotoidea. The protoconch of M. radiata is, as noted above, similar to that of M. costata (Marshall Citation2003, Fig. 25, 26) but has a markedly longer spiral suture visible on the protoconch, with a longer coiled spire.
Crypta turnialis and Crypta opuraensis of Bartrum & Powell (1928:144–145, pl. 27, Fig. 23–25) from Kaawa Creek were considered by Laws (Citation1936:110) and Grant-Mackie (Citation1990) to be conspecific, and Grant-Mackie (1990), as first reviser, selected C. turnialis as the senior synonym. The holotypes are similar in size and shape, but that of C. opuraensis (Bartrum & Powell 1928, pl. 27, Fig. 25; F,K) bears uneroded sculpture of low, narrow, closely spaced radial (that is, spiral) ridges, whereas that of C. turnialis (Bartrum & Powell 1928, pl. 27, Fig. 23, 24; A,B) is smooth because it is a severely abraded specimen, perhaps picked up after eroding out of the outcrop and being abraded on the beach; the paratype is very similar but narrower. The holotype of C. turnialis clearly bears a muscle scar on each side, interior to the end of the septum, confirming a position in Maoricrypta. Grant-Mackie (Citation1990, fig. 330–333) described and illustrated quite high stacks of specimens attached in life position to Glycymerita (Manaia) kaawaensis (Marwick, 1923) and a smaller stack attached to Austrofusa (Zelandiella) propenodosa (Bartrum & Powell, 1928). As Grant-Mackie (Citation1990) also described the very variable, relatively fine radial sculpture present on many Kaawa specimens, and all other Opoitian material I have seen from New Zealand includes specimens in life position in tall stacks, it seems likely that the Kaawa Creek specimens fall within the range of variation of M. radiata. Therefore, both Crypta opuraensis and C. turnialis are taken here to be further synonyms of M. radiata. A juvenile specimen from Kaawa Creek (G; AUGD G7130) examined by SEM has a large, inflated protoconch for a Maoricrypta species, 1.40 mm wide in total and 1.19 mm wide at the junction with the teleoconch, similar to that of M. costata (Marshall Citation2003, Fig. 25, 26) and the species illustrated by Marshall (Citation2003, Fig. 28), but with a larger initiation and longer and more evenly coiled suture than in those Recent species, and with no straight section after the coiled apex.
As noted previously by Saul & Squires (Citation2008:133), a palynoflora was examined from the matrix of Crepidula hochstetteriana Wilckens (Citation1922:5, pl. 1, a,b; F) recently by G Wilson (formerly of GNS pers. comm.), in a sample sawn off the slab bearing the lectotype of C. hochstetteriana designated here. He reported that it contains a middle to late Miocene dinoflagellate assemblage and palynoflora, including pollen of Acacia, a genus with an early Miocene to middle Pleistocene time range in New Zealand (Mildenhall & Pocknall Citation1989:61). The identification as Acacia was confirmed by JI Raine (GNS). This supposedly late Cretaceous species was based on material in pale grey concretions (rather than the dark grey to brown Cretaceous lithologies) from the late Oligocene to middle Miocene (Waitakian–Lillburnian) Waima Formation siltstone (Warren Citation1995:15–16) overlying the Cretaceous rocks at Haumuri Bluff (Warren & Speden Citation1977, map 1). M. hochstetteriana is, therefore, probably another synonym of M. radiata, which is not surprising as the lectotype and the fragmentary paralectotype are large internal moulds of an uncoiled Maoricrypta species resembling some Awatere specimens of M. radiata. However, checking of muscle scars is required on better specimens from middle Miocene rocks, as the type material does not rule out a position in Grandicrepidula, and there is some suggestion on the poor, partly recrystallized holotype mould that only the right muscle scar is present. Other late Cretaceous calyptraeids elsewhere in the world have not reached the evolutionary stage of Crepidula (Louella Saul, Los Angeles County Museum of Natural History pers. comm. 2006) so it is also not surprising from a phylogenetic viewpoint that C. hochstetteriana turns out to have been based on a Miocene specimen. A Miocene origin was pointed out earlier by me (Beu Citation1966) for Zenatia cretacea (Wilckens, Citation1922), another species supposedly from Cretaceous rocks at Haumuri Bluff. The middle Miocene Waima Formation at Haumuri Bluff must be kept in mind as the likely provenance of other supposedly late Cretaceous species described from Haumuri Bluff by Wilckens (Citation1922), but not subsequently re-collected from Cretaceous rocks.
Maoricrypta species, like all other calyptraeids, are sedentary filter-feeders living attached to hard objects, which in some cases include other Maoricrypta shells. Maoricrypta radiata is the middle Miocene (and earlier?) to Opoitian species that lived in large, narrow, irregular to evenly spiral stacks, with many large females at the base and making up much of the stack, males at the top, and individuals progressing through their sexual cycle as they become older through the upper part of the stack. It is the only New Zealand species I am aware of that formed high, obvious stacks in the same manner as the abundant North Atlantic species Crepidula fornicata (Linné, 1758) (Collin Citation2006). One stack of specimens (pointed out to me by JK Williams and VE Neall, Massey University) in Maoricrypta-Struthiolaria shellbeds in Matemateaonga Formation (Opoitian) east of Waiouru military camp, central North Island, formed a complete circle about 20 cm in diameter, and in cross-section resembled the nautiloid cephalopod Aturia. In contrast, all other New Zealand fossil species lived (and the Recent ones described by Marshall [Citation2003] all live) as isolated large females with one or a few much smaller males closely associated, males living on the shell of the female in many cases.
Maoricrypta youngi Powell, Citation1940 (E,J)
Crypta contorta. Hutton 1873b:14 (in part not C. contorta Quoy & Gaimard, 1835; Wanganui record only).
Maoricrypta youngi Powell Citation1940:232, pl. 33, , 13; Powell Citation1946a:75; Powell Citation1979:149, pl. 30, , 12; Marshall Citation2003:121, , 4, 7–9, 12, 27, 30, 70; Spencer et al. Citation2009:204; Maxwell Citation2009:242.
Crepidula youngi. Hoagland Citation1977:394.
Crepidula profunda. Beu & Maxwell Citation1990:259 (in part not Crypta profunda Hutton, 1873).
Type material
Maoricrypta youngi holotype AIM AK70473, from beach, Cape Maria van Diemen, N North Island (not seen).
Other material examined
Marshall (Citation2003:121) stated that Hutton's (1873b:14) specimen identified as ‘Crypta contorta’ from ‘Wanganui (U)’ is a specimen of Maoricrypta youngi. Other specimens occur in Upper Castlecliff Shellbed (OIS 11) on the Castlecliff coast. They are presumably among the specimens recorded as ‘Zeacrypta monoxyla (Less.)’ by Fleming (Citation1953:239) from GS4008, R22/f7396, and GS4185, R22/f6502, both on the Castlecliff coast, where I have observed them not uncommonly. Others are present in the GNS collection from GS4022 re-collection, R22/f6360, Pinnacle Sand (OIS 14) at head of ‘the pinnacles’ gully, Castlecliff (1; J) and GS4014 re-collection, R22/f6513, basal shellbed member of Shakespeare Cliff Sand, Castlecliff (one; G). Similar specimens from a few other units (e.g. GS4013, R22/f6353, Tainui Shellbed, OIS 13, at ‘the buttress’, Castlecliff, one specimen) appear to be M. youngi, but are not certainly distinguishable from weakly sculptured specimens of M. costata when their protoconch characters cannot be observed. Fleming (Citation1953:203, 218, 245) recorded specimens as ‘Zeacrypta monoxyla’ from a range of other late Castlecliffian units at Wanganui (Kupe Formation, OIS 17, two members; Lower Castlecliff Shellbed, OIS 15, GS4028, R22/f6366, coastal section; GS4104, R22/f6518, coastal section; GS4231, S22/f6454, Waikupa Road, off Fordell-Whangaehu highway, E of Wanganui; Pinnacle Sand, OIS 15–14, GS4098, R22/f6425, uppermost 23 cm in ‘the pinnacles’ gully, Castlecliff; Landguard Sand, GS4003, R22/f7394) and any or all of these might have been based on specimens of either M. youngi or M. sodalis Marshall, Citation2003 . The stratigraphic ranges of both these species are poorly constrained at present.
Distribution
Maoricrypta youngi was recorded from Wanganui (Castlecliffian) by Marshall (Citation2003:121), so the small, wide, almost smooth, strongly inflated specimens occurring moderately commonly in Upper Castlecliff Shellbed (OIS 11) and confused with M. profunda by Beu & Maxwell (Citation1990:259) belong in M. youngi. As M. youngi now occurs as far south as western Cook Strait (Marshall Citation2003:121), Castlecliff fossils are within the Recent range of the species. Castlecliff fossils I have seen are all small, strongly inflated and with deeply curved and embayed apertural planes, and apparently lived attached to the ubiquitous gastropod Maoricolpus roseus, cited by Marshall (Citation2003:122) as a common substrate for M. youngi.
Dimensions
NMNZ M.134496, 50 m, off Bream Tail, Northland: H 28.3, W 17.1, D 10.8 mm; NMNZ M.117767, 55 m, off Mahia Peninsula, attached to Charonia: H 22.5, W 16.8, D 9.3 mm; NMNZ M.15633, Falcon Shoal, Wellington Harbour, attached to Maoricolpus, closely resembling Wanganui fossils: H 25.4, W 15.0, D 9.1 mm; H 22.8, W 15.5, D 11.5 mm; H 18.0, W 12.7, D 8.6 mm; GS4022, Pinnacle Sand: H 15.2, W 11.2, D 7.4 mm; GS4014, basal shellbed of Shakespeare Cliff Sand; H 14.4, W 8.9, D 7.1 mm.
Remarks
Marshall (Citation2003:122) pointed out that Maoricrypta youngi differs from the superficially similar M. monoxyla in its brownish rather than white shell, its whitish rather than yellow animal, and its more strongly convex protoconch. It also differs from weakly sculptured specimens of M. costata (living specimens of which can occur alongside M. youngi) in its more paucispiral protoconch, and in the considerably broader start of the first whorl of the protoconch (width of exposed part 330–430 µm, compared with 230–270 µm in M. costata). The protoconch also is completely smooth, apart from commarginal growth lines, in M. costata (Marshall Citation2003, Fig. 25, 26), whereas it is initially smooth but then abruptly adopts crisp radial (spiral) sculpture in M. youngi (Marshall Citation2003, Fig. 27, 30). Their modern ranges and, to some extent, habitats are also distinct, as M. costata occurs at present only in the northeastern North Island, with one record as far south as NE Mahia Peninsula (Marshall Citation2003, ) whereas M. youngi occurs all around the eastern North Island, and through Cook Strait up the west coast as far north as Wanganui (Marshall Citation2003, ). M. costata also occurs attached to the undersides of rocks and to Perna canaliculus (Gmelin, 1791), and a few specimens occur alongside M. youngi on the undersides of Xenophora neozelanica Suter, 1908, whereas M. youngi has been found attached only to other gastropod shells. Marshall (Citation2003) listed as hosts for M. youngi living specimens of Astraea heliotropium (Martyn, 1784), Penion sulcatus (Lamarck, 1816), Maoricolpus roseus, Turbo smaragdus (Gmelin, 1791), Calliostoma (Maurea) granti (Powell, Citation1931) and X. neozelanica, and empty shells of Charonia lampas (Linné, 1758). A fossil specimen of M. costata from Wanganui (GS4014 re-collection, R22/f6513, basal shellbed of Shakespeare Cliff Sand, Castlecliff coast) is still in situ attached to the base of a specimen of Astraea heliotropium (B), showing that these two species can occur on the same substrate fairly frequently.
Genus Grandicrepidula McLean, Citation1995
Grandicrepidula McLean Citation1995:80. Type species (by original designation): Crepidula grandis Middendorf, 1849, Recent, boreal Pacific.
Remarks
Marshall (Citation2003:129) adopted Grandicrepidula for New Zealand species resembling Crepidula but with a single muscle scar visible, under the right edge of the columellar septum (as viewed in standard apertural view, with apex directed dorsally; this is also anatomically the right side of the living animal). No scar is present in most species of Crepidula, and two are present (one on each side) in Maoricrypta. Marshall also stated as a character of the genus ‘septal rim oblique relative to longitudinal axis of shell’, but this character does not hold for all New Zealand fossil species (as judged by the single muscle scar), as in the new species described below the edge of the septum is at right angles to the long axis of the shell. The teleoconch apex is weakly coiled to the right in all species I am aware of. Marshall (Citation2003:118) pointed out that Crepidula densistria Suter, Citation1917 (early Miocene, Oamaru area), Maoricrypta salebrosa Marwick, 1929 (Duntroonian, late Oligocene, Southland) and the Recent species G. collinae Marshall, Citation2003 should be referred to Grandicrepidula. The Oligocene and Miocene species have regular radial sculpture, unlike the smooth Pliocene and Recent species. Examination of GNS collections demonstrated that several other, unnamed species of Oligocene and Miocene age also belong here.
Grandicrepidula hemispherica n. sp. (B–E,G–I)
Type material
Holotype (TM8688; C,D,G,K) and four paratypes (TM8689–TM8692), GS10857, U23/f7049 (= GS10944, U23/f7498A), Maharakeke Mudstone (of Thomson Citation1926), 200–300 m along Maharakeke Road from Highway 2, at the E edge of the Takapau plain 1.5 km SW of Pukeora Hill, W of Waipukurau, S Hawke's Bay; occurring with the Pelicaria convexa form of P. vermis (Martyn, 1784), Austrofusus, taitae (Marwick, 1924), Bonellitia lacunosa (Hutton, 1885) and Paracomitas protransenna (Marshall & Murdoch, Citation1923), in a diverse, typical Hawke's Bay late Nukumaruan fauna. The type material was collected syntopically with specimens of Maoricrypta profunda.
Distribution
So far known only from the type locality.
Dimensions
Holotype: H 25.3, W 20.6, D 10.0 mm; largest paratype (B,E,H), TM8689: H 35.0, W 27.0, D 14.7 mm; paratype TM8690: H 26.0 (incomplete), W 24.5, D 10.2 mm.
Description
Shell moderately small for genus, smooth, highly inflated, wide (width 77–81% of length), with low, evenly inflated dorsum in small adults (height 40% of length, 49% of width in holotype) but slightly taller in large specimens (height 42% of length, 54% of width in largest specimen). Protoconch large (2.75 mm long on holotype), clearly demarcated from teleoconch by shallow groove on most specimens, rounded-triangular, with apex weakly inclined to left and curled under teleoconch; with low median external ridge in most specimens, nearer left than right edge of protoconch, providing triangular shape to whole protoconch; initial whorl coiled under, with initial protoconch suture located at edge of teleoconch and not visible in some specimens, scarcely visible above teleoconch edge in others, or located well above teleoconch edge and clearly visible in still other specimens; a few specimens with 1–3 very low, obscure, accessory ridges parallel to main central one. Teleoconch strongly and evenly inflated, with apex directed weakly towards right in small specimens but increasingly strongly towards right in larger ones, situated well above and slightly anterior to posterior margin in small, evenly oval specimens (3.7 mm above margin in holotype) but overhanging posterior margin slightly in largest specimen. Exterior smooth on smaller specimens, but with irregular, slightly scaly growth ridges on larger specimens. Aperture subcircular to evenly oval, apertural plane flat in largest specimen but weakly arcuate in smaller ones, indicating that small specimens live attached to dorsum of similar shells. Columellar septum occupying a little under half shell height, flat, with lightly and evenly curved anterior margin normal to long axis of teleoconch, curved around more strongly at margins to join side walls of shell; one obvious, small, moderately deeply impressed muscle scar beneath septum edge on right side, but none on left.
Remarks
Grandicrepidula hemispherica n. sp. is provided for nearly circular (when relatively young) to highly inflated, oval, smooth specimens of a previously unnamed species of Grandicrepidula from Maharakeke Road, W of Waipukurau, with the apex an unusually long way from the posterior end of the shell and a weakly triangular protoconch with a few unusual, low radial ridges and, in many specimens, no suture visible at all. The anterior edge of the columellar septum is aligned approximately at right angles to the long axis of the teleoconch. Most specimens with well-preserved protoconchs have a single prominent radial ridge extending from the visible apex to fade out at the end of the protoconch, producing a prominent, rounded-triangular shape for the protoconch as a whole, although a few specimens have weak subsidiary ridges parallel to the prominent one. The holotype has been chosen as a specimen that displays a single relatively prominent ridge and triangular protoconch shape. In some specimens the protoconch suture is aligned with the edge of the subsequent, enveloping teleoconch whorl, and scarcely visible or not visible at all, as in illustrations of the protoconch of Grandicrepidula collinae by Marshall (Citation2003, Fig. 23, 24), whereas in others the initial spiral section is slightly smaller, and a short but obvious suture is revealed on the side of the protoconch above the teleoconch margin, establishing that the extent to which the protoconch suture is revealed is quite variable in (at least some) Grandicrepidula species. Specimens of G. hemispherica n. sp. with the internal muscle scar visible under the anterior end of the columellar shelf display an obvious but relatively small, moderately deeply impressed, semicircular scar on the right side but none on the left, whereas one markedly larger but more shallowly impressed one is obvious on each side in specimens of M. profunda collected at the same locality (identified by their smaller, narrower protoconchs, straight shell apex, and narrower, ‘boat-shaped’ form, and the presence of weak to moderately prominent radial ribs on the exterior of the teleoconch). I have not seen the new species from elsewhere, but it is possibly the second species mentioned under Maoricrypta by Marshall (Citation2003) occurring at Mangahao, N Wairarapa, lacking a protoconch suture. However, it also seems feasible that several other Pliocene–early Pleistocene species of Maoricrypta and Grandicrepidula remain unrecognized in New Zealand, and certainly there are several unnamed Miocene and Oligocene taxa.
A few small specimens of Grandicrepidula hemispherica n. sp. have been collected attached to each other in small piles of 2–3 specimens, but tall stacks have not been seen at the one locality where it has been collected. Most smaller specimens (to about 25–28 mm long) have a weakly curved apertural plane, arched upwards across the central area and down at the anterior and posterior ends, suggesting that they have been attached to other specimens of G. hemispherica n. sp. In contrast, the largest specimen (H 35 mm) has a flat apertural plane and apparently lived attached to a flat surface, such as a rock or an oyster. Possibly this species changes it habitat as it grows. Of New Zealand calyptraeids, the protoconch of Grandicrepidula hemispherica n. sp. most nearly resembles that of G. collinae Marshall, Citation2003 in its lack of obvious fine sculpture, its subtriangular shape (much more marked in G. hemispherica than in G. collinae), weak radial ridge(s), and lack of a visible suture on many specimens. G. hemispherica n. sp. also differs obviously from G. collinae in its smaller size, more evenly inflated shape with a higher dorsum, more evenly subcircular to oval rather than subtriangular apertural outline, and the anterior margin of the columellar septum being normal to the long axis of the teleoconch rather than oblique. One of the most distinctive characters of G. hemispherica n. sp. is the position of the apex, which is well above and a short distance in front of the posterior margin in most specimens, and only overhangs the margin in the largest paratype.
It seems possible that Grandicrepidula hemispherica n. sp. occurs also in eastern Australia, in view of Marshall's (2003):118) statement that ‘all Australian crepiduliform species’ he had examined, other than Maoricrypta immersa (Angas, Citation1865), belong in Grandicrepidula. However, the possibility this provides of recognizing another warm-water migrant to New Zealand during late Pliocene–early Pleistocene time must await a taxonomic study of Australian calyptraeids, an almost completely unstudied group.
Etymology
Latin, hemispherical, half a sphere; intended to refer to the almost circular to evenly oval shape and high dorsum of this species.
Family Capulidae
Genus Zelippistes Finlay, 1926
Zelippistes Finlay Citation1926b:396. Type species (by original designation): Separatista benhami Suter, 1902, Pleistocene and Recent, New Zealand.
Remarks
There has been much confusion in the literature about the status of the superficially similar genera Lippistes Montfort, 1810 and Separatista Gray, Citation1847. To evaluate these genera and Zelippistes Finlay, 1926, I compared material of Lippistes cornu (Gmelin,1791) from South Africa, Separatista helicoides (Gmelin, 1791) from the tropical Pacific, and Zelippistes benhami (Suter, 1902) from northern New Zealand. The results are summarized here.
-
Lippistes cornu (A–C,E,F) and a second South African species (not L. grayi [A. Adams, 1850]; holotype examined [E,F], BMNH 198352,=L. cornu) are highly distinctive, planispirally coiled species (included by Bouchet et al. [Citation2005] in Hipponicidae) with distinctive, cap-shaped, paucispiral, planispiral protoconchs, reflecting lecithotrophic development. The whorl section on spire whorls is almost square, with numerous weak to prominent spiral cords around the outer surface. Specimens of L. cornu from eastern South Africa are evenly but loosely coiled, that is, all whorls are separated from each other evenly and regularly but narrowly, whereas specimens from western South Africa have all whorls closely attached to each other and have a more laterally expanded aperture; the two forms intergrade clinally around the coast. Lippistes is a narrow-whorled, planispiral genus restricted to South Africa.
-
Separatista contains two species of widespread tropical Pacific to southern Australian species of Capulidae resembling Trichotropis Broderip & Sowerby, 1829 and, in particular, Turritropis Habe, 1961 (Kantor & Sysoev Citation2006, pl. 27) in most characters, including its aperture shape, its low to moderately tall spire and its heavily hirsute periostracum, but consistently having three smooth spiral cords around the wide, flat, vertical periphery, and on many (but by no means all) specimens the last quarter to half of the last whorl tends to separate slightly to markedly from the previous whorl. A few specimens (such as the holotype of Zelippistes eccentricus Petuch, Citation1979) have the entire last whorl widely separated, so that it resembles that of Lippistes cornu, but all specimens intergrade. The protoconch is low-spired and paucispiral, with four raised spiral cords, significantly different from those of Lippistes and Zelippistes. The two species are S. helicoides (Gmelin, 1791) (D,H) (= Turbo separatista Dillwyn [1817:867; Wilson Citation1993:166, pl. 22, ],=Separatista chemnitzii A. Adams [1850b:45],=Trichotropis blainvilleanus Petit de la Saussaye [Citation1851:22, pl. 1, ; Macpherson & Gabriel, Citation1962:129, fig. 153; holotype examined in MNHN],=Trichotropis tricarinata Brazier [1878:313; Hedley Citation1901:126, pl. 17, , 23; holotype examined in AMS],=T. gabrieli Pritchard & Gatliff [Citation1899:183, pl. 20, ; Wilson Citation1993:166, pl. 22, a,b],=Lippistes meridionalis Verco [Citation1906b]:221, pl. 9, , 2] [note that Verco [Citation1906b]:220, pl. 9, –9] also recorded ‘Lippistes’ separatista [that is, Separatista helicoides] from nearby localities in South Australia, illustrating the radula and operculum, apparently not realizing that these forms intergrade],=Separatista fraterna Iredale [Citation1936]:293; holotype examined in AMS],=Zelippistes eccentricus Petuch [Citation1979]:5, –9; Poppe Citation2008a, pl. 98, ]; synonymy based on examination of most of the type material), occurring throughout the Indo-West Pacific to southern Australia; and S. gracilenta (Brazier) (Citation1878:313; holotype examined in AMS), a much smaller, narrower, taller-spired species with closely attached whorls, occurring in Queensland. The specimen identified as Trichotropis flavida Hinds, 1843 by Poppe (2008a), pl. 98, ) appears to be conspecific with Australian specimens identified as S. gracilenta, indicating an earlier name and a wide tropical Indo-West Pacific distribution for S. flavida. Separatista is a tall-spired genus with a weakly uncoiled last whorl, closely resembling Trichotropis and Turritropis, that occurs throughout the tropical Indo-West Pacific as far south as southern Australia. It is strongly distinct from Lippistes in size, shape, sculpture, protoconch characters and degree of uncoiling, although the small specimen illustrated here has a lower spire than most other specimens and does not appear as strongly distinct from L. cornu as larger, tall-spired specimens do (D,G; NMNZ M.246409, Onslow, Western Australia; H 6.9, D 10.9 mm).
-
Zelippistes benhami (F, A–D, A,C,E) is a further distinctive capulid that has evolved a laxly coiled last teleoconch whorl from Trichosirius independently in New Zealand. The last whorl is separated from the others similarly to that of some Separatista specimens and large western South African specimens of L. cornu, but this produces a more limpet-like adult. The moderately tall, smooth protoconch, with strongly inflated whorls, and tall early spire whorls are virtually indistinguishable from those of the type species of Trichosirius, T. inornatus (Hutton, 1873), and only the last whorl of Z. benhami adopts a wide, low, weakly uncoiled growth form. Most large specimens have the gap between the spire and last whorl filled by a flange, complex in many specimens, formed by folding the inner lip strongly back on itself, and the aperture and outer lip become irregular on most large specimens. Zelippistes benhami evidently is a Trichosirius relative that lives permanently attached to a hard substrate, and is more strictly sedentary than Trichosirius species. Zelippistes is an initially tall-spired but later weakly uncoiled, monotypic genus restricted to New Zealand. Its distinction from Trichosirius is a little tentative, requiring confirmation from anatomical and molecular studies.
Fig. 17 (A-C,E,F) Lippistes cornu (Gmelin), type species of Lippistes Montfort, 1810, South Africa; A,B, NMP E1147, De Hoop, E of Cape Agulhas, 50 m; A, protoconch, diameter 0.79 mm; B, whole shell of same specimen, partially uncoiled form, diameter 7.7 mm; C, NMP S8243, Aliwal Shoal, off Scottsburg, Natal, 10 m; juvenile, protoconch plus 0.5 teleoconch whorl; width 1.50 mm; E,F, largest syntype of Separatista grayi A. Adams, BMNH 198352, “South Africa”, the western tightly coiled form of L. cornu; H 7.4, D 14.8 mm. (D,G) Separatista helicoides (Gmelin), type species of Separatista Gray, 1847, NMNZ M.246409, Onslow, Western Australia; small specimen, H 6.4, D 10.9 mm.

Fig. 18 Zelippistes benhami (Suter). (A,D) GS4019, R22/f6358, Tainui Shellbed (Castlecliffian, OIS 13), Castlecliff coastal section, Wanganui; H 13.5, D 12.2 mm. (B,C) NMNZ M.84120, Recent, Hole in the Rock, Whangaroa, Northland; H 10.0, D 13.6 mm.

