Abstract
The avifauna from the Glencrieff swamp deposit in North Canterbury, South Island, New Zealand, is described. Radiocarbon ages of moa bones bracket miring at the site to between 10,000 and 12,000 (uncalibrated) years BP. Heavy-footed moa (Pachyornis elephantopus) and eastern moa (Emeus crassus) dominated the moa assemblage at the site, while South Island giant moa (Dinornis robustus) and stout-legged moa (Euryapteryx curtus, formerly E. gravis (in part)) were rare. The total assemblage from the site consists of at least 1896 bones from 18 species of birds, of which nine are extinct and a further three locally extinct. In addition, we report on the discovery of the oldest known moa gizzard contents, the palynology of the Glencrieff deposit and comment on significant recent changes in site preservation conditions that are threatening the continued preservation of this significant fossil deposit.
Introduction
The Glencrieff swamp offers an important insight into the avifauna in Late Glacial North Canterbury (). Several excavations of bones from the site have occurred over the past 40 years (reviewed by Worthy & Holdaway Citation1996). In 1971, a partial skeleton of the South Island giant moa (Dinornis robustus Owen, 1846, CM Av25373) was excavated by Canterbury Museum staff and the Eaves family, the former owners of Glencrieff. The location within Glencrieff where the D. robustus skeleton was excavated from is unknown but is probably from an area close to the swamp outlet away from the main sub-fossil deposits. Further excavations in 1991 by the Eaves family resulted in 356 moa bones being found, which were retained as a private collection that was subsequently lost overseas (THW, unpublished data). In 1993–94, Worthy and Holdaway (Citation1996) conducted three further excavations, mapping the site and the extent of the bone deposits. In order to obtain new palaeontological and taphonomic data for analysing the deposition mechanism at the Glencrieff deposit, an additional sub-deposit, previously identified in 1993–94 was excavated in 2007 (Wood et al. Citation2008b). This paper describes the results of the 2007 excavation and summarizes the avifauna of Glencrieff. Additionally, this paper documents the oldest known moa gizzard contents and the palynology of the Glencrieff deposit, and comments upon recent changes in site preservation conditions since the 1993–94 excavations.
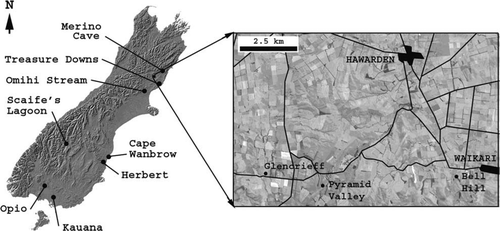
Figure 1 Location of fossil deposits in the South Island discussed in this paper, and location of the Late Glacial Glencrieff miring bone deposit in North Canterbury, South Island, New Zealand (insert).
Site discription
Glencrieff is located on the Wilson's (previously Eaves's) farm near Hawarden, North Canterbury (42°58′07.45′′S, 172°34′01.84′′E, 288 m asl), South Island, New Zealand (). Worthy and Holdaway (Citation1996) and Wood et al. (Citation2008b) described the stratigraphy, taphonomy and extent of the bone deposit.
Methods
Excavation
The bone deposit identified by Worthy and Holdaway (Citation1996) to the south of their 1993–94 excavation was re-located by probing the ground with a metal rod. This deposit was excavated in a pit measuring 5 m×2 m, with an extension of 0.5 m×2 m along the pit's northern face, adjoining the 1993–94 excavation. The pit is equivalent to squares I0–I3, J0–J3 and half of G1 and G2. The exact positions of all crania, sterna, pelves, femora, tibiotarsi, fibulae, tarsometatarsi and preserved gizzard contents were measured, with respect to the x and y axes of the excavation pit, along with their depth, orientation and inclination. These measurements were used by Wood et al. (Citation2008b) to create a three-dimensional reconstruction of the deposit. Excavated remains were identified and accessioned into the Canterbury Museum collections. Gizzard contents are stored frozen at Landcare Research, Lincoln.
Identification
Bones were identified to species level through comparison with reference material held in Canterbury Museum and by application of the identification key published by Worthy (1988a, b; see also Worthy & Holdaway Citation2002; appendix 2). Where possible, preserved gizzard contents were associated with moa species by articulated moa ribs and sterna.
Nomenclature
We follow the taxonomy advocated in the New Zealand Bird Checklist (Gill et al. Citation2010) for extinct and extant New Zealand birds. We use the following geological interval names: Holocene, the present interglacial period, beginning at 10,000 years BP; Late Glacial, a period of rapid climatic and vegetational change, from 14,000 to 10,000 years BP (Worthy & Roscoe Citation2003); and the Otiran Glaciation, the last New Zealand glacial period, from approximately 70,000 to 14,000 years BP, encompassing the Last Glacial Maximum, 29,000–19,000 years BP (Newnham et al. Citation2007).
Collagen preparation and radiocarbon dating
To augment the three published radiocarbon dates available for moa bones from Glencrieff (Worthy & Holdaway Citation1996), five bones of heavy-footed moa (Pachyornis elephantopus (Owen, 1856)) originating from the excavations in 1993–94 were radiocarbon-dated at the University of Wollongong (UW) and the Australian National University (ANU). The stratigraphic position of radiocarbon dated bones within the Glencrieff deposit is shown in and .
Each radiocarbon date was assigned a specific ANU laboratory number. For dates with laboratory numbers ANU1605–1610, collagen preparation was conducted at UW using ultrafiltration, following Bronk Ramsey et al. (Citation2004a, Citationb) and Jacobi et al. (Citation2006), while stepped-combustion and graphitization followed Turney et al. (Citation2008). Graphitized samples were run on the ANU single-stage accelerator mass spectrometer (SSAMS) with the appropriate carbon blanks and known age samples. The following modifications were made to the methodology of Bronk Ramsey et al. (Citation2004a, Citationb) and Jacobi et al. (Citation2006) for dates with laboratory numbers ANU4079–4937 and ANU7612–7625 (combustion, graphitization and radiocarbon dating were all conducted at ANU): The 8 µm Eezi filter centrifugal dialysis step was not included as this is not necessary to retrieve pure collagen (Higham et al. Citation2004); and silver powder was used instead of silver wire in the combustion step, providing a greater surface area to remove contaminants. As noted in the discussion, it was subsequently discovered that the iron catalyst used in the graphitization reaction for dates ANU4079–4937 contributed a limited amount of modern 14C to the sample (percent modern carbon (pMC) 0.24–0.38±0.03–0.04), despite standard cleaning procedures (reduction by hydrogen at 450 °C; Boaretto et al. Citation2009; see results and discussion for further comment). Subsequently, for dates ANU7612–7625, a new modern 14C-free iron catalyst was used (pMC weighted average 0.080±0.007). Combustion, graphitization and radiocarbon dating were conducted using ultra-purified collagen prepared for the dates ANU4079–4937.
Dates are reported as radiocarbon ages, based on Libby T1/2=5568 years, uncorrected for calendar variation, in years before present (i.e. before 1950). Radiocarbon ages were calibrated using the programme OxCal4.0 (available at www.c14.arch.ox.ac.uk) and the IntCal04 (Reimer et al. Citation2004) or the ShCal04 calibration curve (McCormac et al. Citation2004). Calibrated dates are reported as 95% confidence calibrated ages (± 2 SD) in years BP. To independently ascertain the quality of radiocarbon dates, sub-samples of ultra-purified collagen were used to determine the %N,%C and the C:N ratio at UW.
Gizzard contents analysis
Moa gizzard contents were analysed following the methodology of Wood (Citation2007). Sub-samples from eight of the 16 preserved gizzard contents excavated from Glencrieff were wet-sorted using a dissecting microscope at 10x magnification. Identifiable plant macro remains were counted (Appendix 1) and representative examples of seeds, leaves and twigs were stored in 70% ethanol. Seeds were identified using Webb and Simpson (Citation2001). The taxonomic identity of wood was determined following the methodology of Meylan and Butterfield (Citation1978). Wood samples were soaked in water and transverse sections were cut with razor blades. All measurements were made to the nearest 0.1 mm using vernier callipers.
Palynology
Samples for pollen analysis from the Moar (Citation2008) Glencrieff sediment core and preserved moa gizzard contents (from two preserved gizzard contents associated with one P. elephantopus, X09/2/21, and one Emeus crassus(Owen, 1846), X09/2/124) were prepared following the methodology of Moore et al. (Citation1991); hot 10% KOH, 40% HF, bleaching and acetolysis. The sediment core was collected towards the edge of the Glencrieff deposit in shallower strata (see Worthy & Holdaway Citation1996; Fig. 6 for location) with the base of the peat in the core at 80 cm depth. Counts of at least 200 pollen grains were made for each sample. Spores of the coprophilous fungus genus Sporormiella Ellis & Everhart 1892, used as an indicator of vertebrate herbivore biomass (Davis & Shafer Citation2006), were also counted to provide a proxy for large herbivores at the site following the methodology of Wood et al. (Citation2011). Pollen data and the relative abundance of Sporormiella spores are expressed as a percentage of the total pollen sum. Microscopic charcoal fragments were quantified using the point count method of Clark (Citation1982). The results are expressed as a charcoal index (hits per 11 points per field of view, calculated as a percentage of the total pollen sum) to avoid spurious fluctuations resulting from changing accumulation rates.
Abbreviations
New Zealand museums: NMNZ, Museum of New Zealand Te Papa Tongarewa, Wellington (ex National Museum of New Zealand); CM, Canterbury Museum, Christchurch. Minimum numbers of individuals (MNI) were determined for each taxon from the most frequent skeletal element (maximum of left or right side only). In species lists, x/y=number of bones/MNI represented in each deposit. When listing material, bones are sometimes identified as left (L), right (R) or partial (pt) elements.
Results
Avifauna
Approximately 2400 individual bones have been collected from excavations that have been undertaken at Glencrieff since 1971. Of these, 1896 bones have been identified as belonging to 18 species of birds. The unidentified bones include vertebrae, ribs, pubes, ischia and phalanges, which cannot be identified accurately to species but are unlikely to affect the MNI.
Moa fauna
At least 16 individual moa from four species were represented in the 2007 excavation (). These include: P. elephantopus (MNI = 8, including one sub-adult); eastern moa, E. crassus (MNI = 6); D. robustus (MNI = 1); and stout-legged moa, Euryapteryx curtus (Owen, 1846) (a monotypic genus in which, formerly, bones from the South Island and southern North Island were distinguished from E. curtus as E. gravis (Owen, 1870); see Bunce et al. Citation2009; MNI = 1). The E. curtus individual represents the first of this species discovered at Glencrieff. Several of the moa pelves had puncture marks; these are attributed to either predation by Haast's eagle (Aquila moorei (Haast, 1872)), which are hypothesized to penetrate the pelvis with their claws, or to kea (Nestor notabilis Gould, 1856) beak marks during scavenging. At least 51 individual moa are represented in the total fauna from Glencrieff. Specimens from the 2007, Worthy and Holdaway (Citation1996), Eaves, and Canterbury Museum excavations are summarized in and Appendix 2.
Table 1 A summary of moa specimens excavated from Glencrieff in all known excavations.
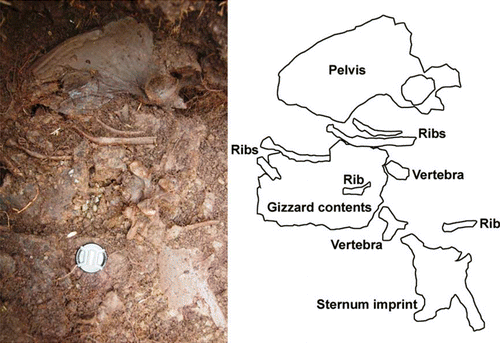
As was observed for the earlier excavation by Worthy and Holdway (1996), most of the bones excavated in 2007 were disarticulated. This is in contrast to the discrete D. robustus skeleton (CM Av25273) that was excavated by Canterbury Museum staff in 1971, from an unknown location within the Glencrieff swamp. This specimen is presumably not from the Eaves, Worthy and Holdaway (Citation1996) or 2007 deposits because disarticulated bones rather than discrete skeletons were excavated from these deposits. Test excavations of the hypothesized location of CM Av25273 near the swamp outlet in 1993–94 revealed a peat matrix that was soft enough to trap individuals. CM Av25273 may represent a significantly younger miring event than the main deposits within Glencrieff. However, we found several incidences of associated and articulated remains during the 2007 excavation. Examples included the pelvis + LR femora + LR tibiotarsi + LR fibulae + LR tarsometatarsi of a P. elephantopus and the vertebrae + pelvis + ribs + sternum +gizzard contents of an E. crassus (). Despite the prevalence of disarticulation, we could assign most leg bones (femora, tibiotarsi and tarsometatarsi) to individual left and right pairs on the basis of comparing within a taxon, the size and shape of muscle and ligamental scars.
Figure 2 Articulated pelvis, ribs, and associated vertebrae, sternum and gizzard contents of an eastern moa (Emeus crassus (Owen, 1846)), Glencreiff, 2007. Lens cap diameter is 58 mm. Note the weathered appearance of the pelvis from this relatively shallow part of the deposit and abundant roots of poplars (Populus Linneaus 1753).
The majority of the known bone deposits along the southern margin of Glencrieff have now been excavated, with probing by a metal rod revealing no fossil bones to the south of the 2007 excavation. The area to the north of the excavations conducted in 1993–94 was not probed in 2007, as the only bones known to remain in situ have previously been reported between squares B2-B3-H and the Eaves deposit (Worthy & Holdaway Citation1996). However, little exploration of the northern half of the deposit was undertaken in 1993–94 (THW, pers. observ.)