Fig. 19 (A,C,E) Zelippistes benhami (Suter); A, protoconch of specimen in Fig. 16F, SEM, dorsal view; C,E, NMNZ M.112179, Recent, 27-37 m, NE of Urupukapuka I., Bay of Islands, Northland, 34°14.10'S, 172°9.00'E, SEM; C, whole shell, diameter 8.4 mm; E, protoconch of C, lateral view. (B,D) Sabia australis (Lamarck), GS2784, T23/f6491, 3-10 m above Torlesse greywacke, Opawe Stream, Pohangina Valley, W Wanganui Basin, early Nukumaruan; B, abraded protoconch, SEM; D, whole shell, height 7.6 mm.

These three taxa have developed a shell form with a partly to completely uncoiled last whorl, reflecting a sedentary life habit, quite independently of each other, from distinct ancestors. This taxonomy will be reviewed elsewhere, and is merely summarized here to substantiate Zelippistes as a distinct genus.
Zelippistes benhami (Suter, 1902) (F, A–D, A,C,E)
Separatista benhami Suter Citation1902a:65, text fig.
Lippistes benhami. Suter Citation1913:297, pl. 15, .
Lippistes benhami var. perornatus Marshall & Murdoch Citation1923:121, pl. 15, , 7 (new synonym).
Zelippistes benhami. Finlay Citation1926b:396; Dell & Ponder Citation1964:64, –7, 11, 14; Powell Citation1979:148, Fig. 37, nos. 1, 2; Morley Citation1996:22–23, , 3; Spencer et al. Citation2009:204.
Zelippistes benhami perornatus. Dell & Ponder Citation1964:69, , 10, 12, 13; Fleming Citation1966:50; Beu & Maxwell Citation1990:408.
Zelippistes perornatus. Maxwell Citation2009:242.
Type material
Separatista benhami, holotype said by Suter (Citation1902a) to be in ‘Otago University Museum, Dunedin’, now in Otago Museum, Dunedin, IV7917 (Ilka Söhle, Otago Museum pers. comm. 3 March 2005), from beach, Cape Maria van Diemen, ‘found by Mr. Rayner, formerly lighthouse keeper at Cape Maria’ (Suter Citation1902a) (not seen). Lippistes benhami var. perornatus, holotype TM8295, from ‘blue clay, Castlecliff’, almost certainly from Tainui Shellbed, Castlecliff coastal section (OIS 13); with one paratype, TM8296. Marshall & Murdoch (Citation1923) mentioned that ‘there are two specimens, one in perfect condition, the other somewhat worn and the aperture imperfect’. They also illustrated the perfect one, referred to here as the holotype, and stated ‘Type to be presented to the Wanganui Museum’. (The combined collection of P Marshall and R Murdoch was purchased by the New Zealand Geological Survey from the Wanganui Museum in 1946, along with H Suter's collection.) The illustrated specimen is considered to be the holotype, because of Marshall & Murdoch's ‘type’ statement.
Other material examined
Castlecliffian: Tainui Shellbed (OIS 13), basal portion of Shakespeare Cliff, Wanganui City (GS4026, R22/f6364, one); Tainui Shellbed, Castlecliff coastal section (GS4097, R22/f6516, one); also three unlocalized ‘Castlecliff’ specimens in GNS, in a fine muddy matrix, probably from Tainui Shellbed; ‘Castlecliff, blue mud W. R. B. Oliver’ (NMNZ M.27300, one large, in a fine sandy mudstone matrix, formation uncertain, but apparently not from Tainui Shellbed; Pinnacle Sand?). Haweran: Landguard Sand (OIS 9), Landguard Bluff, east of Wanganui River (GS4003 re-collection, R22/f7394, one). Recent: 37 lots in NMNZ, ranging from King Bank, N of Three Kings Islands, to beach, Boulder Bay, Motuhora Island, off Whakatane, Bay of Plenty (NMNZ M.33073, three), from cast ashore on beaches to a depth of 710 m. Morley (Citation1996) also recorded specimens from the Three Kings Islands, Spirits Bay, the Bay of Islands and Goat Island Beach, Leigh.
Distribution
Apparently in OIS 13 and 9 only, at Wanganui (the same stages as Stephopoma roseum), but possibly more widespread stratigraphically, as several specimens lack detailed localities; Recent, N Northland to the Bay of Plenty; most Recent material is from the Three Kings Islands, the Bay of Islands and W Bay of Plenty.
Dimensions
Separatista benhami, holotype H 6.5, D 7 mm (Suter Citation1902a); Lippistes benhami perornatus, holotype H 9.6, D 13.4 mm; largest in GNS (unlocalized, ‘Castlecliff’): H (incomplete) 12.3, D 17.5 mm; GS4003 re-collection, Landguard Bluff: H 7.1, D 7.0 mm; large Recent shells: NMNZ M.84120, Hole in the Rock, Whangaroa, Northland: H 10.0, D 13.6 mm; M.21500, beach, Smugglers Bay, Whangarei Heads: H 9.5, D 18.5 mm; M.12134, fish stomach, Bay of Plenty: H 8.7, D 14.3 mm; M.75170, 25 m, outside entrance to Whangaroa Harbour: H 8.5, D 14.8 mm. Recent shells reach up to 21.5 mm in maximum dimension (Powell Citation1979:148).
Remarks
Dell & Ponder (Citation1964) maintained a subspecific distinction between Castlecliff fossils and Northland Recent specimens of Zelippistes benhami on the basis of the fossils having five spiral cords raised more than the others, whereas in Recent specimens they were all said to be subequal. Recent specimens in the large collection in NMNZ (37 lots) are highly variable in shape, coiling irregularity, and sculptural prominence, and there is no doubt that the trivial ‘forms’ Z. benhami and Z. benhami perornatus intergrade; several of the Recent specimens have spiral sculpture identical to that of fossils. Large specimens develop a strongly concave, shelf-like flange on the inner lip, where it bends back strongly over the columella to reach the previous whorl. Dell & Ponder (Citation1964) described the external appearance of the animal, the mantle cavity organs and the foregut of Z. benhami, including the style gland and stomach, and the conspicuous food groove extending from the ctenidium up the mantle cavity roof, to meet another groove running around the right cephalic tentacle, taking food to the proboscis. They also illustrated the short, disc-shaped, sucker-like foot, and commented that it ‘is probably little used for locomotion’ and ‘is almost certainly a sedentary animal’ (Dell & Ponder Citation1964:68), reflecting its filter-feeding habit and its sedentary habitat on cylindrical substrates. However, these aspects of the anatomy and ecology of Trichosirius inornatus have not been described, and need to be compared with those of Z. benhami.
One moderately large specimen of Zelippistes benhami in NMNZ (M.84119, trawled off Great Barrier Island, E of Auckland) was living attached to a black coral tree (Antipathes sp.) when collected. However, Morley (Citation1996:22) recorded a specimen collected living attached to the exterior of the tube of the polychaete worm Galeolaria hystrix, with its aperture near the tubeworm's aperture. Zelippistes, therefore, is the kleptoparasitic species it might be expected to be from its irregular shape and, in particular, irregular aperture, feeding by gathering food particles that escape the tubeworm's feeding. The specimen collected on Antipathes presumably was actually attached to a tubeworm growing on the Antipathes tree. The similarities between the teleoconchs of species of Lippistes, Separatista and Zelippistes no doubt result from them all independently adopting a kleptoparasitic habit on cylindrical hosts, probably tubeworms in all cases.
Zelippistes benhami clearly is another warm-water element in the Castlecliff fauna, a migrant (as planktotrophic larvae) from the northern North Island. Recent records are only from the northeastern North Island warm province. It is of interest, then, that fossil specimens have been collected recently only from Tainui Shellbed (OIS 13) at Castlecliff and from Landguard Sand (OIS 9) at Landguard Bluff, the two formations in Wanganui Basin from which the highest numbers of warm-water species are recorded (Beu Citation2004, Citation2006).
Family Hipponicidae
Genus Sabia Gray, Citation1841
Amalthea Schumacher 1817:56, 181 Type species (by subsequent designation, Gray 1847:157): Amalthea conica Schumacher, Citation1817, Recent, Indo-West Pacific (junior homonym of Amalthea Rafinesque, 1815).
Sabia Gray 1841:94 (no included species). Type species (by subsequent designation, Gray 1847:157, in synonymy of Amalthea): Amalthea conica Schumacher, Citation1817.
Saptadanta Prashad & Rao 1934:2. Type species (by original designation): Saptadanta nasika Prashad & Rao, 1934, Recent, Andaman Islands (= Sabia conica (Schumacher, 1817); see illustrations by Rajagopal 1977, pl. 9, –6).
Remarks
Higo et al. (Citation1999:112) stated that Sabia was first established by Gray (Citation1839), but I have been unable to find the name Sabia in Gray's (1839) work. A reference by Agassiz to its establishment by Gray (1839) was also mentioned by Sherborn (Citation1930:5708), who commented ‘sed non comp.’, that is, he also could not find the name in Gray's work. The first publication of the name, therefore, was by Gray (1841:94). Iredale (Citation1937:251) concluded that Sabia Gray (1841) is a nomen nudum, and that a description was first provided by Reeve (1842). However, Gray (1841:94) gave the following description: ‘In Capulus the foot is flat, with a plaited front edge; in Hipponyx and Sabia it is, as it were, folded on itself, and is unfit for walking upon; the back of the foot of the former of these animals secretes a shelly plate, marked with a horse-shoe shaped muscular impression, like an operculum; and of the latter forms a depression by corroding a space on the surface of the shell to which it is attached of the size of its own shell, and marked with a crescent-shaped ridge, shewing the place where the muscles were affixed; … ’. This is an available description of the new genus Sabia, distinguishing it from Hipponix Defrance, 1819, and the genus name Sabia clearly was made available by Gray (1841). Species were first included by Gray (1847), and there is nothing to indicate that Sabia Reeve (1842:34) was any more than a mention of the name established by Gray (1841); Reeve (1842) did not describe the genus or include any species in it. Iredale (Citation1937:251) also pointed out that Gray (1847:157) provided two type species selections for Amalthea, as he also did for Hipponix. Gray selected Amalthea conica Schumacher, 1817, the sole included species, as type species of Schumacher's (1817) ‘Amalthea α’, and A. ungaricus (Linné, 1758) as type species of ‘Amalthea β’. Ludbrook (Citation1957:49, pl. 4, , 2) and Knudsen (1991, c) both described and illustrated the holotype of Amalthea conica (ZMC 181, no. 1071), showing it to be essentially identical to the southern Australian species usually known as Sabia australis, although with a taller apex and correspondingly longer posterior slope than most Recent Australian specimens. Ludbrook (Citation1957:49) clarified the synonymy of this species and its confusion with Capulus danieli (Crosse, 1858) in South Australia (see Beu Citation2004:192). However, she assumed that Sabia australis is conspecific with Sabia conica, whereas I follow Knudsen's (1991) suggestion and keep these species separate at present.
Sabia australis (Lamarck, 1819) (B,D)
Patella australis Lamarck 1819, part 1, p. 335; Delessert Citation1841, pl. 23, a–c; Deshayes 1843:541 (transferred to ‘Pileopsis de la section des Hipponices’ in footnote).
Hipponix listeri Gray 1827:491.
Hipponyx australis. Deshayes 1832:274; Menke Citation1843:33; Menke Citation1844:61; Crosse Citation1862:21; Tryon 1886:136, pl. 41, –15; Tate 1893:330; Pritchard & Gatliff, Citation1900:198; Tate & May Citation1901:377; Odhner Citation1917:46.
Hipponix australis. Quoy & Gaimard 1835:434, pl. 72, Fig. 25–34; Knudsen Citation1991:642, a,b, 2–11 (with further synonymy, table 1); Wilson Citation1993:161, pl. 22, a–c.
Amalthea australis. H Adams & A Adams 1858:374, pl. 41, ; Watson Citation1886:457.
Capulus danieli. Angas 1865:175 (not Capulus danieli Crosse, 1858; Hedley Citation1902b:601).
Amalthea conica. Angas 1865:175; Tenison Woods 1877:38; Thiele Citation1930:580 (not A. conica Schumacher, 1817).
Pileopsis conica. Maplestone Citation1872:51, pl. 26, Fig. 28.
Capulus australis Hedley Citation1902b:600; Iredale 1924:245; Singleton 1937:392.
Capulus conica. Verco Citation1908:8; Verco 1935:8.
Hipponix conicus. Hedley Citation1916a:40 (in part); May 1921:57; May 1923:57, pl. 26, ; Macpherson Citation1958:30, pl. 26, ; Hartley Citation1958:37, pl. 8, –4; Laws Citation1970:115, –6; Spencer et al. Citation2009:205.
Sabia conica Cotton & Godfrey Citation1938:18; Ludbrook Citation1941:100; Kershaw Citation1955:313; Cotton Citation1959:365; Monger Citation1984:50, illus; Simone Citation2002:85, Fig. 36–38, 83, 83, 329–342.
Sabia australis. Macpherson & Chapple Citation1951:126; Ponder Citation1998:770, .123C,D.
Hipponix (Sabia) conicus. Ludbrook Citation1957:49, pl. 4, –4; Ludbrook Citation1978:123, pl. 13, , 2; Ludbrook Citation1984:82,234,268, o,p; 57t; 64l; pl. 5h.
Sabia wyattae Powell Citation1958b:90, pl. 12, , 4 (new synonym).
Capulonix australis. Iredale & McMichael Citation1962:44.
Hipponyx conicus. Macpherson & Gabriel Citation1962:127, fig. 152; Macpherson Citation1966:217.
Hipponix conicus wyattae. Powell Citation1979:145, pl. 30, , 16; Spencer & Willan, Citation1996:21.
Type material
Patella australis Lamarck, no type material present in MHNG, although a specimen (presumably a syntype) was illustrated by Delessert (1841, pl. 23, a–c), and Rosalie de Lamarck's annotations of Lamarck's copy of ‘Animaux sans vertèbres’ in MHNG indicate that Lamarck owned three specimens; the type material has been missing for many years (Y Finet MHNG pers. comm. 6 November 2007). The specimen illustrated by Delessert (1841, pl. 23, a–c) is apparently not conspecific or even congeneric with the interpretation adopted here, as (although clearly a hipponicid) it has prominent, extremely narrow radial ribs and very wide, flat interspaces, and so is not appropriate for lectotype designation. Further research is required on a possible neotype. Sabia wyattae, holotype AIM AK71238, with one paratype, AK71471, from beach at Tutukaka, Northland, living attached to Penion sulcatus (Lamarck, 1816) (not seen).
Other material examined
Nukumaruan: 3–10 m above Torlesse greywacke, Opawe Stream, Pohangina valley, E Wanganui Basin; early Nukumaruan, with Glycymeris shrimptoni, Myadora waitotarana, Clanculus plebejus, Diloma, Cantharidus, Austrolittorina and diverse Rissooidea (GS2784, T23/f6491, one specimen). Recent: NMNZ M.35225, off Poor Knights Islands, 110 m, 35°29′S, 174°43.5′E, RV ‘Acheron’ (one); M.34817, N of Three Kings Islands, 620 m, 34°01′S, 172°07′E, RV ‘Acheron’ (six); M.33748, off Three Kings Islands, 91 m, 34°11′S, 172°10′E, RV ‘Acheron’ (one); NMNZ M.287803, off Great Mercury I, 24 m (one).
Distribution
As interpreted by Knudsen (1991), Sabia australis is a species living along the coast of southern Australia, from southern Western Australia to southern New South Wales and around Tasmania. A few records (above) are known of empty Recent shells from NE New Zealand, besides the live-collected type material of S. wyattae, and a single fossil is recorded here from late Pliocene (early Nukumaruan) rocks of Wanganui Basin. Fossils were also recorded from the Pleistocene Ellen Point and Glanville Formations in South Australia by Ludbrook (Citation1984). Specimens on which Maxwell (in Campbell et al. Citation1994, table 4.30, p. 143) based his records of Hipponix from Whenuataru Tuff, Pitt Island, Chatham Islands, and that Laws (Citation1936:109) identified as Kaawa Creek specimens of Hipponix centrifugalis Marwick, Citation1931 are more nearly similar to tropical Pacific and eastern Australian material of Pilosabia trigona (Gmelin, 1791), and will be considered in a later paper. Pilosabia centrifugalis (Marwick Citation1931:97, pl. 8, fig. 139, 140), from Lillburnian (middle Miocene) rocks of the Gisborne district, has an unusual combination of relatively coarse radial sculpture over much of the shell, broken up by commarginal ridges towards the margin, and also seems more nearly similar to Pilosabia trigona than to S. australis. Hipponicidae are sedentary filter-feeders living attached to hard substrates, and in view of its severe facies restrictions, the time range in New Zealand of this shallow-water species may well have been much longer and more continuous than the Nukumaruan–Recent records suggest.
Dimensions
GS2784, Opawe Stream: H 2.8, D 7.6 mm.
Remarks
Usages of the name Hipponix (or Sabia) conica or Sabia australis for tropical Pacific records have not been included in the above synonymy, but the synonymy is still remarkably complex, reflecting long-continued doubts about the classification of and name for this species. It should be noted, though, that it would not be surprising if the correct name is eventually determined as Sabia conica; only molecular studies will resolve the taxonomy. It is important to note that this, rather than tropical material of S. conica, is the species identified by Simone (Citation2002:85, Fig. 36–38, 83, 83, 329–342) as Sabia conica in his anatomical evaluation of the Hipponicidae, in which he substantiated Sabia as a distinct genus (he examined material from Cockburn Sound, S Western Australia, and Green Cape, Maria Island, Tasmania). Laws (Citation1970) pointed out several differences in animal coloration, number of egg capsules, and number of larvae per capsule between South Australian specimens and the tropical Pacific specimens reported by Cernohorsky (Citation1968), supporting the distinction of S. australis from S. conica. This led Knudsen (1991) to suggest that S. australis should be maintained as a species separate from S. conica. However, whether this is correct is unknown, as the variation of these characters and whether there is any discontinuity or areas of syntopy in the distribution of Sabia species around the coast of Australia have not been studied; it is quite feasible that the distinguishing characters vary clinally, and all specimens are conspecific.
The specimen now recorded from early Nukumaruan rocks in eastern Wanganui Basin (from the same locality as the new record of Clanculus plebejus, above) is almost identical to the one illustrated by Ponder (Citation1998, .123C,D). Although the apical area has been abraded smooth, a protoconch closely similar to that illustrated by Knudsen (1991), b) is present. The exterior bears about 58 radial costae around the margin, including behind the apex, all relatively flat, wide and closely spaced, and separated by deep, linear grooves. This appears to be a typical specimen of the form identified by Knudsen (1991) as Sabia australis.
Powell (Citation1958b:90, pl. 12, , 4) proposed the name Sabia wyattae for Recent specimens found living on an empty shell of Penion sulcatus cast up on the beach at Tutukaka, NE North Island. Powell distinguished this form from S. australis only by its fewer radial costae, an exceedingly variable character in both S. australis and S. conica. Although no further records have been published, four lots from off northern New Zealand are present in NMNZ (listed above). The specimens illustrated by Powell and the NMNZ specimens are within the range of variation of S. australis as illustrated and described by Knudsen (1991), and indicate that S. australis occurs uncommonly in NE New Zealand. The New Zealand occurrences possibly are pseudopopulations recruited as planktotrophic larvae from southern Australia. If this is so, the single record of Sabia australis in Wanganui Basin early Nukumaruan rocks represents another case of a warm-water mollusc transported as larvae from southern Australia into Wanganui Basin, and it has certainly extended its range to Wanganui briefly from at least the northern North Island.
Family Triviidae
Genus Trivia Broderip, 1837
Trivia Broderip 1837:256. Type species (by subsequent designation, Gray 1847:142): Cypraea europaea Montagu, 1808 (= C. monacha Da Costa, 1778), Pliocene–Recent, NE Atlantic & Mediterranean (Iredale [Citation1912]:34] showed that Trivia was first made available, along with Gray's other manuscript names for Cypraeidae [sensu lato], by Broderip [1837] in his article on Cypraeidae in the ‘Penny cyclopedia’).
Subgenus Ellatrivia Iredale, Citation1931
Ellatrivia Iredale Citation1931:221. Type species (by original designation, Iredale Citation1931:233): Trivia merces Iredale, 1924, late Pliocene–Recent, central and N New Zealand and SE Australia.
Remarks
Several genera have been proposed in Triviidae, but their distinction is very subtle. Cernohorsky (Citation1971:111) referred all New Zealand Triviidae to Ellatrivia Iredale, Citation1931. Maxwell (1992:97) followed Beu & Maxwell (Citation1990:118) in referring all New Zealand Triviidae to Trivia, remarking that there do not seem to be any taxonomically important differences between Trivia monacha (Da Costa, 1778), the type species of Trivia Broderip, Citation1837, and T. pinguior Marwick, Citation1926 (late Eocene, S Canterbury–N Otago). He maintained this position recently (Maxwell Citation2009:242). In contrast, Dolin (Citation2001) recognized Trivia (Trivia), T. (Decoratitrivia), Dolichupis, Trivellona, Niveria, N. (Ellatrivia), Trivirostra, Cleotrivia and Semitrivia as generic groups within Indo-Pacific Triviidae. They are separated by subtle differences in the shape of the basal callus, apertural canals and dorsum, in the prominence of the dorsal sulcus, and in the shape and sculpture of the fossula, a concave columellar area inside the aperture, parallel to the inner lip. Dolin (Citation2001:218) referred the New Zealand and SE Australian shallow-water triviids to Niveria Jousseaume (Citation1884:100) (type species: Cypraea nivea Gray in GB Sowerby I Citation1832 (junior primary homonym of Cypraea nivea Wood, Citation1828; = Niveria nix Schilder, 1922), Recent, western Atlantic), subgenus Niveria (Ellatrivia), expressly including Trivia kaiparaensis Laws, 1939 (early Miocene, Pakaurangi Point, Kaipara Harbour, Northland). However, Fehse (in Poppe Citation2008a:668–681) used only Trivia, Cleotrivia, Dolichupis, Trivellona and Trivirostra for Philippine Islands Triviidae, most of which were assigned to either Trivirostra or Trivellona. Several of the species proposed by Dolin (Citation2001) were assigned to Trivellona by Fehse (in Poppe Citation2008a). All taxa illustrated by Fehse (in Poppe Citation2008a) have either more protruding anterior and posterior extremities or more prominent exterior ribs than in the New Zealand–SE Australian species considered here. The classification of triviids is obviously undecided, differing between authorities. In the circumstances, it seems best to follow Spencer et al. (Citation2009) and retain the usage of the subgenus Trivia (Ellatrivia) for New Zealand species. Trivia zealandica Kirk, Citation1882, discussed below, differs from all other New Zealand triviids in lacking transverse ridges over the dorsum—the very short ridges stop at the dorsal edges of the narrow lateral callus—but the taxonomic significance of this character also is unclear. Illustrations of Trivellona suiduirauti (Lorenz, 1996) by Fehse (in Poppe Citation2008a, pl. 282, a,b, 6a,b) show that the ribs are reduced to faint angulations on the dorsal surface of this species, but do not fade out completely as in T. zealandica. The aperture is also considerably wider and the spire protrudes more in T. suiduirauti than in T. zealandica. The position of the dorsal sulcus (an antero-posterior groove between the dorsal ends of the transverse ridges on some of the more strongly sculptured species) is clearly shown by a smooth area between the surface granules in well-preserved specimens of T. zealandica, and it seems to be a normal Trivia (Ellatrivia) species in all characters other than the length of the ridges. It is assumed here that the loss of dorsal ribs is merely a continuation of the trend towards weakening of the dorsal ribs already seen in such Recent species as T. (Ellatrivia) merces (Iredale, 1924), and Trivia zealandica also is referred to Trivia (Ellatrivia).
Trivia (Ellatrivia) zealandica (Kirk, 1882) (A–C)
Fig. 20 (A-C) Trivia (Ellatrivia) zealandica (Kirk), O32/f8875, Leader siltstone lithofacies (early Castlecliffian, OIS 31 or earlier), Leader River, Parnassus, N Canterbury; height 12.8 mm. (D-F) Trivia (sensu lato) sp., cf. Trivellona paucicostata valeriae (Hart), GS15629, R28/f078, Onoke Formation (early Nukumaruan), bathyal fauna E of Lake Ferry, Palliser Bay; height 9.4 mm. (G-I) Trivia (Ellatrivia) merces (Iredale), abraded specimen, GS689, Y19/f9491, uplifted Holocene terrace, N shore Mahia Peninsula; height 11.6 mm.

Trivia zealandica Kirk 1882:409; Suter Citation1915:11, pl. 8, a,b; Beu & Maxwell Citation1990:351, pl. 47m, q; Maxwell Citation2009:242.
Trivia neozelanica (sic). Hutton 1893:58 (error for T. zealandica Kirk).
Trivia flora Marwick Citation1928:182, fig. 118, 119; Fleming Citation1966:52, fig. 1071, 1072; Beu & Maxwell Citation1990:408; Maxwell Citation2009:242 (new synonym).
Niveria (Ellatrivia) flora. Schilder Citation1941:75.
Ellatrivia zealandica. Fleming Citation1966:52; Cernohorsky Citation1971:113, .
Ellatrivia flora. Cernohorsky Citation1971:113.
Type material
Trivia zealandica, lectotype (of Suter Citation1915:11, pl. 8, a,b) NMNZ M.1815, an incomplete shell, with one paralectotype NMNZ M.1816 (Marshall Citation1996:21), from ‘Petane’, that is, Tangoio Limestone or Mairau Mudstone, formations high in Petane Group (late Nukumaruan) in Matapiro Syncline, 30 km N of Napier, central Hawke's Bay. Marshall (Citation1996:21) discussed the complex history of the type material, and concluded that Cernohorsky (Citation1971:113) was incorrect to regard Suter's (1915) selection as designating a neotype. Trivia flora, holotype TM8575, from Whenuataru Tuff (Waipipian, and perhaps in part Mangapanian), Flowerpot Harbour, Pitt Island, Chatham Islands.
Other material examined
Waipipian: GS12162, CH/f20A, Whenuataru Tuff, E side Flowerpot Bay, Pitt Island (four); GS12163, CH/f13B, Whenuataru Tuff, Motutapu Point, Pitt Island (one fragment); GS12164, CH/f25, Whenuataru Tuff, tip of Taruwhenua Peninsula, Pitt Island (one). Nukumaruan: specimens of Trivia (Ellatrivia) zealandica occur uncommonly in the ‘Petane beds’ (late Nukumaruan formations of Petane Group in Matapiro Syncline, central Hawke's Bay; Beu Citation1995:117–135) in eastern North Island (Okawa Stream, GS1063, V21/f8476, two specimens; Puketapu, Matapiro Syncline, NW of Napier, GS5216, V20/f8517, one; walkway on W side of Roro-o-kuri, Onehunga Road, Ahuriri ‘lagoon’, Napier, Mairau Mudstone, GS12386, V21/f05, three; ‘the watchman’ hill, Ahuriri Lagoon, Napier, Mairau Mudstone, GS11465, V21/f8015A, one; Flat Rock, Tangoio, calcite neomorph fauna in Tangoio Limestone, GS12392, V20/f07, one).