Non-moa avifauna
Eight non-moa bird species were discovered during the 2007 excavation () and have been summarized with previous finds of non-moa bird species in Appendix 2. Notably, Haast's eagle, recorded by Worthy and Holdaway (Citation1996), was missing from the 2007 excavation. Fourteen species of non-moa birds are represented in the total fauna from Glencrieff (see Appendix 2).
Table 2 List of non-moa bird specimens excavated from Glencrieff, 2007.
Radiocarbon ages
Twelve radiocarbon ages were obtained from five P. elephantopus bones (). Radiocarbon ages from bones derived from the 1993–94 excavations were between 11,390±130 yr BP (ANU1610) and 9070±80 yr BP (ANU4937), and are within the range of the three previously published dates (11,898±82 yr BP (NZA4162) to 10,470±130 yr BP (NZA4079); Worthy & Holdaway Citation1996), except ANU4937 at 9070±80 yr BP. Specimens processed using the UF1 method (ultrafiltration, stepped combustion) were radiocarbon dated to between 11,490±80 yr BP (ANU1606) and 10,580±90 yr BP (ANU1605; ). In contrast, the same specimens, using the UF2 method (ultrafiltration, using the contaminated iron catalyst), were significantly younger in two of the three specimens dated (by 190 yr for NMNZ S.32670.9 and by 160 yr for NMNZ S.32670.3), but 340 yr older in the other specimen (NMNZ S.32670.2). Finally, samples processed using the UF3 method (ultrafiltration, silver powder and uncontaminated iron catalyst) returned dates that were younger (by 80 yr for NMNZ S.32670.9, and by 60 yr for NMNZ S.32670.3), 1460 yr older (NMNZ S.32670.8), and the same (NMNZ S.32670.2) as samples processed using the UF1 and UF2 methods. The biases in the radiocarbon ages between the different treatment methods are significant (greater than one sigma) but the differences between radiocarbon dates are not large. To determine whether the radiocarbon dates were stratigraphically concordant, the radiocarbon ages were plotted against the excavation depth of the dated specimens (). The results indicate there is no relationship between excavation depth of the sample and its radiocarbon age from the Worthy and Holdaway (Citation1996) excavation ().
Figure 3 Relationship of radiocarbon age (± 1 SD) to excavation depth for dated moa bones at Glencrieff. Numbers refer to museum accession numbers (), except NZA4162, which refers to the Rafter Radiocarbon Laboratory number because there is no accessioned material for this bone. Preparations under Prep Code are: ABA, acid base acid collagen preparation, used by Worthy and Holdaway (Citation1996); UF1, ultrafiltration with Eezi and Vivaspin filters, stepped-combustion with silver wire; UF2, ultrafiltration with Vivaspin filters, combustion with silver powder, contaminated iron catalyst; UF3, ultrafiltration with Vivaspin filters, combustion with silver powder, modern 14C-free iron catalyst.
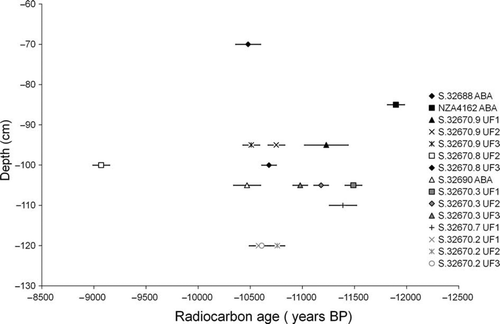
Table 3 Determining the age range of moa bones excavated from Glencrieff by Worthy and Holdaway (Citation1996).
Moa gizzard contents
The preserved contents of 16 individual moa gizzards were recovered during the 2007 excavation and were the first gizzards identified and recovered from Glencrieff (). Unlike moa gizzard contents from the Late Holocene calcareous mires at Pyramid Valley and Treasure Downs, at the Glencrieff and Scaife's Lagoon mires the boundary between the gizzard contents and peat matrix is indistinct, making the identification of gizzard contents somewhat difficult (Burrows et al. Citation1981; Wood Citation2007).
Figure 4 In situ heavy-footed moa (Pachyornis elephantopus (Owen, 1856)) gizzard contents resting between the rib cage and sternum. Between the sternum and gizzard was a matted layer of grass material.

Gizzard contents were characterized by dense accumulations of gizzard stones, and were, in some instances, associated with articulated or associated moa ribs and sterna, allowing identification of the moa species. The gizzard contents were associated with one P. elephantopus, one E. crassus and 14 unidentified moa. Associated bone radiocarbon dates () show that they are the oldest moa gizzard contents found in New Zealand. These samples include the third record of P. elephantopus gizzard contents (Wood Citation2007).
No large seeds or twigs were noted in the eight gizzard content samples analysed from Glencrieff, which is surprising as such material is considered characteristic of both gizzard contents (Burrows et al. Citation1981; Wood Citation2007) and coprolites (Horrocks et al. Citation2004; Wood et al. Citation2008a) from other sites. Small seeds, representing wetland taxa (Limosella Linnaeus 1753, Juncus Linnaeus 1753, Carex Linnaeus 1753, Ranunculus Linnaeus 1753, Colobanthus Bartling 1830 and Chenopodiaceae) were found within the gizzard contents (Appendix 1). Because the sub-sample volume was different between the eight analysed gizzards, the relative abundance of each wetland taxon cannot be compared between gizzards. Matted grass cf. Poaceae leaves were recovered from within the bowl of two sterna, one identified as P. elephantopus (), suggesting that grass formed part of the diet of P. elephantopus.
During the 2007 excavation, a small (approximately 150 mm diameter), discrete deposit of concentrated plant material was collected from amongst the moa bones but was not associated with moa gizzard stones. The sample included at least 74 twigs (identified as Nothofagus sp. cf. menziesii (Hook f.) Oerst 1873, silver beech), with lengths ranging from 9 to 43 mm and normally distributed (mean = 20.4 mm, SD = 8.1 mm). Similar distributions of twig lengths have been reported associated with both moa gizzard contents (Wood Citation2007) and nesting material (Wood Citation2008). The sample also included at least 18 leaf fragments of an unidentified monocot. Unlike some avian species, ratites do not have a crop that acts as a food storage organ. However, ratites do have a proventriculus, which forms part of the stomach and may store and commence food digestion before progression to the gizzard (Angel Citation1996). It is possible that the plant material described above came from the proventriculus.