Trivia (Ellatrivia) zealandica also occurs rarely in early Nukumaruan limestone at Castlepoint, eastern Wairarapa, in rudstone beds (Van der Lingen et al. Citation1985) with common Chama ruderalis (Lamarck, 1819) (= C. huttoni Hector, Citation1886, type locality; Beu Citation2006), alternating with floatstone beds containing the cold-water scallop Zygochlamys delicatula (Beu Citation1995:156) (GS10844, U26/f6037). Van der Lingen et al. (1985) described and illustrated complex liquefaction deformation structures in the floatstone component and low-angle cross-stratification in the interbedded rudstone, in a sequence of two or three sedimentary cycles (stratigraphic column: van der Lingen et al. 1985, ). Beu & Kitamura (Citation1998:146) suggested that deposition of Castlepoint Formation occurred as deep-water, canyon-head debris flows in response to cyclic sea-level changes on the adjacent shelf. It seems likely that the floatstone component was deposited as near-in situ sediment in the canyon head, and the rudstone beds represent coarser material slumped into the site as a result of sediment accumulation on the shelf around the canyon head. Loading of floatstone by the rudstone component and slumping of the entire mass further into the canyon head as a result of sea-level lowering, and perhaps coseismic shaking, would then have produced the liquefaction structures.
In Wanganui Basin, Trivia (Ellatrivia) zealandica has been collected only from early Nukumaruan mudstone a short distance above Hautawa Shellbed, at the disused Hautawa Road, off West Road, north of Hunterville, Rangitikei valley, and from Tewkesbury Formation ca 200 m W of the mouth of Ototoka Stream, Nukumaru Beach, in the Wanganui coastal section (specimen in the collection of Mrs Julie Loveridge, Waitotara). The only other record I am aware of from outside Hawke's Bay is in NMNZ: M.287951, head of gully 800 m S of Pahiatua Trig., Konini, Pahiatua, N Wairarapa (four). Castlecliffian: Castlecliffian specimens are recorded only from eastern New Zealand: Leader River, ‘Mendip Hills’ Station, Parnassus, North Canterbury, where it underlies Potaka Tephra (Warren Citation1995, , 21), and so is older than 1.0 Ma (Shane Citation1994; Shane et al. Citation1996), that is, approximately OIS 29–31 or older (O32/f8875; Beu & Maxwell Citation1990:352, pl. 47m, q; A–C); and Matata and Ohope Beach, Whakatane, Bay of Plenty (OIS 15?): NMNZ M.96442, large bend on Wainui Stock Road, south of Ohope Beach (one); NMNZ M.42960, shellbeds below old reservoir in stream at foot of Whakatane-Ohope Road (one + five fragments); NMNZ, locality F518, Ohinekeao Stream, W of Matata (one fragment).
Distribution
Waipipian (–Mangapanian?), Whenuataru Tuff, Pitt island; Early Nukumaruan, just above Hautawa Shellbed, Wanganui Basin (one specimen), at Castlepoint, East Wairarapa, and near Pahiatua Trig., Konini, N Wairarapa; late Nukumaruan, Wanganui Basin (rare) and Hawke's Bay (uncommon); early Castlecliffian, Mendip Hills station, Leader River, North Canterbury (OIS 29–31 or older), one specimen (A–C); later Castlecliffian at Matata and Ohope Beach, Whakatane (OIS 15?).
Dimensions
O32/f8875, Leader River, Parnassus, early Castlecliffian: H 12.8, D 9.4, H (normal to aperture) 7.4 mm; GS5216, Puketapu, Hawke's Bay: H 11.5, D 8.0, H (normal to aperture) 6.5 mm; GS12386, Roro-o-kuri, Napier: H (incomplete) 11.4, D 9.8, H (normal to aperture) 7.6 mm; holotype of T. flora: H 10.0, D 7.3, H (normal to aperture) 6.0 mm (Marwick Citation1928:482).
Remarks
Trivia (Ellatrivia) zealandica is rendered very distinctive, compared with all the other New Zealand species of Triviidae, because the narrow ridges on the exterior extend only to the dorsal margin of the lateral callus areas, and the superficially smooth, microscopically granulose dorsum is clearly slightly conical in shape. The lateral callus areas also are narrower and more prominent than in other New Zealand species. It is also one of the larger triviids recorded from New Zealand, reaching 13 mm high (specimens of E. merces reach 15.3 mm high: NMNZ M277017, beach, Reef Point, Ahipara, N Northland, H 15.3 mm; NMNZ M277019, beach, Cape Maria van Diemen, Northland, H 15.2 mm; NMNZ M277015, beach, Taupiri Bay, Northland, H 14.9 mm).
It is not quite certain that the relatively small specimen (height 8.4 mm) from ‘Petane’ illustrated by Cernohorsky (Citation1971, ) as Trivia (Ellatrivia) zealandica is conspecific with the significantly larger specimens that usually bear this name. Cernohorsky's figure shows the narrow ridges extending a significant distance up the sides of the dorsum above the lateral callous ridges, whereas they stop at the dorsal edges of the callous ridges in almost all specimens I have examined (but note the comment below on Trivia flora Marwick, Citation1928). However, the specimen agrees in shape, in its narrow aperture, and in its conical dorsum with T. zealandica, and Cernohorsky (1971) described the dorsum as ‘distinctly finely granulose’, so it evidently is a slightly unusual specimen of T. zealandica.
Marwick (Citation1928:428) distinguished Trivia flora, from the Pliocene of Pitt Island, Chatham Islands, from T. zealandica by the ‘bluntly conoid’ rather than rounded shape of the dorsum, and by being ‘not so attenuated anteriorly’. However, the few specimens collected since and the holotype seem indistinguishable from T. zealandica. The shape of the dorsum is identical in specimens from the Chatham Islands and mainland New Zealand, Whenuataru Tuff specimens are only slighty smaller than the largest Hawke's Bay Nukumaruan specimens of T. zealandica, and the shape of the anterior seems within the range of variation of one species. One of the Pitt Island specimens has the narrow ridges extending slightly further up the sides than I have seen in Nukumaruan specimens, slightly above the lateral callous ridges, but the holotype and all othe specimens examined agree with the common form of T. zealandica in this character, and the specimen of T. zealandica illustrated by Cernohorsky (Citation1971, ) has the ridges extending further dorsally than on any Pitt Island specimens I have seen. In my opinion T. flora is simply a synonym of T. zealandica. Therefore, T. zealandica is a considerably longer-ranging species than has been thought previously.
Cernohorsky (Citation1971:115, , 10a) recorded Trivia (Ellatrivia) merces also as a fossil at ‘Petane’ (that is, from a late Nukumaruan formation of Petane Group in the Matapiro Syncline in central Hawke's Bay), on the basis of a single specimen in the Auckland Museum, but I have not seen specimens from Hawke's Bay. The other fossils of T. merces I am aware of are from Te Piki, Cape Runaway (OIS 7) (Grant-Mackie & Chapman-Smith Citation1971:673; Richardson Citation1997:8), and Denby Shellbed (OIS 7) at Waihi Beach, Hawera (Thomson Citation1917:417, as Trivia zealandica; Fleming Citation1953:271). A single specimen (G–I) is also present in GNS collections from a Holocene terrace at Mahia Peninsula (GS689, Y19/f9491, ‘raised beach, north shore’, collected by Alexander McKay), and a fragment is present in a collection from the highest Holocene terrace at Aramoana Beach, S Hawke's Bay coast SE of Waipukurau (V23/f61, 200 m NE of Gibralter Road, V23/372112). T. merces has, therefore, a Nukumaruan–Recent time range, but clearly has a highly unreliable fossil record and is not useful for biostratigraphy, whereas T. zealandica is known in post-Nukumaruan rocks only at Parnassus and Ohope Beach, and has some utility in biostratigraphy.
A further, apparently undescribed early Nukumaruan fossil triviid is represented by a single specimen (D–F) from the bathyal fauna (Beu Citation1967, Citation1969) in Onoke Formation mudstone (early Nukumaruan) in the cliffs east of Lake Ferry, Palliser Bay, collected and presented by Sashia Wood (GS15629, R28/f078). It resembles Trivellona paucicostata valeriae (Hart Citation1996; Dolin Citation2001:219, , 13, 47) in many characters, having relatively sparse, prominent riblets passing across the dorsum with little interruption, that is, longer than in both Trivia (Ellatrivia) zealandica and Trivia merces, the two more common species in New Zealand. Dolin (Citation2001) pointed out that ‘Erato’ tetatua Hart, Citation1996 is based on an immature specimen of Trivellona paucicostata valeriae Hart. The Palliser Bay shell differs from Trivellona paucicostata valeriae in its smaller size, its more elongate shape, and in its fewer, narrower, more widely spaced dorsal ridges, interrupted by a narrow sulcus in the centre of the dorsum. The fossula is scarcely visible, as the specimen was cracked and has been filled with glue. Further specimens are needed to clarify the identity of the Lake Ferry species.
Family Naticidae
Genus Taniella Finlay & Marwick, Citation1937
Taniella Finlay & Marwick Citation1937:48. Type species (by original designation): Natica notocenica Finlay, 1924, early Miocene, New Zealand.
Remarks
Taniella differs from Tanea Marwick, Citation1931 (type species: Natica zelandica Quoy Gaimard, 1835, late Pliocene–Recent, New Zealand) in its significantly smaller size, its markedly lower spire in most specimens, and its straight spire outlines, with a weakly impressed, tangential suture. The apertural characters, including a prominent funicle, are similar in both groups. The relationship between Tanea and Taniella is unclear because Taniella apparently is extinct, and radular characters are critical to understand naticid relationships. The operculum of Taniella notocenica (Finlay, 1924) was described by Finlay & Marwick (Citation1937:48) as ‘almost flat, exteriorly with a thickened, medial concentric [that is, commarginal] ridge (as in many Astraea) the surface inside this smooth, the surface outside finely but conspicuously granulate with traces of numerous subsurface spirals, ending as well-marked granulations on the inner posterior margin; no trace of marginal grooves’. This differs markedly from the operculum of Tanea zelandica, with its two conspicuous marginal grooves and margining ridge, and suggests that Taniella and Tanea are not closely related. Finlay & Marwick (Citation1937) suggested a close relationship between Taniella and Tectonatica Sacco, 1890 (type species: Natica tectula Sacco, 1890, Pliocene, Italy), and the similarity is great enough that generic distinction is uncertain (the distinction was probably based mainly on Finlay's conviction that New Zealand taxa are all restricted endemics). Tectonatica rizzae (Philippi, 1844) (Bouchet & Warén Citation1993:764, fig. 1822, 1823) is very similar to Taniella species in size (8.1–10.6 mm high), its shape, its low spire with straight outlines, its tangential, flat suture, and its obvious umbilical callus (funicle) that leaves a surrounding groove. However, the juvenile illustrated by Bouchet & Warén (Citation1993, fig. 1879) has two narrow but obvious marginal opercular grooves around the margin against the outer lip, reduced in the adult to one groove (Bouchet & Warén Citation1993, fig. 1908). Tectonatica tecta (Anton, 1838) (Recent, Namibia to E Cape Province, South Africa; neotype illustrated by Kilburn Citation1976, b, 8b, 9) has an essentially smooth operculum, apart from a weak callus ridge in the nuclear region, and has a weakly sinuous spire outline similar to that of the tall-spired specimens of Taniella planisuturalis described below. Having none or one rather than two marginal opercular grooves seems to distinguish Tectonatica from Taniella. Also, both T. rizzae and juvenile specimens of T. filosa (Philippi, 1845) (Bouchet & Warén Citation1993, fig. 1880) have a conspicuously thickened umbilical callus at a very small size, the umbilicus is completely filled with callus at all growth stages in T. tecta, and the columellar margins of the opercula of T. rizzae and T. filosa bear obvious transverse ridges, transformed a little in the adult (Bouchet & Warén Citation1993, fig. 1906, 1908) into short ridges down the adapical third of the columellar margin. Taniella seems best retained as a distinct genus, although probably closely related to Tectonatica phylogenetically.
Finlay & Marwick (Citation1937:49) referred Natica subnoae Tate, Citation1893 (Janjukian, Oligocene; Bird Rock, Torquay, Victoria) to Taniella, ‘the only Australian species at present known’. Darragh (Citation1970:197) followed their suggestion. Wilson (Citation1993:218–219) referred several Indo-West Pacific and Australian species to Tectonatica, but most have a significantly taller spire than Taniella notocenica and T. planisuturalis. One Recent species that seems very similar to T. planisuturalis is Natica elkingtoni Hedley & May (Citation1908:119, pl. 23, ) from moderately deep water (180 m) off Cape Pillar, S Tasmania. The spire of N. elkingtoni is slightly taller than that of most specimens of T. planisuturalis, despite all other characters being similar, but that is also true of the New Zealand Miocene fossil species assigned to Taniella, and of a few large specimens of T. planisuturalis described below from shallow-water environments. A close phylogenetic relationship between New Zealand Taniella species and Australian or tropical Pacific species remains possible, but requires further research. If a position in Taniella could be confirmed for N. elkingtoni, it would allow the radula and DNA of Taniella to be assessed, and it would not be surprising if Taniella is a synonym of Tectonatica.
Taniella planisuturalis (Marwick, 1924) (B, G)
Fig. 21 (A) Eunaticina papilla (Gmelin), holotype of Sigaretus (Naticina) cinctus Hutton, CMC M3014, Landguard Sand (Haweran, OIS 9), Landguard Bluff, Wanganui; H 14.8, D 12.8 mm. (B,G) Taniella planisuturalis (Marwick), 2 large, tall-spired specimens, GS11225, V20/f8002, Darkys Spur Formation (late Nukumaruan), Darkys Spur, W of Devils Elbow, Hawke's Bay; B, H 16.8, D 13.7 mm; G, H 18.2, D 15.3 mm. (C-F,H) Semicassis labiata (Perry); C,D, lectotype of Cassis achatina Lamarck and neotype of Cassidea labiata Perry, 1811, MHNG 1100/99; unlocalised (type locality Port Jackson, Sydney, Australia); height 53 mm; E,F, 2 syntypes of Cassis zeylanica Lamarck, unlocalised (South Africa); E, MHNG 1101/4, ventral view, height 81 mm; F, MHNG 1101/3, dorsal view, height 76 mm; H, GS5833, Y14/f7505A, Te Piki Member (Haweran, OIS 7), Te Piki, near East Cape; height 31.3 mm.
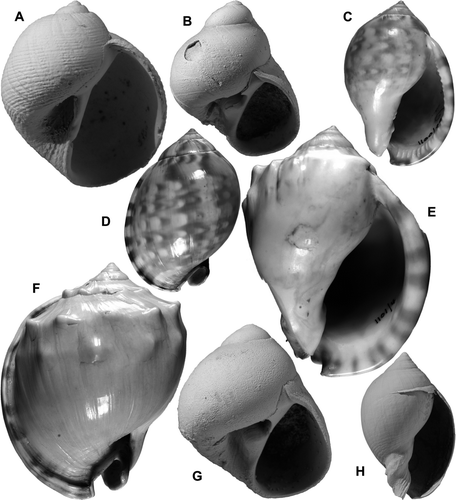
Natica planisuturalis Marwick Citation1924c:550, pl. 55, , 11.
Natica haweraensis Marwick Citation1924c:551, pl. 55, , 7.
Taniella planisuturalis. Finlay & Marwick Citation1937:49; Fleming Citation1966:53, fig. 1987, 1988; Beu & Maxwell Citation1990:321, pl. 41n; Maxwell Citation2009:242.
Taniella haweraensis. Finlay & Marwick Citation1937:49; Fleming Citation1966:53, fig. 1084, 1085.
Type material
Natica planisuturalis, holotype TM6888, from GS1089, V21/f8482, ‘blue clays’ (Okauawa Formation), Okauawa Stream, Kereru Road, inland central Hawke's Bay (late Nukumaruan, early Pleistocene). Natica haweraensis, holotype TM6887, from GS1173, Q21/f6495, Tangahoe Formation, Waihi Beach, Hawera, South Taranaki (Waipipian, mid-Pliocene).
Other material examined
Opoitian: GS8101, Y14/f7618A, Oweka Creek, halfway between Whangaparaoa and Hicks Bay, East Cape (four); GS5626, C46/f9594, Birch's Mill shell lens, Te Waewae Bay, Southland (five); Kaawa Creek, SW Auckland (recorded by Laws (Citation1936, list p. 99) as ‘Tanea notocenica (Finlay)’, the early Miocene type species of Taniella). Waipipian: occurs at Waihi Beach and Waipipi; recorded by Fleming (Citation1953:112) from Middle Waipipi Shellbed (GS1101, Q22/f7303). Mangapanian: recorded from Mangapani and Wilkies Bluff by Laws (1940b: 38); recorded by Fleming (Citation1953:119,120,124) from Mangapani Shellbed (GS4227, R21/f8478), Makokako Sand (GS4221, R21/f9504) and Wilkies Shellbed (GS4124, R22/f6543; GS4224, R22/f6467). Nukumaruan: Fleming (Citation1953:131, 139, 144, 151) recorded Taniella planisuturalis from Hautawa Shellbed, Nukumaru Limestone, Nukumaru Brown Sand, and Waipuru Shellbed in Wanganui Basin. Scott (in Kingma Citation1971, table 10A) also recorded it from six localities in Petane Group in Te Aute Subdivision, but it is much more common and ubiquitous in mudstone and sandstone formations of Petane Group than this record suggests. Castlecliffian: Fleming (Citation1953:174, 181) recorded T. planisuturalis from Butler's Shell Conglomerate (early Castlecliffian, OIS 31; GS4083, R22/f6413, Ototoka coast section, listed in parentheses to indicate thought reworked from older rocks), and Okehu Shell Grit (Castlecliffian, OIS 27; GS4072, R22/f6403, road from Kai-Iwi Station to beach, again in parentheses). Fleming (in Kingma Citation1971:98) also recorded T. planisuturalis from Maraetotara Sand (early Castlecliffian, from shellbeds underlying Potaka Tephra, ca OIS 29–31) at Cape Kidnappers, Hawke's Bay (GS5313, W21/f8520, bend in tributary of Maraetotara River to NW of high terraces; GS5309, W21/f8516, lowest shellbed, 1.6 m above unconformity, coast W of Black Reef). BA Marshall's collections from Ohope Beach, Whakatane (Castlecliffian, about OIS 15?) in NMNZ include the following records of Taniella planisuturalis: M.40267, cutting on Wainui Stock Road, SW shore Ohiwa Harbour (two); M.43035, shellbeds below old reservoir in stream at foot of Whakatane-Ohiwa Road, Ohope (six); M.95323, large bend on Wainui Stock Road, S of Ohope Beach (23).
Distribution
Opoitian, Kaawa Creek, Oweka Creek and Te Waewae Bay, probably more widespread; Waipipian–Nukumaruan and, probably, early Castlecliffian in Wanganui Basin (assuming some OIS 31–27 records are not reworked); up to early Castlecliffian in eastern New Zealand, at Cape Kidnappers (OIS 31–29?) and mid-Castlecliffian at Ohope Beach, Whakatane (OIS 15?). Widespread and common throughout New Zealand Pliocene rocks. Taniella planisuturalis evidently retreated gradually from central New Zealand to the north as temperatures fell after Nukumaruan time, surviving in New Zealand until about OIS 15 in the Bay of Plenty, that is, in the northeastern New Zealand warm province. The only other examples with extensive enough fossil records to reveal such a gradual retreat are (1) Arca cottoni Waghorn, 1926 which, as noted by me (Beu Citation2004:133), is also recorded from Castlecliffian (OIS 15?) rocks at Ohope Beach, Whakatane, but at no other post-Nukumaruan localities in New Zealand, and (2) Trivia (Ellatrivia) zealandica, which also is recorded above from OIS 15 rocks at Whakatane. Arca cottoni might well be a synonym of one of the living tropical western Pacific species (Beu Citation2004:133).
Dimensions
Marwick (Citation1924c:550, 551) stated the dimensions of the holotypes as: N. planisuturalis, H 11, D 11 mm; N. haweraensis, H 10, D 10 mm; he also stated that ‘an imperfect specimen from Waipipi Beach is 20 mm high’; GS11225, V20/f8002, Darkys Spur, Nukumaruan: H 18.2, D 15.3 mm; H 16.8, D 13.7 mm.
Remarks
This small species (commonly up to 12 mm high, reaching 20 mm) easily is recognized by the combination of the flat, tangential suture, in most small specimens having a rather low spire with straight outlines and a relatively squat shape, and the very prominent, clearly demarcated funicle, which is separated from the last whorl by a deep, narrow groove. Some larger specimens from Hawke's Bay Nukumaruan sandstone units (presumably living in shallower water than most; particularly from Darkys Spur Formation, Darkys Spur, off Kaiwaka Road, 5 km WNW of Devils Elbow, N of Napier, GS11225, V20/f8002, three specimens, B,F) develop a much more excert spire with more sinuous outlines than the common low-spired form. The suture of such specimens gradually descends below the periphery of the previous whorl as the shell grows. These specimens resemble Tectonatica species more closely than the usual short-spired form does. Beu & Maxwell (Citation1990:321) suggested that the Waipipian Taniella haweraensis is part of the variation of T. planisuturalis, and the wide range of variation of Hawke's Bay Nukumaruan specimens makes this synonymy certain. T. planisuturalis is widespread in New Zealand in Pliocene and early Pleistocene rocks, of Opoitian to early-mid Castlecliffian age. Their inclusion in the one synonymy list and usage of the name T. planisuturalis for the species by Beu & Maxwell (Citation1990:321) constitute the necessary first reviser's action to make T. planisuturalis the valid name for the species named both N. planisuturalis and N. haweraensis by Marwick (Citation1924c). Specimens definitely in situ are not known in Castlecliffian rocks in Wanganui Basin, although it is possible that some early Castlecliffian records are based on specimens in situ, whereas they occur in early Castlecliffian rocks (OIS 31?–29) at Cape Kidnappers, and in rocks deposited during about OIS 15 at Whakatane. Obviously, this species must be used as a Nukumaruan index fossil with caution, although it seems likely to be limited to Nukumaruan and older rocks in Wanganui Basin and further south.
Genus Eunaticina Fischer, 1885
Naticina Gray Citation1840:147 (nomen nudum); Gray 1847:150. Type species (by original designation): ‘Nat. papilla’ (= Natica papilla Gmelin, 1791), Recent, Indo-West Pacific; Pleistocene, New Zealand (junior homonym of Naticina Guilding, 1834).
Eunaticina Fischer 1885 (in 1880–1887), p. 768; replacement name for Naticina Gray, Citation1847.
Eunaticina papilla (Gmelin, 1791) (A)
Eunaticina papilla. Beu Citation2004:206, D–F,H (with synonymy).
Fig. 22 Argobuccinum pustulosum (Lightfoot). (A) GS4175, Q21/f6499, Denby Shellbed (Haweran, OIS 7), Waihi Beach, Hawera, earliest New Zealand record; height 57.7 mm. (B) BMNH 1973113, Dunker's illustrated syntype of Bursa tumida Dunker, 1862, “New Zealand”, height 92.6 mm. (C) holotype of Ranella polyzonalis Lamarck, 1816, MHNG 1098/79, neotype of Buccinum pustulosum Lightfoot, 1786, of Murex argus Gmelin, 1791, and of Tritonium argobuccinum Röding, 1798; type locality Jeffreys Bay, South Africa; height 71 mm. (D) TM3992, GS5674, J41/f8710, Hillgrove Formation (Last Interglacial, OIS 5e), Old Rifle Butts, S of Cape Wanbrow, S Oamaru; height 96.2 mm. (E) BMNH 1950.11.28.17, largest syntype of Ranella vexillum G. B. Sowerby I, 1835, “Chiloé, Concepción”, Chile; height 94.9 mm.

Eunaticina linneana. Maxwell Citation2009:242.
Type material
Since I (Beu Citation2004:206) discussed the type material and variation of this species and its synonyms, I enquired at CMC about the possible continued existence of the holotype of Sigaretus (Naticina) cincta Hutton (1885:318). Norton Hiller (CMC pers. comm. 11 December 2007) reported that Hutton's holotype is still present in the Museum (CMC M3014, A), and was simply missed out of the catalogue by Bradshaw et al. (Citation1992). As the holotype still exists, the neotype proposed by Beu (Citation2004:209) (AIM AK102482) was unnecessary, and has no status (ICZN Art. 75.8). It should be noted that the holotypes of Mitrellatoma angustata (Hutton, 1886) (see Part 4) and Cominella incisa (Hutton, 1885) have the same history; no type material was listed by Bradshaw et al. (Citation1992), but both holotypes are still in CMC. Obviously the catalogue by Bradshaw et al. (Citation1992) must be used with caution.
Dimensions
Holotype of Sigaretus cinctus: H 14.8, D 12.8 mm; neotype proposed by Beu (Citation2004:209): H 11.3, D 10.2 mm.
Remarks
The synonymy and the time range in New Zealand of Eunaticina papilla were discussed in Part 1 of this series (Beu Citation2004:206). During the search of Henry Suter's notes and photographs in GNS (for the illustration of a syntype of Venericardia zelandica Potiez & Michaux, 1844,=Austrovenus stutchburyi (Wood, Citation1828), sent to Suter by the curator of the Muséum de Douai, France; Beu Citation2006:280), a set of photographic prints was found. These are evidently duplicate prints of the illustrations in several of Suter's New Zealand Geological Survey Paleontological Bulletins, particularly Suter (Citation1915). One of them shows the holotype of Sigaretus (Naticina) cinctus Hutton. This print is considerably larger and clearer than Suter's (1915), pl. 4, ) published figure and demonstrates that the holotype recognized by Suter (Citation1915) definitely is the specimen of E. papilla in CMC (M3014). The dimensions of the holotype were stated by Suter (Citation1915:9) as H 16, D 12.5 mm, substantially agreeing with those of M3014 (H 14.8, D 12.6 mm), indicating that the illustration by Marwick (Citation1924c, pl. 60, , 16) is enlarged ×2, not ×1 as stated in the caption; Marwick's illustration is about 31 mm high. The holotype is a white, slightly chalky shell with a pale yellow stain, closely resembling many other shells from Landguard Sand, and Marwick (1924c:573) was incorrect to doubt its provenance. The holotype of S. cinctus is actually a rather small, inflated specimen of E. papilla, considerably smaller than the several specimens known from Te Piki, near East Cape, although it is larger and a little taller and narrower than the redundant neotype I proposed (Beu Citation2004:209).