Palynology
Radiocarbon dates obtained from the Moar (Citation2008) sediment core from Glencrieff, collected towards the edge of the deposit in shallower strata were 5460±120 yr BP (NZA5711, organic silt, δ13C−29.1) at 65 cm depth, and 9030±160 yr BP (NZA5712, organic silt, δ13C−27.4) at the base of the core (80 cm. See , Footnote 3 for further discussion). The base of the core is therefore probably contemporaneous with the majority of moa bones from Glencrieff (approximately 75–120 cm depth during 1993–94, and 45–104 cm during 2007), which were of Late Glacial age (). The pollen diagram () shows the decline of Phyllocladus Rich. Ex Mirb 1825, Plagianthus J.R. Forst & G. Forst 1776, Poaceae and Pteridium Gleditsch ex Scopoli 1760, which may have been more common near the site towards the Last Glacial Maximum. Prumnopitys taxifolia (D. Don) de Laub 1978 was established in the region by the Late Glacial, and pollen of this species is present throughout the Holocene. Southern beech (reflected by Nothofagus fusca-type pollen) established in North Canterbury around the mid-Holocene. The presence of wetland taxa (Cyperaceae, Myriophyllum Linneaus 1753, Potamogeton Walter 1788) throughout the core reveals that the Glencrieff wetland has been constantly wet since the Late Glacial. The upper 10 cm of the core represents the post-European era. Through this zone, charcoal and pollen of several plant taxa (e.g. bracken (Pteridium esculentum Cockayne 1908) and Poaceae) peak, and pollen of native tall trees and shrubs (e.g. P. taxifolia, Nothofagus Blume 1851, Coprosma J.R. Forst & G. Forst 1776) declines. This zone is also defined by the presence of exotic taxa (e.g. Salix Linneaus 1753, Ulex Linneaus 1753, Pinus Linneaus 1753).
Figure 5 Pollen diagram for sediment core, spot samples (raw data from Appendix 6 in Worthy & Holdaway Citation1996), and sediment samples taken from around concentrations of moa gizzard stones, Glencrieff. * denotes trace values (< 1%). Taxonomic names follow the International Plant Names Index (Citation2010). The grey shaded zone at the top of the figure represents the post-European era.

Pollen assemblages from spot sediment samples (A1/B1 and FO) collected at 75 cm depth from two locations in the bone deposit (see Worthy & Holdaway Citation1996 for raw data and spot locations) most likely represent Late Glacial vegetation at the site, and may be older than the base of the pollen core, as indicated by relatively high percentages of Phyllocladus, Myrsine Linneaus 1753, Coprosma and Asteraceae (probably tree daisies; Olearia Moench 1802), and low Prumnopitys taxifolia. Pollen assemblages for sediment samples obtained from around concentrations of gizzard stones (associated with one P. elephantopus, X09/2/21, and one E. crassus, X09/2/124), and assumed to be part of the gizzard, were similar to those from the spot sediment samples (). The relative abundance of spores of the dung fungus Sporormiella peaks in the upper 10 cm of the Moar (Citation2008) sediment core and reflects intensive sheep farming on the site. Sporormiella was present at low levels in Holocene samples of pre-settlement age but moderate levels in Late Glacial samples ().
Recent changes in site preservation conditions
Some significant changes in site preservation conditions have occurred at Glencrieff in the 14 years between the excavations in 1993–94 and 2007. In 1993, a blue clay layer between the overlying peat and the basement gravels was liquid and under pressure (Worthy & Holdaway Citation1996). In the subsequent 14 years, there has been significant drying and desiccation of the site. The water table has dropped and the blue clay layer has dried out. In addition, there were numerous large desiccation cracks that extended from the soil surface to the water table. In places, the blocks of peat created by these cracks had formed hard concretions around the bones.
Throughout the peat layer, we also found abundant roots from poplar trees (Populus Linneaus 1753) lining the western edge of the site. As in 1993–94, the roots appeared to be preferentially growing on the surfaces of the moa leg bones and throughout the more porous pelves. Bones higher in the stratigraphy were not as well preserved as those at lower levels due to the combination of poplar roots and low water table. Extensive erosion of the bone surface, especially of less dense elements such as pelves, was evident for bones nearer the surface of the 2007 deposit. It is important to note that bones excavated during 1993–94 were whitish in colour, while bones excavated during 2007 were dark brown in colour. It is therefore possible that the dropping of the water table, the desiccation and deterioration of Glencrieff, and differences in the preservation of excavated bones from the 1993–94 and 2007 deposits is due to the poplars. Further radiocarbon dating of bones from the 2007 excavation and comparison of the percentage carbon and nitrogen of bones from the 1993–94 and 2007 excavations would help determine the different preservation states of the deposits.
Discussion
Radiocarbon dating
All the radiocarbon dates obtained from P. elephantopus bones, except ANU 4937, were within the range of previously published dates for the site (Worthy & Holdaway Citation1996). However, the differences in the radiocarbon ages obtained using the three different pre-treatment methods (UF1–3) warrant further discussion.
Stepped combustion (UF1), following the methodology of Turney et al. (Citation2008), was originally designed for radiocarbon dating charcoal (Turney et al. Citation2001), eggshell (Bird et al. Citation2003) and sediment (Gilmour & Pillinger Citation1985), but not bone collagen. In the stepped combustion of bone collagen, labile contaminants are not removed as effectively due to the incremental heating of collagen in oxygen compared to combustion at a constant temperature (UF2–3; Gillespie Citation2008; Turney et al. Citation2008). The variation between the radiocarbon dates obtained using the UF1 versus UF2–3 methods can also be explained by differential removal of glycerol from the ultrafilters before collagen purification, contributing varying amounts of modern carbon to the analysis.
Two samples (specimens NMNZ S.32670.9 and NMNZ S.32670.3) returned older radiocarbon dates (ANU4923 and ANU4925) using the UF2 pre-treatment method (ultrafiltration, contaminated iron catalyst) than the UF3 pre-treatment method (ultrafiltration, silver powder and uncontaminated iron catalyst) (ANU7612 and ANU7614; ). Before each batch of radiocarbon dating the iron catalyst was cleaned using the standard procedure (reduction by hydrogen at 450 °C; Boaretto et al. Citation2009) and blanks were prepared for each batch of iron catalysts. It was subsequently discovered that the blanks for dates ANU4023–4925 contained a high percentage of modern carbon (pMC 0.24–0.38±0.03–0.04), alerting us to the presence of variable amounts of modern 14C contamination. The high blank values were subsequently used to correct the bone radiocarbon dates. If more contamination remained in the blank samples, then subtracting the 14C contribution of the blanks from the radiocarbon-dated bone samples over-compensated, leading to older ages. The ages obtained for bones using the UF3 method are the most reliable because the blank correction was very small.