Family Cassidae
Genus Semicassis Mörch, Citation1853
Semicassis Mörch 1853:112. Type species (by subsequent designation, Harris, 1897:198): Cassis japonica Reeve, 1848 (= Cassis bisulcataSchubert & Wagner, 1829), Miocene to Recent, Indo-West Pacific.
Faurotis Jousseaume Citation1888:188, Type species (by original designation): Faurotis faurotis Jousseaume, 1888, Recent, western Indian Ocean & Red Sea.
Xenophalium Iredale Citation1927:333. Type species (by original designation): Xenophalium hedleyi Iredale, Citation1927 (= Cassidea royana Iredale, 1914), Recent, northern New Zealand, Kermadec Islands and southeastern Australia.
Xenogalea Iredale Citation1927:339. Type species (by original designation): Cassis pyrum Lamarck, 1822, Pleistocene and Recent, New Zealand and southeastern Australia.
Tylocassis Woodring Citation1928:306. Type species (by original designation): Buccinum inflatum Shaw, 1811 (= Buccinum granulatum Born, 1778), Pleistocene and Recent, western Atlantic.
Remarks
The synonymy above is taken from Beu (Citation2005:50), and was based on research on tropical American Neogene Tonnoidea (Beu Citation2010). The minor characters of nodulation and ridges of the inner lip callus separating supposed genus-group taxa such as Semicassis, Tylocassis, Xenophalium and Xenogalea are in my opinion only of specific rank, and no characters of the radula, protoconch or internal anatomy separate these species into distinct genera.
Semicassis labiata (Perry, Citation1811) (C–F,H)
Cassidea labiata Perry 1811, pl. 34, .
Cassis achatina Lamarck 1816, pl. 407, a, b, ‘Liste des objets’ p. 3; Lamarck 1822b:226; Kiener Citation1835:37, pl. 13, Fig. 24; Deshayes 1843:33; Reeve 1848, Cassis, pl. 10, Fig. 28a; Krauss Citation1848:115; Küster 1857:34, pl. 50, , 4; Angas Citation1867:196; Hutton 1873a:20; Hutton 1880:66; GB Sowerby III Citation1892:23; Pritchard & Gatliff Citation1900:189.
Cassis zeylanica Lamarck 1822b:226; Kiener Citation1835:38, pl. 13, Fig. 26; Deshayes 1843:33.
Buccinum achatinum. Wood 1828:105, pl. 22, .
Cassidea achatina. Swainson 1840:299.
Cassis pyrum var. Küster 1857:29, pl. 49, , 4 (in part; references to C. zeylanica Lamarck only).
Cassis (Casmaria) achatina. Tryon 1885:278, pl. 8, fig. 95.
Cassis ceylonica (sic). Tate & May Citation1901:444.
Cassidea labiata. Hedley Citation1902a:27.
Cassis pirum var. intercedens Martens 1904:54, 56, note 12.
Semicassis labiata. Suter in Hutton Citation1904:75; Moss Citation1908:23, pl. 5, ; Beu Citation1976:421; Wilson Citation1993:233, pl. 37, a,b; Richardson Citation1997:9; Spencer at al. 2009:204.
Phalium labiatum. Suter Citation1913:312, pl. 40, ; Hedley Citation1918:M67; Bucknill Citation1924:55, pl. 4, ; Coleman Citation1975:187, fig. 528.
Phalium n. sp. (labiatum auct.) Finlay Citation1926b:400, pl. 20, fig. 62, 63.
Xenogalea labiata. Iredale Citation1927:347:31, ; Cotton Citation1945:251; Macpherson & Gabriel Citation1962:149, fig. 178; Iredale & McMichael Citation1962:56; Anderson Citation1966:243, pl. 10 (egg mass).
Xenogalea insperata Iredale Citation1927:349, pl. 31, ; Iredale & McMichael Citation1962:56.
Xenogalea collactea Finlay Citation1928:246.
Xenophalium labiatum. Powell Citation1928:640, –17; Powell Citation1937:74, pl. 12, ; Penniket & Moon Citation1970:32, pl. 13, .
Xenophalium insperatum. Powell Citation1928:640, .
Cassis achatina craticulata Turton Citation1932:112 (not Cassis craticulata Euthyme, 1884).
Cassis zealanica (sic). Turton Citation1932:112.
Phalium (Xenogalea) labiatum. var. iredalei Bayer Citation1935:109 (new synonym).
Phalium pyrum var. zeylanica Bayer Citation1935:110.
Xenophalium collactea. Powell Citation1937:74, pl. 12, Fig. 23.
Xenogalea inseparata (sic) Cotton Citation1945:251.
Xenophalium (Xenogalea) labiatum Powell Citation1946a:77, pl. 12, ; Powell Citation1958a:101, pl. 12, ; Powell Citation1962:93, pl. 12, .
Xenophalium (Xenogalea) collactea Powell Citation1946a:77, pl. 12, Fig. 23; Powell Citation1958a:101, pl. 12, Fig. 23; Powell Citation1962:93, pl. 12, Fig. 23.
Phalium iheringi Carcelles Citation1953:5, –10 (new synonym).
Phalium (Xenophalium) labiatum. Abbott Citation1968:183, pl. 13, –5, 17–21; pl. 173; pl. 175.
Xenophalium labiatum labiatum. Powell Citation1976b:96, pl. 19, ; Powell Citation1979:159, pl. 34, , 8.
Xenophalium labiatum insperatum. Powell Citation1976b:93, pl. 19, Fig. 23; Powell Citation1979:159, pl. 34, , 10.
Phalium (Casmaria) decipiens. Kilburn Citation1980:195, , 8 (new synonym).
Casmaria decipiens. Kreipl Citation1997:43, pl. 15, fig. 44.
Semicassis (Semicassis) labiata labiata. Kreipl Citation1997:57, pl. 20, fig. 66.
Semicassis (Semicassis) labiata forma collactea. Kreipl Citation1997:57, pl. 20, fig. 66a.
Semicassis (Semicassis) labiata iheringi. Kreipl Citation1997:57, pl. 20, fig. 67, 67a.
Semicassis (Semicassis) labiata zeylanica. Kreipl Citation1997:58, pl. 20, fig. 68, 68a.
Semicassis (Semicassis) labiata forma iredalei. Kreipl Citation1997:58, pl. 20, fig. 68b.
Semicassis (Semicassis) labiata. Maxwell Citation2009:243.
Type material
No type material is known for the taxa proposed by Perry (1811). The Lamarck collection in Geneva contains two specimens (MHNG 1100/99–100; G–I) identified as Cassis achatina, and the annotations by Rosalie de Lamarck on Lamarck's copy of Animaux sans vertèbres indicate that Lamarck owned two specimens (Y Finet MHNG pers. comm. June 2004). In the opinion of Y Finet, it is difficult to be certain that these two specimens are the original Lamarck syntypes, but as there are no other possible syntypes in the Delessert collection that might have belonged to Lamarck, these specimens are accepted as syntypes. The difficulty is the size—Lamarck stated the height as ‘2 pouces 2 lignes’, which is about 58.5 mm, whereas the two specimens in MHNG are 54.5 and 53.0 mm high. Not enough detailed characters are visible to be able to match either of the specimens with the illustration by Lamarck (1816, pl. 407, a, b). However, there is no doubt that specimen MHNG 1100/99 is that illustrated by Kiener (1835, pl. 13, Fig. 24). Abbott (Citation1968:186) designated this specimen as the lectotype of Cassis achatina Lamarck, 1816. Both Lamarck and Kiener indicated the type locality as ‘Nouvelle-Hollande’, and the type locality is here designated as Sydney Harbour, New South Wales. In order to attach the name Cassidea labiata Perry, 1811 unambiguously to this species, the same specimen, MHNG 1100/99 (C,D), is here designated the neotype of Cassidea labiata Perry, 1811. The type material of Cassis zeylanica Lamarck was also said by Abbott (Citation1968:187) to be in MHNG. The Lamarck collection in MHNG contains two specimens identified as Cassis zeylanica Lamarck (MHNG 1101/3–4; D,E) and again, Rosalie de Lamarck's annotations on Lamarck's copy of Animaux sans vertèbres call for two specimens in Lamarck's collection (Y Finet MHNG pers. comm. June 2004). However, again there is little proof that the two segregated specimens are Lamarck's original syntypes, as this species was not illustrated by Lamarck (1816), but both are from the Delessert collection, and one of them appears to be that illustrated by Kiener (1835, pl. 13, Fig. 26). Although they are labelled ‘Ceylan’, as stated by Lamarck (1822b) (that is, Sri Lanka), they are not accompanied by an original Lamarck label. Nevertheless, in the absence of any other possible Lamarck syntypes, these specimens are accepted as the syntypes of Cassis zeylanica Lamarck. Lamarck (1822b:226) stated the dimensions as ‘2 pouces 10 lignes et demie’, or about 77.5 mm; MHNG 1101/3 is 76 mm high, and MHNG 1101/4 is 81 mm high. Xenogalea insperata, two syntypes AMS C53270, from Sydney Harbour, New South Wales (illustrated by Abbott Citation1968, pl. 173). Xenogalea collactea, holotype AIM AK70851, from Opotiki, Bay of Plenty, with one paratype, AIM AK72554 (not seen). Phalium labiatum var. iredalei, holotype RMNH.MOLL.110052, from ‘South Africa’; three paratypes RMNH.MOLL.110053, from Port Elizabeth, South Africa; two paratypes RMNH.MOLL.110054, from Port Elizabeth (photographs sent by J Goud RMNH 5 November 2007). Phalium iheringi, holotype in Museu de Historia Natural de Montevideo, Uruguay, no. 13805, with two paratypes, nos. 16764, 17797; holotype and paratype no. 17797 from beach, Mar del Plata, Argentina; paratype no. 16764 from 37°38′S, 56°20′W, 84 m, ‘Undine’, October 1926 [off Macedo, northern Argentina N of Mar del Plata] (Carcelles Citation1953; not seen). Phalium (Casmaria) decipiens Kilburn, holotype Natal Museum, Pietermaritzburg, NM5422, T2292, from Melville, Natal south coast, South Africa; 4 paratypes examined in Natal Museum, listed by Kilburn (Citation1980:196–197).
Other material examined
Haweran: the earliest New Zealand occurrence of Semicassis labiata I am aware of is in the Te Piki fauna, road cutting 6 km E of Whangaparaoa, near East Cape, OIS 7 (GS5833, Y14/f7505A, H, one immature specimen, the basis of the record by Richardson Citation1997:9). I am not aware of any other fossil record.
Distribution
OIS 7–Recent, largely limited to the NE North Island warm province, although rarely extending as far south as the Kapiti Coast, W Wellington (Bracegirdle Citation2007); Recent in Australia, from southern Queensland (as far north as Moreton Bay; database maps by I Loch, AMS pers. comm.) to NE Victoria and, uncommonly, across the southern coast of Australia to southern Western Australia (Wilson Citation1993:234); South Africa and southern South America, that is, circum-Southern Ocean.
Dimensions
GS5833, Te Piki (immature): H 31.3, D 20.5 mm; adult Recent specimens: RM370, Mangapai, Northland: H 63.1, D 39.8 mm; RM4971, Waihau Bay, E Bay of Plenty: H 74.8, D 47.7 mm.
Remarks
As with several other circum-Southern Ocean tonnoideans, it seems likely that Semicassis labiata occupies the entire Southern Ocean, and the other named, supposedly geographically restricted forms are part of the variation of one species (S. labiata iheringi Carcelles, Citation1953, southern South America; Rios 1994:86, pl. 28, fig. 334; Kreipl Citation1997:57, pl. 20, fig. 67, 67a; S. labiata zeylanica Lamarck, 1822, South Africa; Kilburn & Rippey Citation1982:72,212, pl. 16, a,b; Kreipl Citation1997:58, pl. 20, fig. 68–68b). However, this needs further research, including a molecular study, before it is accepted, particularly as some Southern Ocean tonnoideans are now known to comprise several distinct species—notably in Fusitriton. Specimens of the zeylanica form also have a wider groove between the siphonal fasciole and the previous whorl than in Australian–New Zealand specimens of S. labiata. The South African form named Phalium (Casmaria) decipiens by Kilburn (Citation1980:195, , 8; Kreipl Citation1997, pl. 15, fig. 44) is coloured and sculptured identically to the usual southern Australian and northern New Zealand smooth form of S. labiata, and has a very narrow groove between the siphonal fasciole and the previous whorl. It is possibly a synonym of S. labiata, based on the form that is common in Australia and New Zealand, but evidently is uncommon in South Africa, although the specimen illustrated by Kreipl (Citation1997, pl. 15, fig. 44) has a larger protoconch than Australasian specimens. I (Beu Citation1976) pointed out that S. labiata is virtually indistinguishable from European fossil specimens usually identified as Semicassis laevigata (Defrance, 1817). Landau et al. (Citation2004:48) showed that S. laevigata and S. saburon (Bruguière, 1792) are distinct species, the Miocene–Pliocene species lacking the obvious spiral grooves of the Pleistocene and Recent species S. saburon. In the Australian and New Zealand fauna, the uncommon species S. labiata easily is distinguished from the much more abundant S. pyrum (Lamarck, 1822) by its smaller size, its narrower, more ovoid shape with a more convex spire outline, its smoother exterior, its more consistently nodulose interior of the outer lip and, in particular, by the much narrower groove separating the siphonal fasciole from the previous whorl. The variation is quite marked in all populations of S. labiata, and includes both smooth (‘typical S. labiata’) and nodulose specimens (‘insperata’, ‘collactea’), and specimens with and without short transverse ridges inside the outer lip. The paralectotype of Cassis achatina, MHNG 1100/100 (I), is an unusual specimen combining the characters of the typical labiata and ‘collactea’ forms, with nodules on the periphery of the pale penultimate whorl up until the aperture, then changing on the last whorl to a smooth, brightly coloured surface. Specimens with obvious nodules around the periphery seem mainly to have been collected on soft substrates near rocks, or to wash ashore on sandy beaches, whereas the more common smooth form lives on rocky shores and subtidal rocks. Communal egg-laying by at least seven smooth specimens on subtidal rocks off Sydney (N Coleman pers. comm.), New South Wales, was illustrated by Coleman (Citation1975:187, fig. 528), and the animal illustrated creeping along with foot, siphon and cephalic tentacles expanded on a sand substrate at Jervis Bay, southern New South Wales, in the photograph above (Coleman Citation1975:187, fig. 527) is the ‘insperata’ form of S. labiata, rather than S. pyrum as identified in the caption. Anderson (Citation1966:243, pl. 10) also described communal egg-laying in this species and illustrated a large communal egg mass.
Semicassis labiata is recorded earliest in New Zealand in the Te Piki fauna (OIS 7), although a small, nodulose form of S. pyrum is much more common at Te Piki than S. labiata. The phylogenetic relationship between S. labiata and the Atlantic and Mediterranean Miocene–Pliocene species S. laevigata is very close, but they appear to be distinct species, as S. labiata is narrower and still more smoothly oval than S. laevigata. The first appearance of S. labiata in New Zealand probably is biostratigraphically meaningful, as it was dispersed around the Southern Ocean in the Antarctic Circumpolar Current as planktotrophic larvae from South Africa. However, in New Zealand this species is uncommon everywhere, and it occurs only rarely outside the warm-water NE North Island province, and so will be of biostratigraphical utility only if it should be found in terrace cover-bed faunas in N New Zealand.
An important implication for dispersal times and routes is provided by Semicassis labiata appearing in New Zealand only during OIS 7. This is at least 1 m yr after the more strongly sculptured species S. saburon had evolved in the North Atlantic–Mediterranean area, during Pleistocene time, whereas the smooth Southern Hemisphere specimens are scarcely distinguishable from the extinct European species S. laevigata, last known in early Pleistocene rocks. This suggests that a complex history of stepping-stone dispersal allowed S. labiata to evolve during its migration to the Southern Hemisphere; migration evidently did not take place in a single rapid event during one glacial or interglacial period. Migrations of other tonnoideans to and around the Southern Hemisphere possibly occurred in a similar way. S. laevigata probably reached the warm southeastern Atlantic (and probably also South Africa) near the end of Pliocene time, along with other Mediterranean–West African tonnoideans known to occur as far south as Angola and Namibia, such as Charonia lampas and Aspa marginata (Gmelin, 1791) (Beu Citation2010), before S. saburon had evolved in the Mediterranean. In southern Africa and the South Atlantic, it evolved into the sister species S. labiata. At some later time, S. labiata was dispersed passively as planktotrophic larvae in the Antarctic Circumpolar Current to Western Australia, as Wells & Kilburn (1986) demonstrated for three other restricted South African molluscs, including the tonnoidean Cabestana africana (A. Adams, 1855). It might then have taken a significant further period of time for S. labiata to disperse to eastern Australia and New Zealand. Other, smaller islands along the way, such as St Paul and Amsterdam Islands in the southern Indian Ocean, have certainly provided stepping stones where larvae have been able to survive and breed, for some tonnoideans such as Argobuccinum, Charonia lampas and both Recent Ranella species (Arnaud & Beurois Citation1972). The enormously long planktotrophic larval lives that have been evolved by tonnoideans (as well as Architectonicidae) to allow larval dispersal in ocean currents over wide distances are exemplified by Fustriton oregonensis (Redfield, 1846), which Strathman & Strathman (Citation2007) have shown can survive as veligers for at least 4.5 yr. This provides plenty of scope for Semicassis labiata and Argobuccinum pustulosum (Lightfoot, Citation1786) to be dispersed around the Southern Ocean in the Antarctic Circumpolar Current.
Family Ranellidae
Genus Argobuccinum Herrmannsen, Citation1846
Argobuccinum Herrmannsen 1846:77. Type species (by original designation): Murex argus Gmelin, 1791 (= Buccinum pustulosum Lightfoot, 1786), middle Miocene to Recent, circum-Southern Ocean.
Gondwanula Finlay Citation1926b:399. Type species (by original designation): Bursa tumida Dunker, 1862 (= Buccinum pustulosum Lightfoot, 1786).
Mediargo Terry Citation1968:42. Type species (by original designation): Gyrineum mediocre Dall, 1909, middle Miocene–Pliocene, western United Sates of America.
Remarks.
Mediargo was proposed by Terry (1968:42) for NW American fossil species that are slightly smaller and more weakly sculptured than species previously referred to Argobuccinum. However, as species such as ‘Mediargo’ mediocris (Dall, 1909) are the only known possible ancestors of the Pliocene–Recent species of Argobuccinum, this seems to be a single clade, and in my opinion Mediargo is a synonym of Argobuccinum.
Argobuccinum pustulosum (Lightfoot, 1786) (A–E)
Buccinum pustulosum Lightfoot 1786:88; Dall Citation1921:125.
Murex argus Gmelin 1791:3547; Dillwyn 1817:694.
Tritonium argo-buccinum Röding Citation1798:127.
Cassidea tuberculata Fischer von Waldheim, Citation1807:185; Ivanov & Kantor, Citation1991:82; Ivanov et al. Citation1993:77, pl. 7, .
Ranella polyzonalis Lamarck 1816, pl. 414, a,b; ‘Liste des objets’:4.
Ranella argus. Lamarck 1822b:151; Kiener Citation1841:31, pl. 8, ; Reeve Citation1844, pl. 3, ; Küster & Kobelt 1870:125; 1878:329, pl. 37, , 6.
Triton ranelliformis King Citation1832:347.
Ranella vexillum GB Sowerby II 1835, pl. 84, ; Menke 1843:24; Reeve 1844, pl. 3, .
Ranella kingii Orbigny Citation1841a:451 (unnecessary new name for Triton ranelliformis King, 1832, not a homonym of Triton ranelloides Reeve, 1844).
Argobuccinum argus. Herrmannsen 1846:77; Suter Citation1913:309, pl. 43, ; Dell Citation1963b:218, pl. 2, , 5; JT Smith 1970:458, pl. 39, , 10–12; Arnaud & Beurois Citation1972:869–870.
Ranella ampullacea Valenciennes Citation1858:759,762 (synonym according to Kantor & Sysoev Citation2002:121, A,B).
Bursa tumida Dunker 1862:239, Dunker 1863:56, pl. 18, , 9.
Bursa zelebori Dunker Citation1863:57 (nomen nudum).
Bursa (Apollon) proditor Frauenfeld Citation1865:894.
Bursa (Apollon) prodita. Frauenfeld Citation1867:4, pl. 1, a, 1b.
Ranella (Argobuccinum) argus. Tryon Citation1880b:44, pl. 24, fig. 61; Watson Citation1886:400.
Ranella tristanensis ‘Gray’ Paetel Citation1888:106 (nomen nudum).
Argobuccinum tumidum. Hedley Citation1913:297; Fleming Citation1952:79,84; JT Smith 1970:469, pl. 39, , 3; pl. 41, –4, 8, 9; Powell Citation1979:166, pl. 12, ; Maxwell Citation2009:243.
Gondwanula tumida. Finlay Citation1926b:399.
Argobuccinum (Argobuccinum) ranelliformis ranelliformis (sic). Dell Citation1963b:219, pl. 2, .
Argobuccinum (Argobuccinum) ranelliformis (sic) tumidum. Dell Citation1963b:220, text-.2, pl. 1, –4.
Argobuccinum (Argobuccinum) proditor proditor. Dell Citation1963b:221, pl. 2, ; Beu 1985:56.
Argobuccinum (Argobuccinum) proditor tristanensis (sic). Dell Citation1963b:222, pl. 2, , 6.
Argobuccinum pustulosum. Rehder 1967:16; Kilburn & Rippey 1982:75, pl. 17, a,b; Kilburn Citation1984:8, pl. Af, h, g.; Wilson 1993:241, pl. 41, .
Argobuccinum ranelliforme. JT Smith 1970:462, pl. 39, , 4, 6, 9; pl. 40, , 6, 7, 10–12.
Argobuccinum tristanense. JT Smith 1970:466, pl. 39, , 8: pl. 40, .
Argobuccinum proditor. JT Smith 1970:468, pl. 41, –7, 10.
Argobuccinum ranelliforme ranelliforme. Arnaud & Beurois 1972:869–870.
Argobuccinum ranelliforme tumidum. Arnaud & Beurois 1972:869–870.
Argobuccinum ranelliforme proditor. Arnaud & Beurois 1972:866, , 4; pl. 1, , 4 (= tristanense Dell, 1963; not pl. 1, a,b,=Ranella gemmifera Euthyme, 1886).
Argobuccinum (Argobuccinum) proditor tristanense. Beu Citation1985a:56.
Argobuccinum (Argobuccinum) pustulosum pustulosum. Beu 1985:56, .
Argobuccinum (Argobuccinum) pustulosum ranelliforme. Beu 1985:56.
Agrobuccinum (Argobuccinum) pustulosum tumidum. Beu 1985:56.
Argobuccinum (Argobuccinum) pustulosum. Henning & Hemmen 1993:13, pl. 1, .
Argobuccinum (Argobuccinum) ranelliforme. Henning & Hemmen 1993:14, pl. 1, .
Argobuccinum (Argobuccinum) tumidum. Henning & Hemmen Citation1993:15, pl. 1, .
Argobuccinum (Argobuccinum) proditor. Henning & Hemmen 1993:15, pl. 1, .
Argobuccinum (Argobuccinum) tristanense. Henning & Hemmen 1993:16, pl. 1, .
Argobuccinum pustulosum tumidum. Spencer et al. 2009:206.
Type material
No type material is known for taxa proposed by Lightfoot (1786), Gmelin (1791) or Röding (1798). Cassidea tuberculata, lectotype designated by Ivanov & Kantor (1991:82–83), Zoological Museum, Moscow University no. ZMUM N L-518, without locality; illustrated by Ivanov et al. (Citation1993, pl. 7, ), a typical if rather tall and narrow specimen of Argobuccinum pustulosum (not seen). Ranella polyzonalis Lamarck, holotype MHNG 1098/79 (C), now labelled ‘Ranella argus Lamarck’, without locality. The specimen closely matches the illustration by Lamarck (1816, pl. 414, a,b). The type locality was designated by me (Beu Citation2010) as Jeffreys Bay, eastern Cape Province, South Africa. To attach all earlier names unambiguously to the present species, I (Beu Citation2010) designated the holotype of Ranella polyzonalis Lamarck as the neotype of Buccinum pustulosum Lightfoot, 1786, the neotype of Murex argus Gmelin, 1791, and the neotype of Tritonium argobuccinum Röding, 1798. Triton ranelliforme King, five unregistered presumed syntypes in BMNH, two probable syntypes presented by Captain PP King and three ‘possible’ syntypes from the Broderip collection (labelled B2115 or B2252 inside the aperture), all unlocalized, stated by King (1832:347) to be from ‘Sinum Peñas et oram occidentalum America meridionalis’. Ranella ampullacea Valenciennes, one syntype in MNHN (Kantor & Sysoev 2002, A,B), supposedly from the ‘Straits of Tartary’, between Sakhalin and eastern Russia; a wrongly localized specimen of the South American vexillum form of A. pustulosum, type locality designated by me (Beu Citation2010) as Chiloé Island, Chile. Ranella vexillum G. B. Sowerby II, three syntypes BMNH 1950.11.28.17-19, from ‘Chilöe Id., Concepción, rocky places, 3–5 fathoms’ [6–10 m]; the medium-sized syntype (E; H 85.0, D 50.0 mm) is the specimen illustrated by both GB Sowerby II (1835, pl. 84, ) and Reeve (1844, pl. 3, ). Ranella kingii d'Orbigny, three syntypes BMNH 1854.12.4.533, from Isla Mocha, Chile. Bursa tumida Dunker, Citation1862, three syntypes BMNH 1973113, from ‘New Zealand’; the largest specimen (B) is the one illustrated by Dunker (1863, in 1863–1864, pl. 18, , 9). Ranella (Apollon) proditor Frauenfeld, five syntypes NHMW 689–693, from St Paul Island; the largest syntype (NHMW 689, H 100.9, D 56.6 mm) is the specimen illustrated beautifully by Frauenfeld (1867, pl. 1, a,b). Argobuccinum proditor tristanense Dell, holotype BMNH 1887.2.9.1221, collected on the shore at Tristan de Cunha Island by the ‘Challenger’ Expedition, with four lots of paratypes.
Other material examined
Haweran: the oldest New Zealand fossil record is from Denby Shellbed (OIS 7), Waihi Beach, Hawera (GS4175 re-collection, Q21/f6499, two specimens from different collectors; A); thereafter common in OIS 5e shellbeds, particularly in southern New Zealand (specimens in GNS reference collection: GS9727, N33/f7580, terrace above Cranky Toms Creek, Motunau, N Canterbury; GS14842, N34/f094, Motunau terrace, N Canterbury, OIS 5e; common in GS5674, J41/f8710, Hillgrove Formation, Old Rifle Butts, S of Cape Wanbrow, Oamaru, OIS 5e (D; see Worthy & Grant-Mackie Citation2003); GS158, O32/f9038; GS167, J39/f7477; & GS14697, O32/f0108, Haumuri Bluff high terrace, Marlborough, OIS 5c?) and in many younger deposits (e.g. GS13796, W21/f057; GS13937, W21/f058, terrace at shelter hut, Cape Kidnappers, Hawke's Bay, 2500 yr old; GS12254, CH/f0532, Owenga ‘shell bank’, Chatham Island, Holocene; and several other Holocene localities). The young radiocarbon ages (22,476±352 – 33,711±1215 14C yr) for the avifauna of Hillgrove Formation, Oamaru, listed by Worthy & Grant-Mackie (Citation2003) are either minimum ages, or imply that the non-marine, vertebrate-rich upper part of this formation is much younger than the basal part, with marine fossils, as this youngest marine terrace along the Otago coast is likely to have been cut during OIS 5e (ca 125,000–130,000 yr), like all the main uplifted terraces around New Zealand and much of the rest of the world.
Distribution
OIS 7–Recent, in New Zealand. Recent specimens occur around the entire Southern Ocean, in South Africa, including up the west coast north of Capetown, at St Paul and Amsterdam Islands, at Kerguelen, around southern South America from central Chile (Talcahuano, 36°40′S; Dell Citation1963b:220; but Aldea & Valdovinos [Citation2005] recorded it only as far north as Isla Santa María, 37°00′12″S) south to Cape Horn and north to central Argentina, along the southern coasts of Australia in South Australia, Victoria and Tasmania, and throughout New Zealand (where, however, it is more common in the south) and the Chatham Islands. JT Smith (1970:465) recorded a single fossil specimen in Chile from Navidad Formation (traditionally dated as middle or late Miocene, but concluded by Finger et al. [Citation2007] to be reworked Oligocene to early Miocene fossils in rocks of late Miocene–early Pliocene age). However, as there are no other records from Navidad Formation, particularly in the large collections in the Museo Nacional de Historia Natural, Santiago, the specimen Smith cited possibly bears an incorrect locality. Fossils are common in later Pliocene rocks (at La Cueva and Coquimbo) and are common in Pleistocene terrace cover beds in Chile (Herm Citation1969:71; JT Smith 1970).
Dimensions
Largest New Zealand Recent specimens, RM5691, 120 m, off Taiaroa Head, E Otago: H 135.1, D 81.2 mm; RM4265, beach, Cape Campbell, Marlborough: H 128.1, D 74.1; NMNZ M.19477, Foveaux Strait: H 136.4 mm; large fossil, GS5674, TM3992, Hillgrove Formation, Old Rifle Butts, South Oamaru: H 96.2, D 62.7 mm; GS4175 re-collection, Q21/f6499, Denby Shellbed, Hawera, OIS 7: H 57.7, D 37.7 mm; Bursa tumida Dunker, syntypes: H 92.6, D 56.5 mm; H 91.2, D 55.6 mm; H 81.2, D 53.2 mm; Ranella vexillum Sowerby, syntypes: H 94.9, D 58.3 mm; H 85.0, D 50.0 mm; H 67.3, D 38.4 mm; Argobucinum proditor tristanensis Dell, holotype: H 53.4, D 29.8 mm; largest paratype: H 74.9, D 36.2 mm.
Remarks
The taxonomy of the Recent Southern Ocean Argobuccinum species has long been debated, because of the great similarity of the several named forms around the Southern Ocean and their intergradation in most shell characters. Opinions have ranged from a single variable species (Suter Citation1913, and several earlier authors; Beu Citation2010) to various combinations of species and geographical subspecies at each of the separate landmasses and islands (Dell Citation1963b; JT Smith 1970; Arnaud & Beurois Citation1972). The overlap of characters seen when a large collection is assembled seems to encompass all named forms, and I tentatively consider them all to be conspecific with Argobuccinum pustulosum, awaiting a study of DNA sequences. The only other realistic possibility is that A. pustulosum (South Africa), A. ranelliforme (King, 1832) (South America), A. tumidum (Dunker, 1862) (New Zealand and S Australia), A. proditor (Frauenfeld, 1865) (St Paul & Amsterdam Islands) and A. tristanense Dell, 1963 (Tristan da Cuñha & Nightingale I.) are distinct species.
Argobuccinum pustulosum is a large, robust shell, which preserves well in terrace cover beds (and is common in places, notably along the SE South Island) even when their fauna consists of few, abraded shells. Among the New Zealand late Neogene and Recent Ranellidae, it is easily recognized by its low spire and short, wide shape, its evenly inflated last whorl, its low varices (wider than they are high) at about each 180–190° around the shell, that is, almost aligned up the opposing sides of the spire, and its large, subcircular, weakly armed aperture. The exterior sculpture consists mainly of many low, narrow, finely nodulose spiral cords, although some specimens have several to many small nodules on one or two cords around the periphery, and in a few specimens quite coarse nodules are present over most of the exterior, so that these closely resemble South African specimens of the typical form of A. pustulosum. Fresh shells, including some late Haweran fossils, have a marbled red-brown and blackish colour pattern that becomes speckled with white when the nodules are abraded. Dell (Citation1963b) recorded fossil specimens from a few localities in New Zealand. Subsequent improved dating of these faunas has shown that the sole record older than OIS 5e is that from Denby Shellbed, Hawera (OIS 7). Argobuccinum pustulosum is probably useful for segregating terrace faunas older than OIS 5e (lacking Argobuccinum) and younger than OIS 7 (with Argobuccinum). As with Semicassis labiata, this is probably a biostratigraphically useful first appearance, as Argobuccinum pustulosum was dispersed around the Southern Ocean (originally from South America; the genus evolved in the North Pacific) as planktotrophic larvae in the Antarctic Circumpolar Current, apparently during glacial OIS 8, but evidently was rare in New Zealand (or occurred mainly in the subantarctic zone to the south?) before OIS 5e.
Acknowledgements
Funded by the New Zealand Foundation for Research, Science and Technology through Backbone Fund Contract CO5XO412 (National Paleontology Collection). Dedicated to the late Dick Dell and Charles Fleming, who inspired my life-long interest in malacology and paleontology, and to the late Phil Maxwell, my friend and colleague for 40 years. Many of the new records from Wanganui Basin were collected and recognised by Charles Fleming, who charged me with recording them. Bruce Marshall (NMNZ) provided extensive, critically important data through his gathering and curation of the huge NMNZ collection, as well as many suggestions on taxonomy and biogeography over many years, provided valuable comments on the manuscript, and pointed out the reference by Morley (1996). Many New Zealand and Australian “Wanganui working group” colleagues have contributed locality data and stratigraphy over the years: Bob Carter (James Cook University of North Queensland, Townsville), Tim Naish & Brent Alloway (School of Earth Sciences, Victoria University of Wellington; formerly of GNS), Brad Pillans (ANU), and the late Tony Edwards. Philippe Bouchet, Pierre Lozouet and Virginie Héros (MNHN) generously provided access to the MNHN collections, library, and type material, loaned specimens, and sent photographs of Lamarck's types, and Jacques Le Renard (MNHN) provided information on many Paris Basin fossils (see his website at http://www.somali.asso.fr/fossils/biotaxis.php for thousands of photos of Paris Basin molluscs). Norton Hiller, S. Stollmon and Kate McCaughan (CMC), Todd Landers and Owen Lee (AIM), Jeroen Goud (RMNH), Jack Grant-Mackie and Neville Hudson (AUGD), Anita Eschner (NHMW), Kathe Jensen (ZMC), and in particular, Kathie Way and Amelia McLennan (BMNH) provided many type data and loaned types, and Yves Finet (MHNG) provided information on and photos of Lamarck's types. Other comparative material was loaned or data provided by Dai Herbert (NMP) and Ian Loch, Alison Miller and Janet Waterhouse (AMS). Dennis Gordon (NIWA, Wellington) advised me on polychaete names. Louella Saul (Los Angeles County Museum of Natural History) initiated the investigation of Maoricrypta hochsetetteriana, Graeme Wilson dated it by dinoflagellates, and Roger Tremain (GNS) prepared the sample. Maureen Coomer, Pauline Muir, Denise Garbutt, and particularly Kristin Garbett (GNS Library) found numerous obscure references, Marianna Terezow (GNS) took the light photographs, Philip Carthew (GNS) carried out extensive work on the photos and arranged the plates, and Scott Morgan and James Xia (Industrial Research Ltd, Lower Hutt), Marianna Terezow, and George Scott (GNS) took the SEM micrographs.
Related Research Data
References
- Abbott , RT . 1968 . The helmet shells of the world (Cassidae). Part 1 . Indo-Pacific Mollusca , 2 ( 9 ) : 7 – 202 .
- Abbott , ST and Carter , RM . 1999 . Stratigraphy of the Castlecliffian type section: 10 mid-Pleistocene sequences from the Wanganui coast, New Zealand . New Zealand Journal of Geology and Geophysics , 42 : 91 – 111 .
- Abbott , ST , Naish , TR , Carter , RM and Pillans , BJ . 2005 . Sequence stratigraphy of the Nukumaruan stratotype (Plio-Pleistocene, c. 2.08–1.63 Ma), Wanganui Basin, New Zealand . Journal of the Royal Society of New Zealand , 35 : 123 – 150 .
- Adams A 1853 . Contributions towards a monograph of the Trochidae, a family of gasteropodous Mollusca . Proceedings of the Zoological Society of London 19 : 150 192 [‘1851’, issued Jan–June 1853; Trew (1992); reprinted in part in The Annals and Magazine of Natural History, Series 2, 12: 142–148; 199–213, Aug–Sept 1853] .
- Adams A 1855 . A monograph of Cerithidea, a genus of Mollusca, with descriptions of several new species, from the collection of Hugh Cuming, Esq.; to which are added, descriptions of two new species of Colina, and one of Donax . Proceedings of the Zoological Society of London 22 : 83 87 [‘1854’, issued April 1855] .
- Adams H , Adams A 1853–1858 . The genera of Recent Mollusca, arranged according to their organisation . London, J. van Voorst . Vol. 1, x + 484 p.; vol. 2, 661 p.; vol. 3, 138 pls. [issued in 36 parts; dates (vol. 2, p. 661): vol. 1, pp. 1–256, pls. 1–32, 1853; pp. 257–484, pls. 33–60, 1854; vol. 2: pp. 1–92, pls. 61–72, 1854; pp. 93–284, pls. 73–96, 1855; pp. 285–412, pls. 97–116, 1856; p. 413–540, pls. 117–128, 1857; pp. 541–661, pls. 129–138, 1858; vol. 1, pp. i–x 1858] .
- Adams A , Angas GF 1864 . Descriptions of new species of shells from the Australian seas, in the collection of George French Angas . Proceedings of the Zoological Society of London 1863 : 418 428 [April 1864; Trew 1992] .
- Adegoke , OS . 1967 . Wyattella, a new turritellid genus from the Eocene of Columbia, South America . Journal of Paleontology , 41 : 1095 – 1100 .
- Aldea , C and Valdovinos , C . 2005 . Rocky shore mollusks of south-central Chile (36°–38°S): taxonomy and key of identification . Gayana , 69 : 364 – 396 .
- Allison , RS . 1965 . Apical development in turritellid classification with a description of Cristispira pugetensis gen. et sp. nov . Palaeontology , 8 : 666 – 680 .
- Allison , RS and Adegoke , OS . 1969 . The Turritella rina group (Gastropoda) and its relationship to Torcula Gray . Journal of Paleontology , 43 : 1248 – 1266 .
- Allmon , WD . 1988 . Ecology of Recent turritelline gastropods (Prosobranchia, Turritellidae): current knowledge and paleontological implications . Palaios , 3 : 259 – 284 .
- Allmon , WD . 1992 . Role of temperature and nutrients in extinction of turritelline gastropods: Cenozoic of the northwestern Atlantic and northeastern Pacific. Palaeogeography, Palaeoclimatolgy . Palaeoecology , 92 : 41 – 54 .
- Allmon , WD . 1996 . Systematics and evolution of Cenozoic American Turritellidae (Mollusca: Gastropoda) I: Paleocene and Eocene Coastal Plain species related to “Turritella mortoni Conrad” and “Turritella humerosa Conrad” . Palaeontographica Americana , 59 : 1 – 134 .
- Allmon , WD , Jones , DS , Aiello , RL , Gowlett-Holmes , K and Probert , PK . 1994 . Observations on the biology of Maoricolpus roseus (Quoy & Gaimard) (Prosobranchia: Turritellidae) from New Zealand and Tasmania . The Veliger , 37 : 267 – 279 .
- Almond CS 1980 . The Pliocene-early Pleistocene geology of the Kaniere district, Hokitika . Unpublished MSc thesis, University of Otago . 363 p.
- Anderson , DT . 1966 . Further observations on the life histories of littoral gastropods in New South Wales . Proceedings of the Linnean Society of New South Wales , 90 : 242 – 251 .
- Angas , GF . 1865 . On the marine molluscan fauna of the Province of South Australia: with a list of all the species known up to the present time; together with remarks on their habitats and distributions, etc . Proceedings of the Zoological Society of London , 1865 : 155 – 190 .
- Angas , GF . 1867 . List of species of marine Mollusca found in Port Jackson harbour, New South Wales, and the adjacent coasts, with notes on their habits, etc. – Part I . Proceedings of the Zoological Society of London , 1867 : 185 – 233 .
- Angas , GF . 1871 . A list of additional species of marine Mollusca to be included in the fauna of Port Jackson and the adjacent coasts of New South Wales . Proceedings of the Zoological Society of London , 1871 : 87 – 101 .
- Ardovini , R and Cossignani , T . 2004 . West African seashells (including Azores, Madeira and Canary Is.) , 319 Ancona : l'Informatore Piceno .
- Arnaud , PM and Beurois , T . 1972 . Première signalisation des genres Charonia et Ranella aux Isles Saint-Paul et Amsterdam (Océan Indien) et révision du genre Argobuccinum. (Gastropoda Cymatiidae) . Tethys , 3 : 865 – 874 .
- Bartrum , JA and Powell , AWB . 1928 . Mollusca from Kaawa Creek beds, west coast, south of Waikato River . Transactions of the New Zealand Institute , 59 : 139 – 162 .
- Bax , NJ and Gowlett-Holmes , KL . 2000 . “ Invasion of the New Zealand screwshell ” . In Molluscs 2000 , Edited by: Ponder , WF . 10 Sydney : Malacological Society of Australasia .
- Bax NJ , McEnnulty FR , Gowlett-Holmes KL 2003 . Distribution and biology of the introduced gastropod, Maoricolpus roseus (Quoy and Gaimard, 1834) (Caenogastropoda: Turritellidae) in Australia. Centre for Research on Introduced Marine Pests, CSIRO Marine Research , Technical Report . Hobart, , Tasmania 25 i v , 1–32 .
- Bayer , C . 1935 . Catalogue of the Cassidae in the Rijksmuseum van Natuurlijke Historie . Zoologische Mededeelingen , 18 : 93 – 120 .
- Beu , AG . 1965 . Lower Pleistocene Mollusca from Devil's Elbow, Hawke's Bay . Transactions of the Royal Society of New Zealand Geology , 3 : 139 – 149 .
- Beu , AG . 1966 . The molluscan genera Lutraria, Resania and Zenatia in New Zealand . Transactions of the Royal Society of New Zealand Zoology , 8 : 63 – 91 .
- Beu , AG . 1967 . Deep-water Pliocene Mollusca from Palliser Bay, New Zealand . Transactions of the Royal Society of New Zealand Geology , 5 : 98 – 122 .
- Beu , AG . 1969 . Additional Pliocene bathyal Mollusca from South Wairarapa, New Zealand . New Zealand Journal of Geology and Geophysics , 12 : 484 – 496 .
- Beu , AG . 1970 . Descriptions of new species and notes on taxonomy of New Zealand Mollusca . Transactions of the Royal Society of New Zealand Earth Sciences , 7 : 113 – 136 .
- Beu , AG . 1976 . Arrival of Semicassis pyrum (Lamarck) and other tonnacean gastropods in the Southern Ocean during Pleistocene time . Journal of the Royal Society of New Zealand , 6 : 413 – 432 .
- Beu , AG . 1977 . Ages of some Chlamys delicatula localities in North Canterbury (note) . New Zealand Journal of Geology and Geophysics , 20 : 199 – 203 .
- Beu , AG . 1985a . A classification and catalogue of living world Ranellidae (= Cymatiidae) and Bursidae . Conchologists of American Bulletin , 13 : 55 – 66 .
- Beu , AG . 1995 . Pliocene limestones and their scallops. Lithostratigraphy, pectinid biostratigraphy and paleogeography of eastern North Island late Neogene limestone . Institute of Geological and Nuclear Sciences Monograph , 10 : 1 – 243 .
- Beu AG 1998 . Appendix 2. Age and environment of deposition based on macrofossils in the Bexley testbore, Christchurch Brown LJ . Bexley groundwater testbore results . Institute of Geological and Nuclear Sciences Client Report 62698D.12 . Pp. 18 22 , tables 1–3 .
- Beu , AG . 2004 . Marine Mollusca of oxygen isotope stages of the last 2 million years in New Zealand. Part 1. Revised generic positions and recognition of warm-water and cool-water migrants . Journal of the Royal Society of New Zealand , 34 : 111 – 265 .
- Beu , AG . 2005 . Neogene fossil tonnoidean gastropods of Indonesia . Scripta Geologica , 130 : 1 – 186 .
- Beu , AG . 2006 . Marine Mollusca of oxygen isotope stages of the last 2 million years in New Zealand. Part 2. Biostratigraphically useful and new Pliocene to Recent bivalves . Journal of the Royal Society of New Zealand , 36 : 151 – 338 .
- Beu , AG . 2009 . Before the ice: biogeography of Antarctic Paleogene Mollusca. Palaeogeography, Palaeoclimatology . Palaeoecology. , 234 : 191 – 226 .
- Beu , AG . 2010 . Neogene tonnoidean gastropods of tropical and South America; contributions to the Dominican Republic and Panama Paleontology Projects and uplift of the Central American Isthmus . Bulletins of American Paleontology , 377–378 : 1 – 550 .
- Beu , AG and de Rooij-Schuiling , LA . 1982 . Subgeneric classification of New Zealand and Australian species of Paphies Lesson (Bivalvia: Mesodesmatidae), and names for the two species of tuatua in New Zealand . New Zealand Journal of Zoology , 9 : 211 – 230 .
- Beu , AG and Kitamura , A . 1998 . Exposed coasts vs sheltered bays: contrast between New Zealand and Japan in the molluscan record of temperature change in Plio-Pleistocene cyclothems . Sedimentary Geology , 122 : 129 – 149 .
- Beu AG , Maxwell PA 1990 58 . Cenozoic Mollusca of New Zealand. Drawings by RC Brazier . New Zealand Geological Survey Paleontological Bulletin 1 518 . Available on-line at: http://www.gns.cri.nz/what/earthhist/fossils/BeuMaxwell1990.pdf .
- Bieler , R . 1997 . “ Stephopoma (Caenogastropoda: Siliquariidae) from the Houtman Abrolhos Islands, Western Australia; with a review of the genus ” . In The marine flora and fauna of the Houtman Abrolhos Islands , Edited by: Wells , FE . 255 – 280 . Western Australia, Perth : Western Australian Museum .
- Bieler , R and Simone , LRL . 2005 . “ Anatomy and morphology of Stephopoma nucleogranosum Verco, 1904 (Caenogastropoda: Siliquariidae) from Esperance Bay, Western Australia ” . In The marine flora and fauna of Esperance , Edited by: Wells , FE , Walker , DI and Kendrick , GA . 159 – 175 . Western Australia, Perth : Western Australian Museum .
- Boreham , AUE . 1965 . A revision of F. W. Hutton's pelecypod species described in the Catalogue of Tertiary Mollusca and Echinodermata (1873) . New Zealand Geological Survey Paleontological Bulletin , 37 : 1 – 125 .
- Bouchet , P and Poppe , G . 1988 . Deep-water Volutidae from the New Caledonian region, with a discussion on biogeography . Venus , 47 : 15 – 32 .
- Bouchet , P , Rocroi , JP , Frýda , J , Hausdorf , B , Ponder , WF , Valdés , A and Warén , A . 2005 . Classification and nomenclator of gastropod families . Malacologia , 47 : 1 – 397 .
- Bouchet , P and Warén , A . 1993 . Revision of the northeast Atlantic bathyal and abyssal Mesogastropoda . Bollettino Malacologico Supplemento , 3 : 577 – 840 .
- Bowles , E . 1939 . Eocene and Paleocene Turritellidae of the Atlantic and Gulf Coast plain of North America . Journal of Paleontology , 13 : 267 – 336 .
- Bracegirdle S 2007 . Notes on New Zealand collected species; Semicassis labiata . Cookia (Journal of the Wellington Shell Club, New Zealand) 19 : 19 .
- Bradshaw MA , Lobb AJ , Cave MP , Watson K 1992 . Catalogue of type and figured fossils in the Geology Department, Canterbury Museum . Canterbury Museum Bulletin 8 : i–iv , 1 113 .
- Brazier , JW . 1878 . Mollusca of the Chevert Expedition [part] . Proceedings of the Linnean Society of New South Wales , 2 : 143 – 145 .
- Breward , I . 1990 . “ Fraser, Charles 1823?–1886 ” . In Dictionary of New Zealand biography , Edited by: Oliver , WH . Vol. 1 , 138 – 139 . Wellington : Allen & Unwin and Department of Internal Affairs .
- Broderip , WJ . 1837 . “ Cypraeidae ” . In The Penny Cyclopedia of the Society for the Diffusion of Useful Knowledge , Edited by: Long , G . Vol. 8 , 254 – 259 . London : C Knight & Co. [not seen] .
- Bruguière JG 1792 . Encyclopédie méthodique. Histoire naturelle des vers. Tome premier [part 2] . , Paris, Panckoucke , 345 757 .
- Bucknill , CER . 1924 . Sea shells of New Zealand. Illustrated with original drawings by A. W. B. Powell and photographs from nature , 123 Auckland : Whitcombe and Tombs .
- Campbell HJ , Andrews PB , Beu AG , Maxwell PA , Edwards AR , Laird MG , Hornibrook NdeB , Mildenhall DC , Watters WA , Buckeridge JS , Lee DE , Strong CP , Wilson GJ , Hayward BW 1994 2 . Cretaceous-Cenozoic geology and biostratigraphy of the Chatham Islands, New Zealand, Institute of Geological and Nuclear Sciences Monograph 1 269
- Carcelles , AR . 1953 . Nuevas especies de gastropodos mariños de las republicas oriental del Uruguay y Argentina . Comunicaciones Zoologicas del Museo de Historia Natural de Montevideo , 4 ( 70 ) : 1 – 16 .
- Carter RM , Naish TR 1999 . The high-resolution, chronostratigraphic and sequence stratigraphic record of the Plio-Pleistocene, Wanganui Basin, New Zealand . Institute of Geological and Nuclear Sciences Folio Series 2: folder with two wall charts .
- Cernohorsky , WO . 1968 . Observations on Hipponix conicus (Schumacher, 1817) . The Veliger , 10 : 275 – 280 .
- Cernohorsky , WO . 1971 . Fossil and Recent Cypraeacea (Mollusca: Gastropoda) of New Zealand with descriptions of new species . Records of the Auckland Institute and Museum , 8 : 103 – 129 .
- Cernohorsky , WO . 1972 . Type specimens of Recent and fossil Mollusca described by H. J. Finlay. Part 1. (Scissurellidae – Turbinidae) . Records of the Auckland Institute and Museum , 9 : 231 – 247 .
- Chemnitz JH 1795 11 . Neues systematisches Conchylien-Cabinet, fortgesetzt und nun vollendet von Johann Heironymus Chemnitz … , Nürnberg, der Raspischen Buchhandlung . 20 + 310 p., pls. 174 213 .
- Chenu JC 1859 . Manuel de conchyliologie et de paléontologie conchyliologique. Tome premier . Paris , V. Masson vii + 327
- Clessin S 1901–1912 . Die Familie Vermetidae Küster HC , Kobelt W Systematisches Conchylien-Cabinet von Martini und Chemnitz … neu herausgegeben und vervollständigt von Dr. H. C. Küster. Nach dessen Tode fortgesetzt von Dr. W. Kobelt. Nürnberg, Bauer & Raspe Vol. 6(6), pp. 1 124 [dates (Welter-Schultes 1999): pp. 1–24, pls. 1–12, 1901; pp. 25–80, pl. 13, 1902; pp. 81–104, 1903; pls. 14, 15, 1904; pp. 105–124, 1912] .
- Coleman , N . 1975 . What shell is that? Dee Why West , 308 New South Wales : Paul Hamlyn .
- Collin , R . 2003 . Phylogenetic relationships among calyptraeid gastropods and their implications for biogeography of marine speciation . Systematic Biology , 52 : 618 – 640 .
- Collin , R . 2006 . Sex ratio, life-history invariants, and patterns of sex change in a family of protandrous gastropods . Evolution , 60 : 735 – 745 .
- Cossmann , M . 1904 . Essais de paléoconchologie comparée , Vol. 6 , 261 Paris : M. Cossmann .
- Cotton , BC . 1945 . Southern Australian Gastropoda. Part II. Doliacea . Transactions of the Royal Society of South Australia , 69 : 249 – 262 .
- Cotton , BC . 1959 . South Australian Mollusca. Archaeogastropoda. Handbook of the fauna and flora of South Australia , 449 Adelaide : South Australian Government Printer .
- Cotton , BC and Godfrey , FK . 1934 . South Australian shells (including descriptions of new genera and species). Part XI. Trochidae . The South Australian Naturalist , 15 : 77 – 92 .