Period of deposition
The radiocarbon ages for Glencrieff () indicate that the fossil deposit is about 10,000–12,000 uncalibrated years BP, dating it to the Late Glacial. The age range suggests that Glencrieff started trapping moa approximately 12,000 (uncalibrated) years BP and stopped about 10,000 (uncalibrated) years BP. It is probable that the lower peat layers are older than 12,000 years BP because the peat had to be at least 50 cm deep to mire moa (Wood et al. Citation2008b). There is no correlation between the radiocarbon dates and stratigraphic depth of the samples (, ), supporting the conclusions of Wood et al. (Citation2008b) that periodic peat liquefaction events and raking of fossil bones by the legs of mired moa caused disarticulation and intermingling of bones. If the radiocarbon dates encompass approximately the entire time period of miring at Glencrieff, this would give a minimum average miring rate of one moa per 28 years.
Taphonomy
The horizontal to sub-horizontal alignment of moa bones in the Worthy and Holdaway (Citation1996) and 2007 deposits (Wood et al. Citation2008b), and the discrete nature of the 2007, Eaves and Canterbury Museum deposits, suggest that there were at least four different ‘soft spots’ in the Glencrieff wetland where birds became mired. We interpret the site of the 2007 excavation as a spring up-welling or an extension of the spring outflow channel that flowed along the eastern edge of the mire (Wood et al. Citation2008b).
All the recovered moa bones were probably derived from individuals trapped in the mire. This contrasts with many of the small bird bones that were found (), which were probably from individuals that died around the mire, and were subsequently incorporated into the fossil deposit. These taphonomic observations suggest that an artesian spring developed on the terrace at Glencrieff during the Late Glacial. This facilitated the formation of floating peat deposits. Cracks, rifts, or local thin spots in the peat would have resulted in the miring of moa that attempted to walk over its surface, until the peat surface became solid enough to support moa in the Early Holocene. Rare liquefaction events may also have resulted in bones migrating through the overlying peat into the underlying liquid blue clay and being deposited on the basement gravels.
Moa gizzard contents, dietary and taphonomic implications
Despite the indistinct border between the preserved gizzard contents and the surrounding peat matrix, some interpretations of moa diet can still be inferred. The absence of large seeds and twigs, and the presence of small seeds from wetland taxa (Appendix A) and matted grass may suggest that moa survived for several days after miring by eating plants within their reach, but eventually starved to death or were killed by Haast's eagle (Batcheler Citation1989; Wood Citation2007). Batcheler (Citation1989) hypothesized that preserved moa gizzard contents with large amounts of plant material were probably from individuals that died soon after becoming mired, either by drowning or predation by Haast's eagle, and were incorporated quickly into the fossil deposit. Preserved gizzard contents from Pyramid Valley support this hypothesis, where perhaps mired moa quickly sank out of sight (Burrows et al. Citation1981). In addition, numerous moa pelves found in North Canterbury swamp deposits, including Glencrieff (Worthy & Holdaway Citation1996; this study) have puncture marks attributed to either Haast's eagle claws or kea beak marks. In contrast, Batcheler (Citation1989) hypothesized that preserved moa gizzard contents with no or small amounts of preserved plant material were probably from individuals that survived for a few days after miring and starved to death before becoming incorporated into the fossil deposit, as is the case with sheep and cows trapped in swamps today. In addition, plant material in the gizzard would have been digested and defecated into the surrounding peat matrix, contributing to the moderate levels of Sporormiella in sediment samples from around gizzard stone concentrations from the 2007 excavation. Gizzard stone concentrations with similar amounts of plant material to those from Glencrieff have also been found in Pyramid Valley swamp (Eyles Citation1955).
It is also possible that the plant material in the gizzards has not been preserved. Batcheler (Citation1989) hypothesized that this may be due to prolonged exposure and decomposition on the surface of the deposit after death. However, extensive decomposition of plant material at Glencrieff is considered unlikely, given the presence of material inferred to be preserved proventriculus contents (twigs and leaves), and the matted grass material found between a P. elephantopus sternum and gizzard stones.
Palynology
The pattern of vegetation change at Glencrieff () characterized by the dominance of Phyllocladus, either co-dominant or being replaced by podocarps (e.g. Prumnopitys taxifolia), and followed by beech (Nothofagus) establishment in the mid-late Holocene, is characteristic of pollen records from other eastern South Island sites (e.g. Moar Citation1973; Burrows & Russell Citation1990; Wilmshurst et al. Citation2002). Worthy and Holdaway (Citation1996) argued that the pollen profile from the spot sediment sample A1/B1 (dominant grassland/shrubland vegetation; ) was older than the sediment sample F0 (closed woodland vegetation), albeit from the same depth (75 cm), through comparison with published pollen records (Moar Citation1971; McGlone Citation1988). The pollen profiles from F0 and the moa gizzard content samples () probably reflect the predominant vegetation surrounding Glencrieff during the period of bone deposition. This is in contrast to the basal layers of the Moar (Citation2008) sediment core. The base of the peat was reached at 80 cm depth, whereas the base of the peat over the Worthy and Holdaway (Citation1996) and 2007 deposits was 104–120 cm in depth. Considering Worthy and Holdaway's (1996) hypothesis that Glencreiff developed as a peat dome, thinning towards the edge, the basal layers of the sediment core may not be contemporaneous with bone deposition at Glencreiff.
The increase in charcoal, bracken spores and Poaceae pollen with the coincident decrease in the pollen of native trees and shrubs (e.g. P. taxifolia, Nothofagus, Coprosma) in the upper 10 cm of the sediment core () represent forest clearance and development of agricultural land by Europeans. In addition, the relative abundance of spores of the coprophilous fungus Sporormiella in this section () and the low to moderate levels in pre-settlement layers during the Holocene and Late Glacial, respectively, indicate the presence of native avian herbivores, but at much lower densities compared to current livestock. The low level of Sporormiella spores during the Holocene supports the hypothesis that miring and bone deposition at Glencrieff occurred during the Late Glacial only.
Avifauna and ecological implications
Of the 18 species of birds excavated from Glencrieff, nine are extinct, while a further three are extinct locally (Appendix 2; Wilson Citation2004; Robertson et al. Citation2007). Of the remaining five species, kea (Nestor) and kakariki (Cyanoramphus Bonaparte, 1854) are absent from the lowlands, while kereru (Hemiphaga novaeseelandiae (Gmelin, 1789)) is restricted in this region to Banks Peninsula and lowland forest remnants (Robertson et al. Citation2007). This reflects a significant decline in avifaunal diversity since the arrival of Polynesians (c. AD 1280; Wilmshurst et al. Citation2008) and later Europeans in New Zealand.