- Cotton BC , Godfrey FK 1938 . A systematic list of the Gastropoda . The marine, freshwater and land univalve Mollusca of South and central Australia. Malacological Society of South Australia Publication 1 . 44 p.
- Crosse , H . 1862 . Note sur l’Hipponix danieli, et catalogue des espèces actuellement connues . Journal de Conchyliologie , 10 : 17 – 28 .
- Dall WH 1921 . Species named in the Portland Catalogue . The Nautilus 34 97 100 , 124–132
- Darragh , TA . 1970 . Catalogue of Australian Tertiary Mollusca (except chitons) . Memoirs of the National Museum of Victoria , 31 : 125 – 212 .
- Delessert B 1841 . Recueil de coquilles décrites par Lamarck et non encore figurées . Paris, J. Rothschild; Leipzig, L. A. Kittler ; Williams & Norgate , London . 40 pls. with unpaginated captions, [93] p .
- Dell , RK . 1952 . A revision of the molluscan fauna of the Hurupi beds, southern Wairarapa . Dominion Museum Records in Zoology , 1 : 71 – 86 .
- Dell RK 1955 . Nature in New Zealand. Native shells . AH & AW Reed , Wellington . 64 p. [reprinted 1957, 1960] .
- Dell , RK . 1963a . Notes on some New Zealand Mollusca in the British Museum . Transactions of the Royal Society of New Zealand Zoology , 3 : 171 – 177 .
- Dell , RK . 1963b . The Mollusca of the genus Argobuccinum (Family Cymatiidae) . Transactions of the Royal Society of New Zealand Zoology , 3 : 217 – 228 .
- Dell , RK , Fleming , CA , Marwick , J and Powell , AWB . 1964 . Turritella kanieriensis G. F. Harris, 1897 (Mollusca): proposed designation of a type-specimen under the plenary powers . Bulletin of Zoological Nomenclature , 21 : 447 – 450 .
- Dell , RK and Ponder , WF . 1964 . Notes on the gastropod, Zelippistes benhami (Suter) . Records of the Dominion Museum , 5 : 63 – 72 .
- Deshayes GP 1830–1832 . Encyclopédie méthodique, ou par ordre de matières; par une société de gens de letters, de savants et d'artistes … Histoire naturelle des vers vol. 2(2) Madame Veuve Agasse , Paris 1 594 [1–144, 1830; 145–594, 1832; Evenhuis (2003); Evenhuis & Petit (2003)] .
- Deshayes GP 1835–1845 . Histoire naturelle des animaux sans vertèbres, présentant les charactères généraux et particuliers de ces animaux … . par J. B. P. A. de Lamarck. Deuxième édition … Histoire naturelle des mollusques. Paris, J. B. Baillière. Vol. 6, 1835, iv + 600 p.; vol. 7, 1836, vi + 735 p.; vol. 8, 1838, 660 p.; vol. 9, 1843, 728 p.; vol. 10, 1844, 638 p.; vol. 11, 1845, 665 p. [vol. 6, p. i states that Deshayes is the author of Mollusca] .
- DeVries , TJ . 2007 . Cenozoic Turritellidae from southern Peru . Journal of Paleontology , 81 : 331 – 351 .
- Dillwyn LW 1817 . A descriptive catalogue of Recent shells, arranged according to the Linnaean method; with particular attention to the synonymy . London, J. & A. Arch. Vol. 1, xii + 580 p.; vol. 2, pp 581 1092 + [23].
- Dolin , L . 2001 . Les Triviidae (Mollusca: Caenogastropoda) de l'Indo-Pacifique: Révision des genres Trivia, Dolichupis et Trivellona. In: Bouchet P, Marshall BA ed. Tropical deep-sea benthos, 22 . Mémoires du Muséum National d'Histoire Naturelle , 185 : 201 – 241 .
- Dollfus , GF . 1926 . Critique de classification des turritelles . Bulletin de la Société Géologique de France, Série , 26 ( 4 ) : 27 – 32 .
- Donald , KM , Kennedy , M and Spencer , HG . 2005 . The phylogeny and taxonomy of austral monodontine topshells (Mollusca: Gastropoda: Trochidae), inferred from DNA sequences . Molecular Phylogenetics and Evolution , 37 : 474 – 483 .
- Douvillé , H . 1929 . Les couches à Cardita beaumonti: Fascicule II. Les couches à Cardita beaumonti dans le Sind . Memoirs of the Geological Survey of India, Palaeontologica Indica, new series , 10 ( 3 ) : 26 – 73 .
- Dunker , W . 1862 . Species nonnulae Bursarum vel Ranellarum collectionis Cumingianae . Proceedings of the Zoological Society of London , 1862 : 238 – 240 .
- Dunker W 1863–1864 . Novitates conchologicae. Mollusca marina. Beschreibung und Abbildung neuer oder wenig gekannter Meeres-Conchylien . Cassel, T. Fischer. Part 6, pp. 51–58, pls. 16–18, 1863; part 7, pp. 59–66, pls. 19–21, 1864 [dates: Johnson (1969)] .
- Finger , KL , Nielsen , SN , DeVries , TJ , Encinas , A and Peterson , DE . 2007 . Paleontological evidence for sedimentary displacement in Neogene forearc basins of central Chile . Palaios , 22 : 3 – 16 .
- Finlay , HJ . 1924a . New shells from New Zealand Tertiary beds . Transactions of the New Zealand Institute , 55 : 480 – 494 .
- Finlay , HJ . 1924b . The molluscan fauna of Target Gully: Part 1 . Transactions of the New Zealand Institute , 55 : 495 – 516 .
- Finlay , HJ . 1924c . Additions to the Recent molluscan fauna of New Zealand . Transactions of the New Zealand Institute , 55 : 517 – 526 .
- Finlay , HJ . 1924d . Some necessary changes in names of New Zealand Mollusca . Proceedings of the Malacological Society of London , 16 : 99 – 107 .
- Finlay , HJ . 1926a . New shells from New Zealand Tertiary beds: Part 2 . Transactions of the New Zealand Institute , 56 : 227 – 258 .
- Finlay HJ 1926b . A further commentary on New Zealand molluscan systematics . Transactions of the New Zealand Institute 56 : 320 485 . [‘1927’; issued 23 December 1926]
- Finlay , HJ . 1928 . The Recent Mollusca of the Chatham Islands . Transactions of the New Zealand Institute , 59 : 232 – 286 .
- Finlay , HJ and Marwick , J . 1937 . The Wangaloan and associated faunas of Kaitangata-Green Island Subdivision . New Zealand Geological Survey Paleontological Bulletin , 15 : 1 – 140 .
- Fischer P 1875–1880 . Spécies générale et iconographie des coquilles vivantes, comprenant la collection du Muséum d'Histoire naturelle de Paris, la collection Lamarck, celle du Prince Masséna, (appartenant maintenant à M. le baron Benjamin Delessert), et les découvertes récentes des voyageurs, par L.-C. Kiener … continué par … P. Fischer . Vol. 11. Genres Calcar, Trochus, Xenophora, Tectarius et Risella . Paris, J.-B. Baillière et fils. iii + 480 p. [dates (Sherborn & Woodward 1901): pp 1–96, 1875; 97–144, 1876; 145–240, 1877; 241–336, 1878; 337–463, 1879; index 1880] .
- Fischer P 1880–1887 . Manuel de conchyliologie et de paléontologie conchyliologique, ou histoire naturelle des mollusques vivants et fossiles Paris, F. Savy. xxiv + 1369 p. [fasc. 1, pp. 1–112, 1880; fasc. 2, 3, pp. 113–304, 1881; fasc. 4, pp. 305–416, 1882; fasc. 5, 6, pp. 417–608, 1883; fasc. 7, pp. 609–688, 1884; fasc. 8, 9, pp. 689–896, 1885; fasc. 10, pp. 897–1008, 1886; fasc. 11, pp. 1009–1369, 1887] .
- Fischer von Waldheim G Muséum-Demidoff (mis en ordre systématique et décrit par G. Fischer) ou catalogue … des curiosités de la nature et de l'art, données à l'Université impériale de Moscou par … P. de Demidoff. Moscow, 3, Végéteaux et animaux iv + 330p [not seen; from Woodward p. 578 1807 1904
- Fleming , CA . 1943 . Tertiary Mollusca from Dannevirke Subdivision . Transactions of the Royal Society of New Zealand , 73 : 193 – 208 .
- Fleming , CA . 1944 . Molluscan evidence of Pliocene climatic change in New Zealand . Transactions of the Royal Society of New Zealand , 74 : 207 – 220 .
- Fleming , CA . 1947 . Standard sections and subdivisions of the Castlecliffian and Nukumaruan Stages in the New Zealand Pliocene . Transactions of the Royal Society of New Zealand , 76 : 300 – 326 .
- Fleming , CA . 1952 . A Foveaux Strait oyster bed . New Zealand Journal of Science and Technology , B34 : 73 – 85 .
- Fleming , CA . 1953 . The geology of Wanganui Subdivision, Waverley and Wanganui sheet districts (N137 and N138) . New Zealand Geological Survey Bulletin , 52 : 1 – 362 .
- Fleming , CA . 1966 . Marwick's illustrations of New Zealand shells, with a checklist of New Zealand Cenozoic Mollusca . New Zealand Department of Scientific and Industrial Research Bulletin , 173 : 1 – 456 .
- Frauenfeld , GH Ritter von . 1865 . Zoologische Miscellan. VI. I. Über zwei Meeresschnecken von St. Paul. Verhandlungen der K.K . Zoologisch-Botanischen Gessellschaft in Wien , 15 : 893 – 902 .
- Frauenfeld , GH Ritter von . 1867 . Mollusken. Novara-Expedition . Zoologischer Theil , 2 : 1 – 16 .
- Gammon PR 1997 . Sequence stratigraphy of the Mangaopari Basin, southeastern North Island, New Zealand . Unpublished PhD thesis, Townsville, James Cook University of North Queensland 1 , 170 p.; vol. 2, 282 p.
- Garrard , TA . 1972 . A revision of the Australian Recent and Tertiary Turritellidae (Gastropoda: Mollusca) . Journal of the Malacological Society of Australia , 2 : 267 – 338 .
- Garrard , TA . 1974 . The genus Haustator, subgenus Kurosioia in Australian waters (Turritellidae: Mollusca) . Journal of the Malacological Society of Australia , 3 : 15 – 17 .
- Garrard , TA . 1976 . A supplement to Australian Recent Turritellidae (Gastropoda: Mollusca) . Journal of the Malacological Society of Australia , 5 : 195 – 200 .
- Gmelin JF 1791 . C. a Linné … Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis … editio decima tertia, aucta, reformata, cura J. F. Gmelin. Tomus I, pars VI, Vermes testacea . GE Beer Lipsiae 3021 4120 .
- Goldstien , SJ , Gemmell , NJ and Schiel , DR . 2006 . Molecular phylogenetics and biogeography of the nacellid limpets of New Zealand (Mollusca: Patellogastropoda) . Molecular Phylogenetics and Evolution , 38 : 261 – 265 .
- Golikov AN 1986 . On the systematics and evolution of gastropods of the family Turritellidae from the cold and temperate waters of the Northern Hemisphere . Zoologicheskii Zhurnal 65 : 1140 1150 [in Russian] .
- Graham , J . 1962 . The North Otago shelf fauna Part I – Mollusca . Transactions of the Royal Society of New Zealand Zoology , 2 : 53 – 60 .
- Grant-Mackie , JA . 1990 . “ Stacking and substrate preservation in a New Zealand Pliocene Crepidula (Mesogastropoda) ” . In Evolutionary paleobiology of behavior and coevolution , Edited by: Boucot , AJ . 398 – 404 . Amsterdam : Elsevier. Pp .
- Grant-Mackie , JA and Chapman-Smith , M . 1971 . Paleontological notes on the Castlecliffian Te Piki bed, with descriptions of new molluscan taxa . New Zealand Journal of Geology and Geophysics , 14 : 655 – 704 .
- Gray , JE . 1835 . “ Catalogue of shells collected on the east coast of New Zealand, by the Rev. W. Yate ” . In An account of New Zealand;and of the formation and progress of the Church Missionary Society's mission in the Northern Island , Edited by: Yate , W. 307 – 310 . London : Seeley and Burnside. Pp .
- Gray JE 1839 . Molluscous animals, and their shells Beechey FW The zoology of Captain Beechey's voyage; compiled from the collections and notes made by Captain Beechey, the officers and naturalist of the expedition, during a voyage to the Pacific and Behring's Straits performed in His Majesty's Ship Blossom, under the command of Captain F. W. Beechey, R.N., F.R.S., &c. &c. in the years 1825, 26, 27, and 28, by J. Richardson; … N. A. Vigors; … G. T. Lay, Esq.; E. T. Bennett; … Richard Owen, Esq.; John E. Gray;… the Rev. W. Buckland;… and G. B. Sowerby, Esq. Illustrated by upwards of fifty finely coloured plates by Sowerby. Published under the authority of the Lords Commissioners of the Admiralty. London, HG Bohn. Pp. 102–142, pls. 33–44 [pp. 143–155, continued by GB Sowerby I] .
- Gray JE 1840 . [Mollusca] Synopsis of the contents of the British Museum , edition 42 British Museum , London 105 152 [first issue, ‘ed. 42A’, 16 October 1840; second issue, ‘ed. 42B’, 4 November 1840, pp. 106–156]
- Gray , JE . 1841 . [Mollusca]. In: Synopsis of the contents of the British Museum , edition 43 , 78 – 130 . London : British Museum .
- Gray JE 1842 . [Mollusca]. In: Synopsis of the contents of the British Museum , edition 44 British Museum , London . Pp. 48 92 [ 21 May 1842 ].
- Gray JE 1843 . Catalogue of the species of Mollusca and their shells, which have hitherto been recorded as found at New Zealand, with the description of some lately discovered species Dieffenbach E . Travels in New Zealand; with contributions to the geography, geology, botany and natural history of that country , vol. 2. J. Murray , London . Pp. 228 265 [reprint, Christchurch, Capper Press, 1975] .
- Gray , JE . 1847 . A list of the genera of Recent Mollusca, their synonyma and types . Proceedings of the Zoological Society of London , 15 : 129 – 219 .
- Gray JE 1850 4 . Figures of molluscous animals, selected from various authors. Etched for the use of students by Maria Emma Gray Explanation of plates and list of genera . Longman , London, Brown, Green and Longmans iv + 124 p.
- Gray JE 1853a . A revision of the genera of some of the families of Conchifera or bivalve shells . The Annals and Magazine of Natural History series 2 11 33 44 , 398–402, [January 1853] .
- Gray JE 1853b . Description of a new genus of bivalve shell (Vanganella) and a sea-egg from New Zealand . Proceedings of the Zoological Society of London 19 : 125 126 [29 April 1853] .
- Gray JE 1857 . Guide to the systematic distribution of Mollusca in the British Museum. Part I London , British Museum xii + 230 p.
- Gray ME 1842–1857 . Figures of molluscous animals, selected from various authors. Etched for the use of students . Longman , London, Brown, Green and Longmans . Vol. 1, 1842, pls. 1–78; vol. 2, 1850, pls. 79–199; vol. 3, 1850, pls. 200–312 [vol. 4, by JE Gray, 1850, see above]; vol. 5, 1857, Conchifera and Brachiopoda, pls. 313–381; Explanation of the plates [by JE Gray], pp. 1–13; Systematic arrangement of the figures (continued from vol. IV) [by JE Gray], pp. 15–49 [also issued in a 5-volume edition in 1857] .
- Greenhill , JF . 1965 . New records of marine Mollusca from Tasmania . Papers and Proceedings of the Royal Society of Tasmania , 99 : 67 – 69 .
- Guillaume , L . 1925 . Essai sur la classification des turritelles, ainsi que sur leur evolution et leur migrations, depuis le début des temps tertiaires . Bulletin de la Société Géologique de France, 4me série , 24 : 281 – 311 .
- Hadfield , MG and Strathman , MG . 1990 . Heterostrophic shells and pelagic development in trochoideans: implications for classification, phylogeny and palaeoecology . Journal of Molluscan Studies , 56 : 239 – 256 .
- Hardy , GH . 1915 . List of the Tenison Woods types of Recent Mollusca in the Tasmanian Museum . Papers and Proceedings of the Royal Society of Tasmania for , 1915 : 61 – 73 .
- Harris GF 1897 . Catalogue of Tertiary Mollusca in the Department of Geology, British Museum (Natural History). Part I. The Australasian Tertiary Mollusca . British Museum (Natural History) , London xxvi + 407 p.
- Hart M 1996 . New species of Trivia and Erato (Gastropoda: Triviidae) from northern New Zealand waters . La Conchiglia 28 279 37 39 62.
- Hartley , T . 1958 . Movement by commensal Hipponix conicus (Schumacher) . Journal of the Malacological Society of Australia , 1 ( 2 ) : 37 – 38 .
- Hector J 1886 . Indian and Colonial Exhibition, London, 1886. New Zealand court. Detailed catalogue and guide to the geological exhibits . Government Printer , Wellington . 101 p.
- Hedley , C . 1901 . A revision of the types of the marine shells of the “Chevert” expedition . Records of the Australian Museum , 4 : 121 – 130 .
- Hedley , C . 1902a . Studies on Australian Mollusca. Part VI . Proceedings of the Linnean Society of New South Wales , 1902 : 7 – 29 .
- Hedley , C . 1902b . Studies on Australian Mollusca. Part VII . Proceedings of the Linnean Society of New South Wales , 1902 : 596 – 619 .
- Hedley , C . 1913 . Studies on Australian Mollusca. Part XI. Notes on museums abroad . Proceedings of the Linnean Society of New South Wales , 38 : 258 – 339 .
- Hedley , C . 1916a . A preliminary index of the Mollusca of Western Australia . Journal of the Royal Society of Western Australia , 1 : 3 – 77 .
- Hedley , C . 1916b . Studies on Australian Mollusca. Part XIII . Proceedings of the Linnean Society of New South Wales , 41 : 680 – 719 .
- Hedley C 1918 . A check-list of the marine fauna of New South Wales. Part 1 . Mollusca. Journal of the Royal Society of New South Wales 51 , Supplement: M1 M120 .
- Hedley , C and May , WL . 1908 . Mollusca from one hundred fathoms, seven miles east of Cape Pillar, Tasmania . Records of the Australian Museum , 7 : 108 – 125 .
- Henning , T and Hemmen , J . 1993 . Ranellidae and Personidae of the world , 263 Wiesbaden : C. Hemmen .
- Herm , D . 1969 . Marines Pliozän und Pleistozän in Nord- und Mittel-Chile unter besonderer Berücksichtigung der Entwicklung der Mollusken-Faunen. Zitteliana . Abhandlungen der Bayerischen Staatssammlung für Paläontologie und Historische Geologie , 2 : 1 – 159 .
- Herrmannsen AN 1846–1852 . Indicus generum malacozoorum primordia. Nomina subgenerum, generum, familiarum, tribuum, ordinum, classium: adjectis auctoribus, temporibus, locis systematicis atque literariis, etymus, synonymis . Praetermittuntur Cirripedia, Tunicata et Rhizopoda. Cassell, T. Fischer. Vol. 1, pp. 1–232, 1846; 233–637, 1847; vol. 2, pp. 1–352, 1847; 353–492, 1848; 493–717, 1949; supplement, v + 140 p., 1852 .
- Hickman , CS . 1992 . Reproduction and development of trochacean gastropods . The Veliger , 35 : 245 – 272 .
- Higo , S , Callomon , P and Goto , Y . 1999 . Catalogue and bibliography of the marine shell-bearing Mollusca of Japan. Gastropoda. Bivalvia. Polyplacophora. Scaphopoda , 749 Osaka : Elle Scientific Publications .
- Hoagland , KE . 1977 . Systematic review of fossil and Recent Crepidula and discussion of evolution of the Calyptraeidae . Malacologia , 16 : 353 – 420 .
- Hoagland , KE . 1983 . Notes on type specimens of Crepidula (Prosbranchia: Calyptraeidae) in the Muséum national d'Histoire naturelle, Paris . Proceedings of the Academy of Natural Sciences of Philadelphia , 136 : 1 – 8 .
- Hornibrook , N and de , B . 1982 . Tom Grant-Taylor [obituary] . Geological Society of New Zealand Newsletter , 58 : 80 – 84 .
- Hutton FW 1873a . Catalogue of the marine Mollusca of New Zealand, with diagnoses of the species . Wellington , Colonial Museum and Geological Survey Department Natural History Publication 5. xx + 116 p. [apparently reprinted in 1874; Beu (2006:325)] .
- Hutton FW 1873b . Catalogue of the Tertiary Mollusca and Echinodermata of New Zealand, in the collection of the Colonial Museum . Colonial Museum and Geological Survey Department Natural History Publication 6 , Wellington xvi + 48 p.
- Hutton , FW . 1878 . Révision des coquilles de la Nouvelle-Zélande et des îles Chathams . Journal de Conchyliologie , 24 : 5 – 57 .
- Hutton FW 1880 . Manual of the New Zealand Mollusca. A systematic and descriptive catalogue of the marine and land shells, and of the soft Mollusca and Polyzoa of New Zealand and the adjacent islands . Colonial Museum and Geological Survey Department , Wellington xvi + iv + 224 p.
- Hutton , FW . 1882 . Notes on some branchiate Mollusca . Transactions of the New Zealand Institute , 14 : 162 – 167 .
- Hutton FW 1885 . Descriptions of new Tertiary shells [parts 1 and 2] . Transactions of the New Zealand Institute 17 : 313 324 , 325–332 .
- Hutton , FW . 1887 . The Mollusca of the Pareora and Oamaru Systems of New Zealand . Proceedings of the Linnean Society of New South Wales , 17 : 205 – 237 .
- Hutton , FW . 1893 . “ The Pliocene Mollusca of New Zealand ” . In The MacLeay Memorial Volume , Edited by: Fletcher , JJ . 35 – 92 . Sydney : Linnean Society of New South Wales .
- Hutton FW 1904 . Index faunae Novae Zelandiae . Dulau & Co for the Philosophical Institute of Canterbury , London viii + 372 p. [Mollusca compiled by H Suter, pp. 57–95] .
- Ida , K . 1952 . A study of fossil Turritella in Japan . Geological Survey of Japan Report , 150 : 1 – 62 .
- International Commission on Zoological Nomenclature 1957 . Opinion 479. Validation under the plenary powers of specific names for nine species of the Class Gastropoda occurring in the New Zealand area as published by Martyn (T.) in 1784 in the work entitled The Universal Conchologist (Opinion supplementary to Opinion 456) . Opinions and Declarations Rendered by the International Commission on Zoological Nomenclature 16 : 365 416 .
- International Commission on Zoological Nomenclature 1967 . Opinion 810. Turritella kanieriensis G. F. Harris, 1897 (Gastropoda): designation of a lectotype under the plenary powers . Bulletin of Zoological Nomenclature 24 : 87 88 .
- Iredale , T . 1912 . New generic names and new species of marine Mollusca . Proceedings of the Malacological Society of London , 10 : 217 – 228 .
- Iredale , T . 1915 . A commentary on Suter's “Manual of the New Zealand Mollusca” . Transactions of the New Zealand Institute , 47 : 417 – 497 .
- Iredale , T . 1924 . Results from Roy Bell's molluscan collections . Proceedings of the Linnean Society of New South Wales , 49 : 179 – 278 .
- Iredale , T . 1927 . A review of Australian helmet shells (Family Cassididae – Phylum Mollusca) . Records of the Australian Museum , 15 : 321 – 354 .
- Iredale , T . 1931 . Australian molluscan notes. No. 1 . Records of the Australian Museum , 18 : 210 – 235 .
- Iredale , T . 1936 . Australian molluscan notes. No. 2 . Records of the Australian Museum , 19 : 267 – 340 .
- Iredale , T . 1937 . The Middleton & Elizabeth Reefs, South Pacific Ocean. Mollusca . The Australian Zoologist , 8 : 232 – 261 .
- Iredale T 1956 . History of New South Wales shells. Part 1: Cook and his associates . The Marine Zoologist 1(4), issued in: Proceedings of the Royal Zoological Society of New South Wales 1954–55 : 81 83 .
- Iredale , T and McMichael , DF . 1962 . A reference list of the marine Mollusca of New South Wales . Australian Museum Memoir , 11 : 1 – 109 .
- Ivanov , DL and Kantor , Yu . 1991 . Paul Demidoff's malacological collection in the Zoological Museum of Moscow University , 95 Moscow : Moscow University Press .
- Ivanov , DL , Kantor , YuI , Sysoev , AV and Egorov , RV . 1993 . Type specimens of Mollusca described by G. Fischer von Waldheim in 1807 . The Apex , 8 : 71 – 94 .
- Jansen , P . 1995 . A review of the genus Clanculus Montfort, 1810 (Gastropoda: Trochidae) in Australia, with description of a new subspecies and the introduction of a nomen novum . Vita Marina , 43 : 39 – 62 .
- Jousseaume , F . 1884 . Étude sur la famille des Cypraeidae . Bulletin de la Société Zoologique de France , 9 : 81 – 100 .
- Jousseaume , F . 1888 . Description des mollusques receuillis per M. le Dr Faurot dans le Mer Rouge et le Golfe d'Aden . Mémoires de la Société Zoologique de France , 1 : 165 – 223 .
- Kantor , Yu and Sysoev , AV . 2002 . On the species of molluscs from Russian waters described by Valenciennes in little known publications in 1858 . Ruthenica , 12 : 119 – 123 .
- Kantor Yu , Sysoev AV 2006 . Marine and brackish water Gastropoda of Russia and adjacent countries: an illustrated catalogue . Moscow , A. N. Severtsov Institute of Ecology and Evolution, Russian Academy of Sciences . 371 p. [in English and Russian] .
- Kershaw , RC . 1955 . A systematic list of the Mollusca of Tasmania, Australia . Papers and Proceedings of the Royal Society of Tasmania , 89 : 289 – 355 .
- Kesteven , HL . 1902 . The systematic position of the genus Fossarina, A. Adams and Angas, and of Fossarina varia, Hutton . Records of the Australian Museum , 4 : 317 – 322 .
- Kiener LC 1835 8 . Spécies générale et iconographie des coquilles vivantes, comprenant la collection du Muséum d'Histoire naturelle de Paris, la collection Lamarck, celle du Prince Masséna, (appartenant maintenant à M. le baron Benjamin Delessert), et les découvertes récentes des voyageurs . Famille des purpurifères . Famille des canalifères. Genre casque (Cassis, Lamarck). Paris, Rousseau, and J. B. Baillière . Pp. 1 40 pls. 1–16.