Pachyornis elephantopus and E. crassus dominate the Late Glacial moa assemblage of Glencrieff, with rare D. robustus and E. curtus. When compared to assemblages from other Otiran Glacial–Early Holocene sites in the eastern South Island, it is evident that Glencrieff represents a characteristic fauna of this time period. Other Otiran Glacial–Early Holocene sites in the eastern South Island, including Omihi Stream (Worthy & Holdaway Citation1996), Cape Wanbrow (Worthy & Grant-Mackie Citation2003), Merino Cave (Worthy & Holdaway Citation1995), Herbert (Oliver Citation1949), and Albury Park (Worthy Citation1997a), have similar faunal assemblages to Glencrieff. They are characterized by the dominance of P. elephantopus, with either E. crassus or E. curtus also dominant (depending on the elevation and aspect of the fossil locality), and D. robustus comparatively rare. This is in contrast to Otiran Glacial–Late Glacial sites in Southland, including Opio (NJR, pers. obs.) and Kauana (Worthy Citation1998b), where E. curtus is the dominant moa, with fewer E. crassus, P. elephantopus and D. robustus.
By the Late Holocene, the composition of the avifauna in North Canterbury had altered, as shown by the faunas from Pyramid Valley (Holdaway & Worthy Citation1997) and Bell Hill Vineyard (Allentoft et al. Citation2010). Warming temperatures and the establishment of tall podocarp forest by the Early Holocene and mixed southern beech forest (Nothofagus) and podocarp forest by the Middle to Late Holocene () resulted in significant shrinking of the open tussock grassland/shrubland mosaic habitat (Moar Citation2008) favoured by P. elephantopus (Worthy & Holdaway Citation1995). Late Holocene fossil deposits in North Canterbury, including Pyramid Valley, Bell Hill, Glenmark (Worthy & Holdaway Citation1996) and Treasure Downs (McCulloch Citation1987), are dominated by E. crassus, and either E. curtus or D. robustus (depending on elevation and aspect of the fossil locality), with P. elephantopus comparatively rare. Other Late Holocene sites in the eastern South Island generally follow this trend, including several sites in South Canterbury (Worthy Citation1997a) and North Otago (Worthy Citation1998a). However, Worthy (Citation1998a) reported that Late Holocene sites in Central Otago were dominated by P. elephantopus, with co-dominant E. curtus or E. crassus, reflecting the increased abundance of the open grassland/shrubland mosaic in Central Otago (Wood & Walker Citation2008) favoured by P. elephantopus.
Worthy and Holdaway (Citation1996) found the absence of E. curtus from Glencrieff intriguing because the species was present in low numbers at other Otiran–Late Glacial sites in Canterbury (Worthy Citation1993a). They hypothesized that E. curtus had a preference for hill country where soils were drier, with E. crassus replacing E. curtus at lower altitudes. This is evident when the relative proportions of moa species in the Late Holocene Pyramid Valley and Bell Hill fossil deposits are compared. Pyramid Valley, located in a basin, has an assemblage dominated by E. crassus and D. robustus, with E. curtus and P. elephantopus present but rarer (Holdaway & Worthy Citation1997). Bell Hill, located on a hillside, has significantly more E. curtus than Pyramid Valley, and D. robustus is rare (Allentoft et al. Citation2010). The single E. curtus from Glencrieff is therefore interpreted as a rare individual on the margin of its preferred habitat, and its former absence the stochastic effect of small sample size. This may also apply to rare E. curtus individuals from Otiran–Late Glacial deposits in the wetter northwest Nelson/West Coast regions of the South Island (Worthy Citation1993b, Citation1994, Citation1997b; Worthy & Holdaway Citation1994).
Conclusions
Glencrieff swamp, in the Waikari Basin, North Canterbury, samples a diverse Late Glacial avifauna. Comparisons with other Otiran Glacial–Late Holocene fossil deposits in the eastern South Island indicate that the Glencrieff avifauna represents a characteristic Otiran Glacial–Early Holocene fauna. A deeper sediment core from the middle of the Glencrieff deposit will probably provide a more complete floral record than presented here, and would help determine the vegetation changes occurring during the period of fossil deposition at Glencreiff. The 2007 excavations have also documented the oldest moa gizzard contents yet found (), and the first occurrence of E. curtus at Glencrieff. The seasonal wetting and drying of the deposit and the extraction of water by the poplars surrounding Glencrieff will probably continue to degrade the remaining bones and impair the archival qualities of the sediments. Future radiocarbon dating of moa bones from the 2007 and 1971 Canterbury Museum excavations will determine whether bone deposition within each deposit at Glencrieff occurred during the same time period. Loss of the Eaves collection overseas is unfortunate as radiocarbon dating of this deposit would be important to gain a complete understanding of the taphonomy and deposition at this significant fossil locality.
Acknowledgements
Excavations were supported by funding from the Australian Research Council to Alan Cooper, and the manuscript was prepared under the tenure of an Australian Postgraduate Award (NJR). The 2007 excavation was conducted by I. and M. Dodson, R. Fuller, N. Rawlence, P. Scofield and J. Wood. We are indebted to M. Wilson and the Wilson family for permission to carry out the excavation. We are grateful for the help of Chris Turney, Richard Gillespie, Keith Fifield and Stewart Fallon with collagen preparation and radiocarbon dating. We thank Jeremy Austin, Jessica Metcalf, Maria Zammit and Alan Cooper for advice and editorial help on this manuscript. Finally we thank James Crampton and one anonymous reviewer for peer reviewing this manuscript.
References
- Allentoft , ME , Bunce , M , Scofield , RP , Hale , ML and Holdaway , RN . 2010 . Highly skewed sex ratios and biased fossil deposition of moa: Ancient DNA provides new insights on New Zealand's extinct megafauna . Quaternary Science Reviews , 29 : 753 – 762 .
- Angel , CR . 1996 . A review of ratite nutrition . Animal Feed Science Technology , 60 : 241 – 246 .
- Batcheler , CL . 1989 . Moa browsing and vegetation formations, with particular reference to deciduous and poisonous plants . New Zealand Journal of Ecology , 12 : 57 – 65 .
- Bird , MI , Turney , CSM , Fifield , LK , Smith , MA , Miller , GH , Roberts , RG and Magee , JW . 2003 . Radiocarbon dating of organic- and carbonate-carbon in Genyornis and Dromaius eggshell using stepped combustion and stepped acidification . Quaternary Science Reviews , 22 : 1805 – 1812 .
- Boaretto , E , Wu , X , Yuan , J , Bar-Yosef , O , Chu , V , Pan , Y , Liu , K , Cohen , D , Jiao , T , Li , S , Gu , H , Goldberg , P and Weiner , S . 2009 . Radiocarbon dating of charcoal and bone collagen associated with early pottery at Yuchanyan Cave, Hunan Province, China . Proceedings of the National Academy of Sciences of the United States of America , 106 : 9595 – 9600 .
- Bronk Ramsey , C , Higham , TFG , Bowles , A and Hedges , R . 2004a . Improvements to the pretreatment of bone at Oxford . Radiocarbon , 46 : 155 – 163 .
- Bronk Ramsey , C , Higham , TFG and Leach , P . 2004b . Towards high precision AMS: progress and limitations . Radiocarbon , 46 : 7 – 24 .
- Bunce , M , Worthy , TH , Phillips , M , Holdaway , RN , Willerslev , E , Haile , J , Shapiro , B , Scofield , P , Drummond , A , Kamp , P and Cooper , A . 2009 . The evolutionary history of the extinct ratite moa and New Zealand palaeogeography revealed by ancient DNA . Proceedings of the National Academy of Sciences of the United States of America , 106 : 20646 – 20651 .
- Burrows , CJ , McCulloch , B and Trotter , MM . 1981 . The diet of moas based on gizzard contents from Pyramid Valley, North Canterbury, and Scaife's Lagoon, Lake Wanaka, Otago . Records of the Canterbury Museum , 9 : 309 – 336 .
- Burrows , CJ and Russell , JB . 1990 . Aranuian vegetation history of the Arrowsmith Range, Canterbury. 1: Pollen diagrams, plant macrofossils, and buried soils from Prospect Hill . New Zealand Journal of Botany , 28 : 323 – 345 .
- Clark , RL . 1982 . Point count method estimation of charcoal in pollen preparations and thin sections of sediments . Pollen et Spores , 24 : 523 – 535 .
- Davis , OK and Shafer , DS . 2006 . Sporormiella fungal spores, a palynological means of detecting herbivore density. Palaeogeography, Palaeoclimatology . Palaeoecology , 237 : 40 – 50 .
- Eyles , JR . 1955 . Field notes on the excavations . Records of the Canterbury Museum , 6 : 257 – 260 .
- Gill , BJ , Bell , BD , Chambers , GK , Medway , DG , Palma , RL , Scofield , RP , Tennyson , AJD and Worthy , TH . 2010 . Checklist of the birds of New Zealand, Norfolk and Macquarie Islands, and the Ross Dependency, Antarctica , 4th edition , Wellington : Ornithological Society of New Zealand and Te Papa Press. 464 p .
- Gillespie , R . 2008 . Updating Martin's global extinction model . Quaternary Science Reviews , 27 : 2522 – 2529 .
- Gilmour , I and Pillinger , CT . 1985 . Stable carbon isotopic analysis of sedimentary organic matter by stepped combustion . Organic Chemistry , 8 : 421 – 426 .
- Higham , TFG , Bronk-Ramsey , FJ , Petchey , C , Tompkins , C and Taylor , M . 2004 . “ New AMS radiocarbon determinations from Kokohuia, New Zealand ” . In 14C and Archaeology, Proceedings of the Fourth Symposium, Oxford 2002. Oxford University School of Archaeology Monograph 62 , Edited by: Higham , TFG , Bronk-Ramsey , C and Owen , C . 135 – 151 . Oxford : Oxbow Books .
- Holdaway , RN and Worthy , TH . 1997 . A reappraisal of the late Quaternary fossil vertebrates of Pyramid Valley Swamp, North Canterbury, New Zealand . New Zealand Journal of Zoology , 24 : 69 – 121 .
- Horrocks , M , D'Costa , D , Wallace , R , Gardner , R and Kondo , R . 2004 . Plant remains in coprolites: diet of a subalpine moa (Dinornithiformes) from southern New Zealand . Emu , 104 : 149 – 156 .
- International Plant Names Index 2010 . http://www.ipni.org/ ( accessed 12 August 2010 ).
- Jacobi , RM , Higham , TFG and Bronk Ramsey , C . 2006 . AMS radiocarbon dating of Middle and Upper Palaeolithic bone in the British Isles: improved reliability using ultrafiltration . Journal of Quaternary Science , 21 : 557 – 573 .
- McCormac , FG , Hogg , AG , Blackwell , PG , Buck , CE , Higham , TFG and Reimer , PJ . 2004 . ShCal04 Southern Hemisphere calibration, 0–11.0 Cal Kyr BP . Radiocarbon , 46 : 1087 – 1092 .
- McCulloch B 1987 . New Zealand . In : Mourer-Chauvire C . Letter of information of the Society of Avian Paleontology and Evolution 1 . http://www2.nrm.se/ve/birds/sape/sapenews1.html.en ( accessed 11 August 2010 ).
- McGlone , MS . 1988 . “ New Zealand ” . In Vegetation History , Edited by: Huntley , B and Webb , T III . 557 – 599 . Dordrecht : Kluwer Academic Publishers .
- Meylan , BA and Butterfield , BG . 1978 . The structure of New Zealand woods , Wellington : Department of Scientific and Industrial Research .
- Moar , NT . 1971 . Contributions to the Quaternary history of the New Zealand flora: 6. Aranuian pollen diagrams from Canterbury, Nelson, and North Westland, South Island . New Zealand Journal of Botany , 9 : 80 – 145 .
- Moar , NT . 1973 . Contributions to the Quaternary history of the New Zealand flora: 7. Two Aranuian pollen diagrams from central South Island . New Zealand Journal of Botany , 11 : 291 – 304 .
- Moar , NT . 2008 . “ Late Quaternary Vegetation ” . In The Natural History of Canterbury , Edited by: Winterbourn , M , Knox , G , Burrows , C and Marsden , I . 169 – 192 . Christchurch : Canterbury University Press .
- Moore , PD , Collinson , M and Webb , JA . 1991 . Pollen analysis , 216 Oxford : Blackwell Scientific .
- Newnham , RM , Vandergoes , MJ , Hendy , CH , Lowe , DJ and Preusser , F . 2007 . A terrestrial palynological record for the last two glacial cycles from southwestern New Zealand . Quaternary Science Reviews , 26 : 517 – 535 .
- Oliver , WRB . 1949 . The moas of New Zealand and Australia , Wellington : Dominion Museum Bulletin No. 15 .
- Reimer , PJ , Baillie , MGL , Bard , E , Bayliss , A , Beck , JW , Bertrand , CJH , Blackwell , PG , Buck , CE , Burr , GS , Cutler , KB , Damon , PE , Edwards , RL , Fairbanks , RG , Friedrich , M , Guilderson , TP , Hogg , AG , Hughen , KA , Kromer , B , McCormac , G , Manning , S , Bronk Ramsey , C , Reimer , RW , Remmele , S , Southon , JR , Stuiver , M , Talamo , S , Taylor , FW , van der Plicht , J and Weyhenmeyer , CE . 2004 . INTCAL04 terrestrial radiocarbon age calibration, 0–26 Cal Kya BP . Radiocarbon , 46 : 1029 – 1058 .
- Robertson , CJR , Hyvonen , P , Fraser , MJ and Pickard , CR . 2007 . Atlas of bird distribution in New Zealand 1999–2004 , 533 New Zealand : Ornithological Society of New Zealand .