- Kiener LC 1838–1839a . [As above]. Vol. 9. Famille des canalifères. Genre struthiolaire. (Struthiolaria, Lam.) . Pp. 1 6 , pls. 1, 2 [issued in livr. 27–34, during 1838–1839, not further resolvable; Sherborn & Woodward 1901] .
- Kiener LC 1841 . [As above] Vol. 7. Famille des canalifères. Genre ranelle (Ranella, Lam.) . Pp. 1 40 , pls. 1–15 [dates, Sherborn & Woodward 1901: plates and captions 1841, text 1842] .
- Kiener LC 1842 . [As above]. Vol. 5. Famille des canalifères. Genre cérite (Cerithium, Adanson) . Pp. 1 104 pls. 1–32.
- Kilburn , RN . 1976 . A revision of the Naticidae of southern Africa and Moçambique (Mollusca) . Annals of the Natal Museum , 22 : 829 – 884 .
- Kilburn , RN . 1980 . Taxonomic studies on the marine Mollusca of southern Africa and Mozambique. Part 2 . Annals of the Natal Museum , 24 : 193 – 200 .
- Kilburn , RN . 1984 . Cymatiidae of South Africa. The Strandloper . Bulletin of the Conchological Society of Southern Africa , 218 : 1 – 5 .
- Kilburn , RN and Rippey , E . 1982 . Sea shells of southern Africa , Johannesburg : MacMillan South Africa . ix + 249
- King , LC . 1933 . Tertiary molluscan faunas from the southern Wairarapa . Transactions of the New Zealand Institute , 63 : 334 – 354 .
- King , LC . 1934 . The geology of the lower Awatere District, Marlborough, New Zealand . New Zealand Department of Scientific and Industrial Research Geological Memoir , 2 : 1 – 49 .
- King , PP . 1832 . Description of the Cirrhipedia, Conchifera and Mollusca, in a collection formed by the officers of H.M.S. Adventure and Beagle employed between the years of 1826 and 1830 in surveying the coasts of South America, including the Straits of Magalhaens and the coasts of Tierra del Fuego . The Zoological Journal , 5 : 322 – 349 . (assisted by Broderip WJ)
- Kingma , JT . 1971 . Geology of Te Aute subdivision . New Zealand Geological Survey Bulletin , 70 : 1 – 173 .
- Kirk , TW . 1882 . Description of new Tertiary fossils . Transactions of the New Zealand Institute , 14 : 409
- Knudsen , J . 1991 . “ Observations on Hipponix australis (Lamarck, 1819) (Mollusca, Gastropoda, Prosobranchia) from the Albany area, Western Australia ” . In Proceedings of the third international marine biological workshop: The marine flora and fauna of Albany, Western Australia , Edited by: Wells , FE , Walker , DI , Kirkman , H and Lethbridge , R . 641 – 660 . Perth : Western Australian Museum .
- Kotaka , T . 1959 . The Cenozoic Turritellidae of Japan. Science Reports of the Tohoku University, Sendai, Japan . Second Series (Geology) , 31 ( 2 ) : 1 – 135 .
- Kotaka , T . 1978 . World-wide biostratigraphic correlation based on turritellid phylogeny . The Veliger , 21 : 189 – 196 .
- Krauss CFF 1848 . Die südafrikanischen Mollusken. Ein Beitrag zur Kenntniss der Mollusken des Kap- und Natallandes und zur geographischen Verbreitung derselben, mit Beschreibung und Abbildung der neuen Arten . Ebner & Seubert , Stuttgart . 2 + 140 p.
- Kreipl , K . 1997 . Recent Cassidae , 151 Wiesbaden : C Hemmen .
- Kuroda T , Habe T , Oyama K 1971 . The sea shells of Sagami Bay collected by His Majesty the Emperor of Japan . Tokyo , Maruzen . xvi + 741 + 489 + 51 p.
- Küster HC , Kobelt W 1839–1878 . Die geschwänzten und bewehrten Pupurschnecken (Murex, Ranella, Tritonium, Trophon, Hindsia) . Begonnen von Dr. H.C. Küster [1839–1856], forgesetzt und beendet von Dr. W. Kobelt [1868–1878]. Systematisches Conchylien-Cabinet von Martini und Chemnitz, fortgesetzt von Hofrath Dr. G.H.V. Schubert und Professor J.A. Wagner. Neu heruasgegeben und vervollständigt von Dr. H.C. Küster…Bauer & Raspe, Nürnberg. Vol. 3(2): 1–99, 110–336, pls. A, B, 1–77, 4b, 37a, 38a, 39a [dates (Welter-Schultes 1999): pls 1–12, 1839; pp 1–8, pls 14–18, 1843; pp 9–12, pls 19, 40–50, 1844; pls A, 13, 38, 40, 41, 1855; pp 13–52, 1856; pp 53–60, pls 20–24, 1868; pp 61–99, 110–118, pls 25–35, 37, 1869; pp 119–134, 1870; pp 135–190, pls 36, 37a, 38a, 39, 39a, 51–55, 57, 1871; pp 191–206, pls 56, 58–61, 1872; pp 207–238, pls 62–67, 1876; pp 239–336, pls 68–77, 1878; pls B, 4b, possibly 1855] .
- Lamarck JBPA de M de 1816 . Tableau encyclopédique et méthodique des trois règnes de la nature … Vingt-troisième partie. Mollusques et polypes divers . Mme Veuve Agasse , Paris . Liste des objets représentés dans les planches de cette livraison, 16 p.; pls. 391–488, 431 bis, 431 bis* [14 December 1816; dates and parts: Evenhuis (2003), Evenhuis & Petit (2003)] .
- Lamarck JBPA de M de 1819–1822b . Histoire naturelle des animaux sans vertèbres, présentant les characters généraux et particuliers de ces animaux … Paris, chez l'auteur . Vol. 6, part 1, 1819, 343 p.; part 2, 1822a (April), 252 p.; vol. 7, 1822b (August), 711 p. [dates: Sherborn 1922, p. lxxvii] .
- Landau , BM , Beu , AG and Marquet , R . 2004 . The early Pliocene Gastropoda (Mollusca) of Estepona, southern Spain. Part 5: Tonnoidea, Ficoidea . Palaeontos , 5 : 35 – 102 .
- Laws , CR . 1936 . The Waitotaran faunule at Kaawa Creek, Part 2 . Transactions of the Royal Society of New Zealand , 66 : 99 – 124 .
- Laws , CR . 1940a . The Waitotaran faunule at Kaawa Creek – Part 3 . Transactions of the New Zealand Institute , 69 : 427 – 447 .
- Laws , CR . 1940b . Palaeontological study of Nukumaruan and Waitotaran rocks near Wanganui . Transactions of the Royal Society of New Zealand , 70 : 34 – 56 .
- Laws , HM . 1970 . Reproductive biology and shell site preferences in Hipponix conicus (Schumacher) (Gastropoda: Hipponicidae) . The Veliger , 13 : 115 – 121 .
- Lesson , RP . 1841 . Description de quelques mollusques nouveaux . Annales des Sciences Naturelles, Série2 , 16 : 253 – 256 .
- Lewis , DW . 1976 . Subaqueous debris flows of early Pleistocene age at Motunau, North Canterbury, New Zealand . New Zealand Journal of Geology and Geophysics , 19 : 535 – 567 .
- Lightfoot JT 1786 . Catalogue of the Portland Museum, lately the property of the Duchess Dowager of Portland, deceased: which will be sold at auction, by Mr. Skinner and Co. on Monday the 24th of April, 1786, and the thirty-seven following days, at twelve o'clock, Sundays, and the 5th of June, (the day his Majesty's birth-day is kept) excepted, at her dwelling-house, in Privy-Garden, Whitehall; by order of the acting executrix . London : Skinner and Co . 194 p.
- Lillie , AR . 1953 . The geology of the Dannevirke Subdivision . New Zealand Geological Survey Bulletin , 46 : 1 – 156 .
- Lindberg , DR . 1998 . “ Order Patellogastropoda ” . In Mollusca: the southern synthesis. Fauna of Australia , Edited by: Beesley , PL , Ross , GHB and Wells , A . Vol. 5 , 639 – 652 . Melbourne : CSIRO Publishing. Part B .
- Linné C von 1758 . Systema naturae per regna tria naturae … editio decima, reformata. Tomus 1 . L. Salvii , Holmiae . 823 p. [facsimile reprint, British Museum (Natural History), 1956; see Beu (2006:328) for use of Linné] .
- Lozouet , P . 1986 . Redéfinition des genres Potamides et Pirenella (Gastropoda, Prosobranchia) à partir des espèces actuelles et fossiles: implications phylétiques et biogéographiques . Annales de Paléontologie , 72 : 163 – 210 .
- Ludbrook , NH . 1941 . Gastropoda from the Abattoirs Bore, Adelaide, South Australia, together with a list of some miscellaneous fossils from the bore . Transactions of the Royal Society of South Australia , 65 : 79 – 102 .
- Ludbrook , NH . 1957 . The molluscan fauna of the Pliocene strata underlying the Adelaide Plains. Part 4: Gastropoda (Turritellidae to Struthiolariidae) . Transactions of the Royal Society of South Australia , 80 : 17 – 58 .
- Ludbrook , NH . 1978 . Quaternary molluscs of the western part of the Eucla Basin . Geological Survey of Western Australia Bulletin , 125 : 1 – 286 .
- Ludbrook , NH . 1984 . Quaternary molluscs of South Australia. Department of Mines and Energy , 327 South Australia : Handbook no. 9 .
- Ludbrook , NH and Gowlett-Holmes , KL . 1989 . “ Mollusca. Chapter II. Chitons, gastropods and bivalves ” . In Marine invertebrates of southern Australia Part II. Handbook of the fauna and flora of South Australia , Edited by: Shepherd , BA and Thomas , IM . 504 – 724 . Adelaide : South Australian Government Printer .
- McLean , JH . 1995 . Three additional new genera and two replacement names for northeastern Pacific prosobranch gastropods . The Nautilus , 108 : 80 – 82 .
- McLean , JH . 2009 . Genera of the Crepidula-group: revised definitions based on the characters of the shell muscle and septum (abstract) . Western Society of Malacologists Annual Report , 39 : 24
- Macpherson JH 1958 . An illustrated index of Tasmanian shells, with 50 plates and 1086 species , by W. L. May and revised by J. Hope Macpherson . Tasmanian Government Printer , Hobart . 54+[18] p.
- Macpherson , JH . 1966 . Port Phillip Survey 1957–1963: Mollusca . Memoirs of the National Museum of Victoria , 27 : 201 – 384 .
- Macpherson , JH and Chapple , EH . 1951 . A systematic list of the marine and estuarine Mollusca of Victoria . Memoirs of the National Museum of Victoria , 17 : 107 – 185 .
- Macpherson JH Gabriel CJ 1962 . Marine molluscs of Victoria . Melbourne , Melbourne University Press & National Museum of Victoria . xv + 475 p.
- Maplestone CM 1872 . Notes on Victorian Mollusca and their palates . The Monthly Microscopical Journal August 1 , 1872 : 45 54 .
- Marples , BJ . 1960 . A fossil penguin from the late Tertiary of North Canterbury . Records of the Canterbury Museum , 7 : 185 – 195 .
- Marshall , BA . 1991 . Dates of publication and supraspecific taxa of Bellardi and Sacco's (1873–1904) “I molluschi dei terreni terziarii del Piemonte e della Liguria” and Sacco's (1890) “Catalogo paleontologico del bacino terziario del Piemonte” . The Nautilus , 105 : 104 – 115 .
- Marshall , BA . 1995a . A revision of the Recent Calliostoma species of New Zealand (Mollusca: Gastropoda: Trochoidea) . The Nautilus , 108 : 83 – 127 .
- Marshall , BA . 1995b . Molluscan and brachiopod taxa introduced by F. W. Hutton in The New Zealand Journal of Science . Journal of the Royal Society of New Zealand , 25 : 495 – 500 .
- Marshall , BA . 1996 . Molluscan name-bearing types in the Museum of New Zealand Te Papa Tongarewa . Tuhinga: Records of the Museum of New Zealand Te Papa Tongarewa , 9 : 1 – 85 .
- Marshall , BA . 1998 . The New Zealand Recent species of Cantharidus Montfort, 1810 and Micrelenchus Finlay, 1926 (Mollusca: Gastropoda: Trochidae) . Molluscan Research , 19 : 107 – 156 .
- Marshall , BA . 2003 . A review of the Recent and late Cenozoic Calyptraeidae of New Zealand (Mollusca: Gastropoda) . The Veliger , 46 : 117 – 144 .
- Marshall , BA and Burch , KW . 2000 . The New Zealand Recent species of Muricopsis Bucquoy, Dautzenberg and Dollfus, 1882 (Gastropoda: Muricidae) . The Nautilus , 114 : 18 – 29 .
- Marshall , P and Murdoch , R . 1919 . Some new fossil species of Mollusca . Transactions of the New Zealand Institute , 51 : 253 – 258 .
- Marshall , P and Murdoch , R . 1920 . Some Tertiary Mollusca, with descriptions of new species . Transactions of the New Zealand Institute , 52 : 128 – 136 .
- Marshall , P and Murdoch , R . 1923 . Some Tertiary Mollusca, with descriptions of new species . Transactions of the New Zealand Institute , 54 : 121 – 128 .
- Martens , E von . 1872 . Conchylien von Cook's Reisen . Malacozoologische Blätter , 19 : 1 – 48 .
- Martens E von 1873 . Critical list of the Mollusca of New Zealand contained in European collections, with references to descriptions and synonyms . Wellington , Colonial Museum and Geological Survey Department . iv + v + 51 + viii p.
- Martyn T 1784–1792 . The universal conchologist, exhibiting the figure of every known shell … drawn … and painted … with a new systematic arrangement . T Martyn , London . Vols. 1 & 2, pl. 1–80, 1784; vol. 3, pls. 81–120, 1786; vol. 4, pls. 121–160, 1878; + 2 pl. of medals, etc., 1792 [Woodward 1910, p. 1258; names not available, but some New Zealand names conserved in ICZN Opinion 479 1957] .
- Marwick , J . 1924a . The Struthiolariidae . Transactions of the New Zealand Institute , 55 : 161 – 190 .
- Marwick , J . 1924b . Palaeontological notes on some Pliocene Mollusca from Hawke's Bay . Transactions of the New Zealand Institute , 55 : 191 – 201 .
- Marwick , J . 1924c . The Tertiary and Recent Naticidae and Naricidae of New Zealand . Transactions of the New Zealand Institute , 55 : 545 – 579 .
- Marwick , J . 1926 . New Tertiary Mollusca from North Taranaki . Transactions of the New Zealand Institute , 56 : 317 – 331 .
- Marwick , J . 1928 . The Tertiary Mollusca of the Chatham Islands including a generic revision of the New Zealand Pectinidae . Transactions of the New Zealand Institute , 58 : 432 – 506 .
- Marwick , J . 1931 . The Tertiary Mollusca of the Gisborne district . New Zealand Geological Survey Paleontological Bulletin , 13 : 1 – 177 .
- Marwick , J . 1948 . Lower Pliocene Mollusca from Otahuhu, Auckland . New Zealand Geological Survey Palaeontological Bulletin , 16 : 1 – 38 .
- Marwick , J . 1957a . Generic revision of the Turritellidae . Proceedings of the Malacological Society of London , 32 : 144 – 166 .
- Marwick , J . 1957b . New Zealand genera of Turritellidae, and the species of Stiracolpus . New Zealand Geological Survey Paleontological Bulletin , 27 : 1 – 55 .
- Marwick , J . 1965 . Upper Cenozoic Mollusca of Wairoa district, Hawke's Bay . New Zealand Geological Survey Paleontological Bulletin , 39 : 1 – 83 .
- Marwick , J . 1971 . New Zealand Turritellidae related to Zeacolpus Finlay (Gastropoda) . New Zealand Geological Survey Paleontological Bulletin , 44 : 1 – 87 .
- Maxwell PA 2009 . Cenozoic Mollusca In: Gordon DP . The New Zealand inventory of biodiversity: a Species 2000 symposium review , vol. 1. Canterbury University Press , Christchurch . Pp. 180 190 231–253.
- May , WL . 1921 . A check-list of the Mollusca of Tasmania , 114 Hobart : Tasmanian Government Printer .
- May , WL . 1923 . An illustrated index of Tasmanian shells. With 47 plates and 1052 species , 100 Hobart : Tasmanian Government Printer .
- Menke KT 1843 . Molluscorum Novae Hollandiae specimen, quod ad celebrandum diem jubilaeum vicesimum sextum mensis Februarii anni MDCCCXLIII, quo per vingti qinque annos prefessoris muneri in illustre gymnasio academico Hamburgensi praefuit vir doctissimus, clarissimus Joannes Georgius Christianus Lehmann … Hannover, in libraria aulica Hahniana . 46 p.
- Menke , KT . 1844 . Molluscorum Novae Hollandiae specimen … [review of above] . Zeitschrift für Malakozoologie , 1844 : 52 – 64 .
- Merriam , CW . 1941 . Fossil turritellas from the Pacific Coast region of North America. University of California . Publications of the Department of Geological Sciences , 26 : 1 – 214 .
- Meyer , CP , Geller , JB and Paulay , G. 2005 . Fine scale endemism on coral reefs: archipelagic differentiation in turbinid gastropods . Evolution , 59 : 113 – 125 .
- Mildenhall , DC and Pocknall , DT . 1989 . Miocene–Pleistocene spores and pollen from Central Otago, South Island, New Zealand . New Zealand Geological Survey Paleontological Bulletin , 59 : 1 – 128 .
- Monger , A . 1984 . “ Gastropoda, Prosobranchia ” . In Coastal Invertebrates of Victoria. An atlas of selected species , Edited by: Phillips , DAB , Handreck , CP , Bock , PE , Burn , R , Smith , BJ and Staples , DA . 23 – 66 . Melbourne : Marine Research Group of Victoria in association with the Museum of Victoria .
- Mörch OAL 1853 . Catalogus conchyliorum quae reliquit D. Alphonso d'Aguirra & Gadea, Comes de Yoldi, regis Daniae cubiculariorum princeps, ordis Dannebrogici in prima classe & ordinis caroli tertii eques. Fasciculus primus. Cephalophora. Haec conchylia publica auctione calendris octobribus dividentur . L. Kleini , Hafniae . 160 p.
- Mörch , OAL . 1860 . tudes sur la famille des Vermets (suite) . Journal de Conchyliologie , 8 : 27 – 48 .
- Mörch , OAL . 1861 . Review of the Vermetidae (Part I) . Proceedings of the Zoological Society of London , 30 : 145 – 181 .
- Morgans HEG 1981 . Biostratigraphy of Mikonui-1 offshore well . New Zealand Geological Survey Report PAL 45: 1 22 .
- Morley , MS . 1996 . Zelippistes benhami (Suter, 1902): (Gastropoda: Capulidae) . Poirieria (Auckland Museum Conchology Section) , 19 : 22 – 23 .
- Morley , MS . 2004 . A photographic guide to seashells of New Zealand , 143 Auckland : New Holland Publishers .
- Morton , JE . 1950 . The Struthiolariidae: reproduction, life history and relationships . Transactions of the Royal Society of New Zealand , 78 : 451 – 463 .
- Morton , JE . 1951 . The structure and adaptations of the New Zealand Vermetidae. Part II. The genera Stephopoma and Pyxipoma . Transactions of the Royal Society of New Zealand , 79 : 20 – 42 .
- Morton , JE . 1955 . The evolution of vermetid gastropods . Pacific Science , 9 : 3 – 15 .
- Morton , JE . 1997a . The Strombacea revisited. Pt. I. Struthiolaria and Aporrhais. Poirieria . Auckland Museum Conchology Section , 20 : 1 – 13 .
- Morton , JE . 1997b . The Strombacea revisited. Pt. II. The Strombidae: classic and baroque. Poirieria . Auckland Museum Conchology Section , 21 : 1 – 17 .
- Morton , JE and Keen , AM . 1960 . A new species of Stephopoma (Siliquariidae: Mesogastropoda) from the eastern Atlantic Ocean . Proceedings of the Malacological Society of London , 34 : 27 – 35 .
- Morton , JE and Miller , MC . 1968 . The New Zealand sea shore , 638 London and Auckland : Collins .
- Moss EGB 1908 . [The] Beautiful shells of New Zealand. An illustrated work for amateur collectors of New Zealand marine shells with directions for collecting and cleaning them . Auckland , Collins Bros . 46 +x p. [‘The’ on cover only].
- Murdoch , R . 1900 . Description of some new species of Pliocene Mollusca from the Wanganui district, with notes on other described species . Transactions of the New Zealand Institute , 32 : 216 – 221 .
- Naish , TR , Field , BD , Zhu , H , Melhuish , A , Carter , RM , Abbott , ST , Edwards , S , Alloway , BV , Wilson , GS , Niessen , F , Baker , A , Browne , G and Maslen , G . 2005 . Integrated behind-outcrop borehole and seismic stratigraphic architecture of a cyclothemic, shallow marine depositional system, Wanganui Basin, New Zealand . Journal of the Royal Society of New Zealand , 35 : 91 – 122 .
- Neef G 1967 . The geology of Eketahuna (N.Z.M.S. 1 N153) . Unpublished PhD thesis, Victoria University of Wellington . 397 p.
- Neef , G . 1970 . Notes on the subgenus Pelicaria . New Zealand Journal of Geology and Geophysics , 13 : 436 – 476 .
- Odhner , NHJ . 1917 . Results of Dr E. Mjöbergs Swedish Scientific Expeditions to Australia 1910–1913. XVII. Mollusca . Kungliga Svenska Vetenskapsakademiens Handlingar , 52 ( 16 ) : 1 – 115 .
- Odhner , NHJ . 1924 . Papers from Dr. Th. Mortensen's Pacific expedition 1914–16. XIX. New Zealand Mollusca . Videnskabelige Meddelelser fra Dansk Naturhistorisk Forening i Kjøbenhaven , 77 : 1 – 90 .
- Oliver WRB 1926 . New Zealand species of Calliostoma . Proceedings of the Malacological Society of London 17 : 107 115 [20 December 1926] .
- Olsson , AA . 1964 . Neogene molluscs from northwestern Ecuador , 256 Ithaca : Paleontological Research Institution .
- Orbigny A d’ 1841a 5 [in 1834–1847]. Voyage dans l'Amérique méridionale (le Brésil, la République Orientales de l'Uruguay, la République Argentine, la Patagonie, la République do Chili, la République de Bolivia, la République de Perou), exécuté dans 1826, 1827, 1828, 1829, 1830, 1831, 1832, et 1833. Mollusques . Paris , P. Bertrand . part 3 , pp. i–xliii, 1 758 .
- Orpin , AR , Gammon , PR , Naish , TR and Carter , RM . 1998 . Modern and ancient Zygochlamys delicatula shellbeds in New Zealand, and their sequence stratigraphic implications . Sedimentary Geology , 122 : 267 – 284 .
- Ortmann AE 1902 . Palaeontology, vol. 1, part 2. Tertiary invertebrates Scott WB Reports of the Princeton University Expeditions to Patagonia, 1896–1899 , vol. 4. Princeton, NJ , J. Pierpont Morgan Publishing Foundation , Pp. 47 332 .
- Paetel , F . 1888 . Catalog der Conchylien-Sammlung von Fr. Paetel. Vierte Neubearbeitung. Mit Hinzufügung der bis jetzt publicirten recenten Arten, sowie der ermittelten Synonyma. Erste Abtheilung: Die Cephalopoden, Pteropoden und Meeres-Gastropoden , 639 Berlin : Gebrüder Paetel .
- Palmer , KVW . 1947 . In: Harris GD, Palmer KVW. The Mollusca of the Jackson Eocene of the Mississippi embayment (Sabine River to the Alabama River), second section, part II, Univalves and index . Bulletins of American Paleontology , 30 ( 117 ) : 209 – 466 .
- Paulucci M 1877 . Osservazioni critiche sopra le specie del genere Struthiolaria, Lamarck . Bullettino della Società Malacologica Italiana 2 : 223 232 [text and synonymies largely copied from Deshayes (1843, pp. 534–537)] .
- Penniket , JR and Moon , GJH . 1970 . New Zealand seashells in colour , 112 Wellington : A. H. & A. W. Reed .
- Perry G 1811 . Conchology, or the natural history of shells; containing a new arrangement of the genera and species, illustrated by coloured engravings executed from the natural specimens, and including the latest discoveries . London , W. Miller . 4 p.+unpaginated captions to 61 pls. [editions and taxonomic names: Petit 2003] .
- Petit , RE . 2003 . George Perry's molluscan taxa and notes on the editions of his Conchology of 1811 . Zootaxa , 377 : 1 – 72 .
- Petit , RE . 2006 . Notes on Sowerby's The genera of recent and fossil shells (1821–1834) . Archives of Natural History , 33 : 71 – 89 .
- Petit , RE . 2007 . Lovell Augustus Reeve (1814–1865): malacological author and publisher . Zootaxa , 1648 : 1 – 120 .
- Petit , RE . 2009b . George Brettingham Sowerby, I, II & III: their conchological publications and molluscan taxa . Zootaxa , 2189 : 1 – 218 .
- Petit de la Saussaye , S . 1851 . Notice sur le G. Trichotropis, Sow., et description de coquilles appartenant probablement à ce genre . Journal de Conchyliologie , 2 : 17 – 24 .
- Petterd , WF . 1879 . Critical remarks on the Rev. J. E. Tenison Woods’ “Census of Tasmanian shells” . Journal of Conchology , 2 : 353 – 354 .
- Petuch , EJ . 1979 . Twelve new Indo-Pacific gastropods. Nemouria . Occasional Papers of the Delaware Museum of Natural History , 23 : 1 – 21 .
- Philippi RA 1851 . Centuria quarta testaceorum novarum . Zeitschrift für Malacozoologie 8 : 29 96 123–126.
- Pilkington , MC . 1974 . The eggs and hatching stages of some New Zealand prosobranch molluscs . Journal of the Royal Society of New Zealand , 4 : 411 – 432 .
- Pilkington , MC . 1976 . Descriptions of veliger larvae of monotocardian gastropods occurring in Otago plankton hauls . Journal of Molluscan Studies , 42 : 337 – 360 .
- Pillans , BJ , Alloway , BV , Naish , T , Westgate , J , Abbott , S and Palmer , A . 2005 . Silicic tephras in Pleistocene shallow marine sediments of Wanganui Basin, New Zealand . Journal of the Royal Society of New Zealand , 35 : 43 – 90 .