- Tennyson , A and Martinson , P . 2007 . Extinct birds of New Zealand , 179 Wellington : Te Papa Press . (reprint)
- Turney , CSM , Bird , MI , Fifield , LK , Roberts , RG , Smith , M , Dortch , CE , Grun , R , Lawson , E , Ayliffe , LK , Miller , GH , Dortch , J and Cresswell , RG . 2001 . Early human occupation at Devil's Lair, Southwestern Australia 50,000 years ago . Quaternary Research , 55 : 3 – 13 .
- Turney , CSM , Flannery , TF , Roberts , RG , Reid , C , Fifield , LK , Higham , TFG , Jacobs , Z , Kemp , N , Colhoun , EA , Kalin , RM and Ogle , N . 2008 . Late-surviving megafauna in Tasmania, Australia, implicate human involvement in their extinction . Proceedings of the National Academy of Sciences of the United States of America , 105 : 12150 – 12153 .
- Webb , CJ and Simpson , MJA . 2001 . Seeds of New Zealand: gymnosperms and dicotyledons , 428 Christchurch : Manuka Press .
- Wilmshurst , JM , Anderson , AJ , Higham , TFG and Worthy , TH . 2008 . Dating the late prehistoric dispersal of Polynesians to New Zealand using the commensal Pacific rat . Proceedings of the National Academy of Sciences of the United States of America , 105 : 7676 – 7680 .
- Wilmshurst , JM , McGlone , MS and Charman , DJ . 2002 . Holocene vegetation and climate change in southern New Zealand: linkages between forest composition and quantitative surface moisture reconstructions from an ombrogenous bog . Journal of Quaternary Science , 17 : 653 – 666 .
- Wilson , KJ . 2004 . Flight of the Huia: ecology and conservation of New Zealand's frogs, reptiles, birds and mammals , 411 Christchurch : Canterbury University Press .
- Wood , JR . 2007 . Moa gizzard content analysis: further information on the diet of Dinornis robustus and Emeus crassus, and the first evidence for the diet of Pachyornis elephantopus (Aves: Dinornithiformes) . Records of the Canterbury Museum , 21 : 27 – 39 .
- Wood , JR . 2008 . Moa (Aves: Dinornithiformes) nesting material from rockshelters in the semi-arid interior of South Island, New Zealand . Journal of the Royal Society of New Zealand , 38 : 115 – 129 .
- Wood , JR and Walker , S . 2008 . Macrofossil evidence for pre-settlement vegetation of Central Otago's basin floors and gorges . New Zealand Journal of Botany , 46 : 239 – 255 .
- Wood , JR , Rawlence , NJ , Rogers , GM , Austin , JJ , Worthy , TH and Cooper , A . 2008a . Coprolite deposits reveal the diet and ecology of the extinct New Zealand megaherbivore moa (Aves: Dinornithiformes) . Quaternary Science Reviews , 27 : 2593 – 2602 .
- Wood , JR , Worthy , TH , Rawlence , NJ , Jones , SM and Read , SE . 2008b . A deposition mechanism for Holocene miring bone deposits, South Island, New Zealand . Journal of Taphonomy , 6 : 1 – 20 .
- Wood JR , Wilmshurst JM , Worthy TH , Cooper A 2011 Sporormiella as a proxy for non-mamalian herbivores in island ecosystems . Quaternary Science Reviews 30: 915–920 .
- Worthy , TH . 1988 . An illustrated key to the main leg bones of moas (Aves-Dinornithiformes) , National Museum of New Zealand Miscellaneous Publications Series 17 .
- Worthy , TH . 1993a . A review of fossil bird bones from loess deposits in eastern South Island, New Zealand . Records of the Canterbury Museum , 10 : 95 – 106 .
- Worthy , TH . 1993b . Fossils of Honeycomb Hill , 56 Wellington : Museum of New Zealand Te Papa Tongarewa .
- Worthy , TH . 1994 . Late Quaternary changes in the moa fauna on the West Coast of the South Island, New Zealand . Records of the South Australian Museum , 27 : 125 – 134 .
- Worthy , TH . 1997a . Quaternary fossil fauna of South Canterbury, South Island, New Zealand . Journal of the Royal Society of New Zealand , 27 : 67 – 162 .
- Worthy , TH . 1997b . Fossil deposits in the Hodge Creek Cave System, on the northern foothills of Mt Arthur, Nelson, South Island, New Zealand . Notornis , 44 : 111 – 124 .
- Worthy , TH . 1998a . Quaternary fossil faunas of Otago, South Island, New Zealand . Journal of the Royal Society of New Zealand , 28 : 421 – 521 .
- Worthy , TH . 1998b . The Quaternary fossil avifauna of Southland, South Island, New Zealand . Journal of the Royal Society of New Zealand , 28 : 537 – 589 .
- Worthy , TH and Grant-Mackie , JA . 2003 . Late-Pleistocene avifaunas from Cape Wanbrow, Otago, South Island, New Zealand . Journal of the Royal Society of New Zealand , 33 : 427 – 485 .
- Worthy , TH and Holdaway , RN . 1994 . Quaternary fossil faunas from caves in Takaka valley and on Takaka Hill, northwest Nelson, South Island, New Zealand . Journal of the Royal Society of New Zealand , 24 : 297 – 391 .
- Worthy , TH and Holdaway , RN . 1995 . Quaternary fossil faunas from caves on Mt Cookson, North Canterbury, New Zealand . Journal of the Royal Society of New Zealand , 25 : 333 – 370 .
- Worthy , TH and Holdaway , RN . 1996 . Quaternary fossil faunas, overlapping taphonomies, and palaeofaunal reconstruction in North Canterbury, South Island, New Zealand . Journal of the Royal Society of New Zealand , 26 : 275 – 361 .
- Worthy , TH and Holdaway , RN . 2002 . The lost world of the moa: Prehistoric life of New Zealand , 718 Christchurch : Canterbury University Press .
- Worthy , TH and Roscoe , D . 2003 . Takaka Fossil Cave – a stratified Late Glacial to Late Holocene deposit from Takaka Hill, New Zealand . Tuhinga, Records of the Museum of New Zealand Te Papa Tongarewa , 14 : 41 – 60 .
Appendix 1 Seed counts from sub-samples of sediment associated with gizzard stone concentrations
GC numbers refer to specimen numbers assigned to preserved gizzard contents in the field. Taxonomic names of plants follow the International Plant Names Index (Citation2010).
Table A1.1
Appendix 2 Vertebrate fossils from Glencrieff miring bone deposit
Taxonomic names are as advocated by Gill et al. (Citation2010). Geographical distribution and status of taxa: 1extinct; 2locally extinct; 3restricted geographical distribution; 4widespread geographical distribution; and 5non-applicable. Eaves collection was detailed by Worthy and Holdaway (Citation1996): its listing below indicates that material from this collection was included in the x/y data. Abbreviations: juvenile (juv), subadult (subad), Australian Centre for Ancient DNA (ACAD).