- Pilsbry HA 1888–1889a . Manual of conchology, structural and systematic; with illustrations of the species . By G. W. Tryon, Jr. [continued by H. A. Pilsbry]. Vol. 10. Neritidae, Adeorbidae, Cyclostrematidae, Liotiidae, Phasianellidae, Turbinidae, Trochidae, Stomatiidae, Haliotidae, Pleurotomariidae . G. W. Tryon , Philadelphia . 323 p. [pp. 1–208, 1888; 209–323, 1889].
- Pilsbry HA 1889b-1890 . Manual of Conchology [as above]. Volume 11. Trochidae, Stomatiidae, Pleurotomariidae, Haliotidae . Academy of Natural Sciences of Philadelphia , Philadelphia . 519 p. [pp. 1–208, 1889; 209–519, 1890]., Conchological Section
- Ponder WF 1998 . Family Hipponicidae; Superfamily Capuloidea Beesley PL Ross GJB Wells A Mollusca: the southern synthesis . Fauna of Australia, 5 . Melbourne , CSIRO Publishing . PartB, Pp. 770 771 774–775.
- Poppe GT 2008a . Philippine marine mollusks, vol. I (Gastropoda – Part 1) . ConchBooks , Hachenhei, , Germany . 758 p.
- Poppe GT 2008b . Philippine marine mollusks, vol. II (Gastropoda – Part 2) . ConchBooks , Hachenheim, , Germany . 848 p.
- Powell , AWB . 1924 . Notes on New Zealand Mollusca . New Zealand Journal of Science and Technology , 6 : 282 – 286 .
- Powell , AWB . 1927 . Mollusca from twenty-three fathoms off Ahipara, N. Z . Transactions of the New Zealand Institute , 58 : 295 – 300 .
- Powell , AWB . 1928 . The Recent and Tertiary cassids of New Zealand and a study in hybridisation . Transactions of the New Zealand Institute , 59 : 629 – 642 .
- Powell , AWB . 1931 . Waitotaran faunules of the Wanganui System: and descriptions of new species of Mollusca from the New Zealand Pliocene . Records of the Auckland Institute and Museum , 1 : 85 – 112 .
- Powell , AWB . 1937 . The shellfish of New Zealand. An illustrated handbook , 100 Auckland : Unity Press .
- Powell , AWB . 1940 . The marine Mollusca of the Aupourian province, New Zealand . Transactions of the New Zealand Institute , 70 : 205 – 248 .
- Powell , AWB . 1946a . The shellfish of New Zealand (second, revised and enlarged edition). An illustrated handbook , 106 Christchurch : Whitcombe & Tombs .
- Powell , AWB . 1946b . New species of New Zealand Mollusca from the South Island, Stewart Island and the Chatham Islands . Records of the Auckland Institute and Museum , 3 : 137 – 144 .
- Powell , AWB . 1958a . Shells of New Zealand. An illustrated handbook [third edition] , 202 Christchurch : Whitcombe & Tombs .
- Powell , AWB . 1958b . New Zealand molluscan systematics, with descriptions of new species, part 3 . Records of the Auckland Institute and Museum , 5 : 87 – 91 .
- Powell AWB 1962 . Shells of New Zealand. An illustrated handbook [fourth edition] . Whitcombe & Tombs , Christchurch . 203 p. [reprinted 1967].
- Powell , AWB . 1964 . New Zealand molluscan systematics with descriptions of new species: Part 4 . Records of the Auckland Institute and Museum , 6 : 11 – 20 .
- Powell , AWB . 1976a . On the considerable influx of warm water molluscs that have invaded northern New Zealand waters within recent years . Records of the Auckland Institute and Museum , 13 : 141 – 166 .
- Powell AWB 1976b . Shells of New Zealand. An illustrated handbook [fifth edition] . Whitcombe & Tombs , Christchurch . 154 p.
- Powell , AWB . 1979 . New Zealand Mollusca. Marine, land and freshwater shells , 500 Auckland : Collins .
- Pritchard , GB and Gatliff , JH . 1899 . On some new species of Victorian Mollusca . Proceedings of the Royal Society of Victoria , 11 : 179 – 184 .
- Pritchard , GB and Gatliff , JH . 1900 . Catalogue of the marine shells of Victoria. Part III . Proceedings of the Royal Society of Victoria , 12 : 170 – 205 .
- Pritchard , GB and Gatliff , JH . 1902 . Catalogue of the marine shells of Victoria. Part V . Proceedings of the Royal Society of Victoria , 14 : 85 – 138 .
- Quoy JCR , Gaimard P 1832–1835 . Mollusques Voyage de découvertes de l'Astrolabe, exécuté par ordre du Roi pendant les années 1826, 1827, 1828 et 1829, sous le commandement de M. J. Dumont d'Urville. Paris, J. Tastu. Zoologie, Tomes 2 & 3. Vol. 2, pp. 1–686; vol. 3, Mollusca on pp. 1–644 [of 954] [vol. 2, pp. 1–320, 1832; 321–686, 1833; vol. 3, pp. 1–366, 1834; 367–954, 1835] .
- Reeve LA 1841–1842 1 . Conchologia systematica, or complete system of conchology: in which the lepades and conchiferous Mollusca are described and classified according to their natural organization and habits . Longman , London, Brown, Green and Longmans . 1841, vi + 195 p.; vol. 2, 1842, 337 p.
- Reeve LA 1844 . Monograph of the genus Ranella. Conchologia iconica; a complete repertory of species . Vol. 2: 8 pls. [July 1844] LA Reeve , London [see Petit 2007].
- Reeve LA 1859 . Monograph of the genus Crepidula. Conchologia iconica; a complete repertory of species . Vol. 11: 5 pls. [June 1859] . LA Reeve , London .
- Reid , DG . 1988 . The genera Bembicium and Risellopsis (Gastropoda: Littorinidae) in Australia and New Zealand . Records of the Australian Museum , 40 : 91 – 150 .
- Richardson ES 1997 . Further molluscan records from the Te Piki Member, Cape Runaway, with the description of two new species . Proceedings of the Taupaki Malacological Society 1 : 1 26 [published privately by ES Richardson, The Taupaki Printery, 188 Hunters Road, Taupaki, RD 2, Henderson, New Zealand] .
- Roissy F de 1805 . Histoire naturelle, générale et particulière, des mollusques, animaux sans vertèbres et a sang blanc . Ouvrage faisant suite aux oeuvres de Leclerc de Buffon, et partie du cours complet d'histoire naturelle rédigé par C. S. Sonnini, membre de plusieurs sociétés savantes. Continué par Felix de Roissy . Vol. 6. F. Dufart , Paris . 480 p.
- Röding PF 1798 . Museum Boltenianum, sive catalogus cimeliorum e tribus regnis naturae … pars secunda, continens conchylia sive univalvia, bivalvia et multivalvia . J. C. Trappii , Hamburg . 199 p. [facsimile reprint, London, Sherborn & Sykes, 1906].
- Saul LR 1983 . Turritella zonation across the Cretaceous–Tertiary boundary, California . University of California Publications in Geological Sciences 125 : i x 1–164.
- Saul , LR and Squires , RL . 2008 . Cretaceous trichotropoid gastropods from the Pacific slope of North America: possible pathways to calyptraeid morphology . The Nautilus , 122 : 115 – 142 .
- Schilder , FA . 1941 . Verwandtschaft und Verbreitung der Cypraeacea . Achiv für Molluskenkunde , 73 : 57 – 120 .
- Schumacher , HCF . 1817 . Essai d’ un nouveau système des habitations des vers testacés , 287 Copenhagen : Schultz .
- Scott RJ 1997 . Aspects of the biology of the introduced gastropod Maoricolpus roseus . Bachelor of Antarctic Science (Hons) thesis , Hobart , University of Tasmania . 115 p.
- Shane , PAR . 1994 . A widespread early Pleistocene tephra (Potaka tephra, 1 Ma) in New Zealand: character, distribution and implications . New Zealand Journal of Geology and Geophysics , 37 : 25 – 35 .
- Shane , PAR , Black , TM , Alloway , BV and Westgate , JA . 1996 . Early to Middle Pleistocene tephrochronology of North Island, New Zealand: implications for volcanism, tectonism and paleoenvironments . Geological Society of America Bulletin , 108 : 915 – 925 .
- Sherborn CD 1902 . Index animalium sive index nominum quae ab A.D. MDCCLVIII generibus et speciebus animalium imposita sunt. Sectio prima. A kalendis Ianuariis, MCCCLVIII usquae ad finem Decembris, MDCCC . Cambridge University Press , Cambridge . lix + 1195 p.
- Sherborn CD 1930 . Index animalium, sive index nominum quae ab A.D. MDCCLVIII generibus et speciebus animalium imposita sunt. Sectio secunda, 1801–1850. Part XXIII. Index S littera-serratus . British Museum (Natural History) , London . Pp. 5707 5910 .
- Sherborn CD 1931 . Index animalium, sive index nominum quae ab A.D. MDCCLVIII generibus et speciebus animalium imposita sunt. Sectio secunda, 1801–1850. Part XXVII. Index Trichoscelis-variegatus . British Museum (Natural History) , London . Pp. 6583 6806 .
- Sherborn , CD and Woodward , BB . 1901 . Notes on the dates of publication of parts of Kiener's “Spécies general et iconographie des coquilles vivantes”, etc. (1834–80) . Proceedings of the Malacological Society of London , 4 : 216 – 219 .
- Simone LRL 2002 . Comparative morphological study and phylogeny of representatives of the Superfamily Calyptreaoidea (including Hipponicoidea) (Mollusca, Caenogastropoda) . Biota Neotropica 2(2) : 1 137 . On line at: http://www.biotaneotropica.org.br/v2n2/pt/abstract?article+BN01602022002 .
- Simone , LRL . 2006 . Morphological and phylogenetic study of the Western Atlantic Crepidula plana complex (Caenogastropoda, Calyptraeidae), with description of three new species from Brazil . Zootaxa , 1112 : 1 – 64 .
- Simpson , GG . 1972 . Pliocene penguins from North Canterbury, New Zealand . Records of the Canterbury Museum , 9 : 150 – 182 .
- Smith EA 1874 . Mollusca Richardson J , Gray JE The zoology of the voyage of H.M.S. Erebus & Terror, under the command of Captain Sir James Clark Ross, R.N., F.R.S., during the years 1839 to 1843. By authority of the Lords Commissioners of the Admiralty . E. W. Janson , London . Vol. 2, part 21, pp. 1 7 , pls. 1–4.
- Smith , EA . 1915 . Mollusca. Part I. – Gastropoda Prosobranchia, Scaphopoda, and Pelecypoda. British Antarctic (“Terra Nova”) Expedition, 1910, Natural History Reports . Zoology , 2 : 61 – 112 .
- Sowerby GB I 1821–1834 . The genera of Recent and fossil shells, for the use of students in conchology and geology . GB Sowerby , London . 265 pls. with unnumbered text, issued in 42 parts [index and plate numbers issued by B. Quaritch, London, pp. i–viii; Struthiolaria inermis in part 1, pl. ‘222’, 3, 4, 31 December 1821; Crepidula costata in part 23, pl. ‘152’, figs. 3, 3 June 1824; authorship and dates: Petit 2006, 2009b]
- Sowerby GB II 1832–1841 . The conchological illustrations, or coloured illustrations of all the hitherto unfigured Recent shells . GB Sowerby , London . vi + 116 p., issued in 117 parts [1832–1833, parts 1–34, each with 1 pl; 1833–1841, ‘parts’ 35–200, issued as 83 parts, each with 2 plates; Beu & Maxwell 1990, p. 450; Petit 2009b; Ranella: pls. 84–85, 88–89, 1835; pls. 92–93, 1836] .
- Sowerby GB II 1842 . Monograph of the genus Struthiolaria . Thesaurus conchyliorum, or figures and descriptions of shells . Vol. 1(1), pp. 23 24 , pl. 5. GB Sowerby , London [see Petit 2009b for details of all works by GB Sowerby I–III].
- Sowerby GB II 1874 . Monograph of the genus Trichotropis. Conchologia Iconica; a complete repertory of species . Vol. 19, 2 pls. [August 1874] . LA Reeve , London .
- Sowerby GB II 1883 . Monograph of the Family Calyptraeidae. Thesaurus conchyliorum, or monographs of genera of shells . Vol. 5, parts 39–40, pp. 55 74 , pls. 445–453. GB Sowerby , London (attributed to GB Sowerby II by Petit 2009b:35).
- Sowerby GB III 1892 . Marine shells of South Africa. A catalogue of all the known species with references to figures in various works, and figures of such as are rare, little known, or hitherto unfigured . GB Sowerby , London . iv + 89 p.
- Speight , R . 1913 . List of Tertiary Mollusca in the Canterbury Museum . Records of the Canterbury Museum , 2 : 27 – 55 .
- Spencer HG , Willan RC 1996 . The marine fauna of New Zealand: index to the fauna 3. Mollusca . New Zealand Oceanographic Institute Memoir 105 : 1 125 [‘1995’, issued February 1996] .
- Spencer HG , Marshall BA , Willan RC 2009 . Recent Mollusca Gordon DP The New Zealand inventory of biodiversity: a Species 2000 symposium review . Vol. 1. Canterbury University Press , Christchurch . Pp. 162 168 , 196–219.
- Spengler L 1782 . Beschreibung zwoer seltenen neuen Gattungen südländischer Conchylien, mit erleuchteten Abbildungen . Der Naturforscher 17 : 24 31 pl. 2.
- Strathmann , MF and Strathmann , RR . 2007 . An extraordinarily long larval duration of 4.5 years from hatching to metamorphosis for teleplanic veligers of Fusitriton oregonensis . The Biological Bulletin , 213 : 152 – 159 .
- Suter , H . 1902a . Description of a new species of Separatista from New Zealand . The Journal of Malacology , 9 : 65 – 66 .
- Suter , H . 1902b . List of the species described in F. W. Hutton's Manual of the New Zealand Mollusca, with the corresponding names used at the present time . Transactions of the New Zealand Institute , 34 : 207 – 224 .
- Suter , H . 1906 . Notes on New Zealand Mollusca, with descriptions of new species and subspecies . Transactions of the New Zealand Institute , 38 : 316 – 333 .
- Suter , H . 1910 . List of Recent shells found fossil in New Zealand . Transactions of the New Zealand Institute , 42 : 8 – 13 .
- Suter H 1913 . Manual of the New Zealand Mollusca. With an atlas of quarto plates [1915] . Government Printer , Wellington . xxiii + 1120 p.
- Suter , H . 1914 . Revision of the Tertiary Mollusca of New Zealand, based on type material. Part I . New Zealand Geological Survey Palaeontological Bulletin , 2 : 1 – 64 .
- Suter , H . 1915 . Revision of the Tertiary Mollusca of New Zealand, based on type material. Part II . New Zealand Geological Survey Palaeontological Bulletin , 3 : 1 – 89 .
- Suter , H . 1917 . Descriptions of new Tertiary Mollusca occurring in New Zealand, accompanied by a few notes on necessary changes in nomenclature. Part I . New Zealand Geological Survey Palaeontological Bulletin , 5 : 1 – 93 .
- Sutherland , R , Nathan , S and Turnbull , IM . 1995 . Pliocene-Quaternary sedimentation and Alpine Fault-related tectonics in the lower Cascade Valley, South Westland, New Zealand. With an appendix by AG Beu: Macrofossils of the Teer Formation . New Zealand Journal of Geology and Geophysics , 38 : 431 – 450 .
- Swainson W 1840 . A treatise on malacology; or the natural classification of shells and shell fish . London , Longman, Brown, Green & Longmans . viii + 419 p.
- Tate , R . 1893 . The gastropods of the older Tertiary of Australia. Part IV (including supplement to Part III) . Transactions of the Royal Society of South Australia , 17 : 316 – 345 .
- Tate , R and May , WL . 1901 . A revised census of the marine Mollusca of Tasmania . Proceedings of the Linnean Society of New South Wales , 26 : 344 – 471 .
- Tenison Woods , JE . 1877 . On some new Tasmanian marine shells, 2nd series . Papers and Proceedings of the Royal Society of Tasmania for , 1876 : 131 – 159 .
- Tenison Woods JE 1878 . Census; with brief descriptions of the marine shells of Tasmania and the adjacent islands . Papers and Proceedings of the Royal Society of Tasmania for 1877: 26–57 [volume dated 14 June 1879, but reprint sent to R Tate by Tenison Woods hand-dated 30 April 1878; I Loch, AMS, JE Tenison Woods ms bibliography] .
- Tenison Woods , JE . 1880 . On some Tasmanian Trochidae . Papers and Proceedings of the Royal Society of Tasmania for , 1879 : 59 – 70 .
- Terry JS [later JT Smith] 1968 . Mediargo, a new Tertiary genus in the Family Cymatiidae . The Veliger 11 : 42 44 .
- Thiele J 1929–1935 . Handbuch der systematischen Weichtierkunde. Jena, G Fischer. Erster Band. Teil 1, 1929. Loricata; Gastropoda: Prosobranchia. 376 p. Teil 2, 1931 . Gastropoda : Opisthobranchia, Pulmonata; addenda . Pp. 377 778 . Zweiter Band. Teil 3, 1934. Scaphopoda, Bivalvia, Cephalopoda; addenda and corrigenda. Pp. 779–1022. Teil 4, 1935. General part and corrigenda. Pp. 1023–1154, i–vi [reprint, Amsterdam, A. Asher & Co., 1963; English translation by JS Bhatti, edited by R Bieler & PM Mikkelsen, Washington, Smithsonian Libraries & National Science Foundation, 1992] .
- Thiele J 1930 . Gastropoda und Bivalvia Michaelsen W , Hartmeyer R Die Fauna Südwest-australiens . Ergebnisse der Hamburger Südwest-Australischen Forschungsreise 1905, Band 5, Lieferung 8 : 561 596 . Jena, G. Fischer .
- Thomson , JA . 1917 . The Hawera Series, or so-called “drift formation” of Hawera . Transactions of the New Zealand Institute , 49 : 414 – 417 .
- Thomson , JA . 1926 . Geology of fossil localities near Waipukurau, Hawke's Bay . Transactions of the New Zealand Institute , 56 : 347 – 354 .
- Trew , A . 1992 . Henry and Arthur Adams's new molluscan names , 63 Cardiff : National Museum of Wales .
- Tryon GW 1880b–1881 . A manual of conchology [as above]. Series 1, vol. 3. Tritonidae, Fusidae, Buccinidae . G. W. Tryon , Philadelphia . 310 p. [pp. 1–64, 31 December 1880; 65–310, 1881].
- Tryon GW 1885 . A manual of conchology [as above]. Series 1, vol. 7. Terebridae, Cancellariidae, Strombidae, Cypraeidae, Ovulidae, Cassididae, Doliidae . G. W. Tryon , Philadelphia . 309 p.
- Tryon GW 1886 . A manual of conchology [as above]. Series 1, vol. 8. Naticidae, Calyptraeidae, Turritellidae, Vermetidae, Caecidae, Eulimidae, Turbonillidae, Pyramidellidae . G. W. Tryon , Philadelphia . 461 p.
- Tryon GW . 1887 . A manual of conchology [as above]. Series 1, vol. 9. Solariidae, Ianthinidae, Trichotropidae, Scalariidae, Cerithiidae, Rissoidae, Littorinidae . G. W. Tryon , Philadelphia . 488 p. [Solariidae, by WB Marshall, see above; pp. 3–32].
- Tucker , JK . 2004 . Catalog of Recent and fossil turrids (Mollusca: Gastropoda) . Zootaxa , 682 : 1 – 1295 .
- Turton WH 1932 . The marine shells of Port Alfred, S. Africa . Oxford, Oxford University Press , ; Humphrey Milord , London . xvi + 331 p.
- Valenciennes A 1858 . Note sur une suite intéressante de coquilles rapportées des mers du Japon, et de la Manche de Tartarie, par M. le Dr Barthe. Comptes Rendus Hebdomadaires des Séances de l'Académie des Sciences 44 ( 16 ): 759 763 [A summary appeared also in: L'Institut, Journal Universal des Sciences, et des Sociétés Savantes, en France et a l'Etranger, Ire Section: Sciences Mathémathiques, Physiques et Naturelles 26e Année-No. 1269: 143–144, 1858] .
- Van der Lingen G , Smale D , Field BD 1985 . Liquefaction deformation structures in sediment gravity flow deposits of the Pleistocene Castlepoint Formation, Castlepoint, Wairarapa, New Zealand . New Zealand Geological Survey Report SL8 : 4 21 .
- Vella , P . 1953 . The genus Pelicaria in the Tertiary of East Wairarapa . Transactions of the Royal Society of New Zealand , 81 : 35 – 48 .
- Verco , J . 1906a . Notes on South Australian marine Mollusca, with descriptions of new species. – Part III . Transactions and Proceedings of the Royal Society of South Australia , 30 : 143 – 150 .
- Verco , J . 1906b . Notes on South Australian marine Mollusca, with descriptions of new species. – Part IV . Transactions and Proceedings of the Royal Society of South Australia , 30 : 205 – 224 .
- Verco , J . 1908 . Catalogue of the marine Mollusca of South Australia , 24 Adelaide : J. Verco .
- Verco , J . 1912 . Notes on the marine Mollusca of Western Australia with descriptions of new species, part 2 . Transactions of the Royal Society of South Australia , 36 : 192 – 205 .
- Verco J 1935 . Combing the southern seas. (Edited from the diaries of Sir Joseph Verco by BC Cotton) . Adelaide , Rigby . 174 + 24 p.
- Warren G 1995 . Geology of the Parnassus area, scale 1: 50 000 . Institute of Geological and Nuclear Sciences Geological Map 18 : 1 sheet + 36 p.
- Warren , G and Speden , IG . 1977 . The Piripauan and Haumurian stratotypes (Mata Series, Upper Cretaceous) and correlative sequences in the Haumuri Bluff district, South Marlborough . New Zealand Geological Survey Bulletin , 92 : 1 – 60 .
- Waters , JM . 2008 . Driven by the west-wind drift? A synthesis of southern temperate marine biogeography, with new directions for dispersalism . Journal of Biogeography , 35 : 417 – 427 .
- Watson , RB . 1886 . Report on the Scaphopoda and Gasteropoda collected by H.M.S. Challenger during the years 1873–1876. Report of the Scientific Results of the Voyage of H.M.S. Challenger During the Years 1973–76 . Zoology , 15 : 1 – 756 .
- Webster , WH . 1906 . Additions to the New Zealand fauna . Transactions of the New Zealand Institute , 38 : 309 – 312 .
- Wells , PE . 1986 . An estuarine fossil assemblage from the Mangaoranga Formation (Upper Miocene), Wairarapa, New Zealand . Journal of the Royal Society of New Zealand , 16 : 307 – 317 .
- Welter-Schultes , FW . 1999 . Systematisches Conchylien-Cabinet von Martini und Chemnitz (1837–1920), bibliography of the volumes in Göttingen . Archives of Natural History , 26 : 157 – 203 .
- Wenz W 1938–1944 . Gastropoda. Teil 1: Allgemeiner Teil und Prosobranchia Schindewolf OH Handbuch der Paläozoologie, Band 6. Berlin, Gebrüder Borntraeger . Vol. 1, pp. i–xii, 1–948; vol. 2, pp. 949–1639 [dates (contents, p. xii): pp. 1–480, 1938; pp. 481–720, 1939; pp. 721–960, 1940; pp. 961–1200, 1941; pp. 1201–1506, 1943; pp. 1507–1639, i–xii, 1944; reprint, Berlin, Borntraeger, 1960–1961] .
- Whitelegge , T . 1889 . List of the marine and fresh-water invertebrate fauna of Port Jackson and the neighbourhood . Journal and Proceedings of the Royal Society of New South Wales , 23 : 163 – 323 .
- Wilckens , O . 1922 . The Upper Cretaceous gastropods of New Zealand . New Zealand Geological Survey Palaeontological Bulletin : i–v , 9 : 1 – 42 .
- Williams , ST . 2007 . Origins and diversification of Indo-West Pacific marine fauna: evolutionary history and biogeography of turban shells (Gastropoda, Turbinidae) . Biological Journal of the Linnean Society , 92 : 573 – 592 .
- Williams , ST , Donald , KM , Spencer , HG and Nakano , T . 2010 . Molecular systematics of the marine gastropod families Trochidae and Calliostomatidae (Mollusca: Superfamily Trochoidea) . Molecular Phylogenetics and Evolution , 54 : 783 – 809 .
- Williams ST , Karube S , Ozawa T 2008 . Molecular systematics of Vetigastropoda: Trochidae, Turbinidae and Trochoidea redefined . Zoologica Scripta 2008 : 24 pp; doi: doi: 10:1111/j.1463_6409.2008.00341.x
- Williams , ST and Ozawa , T . 2006 . Molecular phylogeny suggests polyphyly of both turban shells (family Turbinidae) and the superfamily Trochoidea (Mollusca: Vetigastropoda) . Molecular Phylogenetics and Evolution , 39 : 33 – 51 .
- Wilson , B . 1993 . Australian marine shells, 1. Prosobranch gastropods, part one , 408 Kallaroo, Perth : Odyssey Publishing .
- Wood W 1828 . A supplement to the Index Testaceologicus, or a catalogue of shells, British and foreign . London , W. Wood . iv + 59 p.
- Woodring W 1928 . Miocene mollusks from Bowden, Jamaica. Part II. Gastropods and discussion of results . Carnegie Institution of Washington Publication 385 : i–vii, 1 564 .
- Woodward BB 1903–1915 . Catalogue of the books, manuscripts, maps and drawings in the British Museum (Natural History), London . British Museum (Natural History) , London . Vol. 1, 1903, pp. i–viii, 1–500; vol. 2, 1904, pp. 501–1038; vol. 3, 1910, pp. 1039–1494; vol. 4, 1913, pp. 1495–1956; vol. 5, 1915 , pp. 1957 2403 .
- Worthy , TH and Grant-Mackie , JA . 2003 . Late Pleistocene avifaunas from Cape Wanbrow, Otago, South Island, New Zealand . Journal of the Royal Society of New Zealand , 33 : 427 – 485 .
- Zittel , KA . 1864 . Fossile Mollusken und Echinodermen aus Neu-Seeland. In: Hochstetter F von, Hörnes M, Hauer F Ritter von ed. Paläontologie von Neu-Seeland. Beiträge zur Kenntniss der fossilen Flora und Fauna der Provinzen Auckland und Nelson. Novara-Expedition . Geologischer Theil , 1 ( 2 ) : 15 – 68 .